










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkDarwiniana, nueva serie
Print version ISSN 0011-6793On-line version ISSN 1850-1699
Darwiniana vol.48 no.1 San Isidro Jan./June 2010
ECOLOGÍA Y FITOGEOGRAFÍA
Composición, estructura y patrón espacial de un bosque tucumano-boliviano en el departamento de Tarija (Bolivia)
F. Santiago Zenteno-Ruiz1,2 & R. Pablo López2,1
1Herbario Nacional de Bolivia (LPB), Instituto de Ecología, Casilla 10077 Correo Central, La Paz, Bolivia; fred6zenruiz@yahoo.com (autor corresponsal)
2Centro de Análisis Espacial, Instituto de Ecología, Cota Cota, Casilla 10077 Correo Central, La Paz, Bolivia.
Resumen
El bosque tucumano-boliviano, en su sector correspondiente a Bolivia, ha comenzado a ser conocido con mayor detalle en los últimos años. Sin embargo, los estudios realizados han tenido un carácter más cualitativo o se han realizado en superficies pequeñas (0,1 ha); además, la biogeografía de sus zonas más bajas no está del todo clara. Por otro lado, algunos aspectos ecológicos, como los referidos a patrones espaciales de los árboles, no han sido nunca abordados. Nosotros realizamos el estudio de una hectárea de bosque tucumano-boliviano a 700-950 m, y mapeamos todos los árboles y lianas con DAP ≥10 cm a fin de caracterizar cuantitativamente un área representativa de este bosque. El propósito fue conocer su composición, abundancia y estructura. Realizamos el primer estudio de patrones espaciales para este tipo de bosque en Bolivia. Registramos 591 individuos agrupados en 31 familias, 47 géneros y 57 especies. Seis especies fueron las más abundantes: Urera caracasana, Trichilia clausenii, Piper tucumanum, Diatenopteryx sorbifolia, Chrysophyllum gonocarpum y Cupania vernalis. El bosque estudiado puede considerarse parte del bosque tucumano-boliviano más que del Arco Pleistocénico. El área basal total fue de 29.4 m2 /ha. La presencia de lianas fue relativamente baja, lo que indica que se trata de un bosque relativamente poco perturbado. Se trata de un lugar diverso. La riqueza específica resultó comparable a la de los bosques secos de Madidi (La Paz, Bolivia), y superior a la del tucumano-boliviano en el lado argentino, y fue estimada en 74 especies de árboles. La diversidad (3,5 índice Shannon-Wiener) es tan alta como la de muchos bosques más tropicales, e incluso algunos amazónicos. Encontramos un patrón espacial aleatorio en los individuos de la comunidad, y la mayoría de las especies presentó agregación a alguna escala espacial.
Palabras clave. Bosque tucumano-boliviano; Composición específica; Diversidad; Estructura; Función K de Ripley; Patrón espacial; Rarefacción.
Abstract
Composition, structure and spatial pattern of Tucumano-Bolivia no forest in the department of Tarija (Bolivia).
The Tucumano-Boliviano forest, on its Bolivian side, has begun to be studied with greater detail in the last decade. However, the studies that have been conducted have had a more qualitative character or have been undertaken in small areas (0.1 ha); on the other hand, the biogeography of its lower zones is not completely understood. Moreover, some ecological aspects, such as tree spatial patterns, have never been addressed. We conducted a study in one hectare of tucumano-boliviano forest at 700-950 m, and mapped all trees and vines with DBH ≥10 cm in order to characterize quantitatively a representative area of this forest. The purpose was to obtain insight into its floristic composition, species abundance and structure. We carried out the first study on spatial patterns for this type of forest in Bolivia. We recorded 591 individuals belonging to 31 families, 47 genera, and 57 species. Six species were the most abundant and: Urera caracasana, Trichilia clausenii, Piper tucumanum, Diatenopteryx sorbifolia, Chrysophyllum gonocarpum and Cupania vernalis. The forest studied can be classed more as Tucumano-Boliviano forest than as a Pleistocene Arc type forest. Basal area was 29.04 m2 /ha. There was a relatively scarce presence of lianas, which indicates that this is a relatively slightly disturbed community. This is a diverse forest. Species richness turned out to be comparable to that of Madidi dry forests, and greater than that in Tucumano-Boliviano forest in Argentina, and was estimated at 74 tree species. Diversity (3.5 Shannon-Wiener index) is as high as that in many tropical forests, even in Amazonia. We found a random spatial pattern in the individuals of the community, and most of the species exhibited clustering at some spatial scale.
Keywords. Diversity; Rarefaction; Ripleys K-función; Spatial pattern; Species composition; Structure; Tucumano-Boliviano forest.
Introducción
Los bosques semihúmedos de las laderas orientales del sur de Bolivia y norte de Argentina se conocen como bosques tucumano-bolivianos. Poseen elementos florísticos, relaciones biogeográficas y una estacionalidad térmica e hídrica peculiares. Se distinguen florísticamente de los bosques montanos húmedos del norte del codo de los Andes boliviano-peruanos (Ibisch et al., 2003). Su extensión latitudinal es relativamente considerable, ya que se extienden desde los 18º hasta los 27-28º de latitud sur. Se distribuye en una estrecha franja altitudinal que va de los 550 a 3.000 m (a veces alcanza los 3.600 m de altitud) desde el norte de Argentina hasta la latitud de la ciudad de Santa Cruz (Hueck, 1978). Se pueden reconocer varias subformaciones bajo distintas condiciones climáticas y edáficas.
