










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkDarwiniana, nueva serie
versión impresa ISSN 0011-6793versión On-line ISSN 1850-1699
Darwiniana, nueva serie vol.2 no.1 San Isidro jun. 2014
ARTÍCULO
Alcances y limitaciones para la identificación arqueológica de gránulos de almidón de razas nativas de Zea mays (Poaceae) del Noroeste argentino
Irene Lantos1, Marco A. Giovannetti2 & Norma Ratto1
1 Museo Etnográfco J. B. Ambrosetti, Facultad de Filosofía y Letras, Universidad de Buenos Aires, Moreno 350, C1091AAH Ciudad Autónoma de Buenos Aires, Argentina; nratto@flo.uba.ar (autor corresponsal).
2 División Arqueología, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Paseo del Bosque s/n, B1900FWA La Plata, Buenos Aires, Argentina.
Editor Asociado: Fernanda Rodriguez
Original recibido el 14 de agosto de 2013,
aceptado el 21 de marzo de 2014
Abstract
Scope and limitations for the archaeological identification of starch grains from native landraces of Zea mays (Poaceae) in Northwestern Argentina. In this paper we characterized and analyzed microscopically starch grains of eight Zea mays modern landraces from Northwestern Argentina (pisingallo, cristalino colorado, perlita, calchaquí, complejo tropical, dentado amarillo-colorado, chullpi, and capia). We created a reference database of local varieties to explore the diversity in the morphological and metric characters of starch grains at intra and inter-racial levels. The aim of this paper was to search for referential elements that could allow better and more reliable identifications of maize starches in archaeological samples. We used descriptive statistics and multivariate analyses; we applied a statistical treatment to estimate the probability of correct classification of starch grains. Results showed high variability within each landrace and a large superposition between varieties. None of the landraces showed distinctive characteristics that allowed an exclusive discrimination.
Key words: Archaeobotany; Landraces; Starch; Zea mays
Resumen
Se caracterizaron y analizaron microscópicamente los gránulos de almidón de ocho razas nativas de Zea mays del noroeste de Argentina (pisingallo, cristalino colorado, perlita, calchaquí, complejo tropical, dentado amarillo-colorado, chullpi y capia). Se generó una base de datos de referencia con caracteres morfológicos y métricos de los gránulos de almidón de las razas locales para dar cuenta de la diversidad a nivel intra e inter-racial. El objetivo de este trabajo es reconocer elementos referenciales para lograr mejores y más confiables identificaciones de los gránulos de almidón de maíz en muestras arqueológicas. Para este fin se utilizaron herramientas estadísticas descriptivas y se realizaron análisis multivariados; se aplicó un tratamiento estadístico de los datos para estimar la clasificación de los gránulos de almidón por razas. Los resultados mostraron una alta variabilidad para cada raza y una gran superposición entre las razas. Ninguna de las razas de maíz presentó características distintivas que permitan una discriminación en grupos.
Palabras clave: Almidón; Arqueobotánica; Razas nativas; Zea mays
INTRODUCCIÓN
La multiplicidad de razas autóctonas de Zea mays L. y su amplia dispersión a lo largo de la América prehispánica ha sido estudiada en profundidad en las últimas cinco décadas. Una significativa cantidad de trabajos han registrado una amplia diversidad racial a partir de las características morfológicas de cariopsis y marlos a nivel macroscópico (Parodi, 1959; Abiusso & Cámara Hernández, 1974; Itlis & Doebley, 1980; Galinat, 1988; McBird, 1994; Miante Alzogaray & Cámara Hernández, 1996; Rodríguez & Aschero, 2007; Oliszewski & Olivera, 2009).
