










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkDarwiniana, nueva serie
versión impresa ISSN 0011-6793
Darwiniana, nueva serie vol.1 no.2 San Isidro dic. 2013
ARTÍCULO INVITADO
Acerca de la flora de Antofagasta de la Sierra, Catamarca, Argentina. Ambiente y paleoambiente
M. Fernanda Rodríguez
Instituto Nacional de Antropología y Pensamiento Latinoamericano, 3 de Febrero 1378, C1426BJN Ciudad Autónoma de Buenos Aires, Argentina; mfrodriguez18@gmail.com
Editor Asociado: Sandra S. Aliscioni
Original recibido el 4 de junio de 2013,
aceptado el 10 de noviembre de 2013
Resumen
Se presenta una síntesis de los trabajos realizados sobre la identificación, anatomía y usos de especies vegetales, actuales y recuperadas en el marco de las investigaciones arqueobotánicas, llevadas a cabo en Antofagasta de la Sierra durante las campañas arqueológicas realizadas en los años 1994-2000. Se describen las transectas realizadas a partir de los sitios arqueológicos estudiados, las asociaciones vegetales características y se presenta un listado de las especies vegetales halladas. Se describe el uso actual y se señala el nombre vernáculo para cada taxón. Luego, se presenta una descripción anatómica de tallo, raíz u hoja, dependiendo del caso, de cada uno de los taxones presentes en los sitios arqueológicos, identificados por medio de la comparación anatómica con material actual de referencia; se presentan las correspondientes fotomicrografías obtenidas con microscopio óptico y/o microscopio electrónico de barrido. Finalmente, se discute el uso, en la actualidad y en el pasado, de las especies recuperadas en los sitios arqueológicos del área de estudio, destacando aquellos aspectos que suponen cambio y continuidad a través del tiempo.
Palabras clave: Ambiente; Anatomía vegetal; Especies herbáceas; Especies leñosas; Paleoambiente; Puna meridional argentina.
This paper integrates archaeobotanical research conducted in Antofagasta de la Sierra based on the identification, anatomy, and use of both present plant resources and archaeological plant remains, recovered during fieldwork between 1994 and 2000. Transects starting from the archaeological sites, the surrounding plant communities, and the species found are described. Characterization of present plant diversity also included the popular name and actual use of every species. Archaeological plant remains were identified by anatomy in comparison with present plant species in the studied area and their anatomical characters are here described and documented with digital images taken with compound microscope (bright field) and scanning electron microscope. Finally, discussion includes past and present use of every species recovered in the archaeological sites from Antofagasta de la Sierra, highlighting those aspects that imply change or continuity through time.
Keywords: Argentine Southern Puna; Environment; Herbaceous species; Palaeo-environment; Plant anatomy; Woody species.
INTRODUCCIÓN
La localidad de Antofagasta de la Sierra se ubica en la provincia argentina de Catamarca (26° 11' 16'' S y 67° 20' 51.3'' O). El área es una cuenca endorreica que tiene como centro la Laguna de Antofagasta (3450 m s.m.) y alcanza los 5000 m s.m. en las montañas y volcanes que la rodean (Figs. 1 y 2). El clima es árido, de tipo andino puneño, con un promedio anual de precipitaciones de 100 mm concentradas en la estación cálida. Las temperaturas mínimas son menores a los 10°C (Cabrera, 1976; García Salemi, 1986). La radiación solar y la evaporación son elevadas, lo mismo ocurre con la amplitud térmica diaria y anual. El drenaje depende de la fusión de los hielos en el verano.

Fig. 1. Antofagasta de la Sierra, Catamarca. Transectas (señaladas con flechas): T1 - T10. Localidades arqueológicas: Quebrada Seca, Punta de la Peña, Peñas de las Trampas, Peñas Chicas, Peñas Coloradas y Peña de la Cruz. Figura en color en la versión online http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/529/569.

Fig. 2. Microambientes correspondientes a las transectas realizadas a partir del sitio Quebrada Seca 3. A, transecta 1 (vega). B, transecta 2 (vega). C, transecta 3 (pajonal). D, transecta 4 (tolar). Figura en color en la versión online http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/529/569.
Desde el punto de vista fitogeográfico, Antofagasta de la Sierra se ubica en la Puna salada (Troll, 1958) o Puna de Atacama (Baied & Wheeler, 1993), que corresponde a la Provincia Puneña del Dominio Andino (Cabrera, 1976). La vegetación dominante es la estepa arbustiva, no obstante, también se desarrollan la estepa herbácea, psammófila y halófila. En el área se distinguen tres tipos de asociaciones vegetales (Fig. 2): pajonal, tolar y vega (Cabrera, 1976; Cabrera & Willink, 1973; Haber, 1992). El pajonal se desarrolla por encima de los 3800 m s.m., donde abundan diferentes especies de gramíneas de los géneros Festuca L., Pappostipa (Speg.) Romasch., P. M. Peterson & Soreng y Deyeuxia P. Beauv. y otras arbustivas de los géneros Adesmia DC., Baccharis L., Parastrephia Nutt. y Fabiana Ruiz & Pav. También por encima de los 3800 m s.m., se ubica el tolar, donde son frecuentes especies arbustivas y subarbustivas de los géneros Parastrephia y Acantholippia Griseb. La vega se ubica a lo largo del curso del Río Las Pitas y en el fondo de la Quebrada Seca y su cubierta vegetal incluye gramíneas y juncáceas (Rodríguez & Rúgolo de Agrasar, 1999; Rodríguez, 2008).
Es importante citar otras investigaciones en relación con la flora del área. En este sentido, se destacan los relevamientos y listados de especies presentados por Haber (com. per.), Elkin (1987, 1992), Rodríguez (1998), Cuello (2006) y Olivera (2006). Recientemente, Mondini (com. per.) realizó un nuevo estudio en Antofagasta de la Sierra con el objetivo de analizar los isótopos estables de las especies vegetales que crecen en la zona.
Los sitios arqueológicos estudiados en Antofagasta de la Sierra, fueron habitados a lo largo del Holoceno temprano, medio y tardío, durante el Fig. 1. Antofagasta de la Sierra, Catamarca. Transectas (señaladas con flechas): T1 - T10. Localidades arqueológicas: Quebrada Seca, Punta de la Peña, Peñas de las Trampas, Peñas Chicas, Peñas Coloradas y Peña de la Cruz. Figura en color en la versión online http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/529/569.
lapso comprendido entre ca. 10000-400 años AP, por grupos humanos cazadores-recolectores, quienes, a lo largo del tiempo, fueron complementando estas actividades con el pastoralismo y una agricultura incipiente (Rodríguez & Aschero, 2007). El estudio de estas poblaciones reveló la continuidad de las ocupaciones en la microrregión. Algunos sitios, tales como QS3, CS1, PP4, PT1.1, PCz1, PCH1.1, PCH1.3 y CC1A (véase Descripción del área de estudio) muestran tal continuidad, confirmada por el registro radio-carbónico (Aschero, 2010). Es interesante destacar que, aún frente a las condiciones adversas del Altitermal u Holoceno medio (8000-6000 AP), el área continuó siendo habitada, razón por la cual Nuñez et al. (2006) la denominaron eco-refugio. La importancia de esta persistencia espacial se vincula con el proceso posterior de cambio hacia economías de producción pastoril y agrícola a partir de las poblaciones de cazadores-recolectores afincadas en la región. Las mismas constituyeron una base poblacional ineludible para establecer las condiciones de inicio de dicho proceso (Aschero, 2010).
El objetivo general de este trabajo es ofrecer una síntesis de los trabajos realizados por la autora en relación a la identificación, anatomía y usos de especies vegetales en el área de Antofagasta de la Sierra, en el marco de las investigaciones arqueobotánicas allí realizadas. Se detallan los taxones presentes en la flora actual del área y, al mismo tiempo, en los sitios arqueológicos relevados. De este modo, se contrasta el ambiente y el paleoambiente a través de la vegetación, así como también se indaga acerca de los usos de las plantas tanto en el presente como en el pasado.
Descripción del área de estudio: sitios arqueológicos
Los sitios estudiados corresponden a las siguientes localidades arqueológicas del área de Antofagasta de la Sierra (Figs. 1 y 3; Tabla 1):