El bosque tucumano-boliviano (conocido como el bosque yungueño austral sensu stricto) corresponde a la ecorregión mejor conocida de la cliserie (la serie de pisos altitudinales) a latitudes subtropicales. Hay varios trabajos sobre este tipo de bosque, aunque en su mayor parte se trata de documentos inéditos. En Bolivia, se halla emplazado entre unos 650 y 3.000 m, aunque puede en ciertas localidades protegidas subir un poco más (Zenteno et al., 2010).
Estos bosques representan ecosistemas singulares, exclusivos de los Andes de Bolivia y Argentina, pero se encuentran en gran medida degradados, en especial en el segundo país. Son diversos pero en menor medida que los bosques yungueños. Sin embargo, tienen especies y géneros exclusivos. Probablemente, la proporción de especies endémicas de estos bosques sea menor que la de las otras áreas más secas, aunque la diversidad local suele ser mayor en el bosque tucumano-boliviano que en los valles secos.
Esta formación vegetal está presente en la Reserva Nacional de Flora y Fauna Tariquía y, en pequeña medida, en la mitad inferior de la Cuesta de Sama en el departamento de Tarija. Como formaciones vegetales con valor de conservación se pueden mencionar los parches de Polylepis hieronymi Pilg. y la faja boscosa de Podocarpus parlatorei Pilg., ambas situadas en la parte superior del bosque tucumano-boliviano. Las especies importantes son Podocarpus parlatorei (pino del cerro), Polylepis crista-galli Bitter, P. hieronymi (keñua), Alnus acuminata Kunth (aliso), Juglans australis Griseb. (nogal de monte), Phoebe porphyria (Griseb.) Mez (laurel), Blepharocalyx salicifolius (Kunth) O. Berg (barroso), Tabebuia lapacho (K. Schum.) Sandwith (lapacho), Weinmannia boliviensis R.E. Fr., Cedrela lilloi C. DC., C. fissilis Vell. y C. saltensis M.A. Zapater & del Castillo (cedro).
Se han realizado varios estudios en el bosque tucumano-boliviano del sector argentino, pero relativamente pocos en el más diverso lado boliviano. Son particularmente escasos los estudios de la vegetación de esta ecorregión en superficies grandes (una ha o más). Además, la biogeografía de los pisos inferiores del bosque tucumano-boliviano no es del todo clara, ya que hay afinidades andinas (yungueñas), por un lado, y paranenses (Arco Pleistocénico), por otro. Por otra parte, hasta donde conocemos, prácticamente no se han realizado estudios de patrones espaciales en ninguna de nuestras ecorregiones. Los patrones espaciales son un atributo muy importante cuando se empieza a estudiar una determinada comunidad vegetal, pues brindan información sobre los procesos que pueden estar creando dichos patrones. Hay muchos estudios sobre patrones espaciales en regiones templadas del planeta, en regiones tropicales como en Panamá y Costa Rica, y prácticamente inexistentes en el caso de ecorregiones presentes en Bolivia, donde existen numerosas (ca. 700) parcelas permanentes de muestreo (PPM) instaladas.
Los objetivos del trabajo fueron describir la composición, la diversidad y la estructura de una parcela de bosque tucumano-boliviano en Tarija (Bolivia) y evaluar el patrón espacial de la comunidad en general y de las especies dominantes que permite inferir algunos procesos bióticos que actúan sobre estas. Así mismo, este trabajo es uno de los primeros en describir patrones espaciales para una parcela permanente instalada en Bolivia.
Área de estudio
La parcela permanente (PPM) está situada en la Estación Experimental Conchas de la Universidad Misael Saracho, próxima al río Conchas (22º 20' 37,38'' S; 64º 24'15,12'' O). Se encuentra a una altitud de entre 700 y 950 m, en el bosque submontano, provincia Arce, departamento de Tarija. El acceso a la zona de estudio es por la carretera principal Tarija-Bermejo y luego por el camino secundario hasta la comunidad Conchas, a 50 km en línea recta al N de la ciudad de Bermejo (Fig. 1). El clima se caracteriza por tener una precipitación promedio de 700 a 2000 mm por año y tres meses secos (junio-agosto), con una temperatura promedio anual de 5 a 23°C (Ibisch et al., 2003).
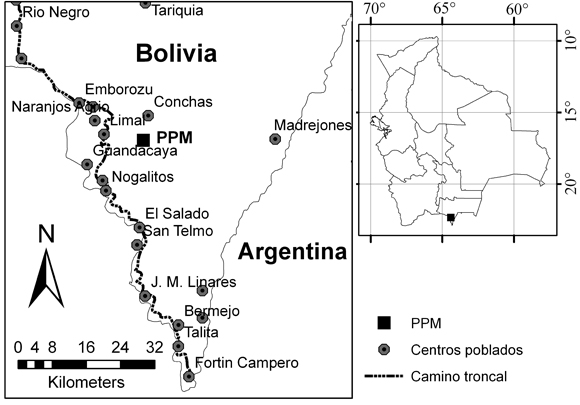
Fig. 1. Ubicación de la parcela permanente de muestreo (PPM) en el bosque tucumano-boliviano.
Material y métodos
La parcela permanente está instalada en una ladera con exposición sudeste, y la muestra fue de una hectárea de superficie dividida en 25 subparcelas de 20 x 20 m (Curtis & McIntosh, 1951). El diseño de la muestra tiene dos líneas principales una que va de sur a norte con una pendiente promedio de 35° y la otra de este a oeste con 15°. Se inventariaron todos los árboles y lianas con un diámetro a la altura del pecho (DAP, a 1.3 m del suelo) igual o mayor a 10 cm, al igual que troncos irregulares (Dallmeier, 1992). Cada individuo fue identificado hasta especie; además, se registraron sus coordenadas en el eje "x" e "y", y se estimó visualmente su altura total y altura de fuste. Las variables ecológicas consideradas fueron posición de copa, forma de copa e infestación de lianas (Dawkins, 1958; Lowe & Walter, 1977; Lamprecht, 1990).