Asimismo, con la finalidad de ampliar líneas de evidencia alternativas y complementarias, se han realizado identificaciones microscópicas de almidones y fitolitos de Zea mays en muestras de sedimentos y artefactos arqueológicos (Babot & Apella, 2003; Coil et al., 2003; Haslam, 2004; Torrence & Barton, 2006; Dickau et al., 2007; Holst et al., 2007; Boyd et al., 2008; Babot, 2009; Giovannetti, 2009; Piperno, 2009; Korstanje & Cuenya, 2010) pero las características morfológicas y métricas a nivel racial permanecen aún inexploradas en los microrrestos, a excepción de investigaciones puntuales sobre almidones de razas de América central y el Caribe. Tal es el caso de Piperno & Holst (1998) que hallaron almidones arqueológicos similares a aquellos de las razas modernas Pepetilla, Nal Tel y Harinoso de Ocho, que se diferenciaron de los almidones de maíz reventón argentino. También, Pagán Jiménez (2007) observó almidones de maíz en muestras arqueológicas que pudo adscribir a razas harinosas o córneas.
Sin embargo, estas aproximaciones metodológicas no están exentas de debate y sobre todo de la necesidad de profundizar los estudios arqueológicos. Específicamente en relación al maíz, la investigación sobre gránulos de almidón antiguo preservados en contextos arqueológicos ha producido algunos cuestionamientos, tal como se evidencia en las identificaciones de fitolitos y almidones de Zea mays en sitios arqueológicos cronológicamente muy tempranos (Staller et al., 2006). El interrogante principal se debe a que los hallazgos más antiguos de dichos microrrestos, incluso en contextos de sociedades cazadoras recolectoras (Babot, 2005), no se correlacionan necesariamente con la presencia de macrorrestos de Zea mays tales como marlos y cariopsis desecados o carbonizados.
Por otra parte, existe otro problema en cuanto a los caracteres propios de la biología de las plantas. En muchos taxones se registra una gran variabilidad de morfologías y tamaños de gránulos de almidón que da cuenta de una multiplicidad de factores genotípicos y fenotípicos, y que pueden dificultar las identificaciones arqueológicas. Un caso representativo es el de los almidones transitorios (Reichert, 1913) de tamaño pequeño y morfología discoidal que son producidos en los cloroplastos de las plantas y que, a diferencia de los almidones de reserva, no están controlados genéticamente. Otros ejemplos de interés podemos tomarlos de almidones de reserva de Phaseolus vulgaris L. (poroto) (Babot et al., 2007), Prosopis L. (algarroba) (Giovannetti et al., 2008), Triticum L. (trigo) (Piperno et al., 2004) y muchos tipos de Zea mays que presentan caras facetadas en la estructura morfológica de los gránulos de almidón.
La construcción de bases de referencia confiables de almidones de maíces nativos actuales tiene el potencial de facilitar la identificación de los materiales arqueobotánicos de escala microscópica. En este sentido, destacamos algunas aproximaciones al estudio de almidones de plantas útiles, tales como los trabajos de construcción de bases referenciales y análisis estadísticos de Torrence et al. (2004) para especies de Papúa Nueva Guinea y de Lentfer (2009) para especies de Indonesia.
Cabe subrayar que, aunque se utilizan técnicas de ADN de manera corriente para el estudio de la variabilidad racial en maíces actuales (Lia et al., 2009; Bracco et al., 2012), el estudio de material genético antiguo en muestras arqueológicas es dificultoso y la tasa de recuperación de restos arqueobotánicos aptos y en buenas condiciones para análisis es relativamente baja (Lia et al., 2007). En contraste, la presencia de gránulos de almidón es muy frecuente en materiales arqueológicos de piedra y cerámica utilizados para la molienda y procesamiento de alimentos. Estos materiales porosos actúan como trampas y protegen a los residuos de gránulos de almidón de los agentes ambientales de degradación y por lo tanto sobreviven en mayor medida. A partir de esto, este trabajo abordó el estudio de almidones fundamentalmente porque constituye un indicador bioarqueológico importante por su ubicuidad y facilidad de recuperación para el análisis.