Fig. 3. Sitios arqueológicos excavados en el área de estudio. A, Punta de la Peña 4 (PP4). B, Cueva Salamanca 1 (CS1). C-D, Quebrada Seca 3 (QS3). Figura en color en la versión online http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/529/569.
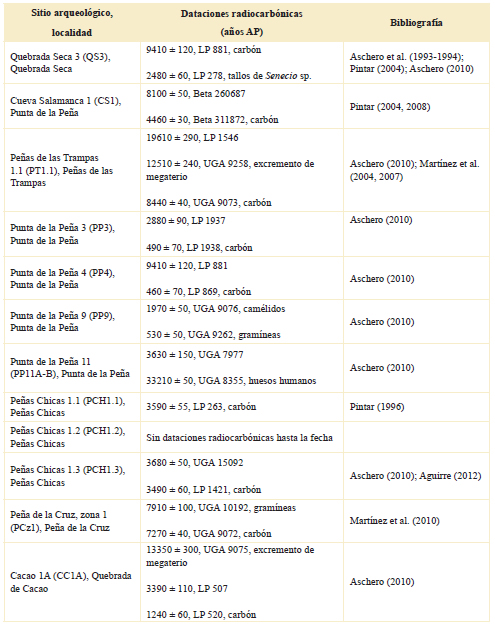
Tabla 1. Sitios arqueológicos y sus correspondientes dataciones radiocarbónicas. Para cada uno de ellos solo se indican algunas dataciones, en la mayoría la más temprana y la más tardía.
1. Localidad arqueológica de Quebrada Seca (3800-4200 m s.m.), en el sector del cauce de Quebrada Seca y afluente del Río Las Pitas. Se estudió aquí el sitio Quebrada Seca 3 (QS3), en la asociación vegetal del pajonal (Elkin, 1992; Aschero et al., 1993-1994; Rodríguez 1998, 2000, entre otros).
2- El sector del curso medio-inferior del Río Las Pitas con las localidades arqueológicas de Peñas de las Trampas, Peñas Chicas, Peñas Coloradas y Punta de la Peña (3500-3800 m s.m.) y los sitios: Cueva Salamanca 1, Peñas de las Trampas 1.1, Punta de la Peña 3, Punta de la Peña 4, Punta de la Peña 9, Punta de la Peña 11 A y B y Peñas Chicas 1.1, 1.2 y 1.3 (CS1, PT1.1, PP3, PP4, PP9, PP11A-B, PCH1.1, PCH1.2, PCH1.3, respectivamente) (Aschero, 1999; Martínez et al., 2004, 2007, 2010; Pintar, 2004, 2008; Rodríguez et al., 2003, 2006; Rodríguez & Aschero, 2007; entre otros ). A este sector corresponde la asociación vegetal del tolar.
3- La Quebrada del Río Ilanco, seco en la actualidad, en donde se localizaron aleros con ocupaciones humanas durante las prospecciones realizadas en diciembre de 1998 en la localidad arqueológica de Peña de la Cruz. De estos sitios interesa especialmente Peña de la Cruz, zona 1 (PCz1), ubicado a 3663 m s.m. en la asociación vegetal del tolar (Martínez, com. per.).
4- La Quebrada de Cacao (4000 m s.m.). Aquí se estudió el sitio Cacao 1A (CC1A) con destacadas representaciones de arte rupestre (Olivera et al., 2003). Asociación vegetal: vega.
MATERIALES Y MÉTODOS
Se realizaron diez transectas a partir de los sitios arqueológicos estudiados (Rodríguez, 2004a) (Fig. 1; Tabla 2). En cada transecta se documentaron las especies vegetales y se depositó un ejemplar de referencia en el herbario del Instituto de Botánica Darwinion, SI (Thiers, 2013). En algunos pocos casos se consideraron especímenes colectados por otros autores (indicado en la Tabla 2). Asimismo, se registraron las asociaciones de especies vegetales más frecuentes en las transectas realizadas (Fig. 2), siguiendo el criterio de Cabrera (1976).
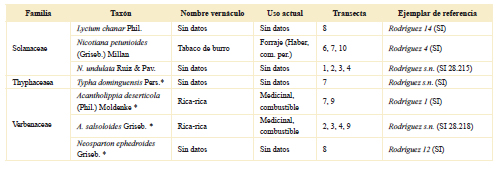
Tabla 2. Taxones hallados en el área de estudio ordenados por familia; se indica nombre vernáculo, uso actual, transecta donde se coleccionó y ejemplar de referencia. El asterisco destaca las especies que fueron recuperadas en los sitios arqueológicos del área de estudio.


Las cuatro primeras transectas se realizaron a partir de la localidad de Quebrada Seca (3800-4200 m s.m.) como se indica a continuación: Transecta 1. Vega de Quebrada Seca - Ojo de Quebrada Seca (se denomina así a la aguada en la que nace la vega). Asociación vegetal: vega (Figs. 1 y 2 A). Transecta 2. Vega de Quebrada Seca - Las Juntas. En este punto se unen los cursos de agua que corren por la Quebrada de Real Grande y por Quebrada Seca para formar el curso medio del Río Las Pitas. Asociación vegetal: vega (Figs. 1 y 2 B). Transecta 3. Quebrada Seca - Quebrada de Real Grande, pasando por la Quebrada de las Vizcachas y la pampa que separa a las dos primeras quebradas. Asociación vegetal: pajonal (Figs. 1 y 2 C). Transecta 4. Quebrada Seca - Punta de la Peña y Peñas Chicas. Asociación vegetal: pajonal y tolar (Figs. 1 y 2 D).
Las transectas 5 y 6 se vincularon con la localidad de Punta de la Peña (3600-3800 m s.m.): Transecta 5. Punta de la Peña - Las Juntas, siguiendo el curso del Río Las Pitas (3600 - 3800 m s.m.). Asociación vegetal: vega. Transecta 6. Punta de la Peña - Peñas Chicas - Peñas Coloradas, márgenes del Río Las Pitas. (3600 m s.m.). Asociación vegetal: vega.
Las restantes transectas (7 a 10) se relacionaron con la Cuenca del Río Las Pitas (7-9) y con la Quebrada del Río Ilanco (10): Transecta 7. Laguna de Antofagasta - Volcán Antofagasta (3400 m s. m). Asociación vegetal: borde de laguna. Transecta 8. Río Punilla - Río Miriguaca (3400 m s.m.). Asociación vegetal: vega. Transecta 9. Quebrada de Cacao (4000 m s.m.). Asociación vegetal: vega. Transecta 10. Quebrada del Río Ilanco - Vega de Ilanco (3500 - 3900 m s.m.). Asociación vegetal: tolar y vega.
Para cada especie hallada en el área se indica el uso actual y el nombre vernáculo (Tabla 2). Esta información se obtuvo a partir de entrevistas realizadas a antiguos pobladores de la zona. Es importante aclarar que no se trató de un trabajo etnobotánico sistemático, sino que las entrevistas se llevaron a cabo con el objetivo de obtener datos que permitieran inferir los usos de las especies halladas en el registro arqueobotánico del área (Rodríguez, 1998).
Los restos vegetales arqueológicos hallados en los sitios estudiados fueron identificados a partir del material vegetal actual de referencia. Se confeccionó una histoteca de referencia para el material actual y el arqueológico.
Se realizó el estudio anatómico de aquellos taxones presentes tanto en los sitios arqueológicos del área de estudio como en las transectas. Teniendo en cuenta el uso de los taxones en el pasado, los mismos se agrupan en dos categorías: especies leñosas (combustibles y/o confección de tecnofacturas) y especies herbáceas (pisos de ocupación, confección de artefactos y algunas comestibles como la quínoa). Este agrupamiento resultó útil en investigaciones etnobotánicas y arqueobotánicas posteriores.
Se presenta una descripción anatómica (sobre la base de tallos, raíz u hojas) de cada uno de los taxones con las correspondientes fotomicrografías obtenidas con microscopio óptico y/o microscopio electrónico de barrido (MEB). Estas descripciones se realizan a partir de los especímenes actuales y arqueológicos cuyas características coinciden. Las descripciones anatómicas de tallos y, en un caso también de la raíz, constituyen una síntesis realizada a partir de trabajos previos (Rodríguez, 2000, 2004b y 2008). En el caso de los taxones herbáceos las descripciones anatómicas de tallos u hojas, según las especies, se basan en trabajos previos (Rodríguez, 1999, 2008; Rodríguez et al., 2003, 2006), con la excepción de Festuca chrysophylla, F. dissitiflora, F. orthophylla, F. weberbaueri, Pappostipa frigida y Puccinellia frigida, que se presentan por primera vez en este trabajo.
Técnicas de anatomía utilizadas para material actual y arqueológico
Del material de las especies leñosas coleccionadas en el área y de aquellas recuperadas en los sitios arqueológicos, se tomaron trozos pequeños de tallo o de raíz, se hirvieron con gotas de detergente de uso comercial y se colocaron en alcohol 70º. Luego se efectuaron cortes histológicos longitudinales (tangenciales y radiales) y transversales con micrótomo de deslizamiento. Se seleccionaron los mejores cortes bajo lupa binocular, se vaciaron en hipoclorito de sodio (lavandina) y se colorearon con safranina "fast-green". Por último, se montaron en Bálsamo de Canadá (D'Ambrogio de Argüeso, 1986; Rodríguez, 1996-1998). Estos cortes histológicos fueron observados y analizados con microscopio óptico. Para las especies herbáceas se siguieron los mismos pasos, pero los cortes se realizaron a mano alzada.
Para las observaciones con microscopía electrónica de barrido (MEB), se separaron pequeños fragmentos de material y se sumergieron en alcohol absoluto (100%); luego se colocaron en nitrógeno líquido hasta alcanzar el punto de congelación. Luego de unos minutos, se realizaron cortes gruesos, dentro de la cubeta con nitrógeno, utilizando hojas de afeitar y un martillo para definir el plano de fractura. Para muestras de carbón arqueológico, el procedimiento consistió en cortar pequeños trozos en distintas direcciones (transversal y longitudinal) bajo lupa binocular. Los cortes o fragmentos obtenidos, fueron montados en platinas impregnadas con carbono y metalizados con oro-paladio (60-40).
RESULTADOS
En la Tabla 2 se detalla el listado de las especies del área en estudio, para cada taxón se indica la familia a la cual pertenece, el nombre vernáculo, el uso actual, las transectas correspondientes y el/los ejemplares de referencia. Las especies que fueron halladas tanto en las transectas como en los sitios arqueológicos, se indican con asterisco en la Tabla 2. Para la clasificación y los nombres científicos se siguió el Catálogo de las Plantas Vasculares del Cono Sur (Zuloaga et al., 2008).
A continuación se presentan las descripciones anatómicas de las especies ordenadas bajo las dos categorías: 1) especies leñosas: combustibles y/o confección de tecnofacturas, y 2) especies herbáceas: pisos de ocupación, confección de artefactos y algunas comestibles como la quínoa. Dentro de estas categorías, las especies se ordenaron por familia. Para las especies leñosas se describe la anatomía del tallo, y también raíz en el caso de Adesmia horrida; para las especies herbáceas se describe la lámina o el tallo. Las descripciones se realizaron sobre la base de los restos arqueológicos y del material actual de referencia.
1) Especies leñosas: combustibles y/o confección de tecnofacturas
Las especies leñosas recuperadas en los sitios arqueológicos del área crecen actualmente en el pajonal y en el tolar (Fig. 2), por lo tanto, estas asociaciones vegetales fueron las probables fuentes de aprovisionamiento de combustible. Estas especies pertenecen a las familias Asteraceae, Brassicaceae, Chenopodiaceae, Ephedraceae, Fabaceae, Solanaceae y Verbenaceae (Tabla 2), siendo la familia Asteraceae la que presenta más variedad de taxones y la familia Fabaceae la mejor representada en relación con la cantidad de material recuperado perteneciente a Adesmia horrida.
Familia Asteraceae
Baccharis salicifolia
Corte transversal de tallo (Figs. 4 C-D). Anillos de crecimiento marcados. Porosidad sub-circular. Vasos de contorno anguloso, paredes delgadas y diámetro pequeño en series radiales cortas y largas, en series tangenciales y en racimos; también en disposición dendrítica. Fibras abundantes de paredes poco engrosadas. Parénquima axial paratraqueal en bandas confluentes.