Para los análisis cuantitativos y cualitativos de los datos, se calcularon las siguientes variables relativas (por especie): densidad (Den), dominancia (AB en m2/ha), frecuencia (Frec) y el índice de valor de importancia de especies (IVI; Mueller-Dombois & Ellenberg, 1974) que es la suma de las tres variables relativas mencionadas anteriormente. Por otro lado, se analizó la distribución vertical y horizontal (altura y dominancia) y algunas variables ecológicas representadas en los individuos de la parcela con respecto a la posición y a la forma de la copa. Se consideraron cinco categorías de posición de copa con respecto a la luz solar (Dawkins, 1958): emergente, libre de competencia lateral (1); dominante, con competencia lateral (2); codominante, parcialmente sombreada por otras copas (3); intermedia, solo una porción de la copa expuesta a la luz (4); y suprimida, con ausencia de luz (5). Con respecto a la simetría de la copa (forma de la copa) se consideraron cinco categorías: simétrica o circular (1); levemente asimétrica (2); asimétrica o rala (3); marcadamente asimétrica y con pocas ramas (4); degradada o suprimida (5). La infestación de lianas (Lowe & Walter, 1977) consignó cuatro categorías: árbol libre de lianas (1); lianas solamente en el fuste (2); lianas en el fuste y levemente en la copa (3); lianas en la totalidad del fuste y copa (4). La diversidad florística fue evaluada mediante el índice de diversidad de Shannon-Wiener (H'):
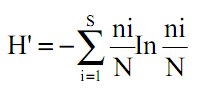
(Magurran, 1988). Para tener una estimación de la riqueza de la zona de estudio, se emplearon rarefacciones basadas en muestras con el estimador de riqueza Chao 2 (Gotelli & Colwell, 2001). Para ello se emplearon las 25 subparcelas en que se dividió la hectárea estudiada. Se utilizó el programa Estimates 8.0.0 (Colwell, 2006).
Para el estudio de los patrones espaciales se empleó la función K de Ripley (Fortin & Dale, 2005), que requiere las posiciones mapeadas de puntos (árboles, en este caso). El método consiste en realizar recuentos de los eventos (puntos) presentes en torno a cada uno de los puntos mapeados dentro de círculos de radio creciente. A diferencia de los métodos tradicionales, que solo permiten conocer los patrones espaciales a escalas pequeñas, este permite conocer una determinada distribución espacial a múltiples escalas. Esto resulta tanto más importante cuanto que el patrón espacial raramente es el mismo según se cambie de escala. De esta manera se pueden detectar diferentes procesos que actúan a diferentes niveles espaciales. Otra ventaja del método es que, para realizar las pruebas de significación, recurre a un proceso de aleatorizaciones repetidas (19, 99 o más veces), con el cual se crean intervalos de confianza dentro de cuyos límites el patrón estudiado se considera aleatorio. Para analizar los patrones espaciales empleamos 999 simulaciones Monte Carlo para determinar las bandas de confianza del 95%, utilizando el programa llamado Programita (Wiegand & Moloney, 2004). Diferentes modelos nulos existen en función de la pregunta biológica que se realice. El más simple es el de aleatoriedad espacial completa (CSR, de sus siglas en inglés), que comprueba si la distribución del organismo estudiado es aleatoria, agregada o uniforme. Existen otros más complejos, como el de independencia (Wiegand & Moloney, 2004), mediante el cual se pone a prueba la existencia o no de relación espacial entre dos tipos de puntos (en nuestro caso dos especies). Nosotros realizamos un análisis de patrones espaciales como un estudio exploratorio de algunos procesos que podrían estar actuando en la zona de estudio y para describir más exhaustivamente la estructura horizontal. Siguiendo procedimientos estándar en el estudio de bosques, sólo incluimos los árboles grandes (adultos) y vivos de la comunidad. Además, encontramos muchas más especies de las previstas, lo que hizo que el tamaño de la muestra (n) por especie fuese bajo (para estudios de patrones espaciales se recomienda un n de aproximadamente 60 individuos, T. Wiegand, com. pers.). Nosotros evaluamos los patrones espaciales sólo de las especies más abundantes (n desde 32 para Cupania vernalis Cambess, hasta 51 para Urera caracasana (Jacq.) Gaudich. ex Griseb.).
Resultados
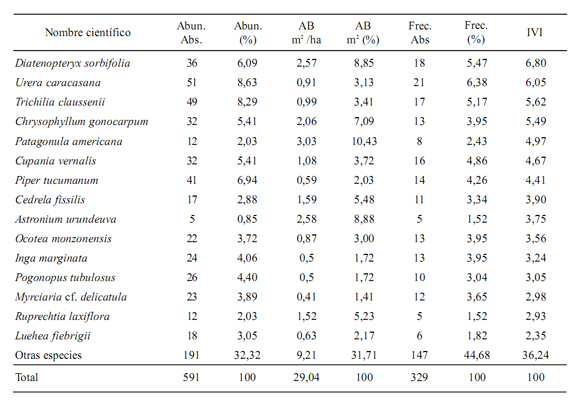
Diversidad florística. Los individuos con mayor densidad fueron Urera caracasana (8,63%), Trichilia claussenii C. DC. (8,29%), Piper tucumanum C. DC. (6,94%) y Diatenopteryx sorbifolia Radlk. (6,09%). Las demás especies se encontraron por debajo del 6%. El área basal o dominancia mayor fue para Patagonula americana L. (10,43%), Astronium urundeuva (Allemão) Engl. (8,88%), Diatenopteryx sorbifolia (8,85%) y Chrysophyllum gonocarpum (Mart. & Eichler ex Miq.) Engl. (7,09). Las demás especies presentaron una dominancia menor al 6%. Las especies mejor distribuidas (frecuentes) en la parcela son Urera caracasana (21/25), Diatenopteryx sorbifolia (18/25) y Trichilia claussenii (17/25). Las demás presentaron frecuencias por debajo del 16/25. El índice de valor de importancia por especie o mayor peso ecológico fue para Diatenopteryx sorbifolia (6,8%), Urera caracasana (6,05%), Trichilia claussenii (5,62%) y Chrysophyllum gonocarpum (5,49%). Las demás estuvieron por debajo del 5% (Tabla 1).