Giovannetti et al. (2012) publicaron un análisis acotado de cuatro razas nativas de maíz (capia, pisingallo, dentado amarillo-colorado y chullpi) procedentes del noroeste de la Argentina a partir de una muestra reducida de almidones (N total=195). Como conclusión preliminar, corroborada por análisis estadísticos descriptivos y multivariados, se observó una gran variabilidad morfológica y de tamaño en cada raza y un alto solapamiento entre ellas; se demostró una superposición parcial entre las razas dentado, chullpi y pisingallo, mientras que la raza capia pudo ser diferenciada en mayor medida. En el presente trabajo se planteó ampliar el tamaño de la muestra, tanto de razas como de cantidad de cariopsis y gránulos de almidón con respecto al estudio preliminar citado al comenzar este párrafo.
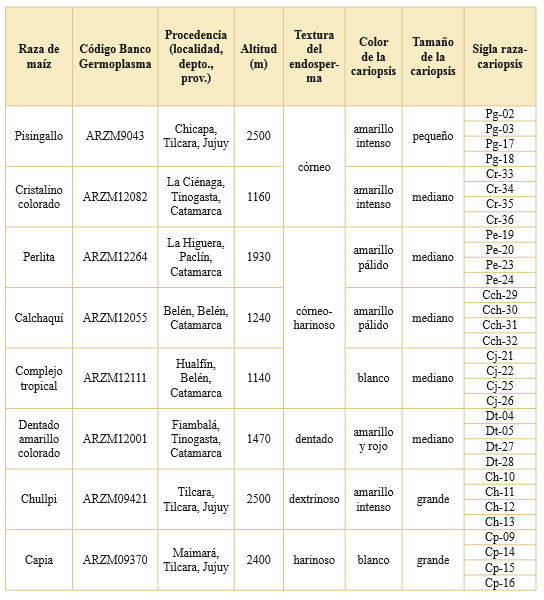
El objetivo del presente estudio fue caracterizar un conjunto amplio de gránulos de almidón de razas nativas de maíz del Noroeste argentino. Dado que esta especie presenta una gran variabilidad intra e interracial, que se evidencia en múltiples morfologías, dimensiones y adaptabilidad ambiental, seleccionamos ocho razas nativas y cultivadas en las provincias de Catamarca y Jujuy (Tabla 1) para generar una muestra representativa tanto a nivel cuantitativo como cualitativo que permita identificar elementos distintivos para cada raza, sobre la base de análisis descriptivos y estadísticos.
Tabla 1. Composición de la muestra de maíces nativos analizada. El tamaño de las cariopsis fue categorizado según su longitud en grande (2 a 1,5 cm), mediano (1,5 a 1 cm) y pequeño (1 a 0,5 cm).
Por lo tanto, el aporte de este trabajo a la arqueología es la caracterización de almidones de maíz del Noroeste argentino, que pueda servir en la comparación con otras razas regionales. En este sentido, buscamos generar elementos referenciales para mejores y más confiables identificaciones de los gránulos de almidón de maíz en muestras arqueológicas. Finalmente, este trabajo se enfoca en los alcances y las limitaciones de esta técnica para la identificación de razas nativas en muestras arqueológicas.
MATERIALES Y MÉTODOS
Conformación de la muestra
Las muestras de cariopsis de las razas nativas actuales estudiadas se encontraban depositadas en el Banco de Germoplasma del Instituto Nacional de Tecnología Agropecuaria INTA-Pergamino y fueron producto de las recolecciones sistemáticas realizadas en diferentes zonas del Noroeste argentino en las décadas de 1960 y 1970. El Banco clasificó las muestras por raza de acuerdo con los descriptores fenotípicos de grano y marlo, como textura, color, forma, ancho, largo y número de filas en el marlo (Solari & Gómez, 1997; Ferrer et al., 1998). Las muestras fueron secadas a temperatura ambiente y presión atmosférica con control de humedad, preservadas en bolsas de papel selladas y guardadas a 5-10 °C y 40-50% de humedad relativa.