Fig. 4. Material actual, fotomicrografías tomadas con microscopio óptico. A-B, Baccharis tola. A, tallo en CT. B, tallo en CLTg. C-E, B. salicifolia. C-D, tallo en CT. E, tallo en CLTg. Figura en color en la versión online http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/529/569.
Corte longitudinal tangencial (Fig. 4 E) y radial de tallo. Elementos vasculares relativamente cortos comunicados entre sí mediante placas de perforación simples y oblicuas. Sistema radial heterogéneo y radios heterocelulares.
Baccharis tola
Corte transversal de tallo (Fig. 4 A). Anillos de crecimiento no marcados. Porosidad sub-circular. Vasos de contorno anguloso, de paredes delgadas y diámetro pequeño, principalmente en disposición dendrítica; también en series radiales cortas y largas, en series tangenciales y en racimos. Fibras muy abundantes de paredes poco engrosadas. Parénquima axial abundante paratraqueal dispuesto en bandas confluentes, acompañando a los vasos en su recorrido dendrítico.
Corte longitudinal tangencial (Fig. 4 B) y radial de tallo. Elementos vasculares relativamente cortos; se comunican entre sí mediante placas de perforación simples y oblicuas. Sistema radial heterogéneo y radios heterocelulares con células verticales, cúbicas y procumbentes, uniseriados y pluriseriados con 3-4 células.
Parastrephia lucida
Corte transversal de tallo (Fig. 5 A). Anillos de crecimiento no marcados. Porosidad difusa. Vasos de contorno circular y diámetro reducido solitarios y en racimos, dispuestos entre las fibras. Fibras relativamente abundantes de paredes engrosadas. Parénquima axial paratraqueal escaso.
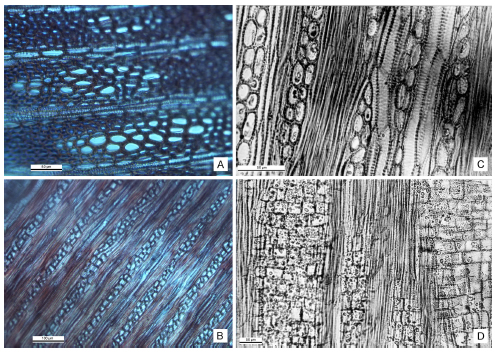
Fig. 5. Material actual, fotomicrografías tomadas con microscopio óptico. A-B, Parastrephia lucida. A, tallo en CT. B, tallo en CLTg. C-D, P. quadrangularis. C, tallo en CLTg. D, tallo en CLRd. Figura en color en la versión online http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/529/569.
Corte longitudinal tangencial (Fig. 5 B) y radial de tallo. Elementos vasculares cortos; se comunican entre sí mediante placas de perforación simples y oblicuas, y por puntuaciones areoladas alternas; fibras con puntuaciones rebordeadas. La mayor parte de los radios son pluriseriados, otros son uniseriados y algunos presentan porciones uniseriadas y biseriadas determinando un sistema radial heterogéneo. Los radios son heterocelulares con células cúbicas y procumbentes.
Parastrephia quadrangularis
Corte transversal de tallo. Anillos de crecimiento no marcados. Porosidad difusa. Vasos de contorno circular y diámetro reducido, solitarios; en racimos, en series radiales cortas y largas. Fibras con tendencia dendrítica, abundantes y de paredes engrosadas. Parénquima axial paratraqueal y escaso.
Corte longitudinal tangencial (Fig. 5 C) y radial (Fig. 5 D) de tallo. Elementos vasculares en general cortos y se comunican entre sí mediante placas de perforación simples algo inclinadas y lateralmente por puntuaciones areoladas alternas. Fibras con puntuaciones escasas. Sistema radial heterogéneo; radios muy altos y heterocelulares; multiseriados con porciones uniseriadas; células verticales y procumbentes.
Senecio santelicis
Corte transversal de tallo (Fig. 6 A). Anillos de crecimiento marcados. Porosidad con tendencia a sub-circular. Vasos de contorno circular y paredes gruesas; solitarios, en racimos y en series cortas y largas y algunas con tendencia dendrítica. Fibras abundantes de paredes engrosadas. Fibrotraqueidas de otoño que delimitan los anillos de crecimiento. Parénquima axial paratraqueal bandeado que acompaña a los vasos en su recorrido.

Fig. 6. Material actual, fotomicrografías tomadas con microscopio óptico. A-B, Senecio santelicis. A, tallo en CT. B, tallo en CLTg. C-D, Neuontobotrys tarapacana. C, tallo en CT. D, tallo en CLTg. Figura en color en la versión online http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/529/569.
Corte longitudinal tangencial (Fig. 6 B) y radial de tallo. Elementos vasculares medianos y algunos de ellos presentan apéndices. Se comunican mediante placas de perforación simples, algo inclinadas y por puntuaciones areoladas alternas. Fibras medianas con paredes gruesas y puntuaciones simples. Radios escasos y homocelulares con células verticales; los radios son uniseriados o parcialmente biseriados, por lo tanto el sistema radial es heterogéneo. También hay radios unicelulares.
Familia Brassicaceae
Neuontobotrys tarapacana
Corte transversal de tallo (Fig. 6 C). Anillos de crecimiento poco marcados. Porosidad difusa con tendencia a subcircular. Vasos de contorno circular; solitarios, en racimos y en series cortas y largas. Fibras abundantes de paredes gruesas. Parénquima axial paratraqueal escaso.
Corte longitudinal tangencial (Fig. 6 D) y radial de tallo. Elementos vasculares relativamente medianos y de diámetro reducido; se comunican mediante puntuaciones areoladas alternas y simples y por placas de perforación oblicuas. Radios muy altos y heterocelulares con células verticales y cúbicas; sistema radial homogéneo.
Familia Chenopodiaceae
Atriplex imbricata
Corte transversal (Fig. 7 C) de tallo. Anillos de crecimiento ausentes. Crecimiento secundario anómalo; células del xilema dispuestas hacia adentro y hacia afuera. Fibras muy abundantes, de paredes gruesas.
Corte longitudinal tangencial (Fig. 7 D-E) y radial de tallo. Elementos vasculares medianos comunicados entre sí mediante placas de perforación, oblicuas y simples. Puntuaciones simples, areoladas y alternadas. Sistema radial homogéneo.
Familia Ephedraceae
Ephedra multiflora
Corte transversal de tallo (Fig. 7 F-G). Anillos de crecimiento marcados. Porosidad difusa. Vasos solitarios de contorno anguloso. Fibrotraqueidas de paredes gruesas. Parénquima axial paratraqueal escaso.