Tabla 1. Las 15 especies más importantes de la PPM ordenadas por el índice de valor de importancia (IVI).
El estimador de riqueza Chao 2 nos indica que la región posee unas 74 especies. Esto significa que en la zona de estudio se detectaron un 77% del total de las especies esperadas (Fig. 2). La diversidad, comparada con los bosques montanos húmedos y secos de Yungas, es relativamente alta, y solo ligeramente menor que la del bosque amazónico inundable (Tabla 2).
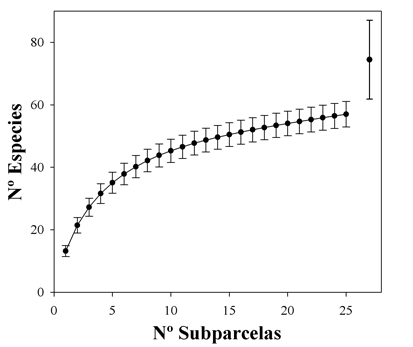
Fig. 2. Curva de rarefacción (más una desviación estándar) del área de estudio. El símbolo (más su desviación estándar) arriba a la derecha representa el estimador de riqueza Chao 2 para 25 subparcelas. El hecho que no se sobrepongan los intervalos de confianza del estimador y de la curva obtenida con los datos de campo indica que para tomar muestras adecuadamente estas comunidades tucumano-bolivianas a bajas altitudes se requieren más de una hectárea. Este es otro indicador de la alta diversidad de la zona.
Tabla 2. Diversidad florística en tres tipos de bosque: amazónico, montano húmedo y seco (en PPM de 1 ha).
Estructura horizontal y vertical. La curva de distribución de individuos por clase diamétrica a intervalos de 10 cm registró una gran cantidad de individuos en la clase inferior, decreciendo significativamente el número a medida que se incrementa el diámetro (Fig. 3A). La curva adopta una forma de "J" invertida, la que es característica de los bosques tropicales. La mayoría de individuos (65,65%) se registraron en la clase diamétrica de 10 a 20 cm, seguida del 17,6% en la clase de 20 a 30 cm. El resto de los individuos correspondió a las clases mayores a 30 cm (16,75%). La mayoría de las especies se registraron en la clase de 10 a 20 cm, exceptuando a Astronium urundeuva, Croton piluliferus Rusby, Ficus maroma A. Cast., Parapiptadenia excelsa (Griseb.) Burkart, Phoebe porphyria (Griseb.) Mez, Tabebuia ochracea (Cham.) Standl., Tipuana tipu (Benth.) Kuntze, Zanthoxylum rhoifolium Lam. y Zanthoxylum sp. Los árboles más gruesos son Astronium urundeuva y Patagonula americana L.
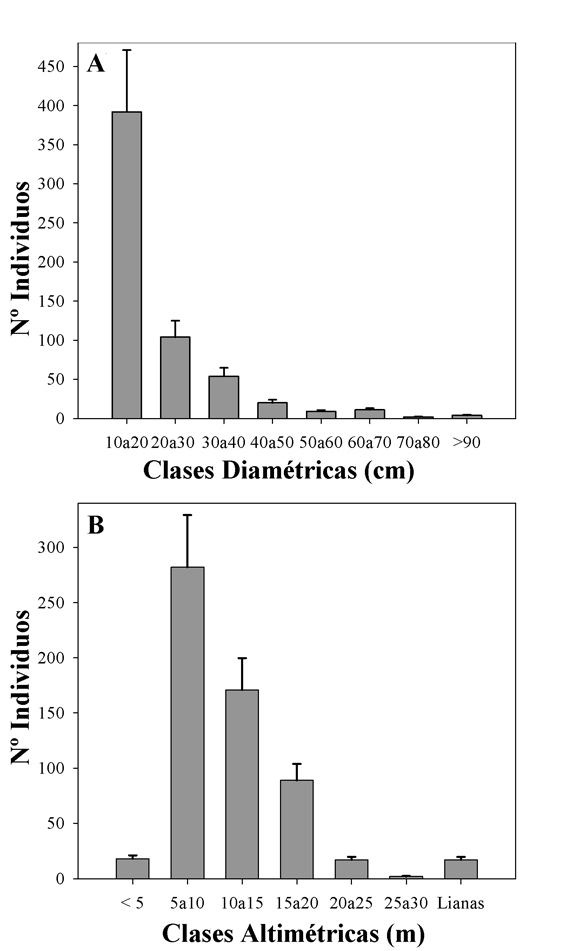
Fig. 3. Curva de distribución de individuos por clases. A, diamétricas. B, altimétricas (media ± desviación estándar).
En base a las alturas totales de los árboles se observan 6 estratos diferentes dentro del perfil vertical del bosque (Fig. 3B). Se puede observar que el máximo número de especies y de árboles se encuentra en el estrato 5 a 10 m (47,32%). En el estrato inferior se tiene un 3,02% y en el estrato superior un 0,34% de los individuos. Las especies que dominan los estratos inferiores son Urera caracasana, Allophylus edulis (A. St.-Hil., Cambess. & A. Juss.) Radlk., Cupania vernalis, Piper tucumanum, Casearia sylvestris Sw. y Sebastiania fiebrigii Pax. Los árboles mas altos son Patagonula americana y Anadenanthera colubrina (Vell.) Brenan.