En este estudio se seleccionaron ocho razas de maíces nativos del Noroeste argentino (Tabla 1, Fig. 1), cuatro fueron recolectados en la provincia de Jujuy entre 1963 y 1968, y las otras cuatro en la provincia de Catamarca en 1977, clasificados por el Banco de Germoplasma (INTA) como: pisingallo (ARZM9043), cristalino colorado (ARZM12082), perlita (ARZM12264), calchaquí (ARZM12055), complejo tropical (ARZM12111), dentado amarillo colorado (ARZM12001), chullpi (ARZM09421) y capia (ARZM09370). A su vez, fueron clasificados por la textura dominante de su endosperma en los siguientes grupos: córneo, córneo-harinoso, dentado, dextrinoso y harinoso (Abiusso & Cámara Hernández, 1974; Molestina, 1987; Oliszewski, 2008). Se seleccionaron cuatro cariopsis por raza y se analizaron en total 800 gránulos de almidón procedentes de 32 cariopsis de maíces nativos actuales.

Fig. 1. Cariopsis de las ocho razas de maíz estudiadas. A, capia; B, chullpi; C, pisingallo; D, calchaquí; E, cristalino colorado; F, dentado amarillo-colorado; G, perlita; H, complejo tropical. Escala = 10 cm. Figura en color en la versión en línea http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/551/585.
Las cariopsis fueron seccionadas con bisturí y se identificaron las dos zonas del endosperma por sus texturas córnea y harinosa, y se trabajó solamente con la porción del endosperma harinoso que permitió una fácil separación e individualización de los gránulos de almidón. No se estudiaron los almidones del endosperma compacto porque la única manera de individualizarlos es por molienda, la cual produce alteraciones en los gránulos de almidón y no cumple con los requisitos de integridad de la muestra requeridos en este estudio.
Las muestras fueron extraídas con un raspado suave y continuo con instrumento de metal sobre la zona del endosperma harinoso. Este método minimiza las alteraciones producidas por la manipulación durante el montaje de la muestra y es de uso corriente en los estudios microscópicos de gránulos de almidón (Lavalle & Mengascini, 2007). El procedimiento se realizó en un ambiente controlado, limpio y hermético para evitar contaminación y se utilizaron guantes sin polvo; además, el instrumento de extracción metálico fue esterilizado antes de cada uso. Todo el material de vidrio utilizado fue nuevo y enjuagado con alcohol previo a su uso. El montaje se realizó con una solución de glicerol: agua 1:1 (Cortella & Pochettino, 1994; Coil et al., 2003).
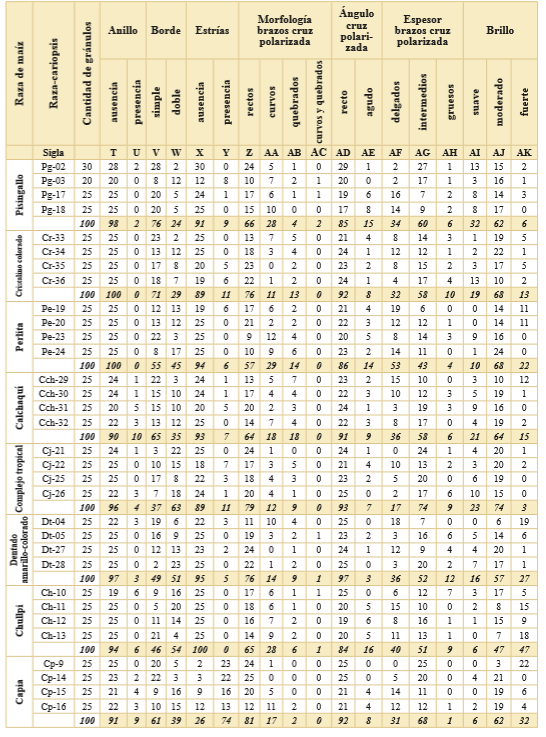
De cada raza de maíz actual se analizaron 100 gránulos de almidón elegidos al azar, los cuales fueron observados con microscopio óptico y caracterizados según las variables detalladas en la Tabla 3. Se excluyeron del estudio los gránulos de almidón que presentaran alteraciones propias del manejo instrumental como el corte del bisturí o el raspado posterior.
Tabla 3. Registro de variables de los gránulos de almidón por cariopsis de cada una de las razas de maíz analizadas.