Fig. 7. Material actual. A-B, Acantholippia salsoloides, fotomicrografías tomadas con microscopio óptico. A, tallo en CT. B, tallo en CLTg. C-E, Atriplex imbricata, fotomicrografías tomadas con MEB. C, tallo en CT. D-E, tallo en CLTg (tomadas de Aguirre, 2012). F-G. Ephedra multiflora, fotomicrografías tomadas con microscopio óptico de tallo en CT. H-J, Neosparton ephedroides, fotomicrografías tomadas con MEB. H, tallo en CT. I, tallo en CLTg. J, tallo en corte longitudinal radial (CLRd) (tomadas de Aguirre, 2012).
Corte longitudinal tangencial y radial de tallo. Los elementos vasculares se comunican entre sí mediante puntuaciones areoladas y placas de perforación oblicuas. Sistema radial heterogéneo; radios heterocelulares.
Familia Fabaceae
Adesmia horrida
Corte transversal de tallo (Fig. 8 A). Anillos de crecimiento marcados. Porosidad sub-circular. Vasos de contorno circular y paredes gruesas en series tangenciales y en disposición dendrítica, también en racimos, series radiales cortas y largas y unos pocos vasos solitarios. Fibras abundantes con paredes engrosadas. Parénquima axial paratraqueal bandeado confluente, parénquima radial cristalífero.

Fig. 8. Material actual, fotomicrografías tomadas con microscopio óptico de Adesmia horrida. A, tallo en CT. B, raíz en CT. C-D, tallo en CLTg. E, tallo en CLRd.
Corte transversal de raíz (Fig 8. B). Anillos de crecimiento marcados. Porosidad sub-circular. Vasos de contorno circular y paredes gruesas, dispuestos en series tangenciales muy marcadas que alternan con paquetes de fibras; pocos vasos solitarios. Fibras muy abundantes con paredes engrosadas. Parénquima axial paratraqueal bandeado confluente, parénquima radial cristalífero.
Corte longitudinal tangencial de tallo (Fig 8 C) y de raíz (Fig. 8 D). Elementos vasculares relativamente cortos; se comunican entre sí por puntuaciones areoladas alternas y por placas de perforación, simples y oblicuas. Sistema radial heterogéneo y radios heterocelulares. Pocos radios uniseriados y numerosos pluriseriados con 2-3 células de ancho en el tallo y 3-10 en la raíz.
Corte longitudinal radial de raíz (Fig. 8 E). Células radiales parenquimáticas rectangulares, cúbicas y procumbentes.
Familia Solanaceae
Fabiana bryoides
Corte transversal de tallo (Fig. 9 A). Anillos de crecimiento marcados. Porosidad sub-circular. Vasos de contorno anguloso y paredes delgadas; principalmente solitarios, en series radiales cortas, largas y en series tangenciales. Parénquima axial escaso, paratraqueal y apotraqueal difuso.

Fig. 9. Material actual, fotomicrografías tomadas con microscopio óptico. A-B, Fabiana bryoides. A, tallo en CT. B, tallo en CLTg. C-E, F. punensis. C-D, tallo en CT. E, tallo en CLTg. Figura en color en la versión online http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/529/569.
Corte longitudinal tangencial (Fig. 9 B) y radial de tallo. Elementos vasculares de tamaño variado; se comunican mediante placas de perforación simples y más o menos oblicuas. Fibrotraqueidas de paredes delgadas y puntuaciones areoladas. Radios heterocelulares y sistema radial heterogéneo.
Fabiana punensis
Corte transversal de tallo (Fig. 9 C-D). Anillos de crecimiento marcados. Porosidad sub-circular. Vasos de contorno anguloso y paredes delgadas; solitarios, en series radiales cortas, largas y en series tangenciales. Fibras con puntuaciones simples. Parénquima axial escaso, apotraqueal difuso y en algunos casos paratraqueal bandeado.
Corte longitudinal tangencial (Fig. 9 E) y radial de tallo. Elementos vasculares de tamaño variado; se comunican entre sí mediante placas de perforación simples y oblicuas; presentan S3 en hélice. Radios heterocelulares con células cúbicas y procumbentes y sistema radial homogéneo (radios uniseriados).
Familia Verbenaceae
Acantholippia deserticola
Corte transversal de tallo (Fig. 10 A-B). Anillos de crecimiento marcados. Porosidad sub-circular; vasos de contorno circular y paredes gruesas en series cortas (2-3 elementos de ancho) radiales y tangenciales; también existen algunas series largas (6 elementos), racimos y vasos solitarios. Fibras de paredes gruesas y frecuentemente aparecen rodeando a los vasos. Parénquima axial paratraqueal bandeado.

Fig. 10. Material actual, fotomicrografías tomadas con microscopio óptico de Acantholippia deserticola. A-B, tallo en corte transversal (CT). C-D, tallo en corte longitudinal tangencial (CLTg).
Corte longitudinal tangencial (Fig. 10 C-D) y radial de tallo. Elementos vasculares relativamente largos, se comunican entre sí mediante puntuaciones areoladas y placas de perforación simples y oblicuas. Sistema radial heterogéneo y radios heterocelulares, uniseriados y pluriseriados (2-4 células). Espesamientos helicoidales en las paredes de los elementos vasales.
Acantholippia salsoloides
Corte transversal de tallo (Fig. 7 A). Anillos de crecimiento marcados. Porosidad sub-circular; vasos de contorno circular y paredes más o menos gruesas, dispuestos en series cortas (2 - 3 elementos de ancho) radiales y tangenciales; también existen algunas series largas (5 elementos), racimos y algunos vasos solitarios. Fibras de paredes gruesas. Parénquima axial paratraqueal bandeado.
Corte longitudinal tangencial (Fig.7 B) y radial de tallo. Elementos vasculares relativamente largos y medianos, se comunican entre sí mediante puntuaciones areoladas y placas de perforación simples y oblicuas. Sistema radial heterogéneo y radios heterocelulares.
Neosparton ephedroides
Corte transversal de tallo (Fig. 7 H). Anillos de crecimiento marcados. Porosidad sub-circular. Vasos de contorno anguloso dispuestos en series cortas o largas. Fibras de paredes delgadas. Parénquima apotraqueal difuso y paratraqueal bandeado.
Corte longitudinal tangencial (Fig. 7 I) y radial (Fig. 7 J) de tallo. Los elementos vasculares se comunican entre sí mediante placas de perforación simples y oblicuas. Radios heterocelulares constituidos por células cúbicas y procumbentes.
2) Especies herbáceas: pisos de ocupación, confección de artefactos y comestibles
Las especies herbáceas recuperadas en los sitios arqueológicos del área pertenecen a las familias Chenopodiaceae, Cyperaceae, Juncaceae, Poaceae y Typhaceae (Tabla 2). La familia Poaceae es la mejor representada en cuanto a cantidad de taxones y variedad de usos. El género Deyeuxia predomina en las camadas que constituyeron los pisos de ocupación en los sitios arqueológicos y al mismo tiempo es el más utilizado en la confección de tecnofacturas (Fig. 13).

Fig. 13. Fotomicrografías tomadas con microscopio óptico. A, material arqueológico. B-G, material actual. A-B, Deyeuxia eminens var. eminens. A, hoja a nivel de la lámina en CT, recuperado en el sitio PP4. B, caña florífera en CT. C-E, D. eminens var. fulva. C, hoja a nivel de la lámina en CT. D, detalle de C. E, caña florífera en CT. F-G, D. rigescens. F, hoja a nivel de la lámina en CT. G, caña florífera en CT. Figura en color en la versión online http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/529/569.
Familia Chenopodiaceae
Chenopodium quinoa
Corte transversal de una rama florífera (Fig. 11 A). Contorno sub-circular. Epidermis seguida por varias capas de células suberificadas. Parénquima incoloro con cristales tipo drusas; tejido esclerenquimático dispuesto en varias capas contínuas. Haces vasculares abiertos subtriangulares colaterales Médula parenquimática de gran desarrollo y parénquima interfascicular con drusas (Rodríguez, 2008).

Fig. 11. Fotomicrografías tomadas con microscopio óptico. A-C, material actual. D, material arqueológico. A, Chenopodium quinoa, tallo en CT. B, Scirpus asper, tallo en CT. C, Juncus balticus sulsp. mexicanus, tallo en CT. D, Typha domingensis, tallo en CT, recuperado en el sitio PP4. Abreviaturas: ae, aerénquima; ec, esclerénquima; hv, haz vascular; pa, parénquima.
Familia Cyperaceae
Scirpus asper
Corte transversal del tallo (Fig. 11 B). Contorno cuadrangular. Epidermis simple seguida de una hipodermis parenquimática; esclerénquima con células de paredes gruesas rodeando los haces vasculares, constituidos por dos vasos de metaxilema y uno de protoxilema poco desarrollados y floema más abundante; haces rodeados también por una capa de células parenquimáticas. Parénquima medular abundante (Rodríguez et al., 2006).
Familia Juncaceae
Juncus balticus subsp. mexicanus
Corte transversal del tallo (Fig. 11 C). Contorno circular. Epidermis formada por una sola capa de células, seguida por varias capas de esclerénquima. Haces vasculares en dos o tres hileras con protoxilema, metaxilema y floema notorios; haces en formación incluidos en esclerénquima. Parénquima cortical escaso. Espacios aeríferos entre las hileras de haces. Médula hueca (Rodríguez, 2008).
Familia Poaceae
Cortaderia speciosa
Corte transversal de la hoja a nivel de la lámina (Fig. 12 A-B). Contorno en forma de "U" muy abierta a expandido. Haces vasculares de contorno circular, con abundantes fibras floemáticas, vasos del metaxilema de gran diámetro; laguna protoxilemática en los haces de mayor tamaño (Rodríguez, 1999), rodeados por vainas parenquimáticas que se extienden hacia ambas epidermis (haces trabados). Engrosamientos en el ala de la hoja con la gran cantidad de fibras que rodean casi por completo a los haces vasculares hacia la epidermis adaxial. Surcos bien marcados, rodeados por clorénquima limitado por fibras subepidérmicas adaxiales. Abundantes aguijones en epidermis adaxial.
Deyeuxia curvula
Corte transversal de la hoja a nivel de la lámina (Fig. 12 C-D). Contorno redondeado. Haces vasculares en general en número de siete, rodeados por vaina mestomática notable y parenquimática inconspicua. Esclerénquima discontinuo formando paquetes subepidérmicos en la cara adaxial de las costillas; esclerénquima subepidérmico abaxial formado por una o dos capas de fibras y marginal formado por dos o tres capas de fibras. Mesofilo con clorénquima homogéneo. Costillas y surcos en cara adaxial. Células epidérmicas adaxiales con pared tangencial externa engrosada, células buliformes de escaso desarrollo, ausentes en la cara abaxial. Pelos largos rígidos y cortos en la porción distal de las costillas. Estomas en las caras laterales de los surcos. Células epidérmicas abaxiales con la pared tangencial externa notablemente engrosada (Rodríguez et al., 2003).