Posición de la copa. Con respecto a la exposición lumínica de las copas se tiene al 13,33% de los individuos en la categoría 1 (copa emergente con luz directa), 18,25% en la categoría 2 (plena iluminación superior), 29,47% en la categoría 3 (alguna iluminación superior), 34,04% en la categoría 4 (alguna luz lateral) y el 4,91 en la categoría 5 (ausencia de luz directa). Agrupando las categorías que indican una débil exposición a la luz (3, 4 y 5), se tiene el 68,42% de los individuos en esta condición, y el 31,58% de los individuos goza de buena exposición a la luz.
Las especies con mejor posición de copa son Cedrela fissilis, Ocotea monzonensis Mez, Myroxylon peruiferum L. f., Anadenanthera colubrina, Cupania vernalis, Nectandra sp., Parapiptadenia excelsa, Patagonula americana, Ruprechtia laxiflora Meisn., Astronium urundeuva, Solanum riparium Pers., Inga marginata Willd., Blepharocalyx salicifolius, Cordia trichotoma (Vell.) Arráb. ex Steud., Lonchocarpus lilloi (Hassl.) Burkart, Chrysophyllum gonocarpum, Chrysophyllum marginatum (Hook. & Arn.) Radlk., Croton piluliferus, Myrciaria cf. delicatula (DC.) O. Berg, Phoebe porphyria (Griseb.) Mez, entre otros. Las especies Urera caracasana, Pogonopus tubulosus (A. Rich.) K. Schum., Cupania vernalis, Trichilia claussenii, Diatenopteryx sorbifolia, Luehea fiebrigii Burret, Myrciaria cf. delicatula, Roupala meisneri Sleumer, Casearia sylvestris, Inga marginata, Myrcianthes pungens (O. Berg) D. Legrand y Piper tucumanum presentan una copa sin incidencia de luz directa.
Forma de copa. Según la forma de la copa se observó que el 0,18% de los individuos se encuentran en la categoría 1 (copa perfecta), 47,89% en la categoría 2 (copa buena), 47,02% en la categoría 3 (copa tolerable), 2,81% en la categoría 4 (copa pobre), y en la categoría 5 (muy pobre), el 2,11%. El 48,07% de los individuos presentó una copa bien formada (categoría 1 y 2). La especie con mejor forma de copa es Anadenanthera colubrina. Las especies que presentaron individuos en la categoría 5 (muy pobre) son Urera caracasana, Cupania vernalis y Sebastiania fiebrigii. Asimismo, las 15 especies principales (ver Tabla 1) registraron categorías 2 y 3 (buena y tolerable) con excepción de Astronium urundeuva que solo registró forma de copa buena.
Infestación de lianas. La mayoría de las especies presenta ausencia de lianas en un 21,23% (nivel 1) y tolerable en un 44,39% (nivel 2), y en su conjunto suman 65,61% de los individuos que no están afectados por las lianas y 26,32% (nivel 3) presentan lianas en el fuste y copa.
Patrón espacial. El patrón espacial del conjunto de individuos de la comunidad se presenta aleatorio a todas las escalas espaciales consideradas, salvo la más pequeña, donde exhibe una ligera tendencia a la agregación (Fig. 4). El análisis por separado de las clases de tamaño (diámetro) muestra un patrón también aleatorio, aunque es interesante notar que la distribución espacial de los árboles más pequeños tiende un poco más a la agregación (Fig. 4). En cambio, los patrones espaciales de las especies con mayor densidad en la PPM muestran por lo menos una tendencia a la agregación a alguna escala espacial (Fig. 5). Por otro lado, no parecen existir relaciones espaciales de atracción o repulsión entre las especies más abundantes, salvo para el par Cupania-Trichilia (atracción) (Fig. 6).

Fig. 4. Patrón espacial general (todos los individuos) y por clase diamétrica en la PPM. Las líneas gruesas muestran las funciones de distribución empíricas (función L de Ripley), y las líneas discontinuas señalan los intervalos de confianza del 95% para la hipótesis de aleatoriedad. Cuando la curva gruesa sale de los limites del intervalo de confianza por arriba, este indica la existencia de un patrón agregado; y por debajo, de un patrón uniforme.
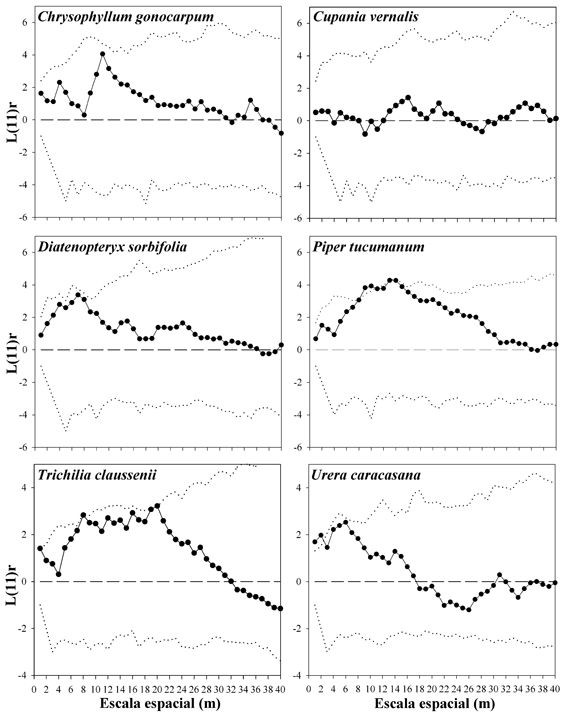
Fig. 5. Patrón espacial de las especies más abundantes en la PPM. Las curvas tienen el mismo significado que en la Fig. 4.