Selección y análisis de variables
Se registraron 11 variables y sus estados, 37 en total, para los 800 gránulos de almidón de las ocho razas de maíz nativo actual estudiadas (Tabla 3).
Se analizaron 100 gránulos de almidón por raza, las frecuencias son equivalentes a los porcentajes en que se presenta cada estado de variable. Cada uno de los nombres de dichos estados fue abreviado con una sigla (A - AK).
La definición, caracterización y atributos de las variables fueron extensamente desarrolladas por Giovannetti et al. (2012). Las variables seleccionadas para este análisis caracterizan los rasgos morfológicos y métricos de los gránulos de almidón. Se tuvieron en cuenta las variables clásicas, tales como la morfología general, el tamaño y los atributos del hilio (Reichert, 1913; Loy, 1992). También se incluyeron variables utilizadas en estudios más recientes (Loy, 1992; Babot, 2007; Pagán Jiménez, 2007): presencia de anillos y estrías, tipos de bordes y brillo de la cruz bajo luz polarizada. Se usaron además algunas variables propuestas especialmente para este estudio que describen en mayor detalle el carácter de los brazos de la cruz de polarización: morfología, ángulo y espesor.
Registro microscópico de los gránulos de almidón y tratamiento estadístico
Las observaciones microscópicas fueron realizadas con microscopio Lancet XSZ-148 a 400 aumentos alternando luz normal con polarizada. Para este último caso se mantuvo constante tanto la intensidad de la luz de entrada del microscopio (nivel siete) como la apertura del diafragma (nivel cinco). Se tomaron fotografías micrográficas con una cámara Samsung modelo HZ10W 10.2 mega-pixeles equipada con un adaptador para el tubo ocular del microscopio. El análisis digital de las imágenes se hizo con el programa ImageJ 1.47 (National Institutes of Health, 2013).
Para el tratamiento numérico de los datos se aplicó una estadística descriptiva e inferencial, univariada y multivariada (Shennan, 1992; Blalock, 1996; Drennan, 1996; Moschetti et al., 2003). La mayor parte de las variables del gránulo son cualitativas nominales y para su análisis estadístico fue necesario tratarlas al nivel de la cariopsis previa transformación. Además, se tuvo en cuenta la proporción entre la cantidad de casos y la cantidad de variables a interrelacionar en el análisis multivariado; por ejemplo, estos análisis no pueden realizarse con una muestra de 32 casos (cariopsis) y 37 estados de variables. Todos estos aspectos determinaron el uso de un procedimiento para la transformación de los datos y/o la aplicación combinada de distintos métodos estadísticos multivariados, que se explican más abajo. Los datos fueron analizados con los programas SPSS 19.0 (IBM, 2010) y PAST 2.17c (Hammer et al., 2001).
De las 11 variables consideradas (Tabla 3) solo una es continua (diámetro máximo del gránulo) y permitió calcular la media, la desviación estándar, mínimos y máximos, asimetría, curtosis y coeficiente de variación. Las restantes variables presentan estados cualitativos que fueron útiles para determinar si los valores de una variable particular diferían o no entre dos o más grupos. Para éstas, se usó la prueba no paramétrica de Kruskal-Wallis (K-W) porque no requiere el cumplimiento de los supuestos de igualdad de varianzas, así como tampoco la distribución normal de de las variables. Para ello, las frecuencias de los estados de cada una de las variables registradas en las 32 cariopsis fueron asignadas a un “ranking” en función de su abundancia relativa, considerándose el rango consignado en la Tabla 2. Igual criterio se aplicó a la única variable cuantitativa (diámetro máximo) por no cumplir con los supuestos citados. De esta manera, se generó un rango jerárquico de abundancia relativa para los 37 estados que caracterizan cada cariopsis, oscilando los valores entre uno (cuando no hay registro del estado) y siete (cuando el estado es mayor a 25 registros). Esto permitió la creación de una matriz compuesta por 32 filas (cariopsis) y 37 columnas (los estados de cada variable). La hipótesis nula sostuvo que las cariopsis pertenecen a la misma población de maíces, considerando cada uno de los estados que toman las variables del análisis (para consultar variables y estados ver Tabla 3). La hipótesis alternativa fue la contraria. De esta manera, la abundancia relativa “rankeada” de cada estado de las variables fue contrastada contra dos niveles de agrupación: (a) razas de maíces (8 grupos) y (b) la clasificación por textura del endosperma (5 grupos). Se determinó como nivel de significación p= 0,05.