Fig. 12. Material actual, fotomicrografías tomadas con microscopio óptico. A-B, Cortaderia speciosa, hoja a nivel de la lámina en CT. C-E, Deyeuxia curvula. C-D, hoja a nivel de la lámina en CT. E, caña florífera en CT. F-H, Deyeuxia deserticola. F, hoja a nivel de la lámina en CT. G, detalle de F a nivel del margen foliar. H, caña florífera en CT. Figura en color en la versión online http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/529/569.
Corte transversal de la caña florífera (Fig. 12 E). Caña hueca de contorno elíptico. Epidermis uniestratificada, con la pared tangencial externa más engrosada. Haces vasculares dispuestos en dos series. Escleréquima subepidérmico inconspicuo formado por una o dos capas de fibras. Parénquima cortical muy abundante formado por células que aumentan gradualmente de tamaño hacia la parte medular (Rodríguez et al., 2003).
Deyeuxia deserticola
Corte transversal de la hoja a nivel de la lámina (Fig. 12 F-G). Contorno redondeado. Haces vasculares primarios en general en número de seis, rodeados por vaina mestomática notable y parenquimática inconspicua. Esclerénquima subepidérmico abaxial continuo formado por una o dos capas de fibras. Mesofilo con clorénquima homogéneo. Costillas y surcos presentes en la cara adaxial, costillas triangulares semejantes; costillas marginales con las caras internas más o menos paralelas entre sí. Células epidérmicas adaxiales con pared tangencial externa engrosada, células buliformes inconspicuas dispuestas en grupos regulares en los surcos y ausentes en la cara abaxial. Pelos largos rígidos y cortos en la porción distal de las costillas. Estomas en caras laterales de los surcos. Células epidérmicas abaxiales con pared tangencial externa engrosada (Rodríguez et al., 2003).
Corte transversal de la caña florífera (Fig. 12 H). Caña hueca de contorno elíptico. Epidermis uniestratificada, con la pared tangencial externa más engrosada. Haces vasculares dispuestos en dos series, rodeados por una vaina parenquimática de paredes engrosadas. Escleréquima conspicuo formado por cuatro capas de fibras. Parénquima cortical abundante formado por células que aumentan gradualmente de tamaño hacia la parte central (Rodríguez et al., 2003).
Deyeuxia eminens var. eminens
Corte transversal de la hoja a nivel de la lámina (Fig. 13 A). Contorno en forma de "U". Haces vasculares generalmente en número de nueve, rodeados por vaina mestomática notable y parenquimática inconspicua. Esclerénquima discontinuo en la cara adaxial de las costillas; esclerénquima abaxial subepidérmico formado por tres o cuatro capas de fibras en vinculación con los haces primarios; esclerénquima marginal formado por dos o tres capas de fibras. Mesofilo con clorénquima homogéneo. Costillas y surcos profundos en la cara adaxial. Células epidérmicas adaxiales con pared externa tangencial engrosada alternando con células papilosas; células buliformes conspicuas dispuestas en grupos regulares en los surcos y ausentes en la cara abaxial. Pelos largos rígidos y cortos dispuestos en la porción distal de las costillas. Papilas notablemente desarrolladas. Estomas presentes en las caras laterales de los surcos. Células epidérmicas abaxiales con la pared tangencial externa engrosada; aguijones presentes (Rodríguez et al., 2003).
Corte transversal de la caña florífera (Fig. 13 B). Caña hueca de contorno elíptico. Epidermis uniestratificada, con la pared tangencial externa más engrosada. Haces vasculares dispuestos en dos series, rodeados por una vaina parenquimática. Esclerénquima subepidérmico conspicuo formado por cinco o seis capas de fibras. Parénquima cortical abundante formado por células que aumentan gradualmente de tamaño hacia la parte central (Rodríguez et al., 2003).
Deyeuxia eminens var. fulva
Corte transversal de la hoja a nivel de la lámina (Fig. 13 C-D). Contorno redondeado. Haces vasculares libres generalmente en número de siete, rodeados por vaina mestomática notable y parenquimática inconspicua. Esclerénquima discontinuo formando paquetes subepidérmicos en la cara adaxial; esclerénquima abaxial subepidérmico formado por tres o cuatro capas de fibras. Mesofilo con clorénquima homogéneo. Costillas y surcos profundos en la cara adaxial, costilla media triangular y las laterales redondeadas y las marginales con las caras internas más o menos paralelas entre sí. Células epidérmicas adaxiales con pared tangencial externa engrosada; células papilosas presentes, células buliformes conspicuas en la cara adaxial. Pelos rígidos y papilas prominentes. Estomas presentes en las caras laterales de los surcos. Células epidérmicas abaxiales con pared tangencial externa engrosada alternando con aguijones (Rodríguez et al., 2003).
Corte transversal de la caña florífera (Fig. 13 E). Caña hueca de contorno subcircular con costillas redondeadas incipientes. Epidermis uniestratificada, con pared tangencial externa más engrosada. Haces vasculares dispuestos en dos series, rodeados por una vaina parenquimática de paredes engrosadas. Esclerénquima abaxial conspicuo formado por cinco o seis capas de fibras. Clorénquima subepidérmico representado por dos grupos de células. Parénquima fundamental escaso (Rodríguez et al., 2003).
Deyeuxia rigescens
Corte transversal de la hoja a nivel de la lámina (Fig. 13 F). Contorno en forma de "V". Haces vasculares trabados, generalmente en número de 13-15, rodeados por vaina mestomática notable y parenquimática inconspicua, poco desarrollada, sin cloroplastos especializados. Esclerénquima discontinuo en vinculación con los haces vasculares; esclerénquima marginal formado por dos o tres capas de fibras. Mesofilo con clorénquima difuso. Costillas y surcos profundos en la cara adaxial. Células epidérmicas adaxiales con paredes delgadas alternando con células papilosas redondeadas y aguijones pequeños; células buliformes conspicuas dispuestas en grupos regulares en los surcos y ausentes en la cara abaxial. Estomas presentes en las caras laterales de los surcos. Células epidérmicas abaxiales con pared tangencial externa engrosada (Rodríguez et al., 2003).
Corte transversal de la caña florífera (Fig. 13 G). Caña hueca de contorno redondeado. Epidermis uniestratificada, con la pared tangencial externa más engrosada. Haces vasculares dispuestos en tres o cuatro series alternas, rodeados por una vaina parenquimática de paredes engrosadas. Escleréquima subepidérmico conspicuo formado por ocho a diez capas de fibras. Parénquima fundamental abundante formado por células que aumentan gradualmente de tamaño hacia la parte central (Rodríguez et al., 2003).
Festuca chrysophylla
Corte transversal de hoja a nivel de la lámina. (Fig. 14 A). Contorno en forma de "U". Haces vasculares generalmente en número de doce, tres primarios trabados por fibras hacia la cara abaxial y parénquima incoloro hacia la cara adaxial, dos secundarios y el resto terciarios, libres, más próximos a la cara abaxial, todos rodeados por vaina parenquimática. Esclerénquima difuso formando pequeños paquetes subepidérmicos dispersos en la cara adaxial de las costillas y en la cara abaxial, en donde se ubica a la altura de los haces vasculares; esclerénquima marginal formado por dos o tres capas de fibras. Mesofilo homogéneo. Costillas y surcos profundos en la cara adaxial. Costillas truncadas, grandes y pequeñas alternas; costillas marginales con las caras internas no paralelas entre sí. Células epidérmicas adaxiales con pared tangencial externa delgada; células buliformes conspicuas dispuestas en los surcos. Pelos y aguijones en la cara adaxial. Células epidérmicas abaxiales con la pared tangencial externa poco engrosada.