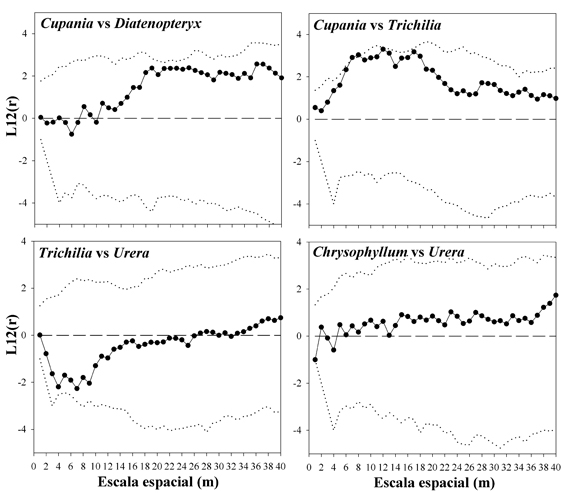
Fig. 6. Patrón espacial de pares de especies (atracción-repulsión), en la PPM. Cuando la curva gruesa excede los limites del intervalo de confianza (curva punteada) por la parte superior, existe atracción entre las especies; si es por la parte inferior, existe en cambio repulsión.
Discusión
La zona de estudio se encuentra en un bosque montano de transición semihúmedo, que ocupa sectores entre 400 y 900 m en el pedemonte de serranías de escasa altitud (ca. 2000 m). Los estudios de Brown & Grau (1993, 1995), Brown et al. (2001) y Malizia et al. (2006) indican que algunas especies registradas en este estudio son indicadoras de la selva pedemontana. Tal es el caso de especies caducifolias dominantes, como Anadenanthera colubrina, Myroxylon peruiferum, Cordia trichotoma, Patagonula americana, Calycophyllum multiflorum Griseb. y Astronium urundeuva. Sin embargo, no se registraron Phyllostylon rhamnoides (J. Poiss.) Taub. ni Tabebuia impetiginosa (Mart. ex DC.) Standl. en la PPM, que son citados por los mismos autores, y sí se registraron otras especies de los géneros Blepharocalyx, Chrysophyllum, Inga, Nectandra, Ocotea y Piper, que son indicadoras del bosque tucumano-boliviano. Esto sugiere que las comunidades de esta región son todavía una variante del tucumano-boliviano más que bosques secos del llamado Arco Pleistocénico (Prado & Gibbs, 1993), los cuales posiblemente son más propios de las regiones más externas de los Andes (por ejemplo, Villa Montes, Yacuiba, ambos en el departamento de Tarija, Bolivia).
Los datos obtenidos muestran una semejanza con la composición y estructura con otras parcelas permanentes realizadas en el noroeste de Argentina y sur de Bolivia a altitudes de 600 a 1.100 m (Malizia, datos inéditos) y una riqueza mayor de nuestra parcela a nivel familiar y específico en comparación con los promedios por hectárea para estos taxones reportados por Malizia et al. (2006). Las especies mas importantes dentro de esta formación en parámetros de densidad, cobertura y frecuencia fueron Diatenopteryx sorbifolia, Urera caracasana, Trichilia claussenii, Chrysophyllum gonocarpum, Patagonula americana, Cupania vernalis y Piper tucumanum. En las PPMs reportadas por Malizia et al. (2006) en la selva pedemontana (ca. 600 m), las especies principales fueron Anadenanthera colubrina, Phyllostylon rhamnoides, Trichilia hieronymi (= T. claussenii) y Parapiptadenia excelsa, que concuerdan en composición con la parcela del presente estudio, aunque con densidades específicas diferentes. Phyllostylon rhamnoides no se registró en nuestro trabajo. Estas diferencias se pueden deber a que Malizia trabajó cerca de la llanura, en el verdadero bosque de transición, donde predominan las especies caducifolias. También se pueden invocar razones de una historia distinta de las dos regiones. Por otro lado, Malizia (Datos Red Subtropical de Parcelas Permanentes de la Fundación ProYungas) caracterizó, como especies frecuentes de un intervalo altitudinal de 550 a 829 m en 26 PPMs, a Parapiptadenia excelsa y Anadenanthera colubrina, con una frecuencia de 25/26 y 23/26, respectivamente; Myroxylon peruiferum, Ocotea puberula (Rich.) Nees, Terminalia triflora (Griseb.) Lillo y Urera caracasana (22/26); Trichilia hieronymi (21/26); Cupania vernalis (19/26); Chrysophyllum gonocarpum y Diatenopteryx sorbifolia (18/26); Piper tucumanum (16/26), lo cual concuerdan con las seis especies en nuestro estudio que están mejor representadas en la parcela, aunque no con las dos especies principales mencionadas por Malizia (Parapiptadenia excelsa y Anadenanthera colubrina), que son características del bosque pedemontano y que son especies pioneras que se desarrollan con mucho éxito después de una perturbación. De hecho, esta característica ha originado una estructura boscosa con pocas perturbaciones por la poca incidencia de lianas y competencia con los árboles por la luz, humedad y nutrientes. Asimismo, la baja densidad de especies pioneras registradas en nuestra parcela, nos indica que la zona de estudio contiene un bosque relativamente poco perturbado.