Tabla 2. Transformación de los estados de las variables cualitativas de las cariopsis (N=32) a escala ordinal

La prueba no paramétrica de K-W permitió identificar aquellos estados de variables que diferenciaban a los maíces a uno y otro nivel de agrupación considerado. Esos estados fueron seleccionados como nuevas variables para analizarlas a través de un tratamiento numérico multivariado al nivel del gránulo (N = 800). Primero se realizó un Análisis de Correspondencia Múltiple (ACM), ya que esta técnica permite analizar la relación existente entre variables categóricas, como así también conocer la forma en que está estructurada esa relación. Para cada gránulo se registró la presencia o ausencia de las nuevas variables seleccionadas. Luego, sobre la base de los ejes dimensionales obtenidos por ACM, se realizó un Análisis Discriminante (AD) para corregir la conformación de los grupos mediante la distancia de Mahalanobis. La utilidad de esta medida de distancia radica en que es una forma de determinar la similitud entre dos variables aleatorias multidimensionales (Muñóz Salas, 1998; Portillo & Salas, 2008). Además, el AD contiene, en las primeras funciones discriminantes, la expresión de las mayores cargas de cada variable. De esta manera, cada caso es asignado a un grupo pronosticado con una probabilidad determinada de pertenencia.
Los resultados de este trabajo se presentan en tres secciones: (a) análisis descriptivo de los datos cualitativos y cuantitativos de los gránulos de almidón (N= 800) registrados microscópicamente en cada raza; (b) test de hipótesis no paramétricos para explorar la variabilidad en la muestra de cariopsis (N = 32); y (c) análisis multivariado (Análisis de Correspondencia Múltiple y Análisis Discriminante) para conocer la estructura subyacente de la matriz de datos a nivel de los gránulos de almidón (N = 800) y para evaluar la factibilidad de diferenciar estadísticamente los almidones de las distintas razas de maíz.
RESULTADOS
Análisis descriptivo de los datos cualitativos y cuantitativos
Las características morfológicas y métricas de los gránulos de almidón de cada raza de maíz pueden observarse en la Fig. 2. En la Tabla 3 se presenta el registro de los estados de las variables de los gránulos de almidón en las cariopsis de cada raza analizada. De la Tabla 3 se desprenden las siguientes observaciones:
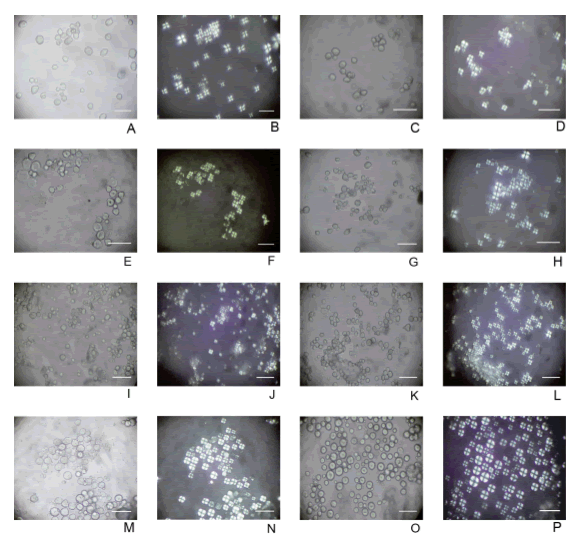
Fig. 2. Microfotografías de gránulos de almidón de cada raza de maíz, tomadas a 400 aumentos con luz normal y polarizada: A, pisingallo, luz normal; B, pisingallo, luz polarizada; C, cristalino colorado, luz normal; D, cristalino, colorado, luz polarizada; E, perlita, luz normal; F, perlita, luz polarizada; G, calchaquí, luz normal; H, calchaquí, luz polarizada; I, complejo tropical, luz normal; J, complejo tropical, luz polarizada; K, dentado amarillo colorado, luz normal; L, dentado amarillo colorado, luz polarizada; M, chullpi, luz normal; N, chullpi, luz polarizada; O, capia, luz normal; P, capia, luz polarizada. Escala = 50 µm. Figura en color en la versión en línea http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/551/585.