Fig. 14. Material actual, fotomicrografías tomadas con microscopio óptico de CT de hoja a nivel de la lámina. A, Festuca chrysophylla. B, F. ortophylla. C, F. dissitiflora. D, F. weberbaueri. E, Puccinellia frigida. F, Pappostipa frigida. G, P. vaginata. Abreviaturas: co, costilla; su, surco. Figura en color en la versión online http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/529/569.
Festuca dissitiflora
Corte transversal de hoja a nivel de la lámina (Fig. 14 C). Contorno en forma de "V". Haces vasculares generalmente en número de quince, tres primarios trabados por fibras hacia la cara abaxial y parénquima incoloro hacia la cara adaxial, cuatro secundarios semi-trabados por fibras a la cara abaxial y el resto libres, más próximos a la cara abaxial, todos rodeados por vaina parenquimática desarrollada. Esclerénquima continuo con células más engrosadas en la cara adaxial de las costillas; esclerénquima abaxial subepidérmico formado por una capa continua de fibras; paquetes de fibras a la altura de los haces vasculares, vinculados con los haces primarios y secundarios; esclerénquima marginal formado por dos o tres capas de fibras. Mesofilo homogéneo. Costillas y surcos profundos en la cara adaxial, costillas truncadas, grandes y pequeñas alternas; costillas marginales con las caras internas más o menos paralelas entre sí. Células epidérmicas adaxiales con pared tangencial externa engrosada; células buliformes conspicuas dispuestas en los surcos. Estomas presentes en las caras laterales de los surcos. Pelos y aguijones en la cara adaxial. Células epidérmicas abaxiales con la pared tangencial externa algo engrosada.
Festuca orthophylla
Corte transversal de hoja a nivel de la lámina (Fig. 14 B). Contorno en forma de "U". Haces vasculares generalmente en número de doce, cinco primarios trabados por fibras hacia la cara abaxial y parénquima hacia la cara adaxial, los restantes secundarios, semi-trabados por fibras a la cara abaxial, todos rodeados por vaina parenquimática poco desarrollada hacia la cara abaxial formada por una o dos capas de células hacia la cara adaxial. Esclerénquima formando paquetes subepidérmicos en la cara adaxial de las costillas; esclerénquima abaxial subepidérmico formado por cuatro o cinco capas de fibras, continuo en vinculación con los haces vasculares; esclerénquima marginal formado por tres o cuatro capas de fibras engrosadas. Mesofilo homogéneo. Costillas truncadas, grandes y surcos no muy profundos en la cara adaxial; costillas marginales con las caras internas más o menos paralelas entre sí. Células epidérmicas adaxiales con pared tangencial externa engrosada; células buliformes conspicuas dispuestas en los surcos. Estomas presentes en las caras laterales de los surcos. Pelos muy abundantes y aguijones en la cara adaxial. Células epidérmicas abaxiales con la pared tangencial externa algo engrosada.
Festuca weberbaueri
Corte transversal de hoja a nivel de la lámina (Fig. 14 D). Contorno en forma de "U". Haces vasculares generalmente en número de nueve, cinco primarios trabados por fibras hacia la cara abaxial y parénquima incoloro hacia la cara adaxial, los restantes secundarios, libres, más próximos a la cara abaxial, todos rodeados por vaina parenquimática desarrollada. Esclerénquima formando paquetes subepidérmicos en la cara adaxial de las costillas; esclerénquima abaxial subepidérmico formado por una o dos capas de fibras, continuo en vinculación con los haces primarios; esclerénquima marginal formado por dos o tres capas de fibras. Mesofilo homogéneo. Costillas y surcos profundos en la cara adaxial, costillas truncadas y grandes; costillas marginales con las caras internas más o menos paralelas entre sí. Células epidérmicas adaxiales con pared tangencial externa algo engrosada; células buliformes conspicuas dispuestas en los surcos. Estomas presentes en las caras laterales de los surcos. Pelos y aguijones en la cara adaxial. Células epidérmicas abaxiales con la pared tangencial externa engrosada.
Pappostipa frigida
Corte transversal de hoja a nivel de la lámina (Fig. 14 F). Contorno en forma de "U". Haces vasculares generalmente en número de ocho, tres primarios trabados por fibras hacia la cara abaxial y parénquima incoloro hacia la cara adaxial, los restantes secundarios, libres. Esclerénquima formando pequeños paquetes subepidérmicos en la cara adaxial de las costillas; esclerénquima abaxial subepidérmico formado por tres o cuatro capas de fibras, continuo, formando paquetes en vinculación con los haces primarios; esclerénquima marginal formado por tres o cuatro capas de fibras de paredes engrosadas. Mesofilo homogéneo. Costillas y surcos poco marcados en la cara adaxial, costillas truncadas y grandes; costillas marginales con las caras internas más o menos paralelas entre sí. Células epidérmicas adaxiales con pared tangencial externa engrosada; células buliformes conspicuas dispuestas en los surcos y ausentes en la cara abaxial. Pelos y aguijones en la cara adaxial. Células epidérmicas abaxiales con la pared tangencial externa engrosada.
Pappostipa vaginata
Corte transversal de hoja a nivel de la lámina (Fig. 14 G). Contorno en forma de "U". Haces vasculares generalmente en número de doce, rodeados por vaina parenquimática desarrollada. Esclerénquima continuo formando paquetes subepidérmicos en la cara adaxial de las costillas; esclerénquima abaxial subepidérmico formado por tres o cuatro capas de fibras; esclerénquima marginal formado por dos o tres capas de fibras. Mesofilo homogéneo. Costillas y surcos profundos en la cara adaxial, costillas truncadas, grandes y pequeñas alternas; costillas marginales con las caras internas más o menos paralelas entre sí. Células epidérmicas adaxiales con pared tangencial externa engrosada; células buliformes conspicuas dispuestas en los surcos y ausentes en la cara abaxial. Estomas presentes en las caras laterales de los surcos. Pelos y aguijones en cara adaxial. Células epidérmicas abaxiales con la pared tangencial externa algo engrosada (Rodríguez et al. 2006).
Puccinellia frigida
Corte transversal de hoja a nivel de la lámina (Fig. 14 E). Contorno irregular, ligeramente en forma de "U". Haces vasculares generalmente en número de diez, seis primarios y cuatro secundarios, todos semi-trabados por fibras hacia la cara abaxial. Esclerénquima formando paquetes subepidérmicos en la cara adaxial de las costillas; esclerénquima abaxial subepidérmico muy marcado y continuo, en vinculación con los haces vasculares; esclerénquima marginal formado por dos o tres capas de fibras. Mesofilo homogéneo. Costillas y surcos más o menos profundos hacia ambas caras, costillas redondeadas, más grandes en la cara adaxial; costillas marginales irregulares. Células epidérmicas adaxiales con pared tangencial externa engrosada; células buliformes dispuestas en los surcos. Aguijones escasos en la cara adaxial. Células epidérmicas abaxiales con la pared tangencial externa engrosada.
Familia Typhaceae
Typha domingensis
Corte transversal del tallo (Fig. 11 D). Contorno redondeado. Epidermis uniestratificada seguida por un esclerénquima muy desarrollado. Haces vasculares abundantes incluidos en el parénquma fundamental, conformados por protoxilema y metaxilema bien desarrollados, floema orientado hacia la periferia y rodeados por esclerénquima (Rodríguez, 2008).
DISCUSIÓN
Los resultados obtenidos en los trabajos que aquí se sintetizan, permiten contrastar el ambiente y el paleoambiente del área de Antofagasta de la Sierra a través del estudio de la flora actual, las entrevistas a antiguos pobladores de la zona y el análisis del registro arqueobotánico.
Todas las especies locales que se recuperaron en los sitios arqueológicos crecen actualmente en el área; sin embargo, debido a aspectos vinculados con la preservación y a problemas tafonómicos, no todas las especies actuales fueron halladas en dichos sitios hasta el momento. Tal es el caso de los taxones comestibles de los que rara vez se encuentran restos arqueológicos; por lo cual es necesario recurrir a los análisis isotópicos sobre huesos o al estudio de coprolitos humanos para estudiar la dieta y asegurar o descartar su consumo. No obstante, es importante destacar la presencia de especies domésticas comestibles tales como Chenopodium quinoa en el registro arqueobotánico del área. Se recuperaron tallos en los sitios Punta de la Peña 4 (PP4) y Peñas Chicas 1.3 (PCH1.3) y una considerable cantidad de semillas en el primero de ellos, las cuales fueron fechadas utilizando el método AMS ("Acelerator Mass Espectrometry") arrojando antigüedades de 760±40, UGA-15089 y 690±50, UGA-15090 (Rodríguez et al., 2006; Aguirre, 2007, 2012). En la actualidad, esta especie es cultivada en el área y se cuenta con una valiosa información etnobotánica al respecto (Aguirre, 2012). Asimismo, se hallaron microrrestos pertenecientes a este taxón, tales como granos de almidón de semillas en artefactos líticos en los sitios QS3 y PCH1.1 y fitolitos de calcio y granos de almidón de semillas en el sitio PP9 (Babot, 2004).