Existen más familias y especies registradas en la PPM estudiada que en el bosque tucumano-boliviano citado por Malizia et al. (2006), 57 especies y 31 familias en nuestra parcela frente a 39 ±2,9 especies y 22,1 ±11,5 familias para el caso de los bosques estudiados por Malizia (n=8 ha y 78 especies) y casi tantas como en los bosques secos tropicales de la diversa zona del parque Madidi (La Paz), que tiene de 24 a 27 familias y 55 a 60 especies (Cayola et al., 2005; Paredes, 2008; Uzquiano, 2008). El valor calculado del índice de diversidad en la PPM es relativamente alta (H'=3,5) comparada con algunos bosques montanos: del tucumano boliviano de la Argentina, y del norte de Bolivia (los Yungas). Por otro lado, si la comparamos con algunos trabajos en el bosque amazónico, la diversidad registrada es similar para el bosque de tierra firme y relativamente baja en comparación con el bosque inundable. El elevado índice de diversidad (H') es debido a una alta densidad y no al número elevado de especies. En la PPM se encontraron 12 especies raras (un individuo) y alrededor de 27 especies con menos de 10 individuos y seis especies con más de 30 individuos, siendo estas las 6 principales especies que están mejor distribuidas en la PPM. Todo esto indica que el bosque tucumano-boliviano de Tarija a altitudes inferiores a los 1000 m es una comunidad vegetal diversa.
El valor estimado de riqueza (Chao 2) es similar al número total de especies para la selva pedemontana reportado por Malizia et al. (2006). Sin embargo, Malizia obtuvo esa riqueza muestreando en ocho parcelas de una hectárea distribuidas en diferentes lugares, lo que brinda una idea del tamaño de muestra necesario para registrar la mayoría de las especies.
Las familias mejor representadas en términos de densidad, cobertura y diversidad fueron Sapindaceae y Meliaceae; la especie mejor representada fue Urera caracasana, salvo para el criterio de cobertura, parámetro para el cual los individuos con mayor área basal fueron Patagonula americana y Astronium urundeuva. El valor de área basal hallado de 29,04 m2 /ha es un poco superior al reportado en el bosque de tierra firme e inundable amazónico, con 26,8 y 25,6 m2 /ha, respectivamente, y al reportado por Malizia et al. (2006), de 25,7±1.3 a 27,5±2,1 m2 /ha; muy similar a los registrados por otros autores en el bosque seco del valle del Tuichi (La Paz, Bolivia), de 20 a 35.4 m2 /ha (Cayola et al., 2005; Paredes, 2008; Uzquiano, 2008). Este es otro indicador del estado relativamente maduro del bosque.
Las especies con mayor densidad en la PPM como Urera caracasana, Trichilia claussenii y Piper tucumanum, podrían estar mejor adaptadas a las variables físicas (topografía) y climáticas (temperatura y precipitación), que son las que mas influyen en la distribución espacial del bosque montano. Por otro lado, el tipo de semillas puede ayudar a una mejor dispersión en algunas especies que están adaptadas para un mayor éxito en su establecimiento por ejemplo, aquenios en Urera caracasana con distribución continua desde México a Sudamérica y registrada en la amazonia hasta los bosques montanos semideciduos y en vegetación secundaria. Trichilia claussenii (cápsula) presenta una dispersión zoocorica y es una especie de amplia distribución y característica de los bosques del subtrópico de tierras bajas (Brazil, Paraguay, noreste de Argentina) y de los bosques montanos del sur de Bolivia y noroeste argentino. Piper tucumanum (drupa) característico del sotobosque y rápido crecimiento conocida solamente en el bosque tucumano-boliviano. Otro factor para el éxito de estas especies es debido a que comparten patrones similares de exigencias de radiación lumínica, regeneración y crecimiento, clasificándose como esciófitas parciales y/o totales por ser de estratos inferiores y ser tolerantes a la poca luz para su desarrollo. Asimismo, las seis especies mejor representadas en la parcela presentan una buena exposición y forma de copa, y resultaron ser las que están mejor adaptadas y son tolerantes a la sombra.
Para el caso de los patrones espaciales de las especies, en principio creímos que un área de estudio de una hectárea podría ser lo suficientemente grande para realizar estos estudios. Así, el conjunto de árboles, de la hectárea permitió establecer claramente el patrón espacial aleatorio, que en general es característico de diferentes tipos de bosque (Mueller-Dombois & Ellenberg, 1974). De hecho, los métodos de distancia que se emplean para calcular densidad tienen como supuesto esa distribución aleatoria de los individuos de una comunidad independientemente de la especie. No obstante, como ya se demostró, la diversidad de la comunidad estudiada resultó ser bastante elevada. Esto implicó que se encontraron pocos individuos por especie. Nosotros realizamos el análisis del patrón espacial de las más abundantes, pero incluso así, los tamaños muestrales fueron relativamente bajos. Lo que indican estos análisis es que la mayoría de las especies tienen agregación a alguna escala (que en general va de pequeña, como en U. caracasana, a intermedia, como en P. tucumanum). Probablemente éstas se relacionan con las distancias de dispersión de los propágulos. No tenemos una explicación plausible para la asociación entre C. vernalis y T. clausenii. Podría tratarse de similares requerimientos abióticos de ambas especies. Los datos de este estudio pueden servir de referentes para realizar estudios más específicos encaminados a explicar estos patrones.
Agradecimientos
Al Herbario Nacional de Bolivia, Centro de Análisis Espacial, Estación Experimental Río Conchas dependiente de la Facultad de Ciencias Agrícolas y Forestales (FCAF), al proyecto Diagnóstico de la Biodiversidad PEA-Bermejo, Orlando Orerazo (FCAF), Lucio Malizia y Violeta Carilla (Fundación Proyungas). AYasmani Balanza, Jonny Flores, Víctor Hugo Schmidt, Alexandy Sejas (UJMS Tarija), Abraham Poma (UMSA La Paz) y Mario Chavarria de la Comunidad Río Conchas. Agradecemos a los revisores anónimos quienes realizaron correcciones en el texto.
Bibliografía
1. Bascope, F. & P. Jorgensen. 2005. Caracterización de un bosque montano húmedo: Yungas, La Paz. Ecología en Bolivia 40: 365-379. [ Links ]
2. Brown, A. D. & H. R.Grau. 1993. La naturaleza y el hombre en las selvas de montaña. Proyecto GTZ - Desarrollo Agroforestal en Comunidades Rurales del Noroeste Argentino. Salta. [ Links ]
3. Brown, A. D. & H. R. Grau (eds.). 1995. Investigación, conservación y desarrollo en selvas subtropicales de montaña. Laboratorio de investigaciones ecológicas de las Yungas. Universidad Nacional de Tucumán. Tucumán. [ Links ]
4. Brown, A. D.; H. R. Grau, L.R. Malizia & A. Grau. 2001. Argentina, en M. Kappelle & A.D. Brown (eds.) Bosques nublados del Neotropico. Pp. 623-659. INBio. Costa Rica. [ Links ]
5. Cabrera, H. 2005. Diversidad florística de un bosque montano en los Andes tropicales del noroeste de Bolivia. Ecología en Bolivia 40: 380-395. [ Links ]
6. Cayola, L.; A. Fuentes & P. Jorgensen. 2005. Estructura y composición florística de un bosque seco subandino yungueño en el valle del Tuichi, Área Natural de Manejo Integrado Madidi. Ecología en Bolivia 40: 396-417. [ Links ]
7. Colwell R.K. 2006. EstimateS: Statistical estimation of species richness and shared species from samples. Disponible en http://purl.oclc.org/estimates. [ Links ]
8. Curtis, J. T. & R. P. McIntosh. 1951. An upland forest continuum in the praire-forest border Region of Wisconsin. Ecology 32: 476-496. [ Links ]
9. Dallmeier, F. (ed.). 1992. Long-term monitoring of biological diversity in tropical forest areas: methods for establishment and inventory of permanent plots. Paris: MAB Digest 11. UNESCO. [ Links ]
10. Dawkins, H. C. 1958. The management of natural tropical high-forest with special reference to Uganda. Oxford: Imperial Forestry Institute. Paper N° 34. University of Oxford. [ Links ]
11. Fortin, M. J. & M. Dale. 2005. Spatial analysis: a guide for ecologists. Cambridge University Press, Cambridge. [ Links ]
12. Gotelli, N. J. & R. K. Colwell. 2001. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters 4: 379-391. [ Links ]
13. Hueck, K. 1978. Los bosques de Sudamérica. Berlín: Agencia Alemana de Cooperación Técnica (GTZ). [ Links ]
14. Ibisch, P. L.; S. G. Beck, B. Gerkmann, & A. Carretero. 2003. Ecorregiones y Ecosistemas, en P. L. Ibisch & G. Mérida (eds.). Biodiversidad: La riqueza de Bolivia. Estado del conocimiento y conservación. Pp. 47-88. Santa Cruz de la Sierra: Ministerio de Desarrollo Sostenible, Editorial FAN. [ Links ]
15. Lamprecht, H. 1990. Silvicultura en los trópicos: los ecosistemas forestales en los bosques tropicales y sus especies arbóreas, posibilidades y métodos para un aprovechamiento sostenido. Berlín: Sociedad Alemana de Cooperación Técnica (GTZ). [ Links ]
16. Lowe, R. & P. Walker. 1977. Classification of canopy, stem, crow status and climber infestation in natural tropical forest in Nigeria. Journal of Apliedd Ecology 14: 897-903. [ Links ]
17. Magurran, A. E. 1988. Ecological diversity and its measurement. London: Chapman and Hall. [ Links ]
18. Malizia, L. R.; C. Blundo & S. Pacheco. 2006. Diversidad, estructura y distribución de bosques con cedro en el noroeste de Argentina y sur de Bolivia, en S. Pacheco & A. Brown (eds.). Ecología y producción de cedro (genero Cedrela) en las Yungas australes. Pp. 83-104. San Miguel de Tucumán: Laboratorio de investigaciones ecológicas de las Yungas y ProYungas. [ Links ]
19. Mueller-Dombois, D. & H. Ellenberg, 1974. Aims and methods of vegetation ecology. Nueva York: John Wiley & Sons. [ Links ]
20. Paredes, S. 2008. Estructura y composición florística de un bosque subandino en el Parque Nacional y Área de Manejo Integrado Madidi. Tesis de Licenciatura, Facultad de Agronomía, Universidad Mayor de San Andrés. La Paz. [ Links ]
21. Prado, D. E. & P. E. Gibbs 1993. Patterns of species distributions in the dry seasonal forests of South America. Annals of the Missouri Botanical Garden 80: 902-927. [ Links ]
22. Poma, A. S. 2007. Estructura y composición florística en dos parcelas permanentes en el bosque amazónico de tierra firme e inundable, en el norte del Parque Nacional Madidi, La Paz. Tesis de Licenciatura, Facultad de Agronomía, Universidad Mayor de San Andrés. La Paz. [ Links ]
23. Uzquiano, J. R. 2008. Aspectos estructurales y florísticos de un bosque seco subandino al sur del Área Natural de Manejo Integrado Madidi, La Paz. Tesis de Licenciatura, Facultad de Ciencias Puras y Naturales, Universidad Mayor de San Andrés. La Paz. [ Links ]
24. Wiegand, T. & A. Moloney. 2004. Rings, circles and nullmodels for point pattern analysis in ecology. OIKOS 104:209-229. [ Links ]
25. Zenteno-Ruiz F. S.; S. Beck, R. López & S. Gallegos. 2010. Caracterización de la vegetación y de la diversidad florística de la cuenca alta del río Bermejo, Tarija: Implicaciones para su conservación, en Beck et al. (eds.). Biodiversidad y ecología en Bolivia - Simposio de los 30 años del Instituto de Ecología. Pp. 162-173. La Paz: Instituto de Ecología, Universidad Mayor de San Andrés. [ Links ]