Un dato significativo es que en el registro arqueobotánico de los sitios estudiados se recuperó una elevada cantidad de especies no locales, que crecen en zonas situadas hasta ca. 600 km de distancia, tales como Acrocomia aculeata (Jacq.) Lodd. ex Mart. (= Acrocomia chunta Covas & Ragonese), Chusquea lorentziana Griseb., Prosopis alba Griseb., P. torquata (Cav. ex Lag.) DC, Rhipidocladum neumanii Suleckic, Rúgolo & L. G. Clark y Salix humboldtiana Willd. Estas especies habrían sido utilizadas para confeccionar tecnofacturas, como cordeles, nudos y astiles para puntas de proyectil, entre otras (véase Rodríguez, 1998, 1999, 2005, 2008; Rodríguez & Aschero, 2005).
Es interesante analizar y discutir el uso actual y aquel que pudieron haber tenido en el pasado las especies vegetales y que se desprende del contexto en el que fueron recuperadas en los sitios arqueológicos. Por un lado, el uso de las especies vegetales como combustible en el pasado no ofrece dudas, es decir que éste se puede afirmar debido a que las mismas son halladas formando parte de estructuras de combustión, tales como fogones y/o lentes carbonosas de menor tamaño. Varias especies son utilizadas en la actualidad como combustible en el área según la información brindada por los pobladores entrevistados (Rodríguez, 1998), las cuales también fueron halladas en casi todos los sitios arqueológicos estudiados: Acantholippia deserticola, Adesmia horrida, Baccharis tola, Ephedra nultiflora, Fabiana bryoides, F. punensis, Neosparton ephedroides, Parastrephia lucida, P. quadrangularis y Senecio santelicis. El taxón más frecuente en los fogones de los sitios es A. horrida, esto coincide con su abundancia en el área y con el uso actual por los pobladores de Antofagasta de la Sierra.
Algunas de las especies mencionadas en el párrafo anterior tienen en la actualidad otros usos que no podemos confirmar en el pasado. Esto ocurre por ejemplo con Baccharis tola, utilizada también en nuestros días con fines medicinales; Adesmia horrida, como forraje; Fabiana bryoides, como infusión, sahumador y para delimitar corrales; F. punensis también forrajera y medicinal y Acantholippia salsoloides, actualmente medicinal (Tabla 2).
Es importante señalar que la única especie utilizada en el presente como combustible que no fue recuperada en los sitios del área hasta el momento es Adesmia echinus C. Presl, (Añawa negra). Sin embargo, hay otros taxones hallados en las estructuras de combustión arqueológicas que no fueron mencionados como combustible por los pobladores del área entrevistados. Tal es el caso de Neuontobotrys tarapacana (Chuchar) y Atriplex imbricata (Cachiyuyo) usadas en el presente como forraje, que pudieron haber servido en el pasado para comenzar la combustión por el reducido diámetro de los tallos, especialmente en el primer caso.
Por otra parte, Senecio puchii (Chachacoma) tiene uso medicinal como digestivo en nuestros días. En el registro arqueobotánico de algunos sitios se encontraron pequeños trozos de tallos cuyo uso no se puede predecir a partir de las evidencias disponibles. Algo semejante ocurre con Hoffmannseggia eremophila (Algarrobita) de la cual se hallaron restos de flores en sitios tales como Cueva Salamanca 1 y Quebrada seca 3 (Rodríguez & Deginani, 1994- 1995). Los entrevistados la describieron como comestible, lo cual no se puede confirmar para tiempos prehistóricos sin realizar otro tipo de análisis.
La familia Poaceae evidencia interesantes cambios a través del tiempo en relación con el uso de las distintas especies que crecen en el área. Si bien en la Tabla 2 se mencionan varios taxones de esta familia a los cuales los entrevistados hicieron especial referencia en el campo (Deyeuxia chrysophylla, D. velutina var. nardifolia, Festuca sp. y Pappostipa sp.), denominan en general pastos o paja a todas las gramíneas y destacan su utilidad para techar las viviendas y como forraje. El hecho es que actualmente no se utilizan para confeccionar tecnofacturas como en el pasado y esto muestra una importante diferencia en perspectiva diacrónica. En varios sitios del área, tales como Quebrada seca 3 y Punta de la Peña 4 (ca. 2500-9000 años AP) se recuperaron cordeles, sogas y nudos elaborados utilizando distintas especies del género Deyeuxia (véase Rodríguez & Rúgolo de Agrasar, 1999; Rodríguez et al., 2003). Se destaca también un fardo funerario hallado en Quebrada Seca 3 (ca. 4500 años AP), constituido con D. eminens (Rodríguez & Rúgolo de Agrasar, 1999). Asimismo, los pisos de ocupación de los sitios arqueológicos del área están conformados por camadas de las especies de gramíneas de los géneros Deyeuxia, Festuca, Pappostipa y Puccinellia. Esta función podría homologarse con la construcción actual de techos ya que se refiere al uso vinculado con el lugar de habitación o vivienda.
Aún más llamativo es el uso en el pasado de Cortaderia speciosa y la falta de información de los pobladores actuales del área. En varios sitios del área de estudio se encontraron fragmentos de cestería que corresponden a esta especie, de los que se destaca una cesta decorada (ca. 2500 años AP) hallada en el sitio Quebrada Seca 3 (Pérez de Micou & Ancibor, 1994; Rodríguez, 1998).
También se registraron cambios vinculados con el uso en la especie Juncus balticus subsp. mexicanus (Unquillo) utilizada en el presente como forraje y en el pasado posiblemente a modo de aguja (Rodríguez, 2008) y para conformar los pisos de ocupación en algunos sitios en los que también se halló Scirpus asper.
Por último, a partir de este análisis que evidencia semejanzas y diferencias en el uso de las especies vegetales en el pasado y en la actualidad, se delimita un proceso en el tiempo que incluye continuidad y cambios. Es interesante advertir entonces aquellos comportamientos que siguen teniendo lugar entre los pobladores del área, tales como la recolección de determinadas especies como combustible y, al mismo tiempo, aquellos que se abandonan, como ocurre con la confección de tecnofacturas y en especial el caso de la cestería.
CONCLUSIONES
A través de las investigaciones que se integran en este trabajo, es posible evaluar los valiosos aportes de la Botánica dentro del campo de la Arqueología, específicamente de la Arqueobotánica. Los estudios anatómicos permiten identificar las especies que se recuperan en los sitios arqueológicos con escasos órganos vegetativos y sin órganos reproductivos. De este modo, la anatomía comparada hace posible conocer los taxones utilizados en el pasado prehistórico y, a partir de esta información, inferir las posibles áreas de explotación de recursos vegetales, así como también los probables usos de los mismos. En el último caso, es necesario considerar la distribución de cada especie en el sitio arqueológico en el que fue hallada y, al mismo tiempo, la información etnobotánica obtenida en el área de estudio o en otras semejantes, en los casos en los que esto sea posible. Por otra parte, las especies que no crecen en el área próxima a los sitios en los que se recuperan, constituye un dato sumamente interesante para proponer circuitos de movilidad e intercambios que pudieron haber llevado a cabo los grupos humanos en el pasado. Es importante aclarar que en esta investigación se consideraron especies no locales a aquellas que crecen más allá de los 30 km de distancia a partir de un sitio o conjunto de sitios. Esta distancia se estimó considerando la elevada movilidad que caracterizó a los grupos cazadores-recolectores que habitaron en el área de estudio.
Finalmente, este trabajo pretende ser una guía útil tanto para estudios arqueobotánicos como botánicos que se lleven a cabo en la Puna meridional o en otras áreas y/o regiones que presenten una vegetación similar.
AGRADECIMIENTOS
Mi agradecimiento a M. Gabriela Aguirre por facilitarme las fotomicrografías de Atriplex y Neosparton y a Mariana Valente del Instituto de Botánica Darwinion por la diagramación de las figuras. Y muy especialmente a los revisores externos y del Comité editorial de Darwiniana, ya que sin duda sus sugerencias y aportes contribuyeron para enriquecerlo.
BIBLIOGRAFÍA
1. Aguirre, M. G. 2007. Arqueobotánica del sitio Peñas Chicas 1.3 (Antofagasta de la Sierra, Catamarca, Argentina), en B. Marconetto, P. Babot & N. Oliszewski (eds.), Paleoetnobotánica del Cono Sur: Estudios de casos y propuestas metodológicas, pp. 179-196. Córdoba: Ferreyra Editor. [ Links ]
2. Aguirre, M. G. 2012. Recursos vegetales: uso, consumo y producción en la Puna meridional argentina (5000-1500 AP). Tesis doctoral. Universidad Nacional de La Plata, Facultad de Ciencias Naturales y Museo. [ Links ]
3. Aschero, C. A. 1999. El arte rupestre del desierto puneño y el noroeste argentino, en J. Berenguer (ed.), Arte Rupestre en los Andes de Capricornio, pp. 97-135, Santiago de Chile: Museo Chileno de Arte Precolombino y Banco de Santiago. [ Links ]
4. Aschero, C. A. 2010. Arqueologías de Puna y Patagonia centro-meridional: comentarios generales y aporte al estudio de los cazadores-recolectores puneños en los proyectos dirigidos desde el IAM (1991-2009), en P. Arenas, C. Aschero & C. Taboada (eds.), Rastros en el camino…Trayectos e Identidades de una Institución. Homenaje a los 80 años del IAM- UNT, pp. 257-293. San Miguel de Tucumán: Instituto de Arqueología y Museo, EDUNT.
5. Aschero, C. L.; L. Manzi & A. Gómez. 1993-1994. Producción lítica y uso del espacio en el nivel 2b4 de Quebrada Seca 3. Relaciones de la Sociedad Argentina de Antropología XIX: 191-214. [ Links ]
6. Babot, M. P. 2004. Tecnología y utilización de artefactos de molienda en el Noroeste Prehispánico. Tesis doctoral. Universidad Nacional de Tucumán. Facultad de Ciencias Naturales e Instituto Miguel Lillo. [ Links ]
7. Baied, C. & J. Wheeler. 1993. Evolution of high Andean Puna ecosystems: Environment, climate and culture change over the last 12000 years in Central Andes. Mountain Research and Development 13: 145-156. [ Links ]
8. Cabrera, A. L. 1976. Regiones fitogeográficas argentinas. Enciclopedia Argentina de Agricultura y Jardinería, vol. 2, parte 1. Buenos Aires: ACME. [ Links ]
9. Cabrera, A. L. & A. Willink. 1973. Biogeografía de América Latina. Serie de Biología. Monografía 13. Washington D. C.: Secretaría General de la Organización de los Estados Americanos. [ Links ]
10. Cuello, A. S. 2006. Guía ilustrada de la flora de Antofagasta de la Sierra - Catamarca (Puna Meridional Argentina). Informe del curso de entrenamiento para la obtención del grado de Licenciatura en Ciencias Biológicas. Universidad Nacional de Tucumán. [ Links ]
11. D'ambrogio de Argüeso, A. 1986. Manual de técnicas en Histología vegetal. Buenos Aires: Editorial Hemisferio Sur. [ Links ]
12. Elkin, D. C. 1987. Análisis de áreas de explotación de recursos en la cuenca media y superior del río Las Pitas (Antofagasta de la Sierra, Catamarca) y su relación con la evidencia arqueológica de cazadores‑recolectores. Tesis de Licenciatura. Universidad de Buenos Aires, Facultad de Filosofía y Letras. [ Links ]
13. Elkin, D. C. 1992. Explotación de recursos en relación al sitio acerámico Quebrada Seca 3, Antofagasta de la Sierra, Puna de Catamarca. Shincal 2:1-14. [ Links ]
14. García Salemi, M. A. 1986. Geomorfología de regiones secas: Antofagasta de la Sierra, Provincia de Catamarca. Centro de Estudios de Regiones Secas 4:5-13. [ Links ]
15. Haber, A. 1992 Pastores y pasturas. Recursos forrajeros en Antofagasta de la Sierra (Catamarca) en relación a la ocupación Formativa. Shincal 2: 15-23. [ Links ]
16. Martínez, J. G.; C. A. Aschero, J. E. Powell & M. F. Rodríguez. 2004. First evidence of extinct megafauna in the Southern Argentinian Puna. Current Research in the Pleistocene 21: 104-107. [ Links ]
17. Martínez, J. G.; C. A. Aschero, J. E. Powell & P. Tchilinguirián. 2007. A gap between extinct Pleistocene megafaunal remains and Holocene burial contexts at archaeological sites in the Southern Argentinian Puna. Current Research in the Pleistocene 24: 60-62. [ Links ]
18. Martínez, J.; M. Mondini, E. Pintar & M. C. Reigadas. 2010. Cazadores-recolectores tempranos de la Puna Meridional Argentina: avances en su estudio en Antofagasta de la Sierra (Pleistoceno final-Holoceno temprano/medio). Actas del XVII Congreso Nacional de Arqueología Argentina, 11-15 de octubre de 2010, Mendoza (Argentina), Tomo 4 (32): 1691-1696. [ Links ]
19. Núñez, L.; I. Cartajena, C. Carrasco, P. de Souza & M. Grosjean. 2006. Emergencia de comunidades pastoralistas formativas en el sureste de la Puna de Atacama. Estudios Atacameños 32: 93-117. [ Links ]
20. Olivera, D. 2006. Recursos bióticos y subsistencia en sociedades agropastoriles de la Puna meridional argentina. Comechingonia 9: 19-56. [ Links ]
21. Olivera, D.; A. S. Vidal & L. G. Grana. 2003. El sitio Cueva Caco 1A: Hallazgos, espacio y proceso de complejidad en la Puna meridional (ca. 3000 años A. P.). Relaciones de la Sociedad Argentina de Antropología XXVIII: 257-270. [ Links ]
22. Pérez de Micou, C. B. & E. Ancibor, 1994. Manufactura cestera en sitios arqueológicos de Antofagasta de la Sierra, Catamarca. República Argentina. Journal de la Société des Américanistes 80: 207-216. [ Links ]
23. Pintar, E. 1996. Prehistoric Holocene Adaptations to the Salt Puna of Northwest Argentina. PH.D. diss., Southern Methodist University. [ Links ]
24. Pintar, E. 2004. Cueva Salamanca 1: ocupaciones altitermales en la puna sur. Relaciones de la Sociedad Argentina de Antropología XXIX: 357-366. [ Links ]
25. Pintar, E. 2008. High altitude deserts: hunters-gatherers from the Salt Puna, North West Argentina. International Journal of South American Archaeology 2:47-55 [ Links ]
26. Rodríguez, M. F. 1996-1998. Propuesta metodológica para el análisis de macrovestigios vegetales. Presentación de un caso: Quebrada Seca 3, nivel 2b (12). Palimpsesto 5: 238- 248. [ Links ]
27. Rodríguez, M. F. 1998. Arqueobotánica de Quebrada Seca 3: Recursos vegetales utilizados por cazadores-recolectores durante el período Arcaico en la Puna Meridional Argentina. Tesis Doctoral. Universidad de Buenos Aires, Facultad de Ciencias Exactas y Naturales. [ Links ]
28. Rodríguez, M. F. 1999. Arqueobotánica de Quebrada Seca 3 (Puna Meridional Argentina): Especies vegetales utilizadas en la confección de artefactos durante el Arcaico. Relaciones de la Sociedad Argentina de Antropología XXIV: 159-184. [ Links ]
29. Rodríguez, M. F. 2000. Woody plant species used during the Archaic period in the Southern Argentine Puna. Archaeobotany of Quebrada Seca 3. Journal of Archaeological Science 27: 341-361. [ Links ]
30. Rodríguez, M. F. 2004a. Cambios en el uso de los recursos vegetales durante el Holoceno en la Puna meridional argentina. Chungara Revista de Antropología Chilena (Volumen especial): 403-413. [ Links ]
31. Rodríguez, M. F. 2004b. Woody plant resources in the Southern Argentine Puna. Punta de la Peña 9 archaeological site. Journal of Archaeological Science 31: 1361-1372. [ Links ]
32. Rodríguez, M. F. 2005. Human evidence during Middle Holocene in the Salty Argentine Puna. Archaebotanical record analyzes. Quaternary International 132: 15-22. [ Links ]
33. Rodríguez, M. F. 2008 Recursos vegetales y tecnofacturas en un sitio arqueológico de la Puna meridional argentina, Área centro - sur andina. Darwiniana 46: 240-257. [ Links ]
34. Rodríguez, M. F. & N. Deginani. 1994-1995. Recursos vegetales utilizados en Quebrada Seca 3 (Puna Meridional Argentina) a comienzos del Arcaico Tardío. Palimpsesto 4: 122-126. [ Links ]
35. Rodríguez, M. F. & Z. E. Rúgolo de Agrasar. 1999. Deyeuxia eminens (Poaceae: Agrostideae) en un sitio arqueológico de la Puna Meridional Argentina (Provincia de Catamarca). Darwiniana 37: 229-242. [ Links ]
36. Rodríguez, M. F.; Z. E. Rúgolo de Agrasar & C. A. Aschero. 2003. El género Deyeuxia (Poaceae, Agrostideae) en sitios arqueológicos de la Puna meridional argentina. Provincia de Catamarca. Chungara Revista de Antropología Chilena 35: 51-72. [ Links ]
37. Rodríguez, M. F. & C. A. Aschero. 2005. Acrocomia chunta (Arecaceae) Raw material for cord making in the Argentinean Puna. Journal of Archaeological Science 32: 1534- 1542. [ Links ]
38. Rodríguez, M. F.; Z. E. Rúgolo de Agrasar & C. A. Aschero. 2006. El uso de las plantas en unidades domésticas del sitio arqueológico Punta de la Peña 4, Puna meridional argentina. Chungara Revista de Antropología Chilena 38: 257-271. [ Links ]
39. Rodríguez, M. F. & C. A. Aschero. 2007. Archaeological evidences of Zea mays L. (Poaceae) in the Southern Argentinean Puna (Antofagasta de la Sierra, Catamarca. Journal of Ethnobiology 27: 256-271. [ Links ]
40. Thiers, B. 2013. [De actualización constante, consulta 2010] Index Herbariorum: a global directory of public herbaria and associated staff. New York Botanical Garden's Virtual Herbarium, http://sweetgum.nybg.org/ih. [ Links ]
41. Troll, C. 1958. Las culturas superiores andinas y el medio geográfico. Revista del Instituto de Geografía (Lima) 5: 3-55. [ Links ]
42. Zuloaga, F. O.; O. Morrone & M. Belgrano. 2008. Catálogo de las Plantas Vasculares del Cono Sur (Argentina, Sur de Brasil, Chile, Paraguay, Uruguay). Missouri: Missouri Botanical Garden Press. [ Links ]