Pisingallo. El conjunto de almidones de la raza pisingallo presentó un tamaño promedio de 12,14 mm y el rango entre 6-9 mm fue el más poblado. Las morfologías fueron mayormente circulares (71%), con menor incidencia de ovaladas (14%), y gránulos de almidón con caras rectas o facetados (10%), campanuliformes (4%) e irregulares (1%).
Los hilios fueron por lo general puntiformes (60%) y en posición central o subcentral (70%). En menor proporción se encontraron ejemplares con hilios no visibles (32%) y filiformes (8%) y ausencia total de trífidos. Solo un 2% de los gránulos de almidón presentaron anillos de crecimiento y un 9% estrías radiales, mientras que el 76% tuvo bordes simples. En cuanto a los rasgos de la cruz de polarización, un 66% presentó brazos rectos y un 28% curvos, mayormente posicionados en ángulo recto (85%) y de espesor intermedio (60%) o delgado (34%). El brillo de la cruz de polarización fue mayormente moderado (62%) a suave (32%).
Cristalino colorado. El conjunto de almidones de la raza cristalino colorado presentó un tamaño promedio de 12,87 mm y el rango entre 9-12 mm fue el más poblado. Las morfologías fueron mayormente circulares (50%) con menor incidencia de gránulos de almidón con una o dos caras rectas (28%) ovalados (11%), poliédricos (8%) e irregulares (3%). Los hilios fueron por lo general puntiformes (50%) y en posición central o subcentral (82%). En menor proporción se encontraron ejemplares con hilios filiformes (34%), no visibles (14%) y trífidos (2%). No se presentaron anillos de crecimiento y un 11% tuvo estrías radiales, mientras que el 71% tuvo bordes simples. En cuanto a los rasgos de la cruz de polarización, un 76% presentó brazos rectos, 13% quebrados y 11% curvos, mayormente posicionados en ángulo recto (92%) y de espesor intermedio (58%) o delgado (32%). El brillo de la cruz polarización fue mayormente moderado (68%) aunque en iguales proporciones apareció suave (19%) y fuerte (19%).
Perlita. El conjunto de almidones de la raza perlita presentó un tamaño promedio de 13,04 mm y el rango entre 9-12 mm fue el más poblado. Las morfologías fueron mayormente circulares (52%) con menor incidencia de gránulos de almidón ovalados (28%), poliédricos (9%), con caras rectas (8%) irregulares (2%) y campanuliformes (1%). Los hilios más frecuentes fueron puntiformes (59%), filiformes (27%), no visibles (13%) y trífidos (1%). Se encontraron mayormente en posición central o subcentral (82%) con menor proporción de casos con hilios excéntricos (36%). No se presentaron anillos de crecimiento y un 6% tuvo estrías radiales. Los bordes dobles representaron el 45% y los simples el 55%. En cuanto a los rasgos de la cruz de polarización, un 57% presentó brazos rectos, 29% curvos y 14% quebrados, mayormente posicionados en ángulo recto (86%) y de espesor intermedio (58%) o delgado (32%). El brillo de la cruz polarización fue mayormente moderado (68%) mientras que un 22% presentó brillo fuerte.
Calchaquí. El conjunto de almidones de la raza calchaquí presentó un tamaño promedio de 12,66 mm, siendo el rango entre 9-12 mm el más poblado. Las morfologías fueron mayormente circulares (53%) con menor incidencia de gránulos de almidón ovalados (17%), con caras rectas (15%), irregulares (12%), campanuliformes (2%) y poliédricos (1%). Los hilios más representados fueron los puntiformes (64%), seguidos por los no visibles (20%) y los filiformes (16%). Se ubicaron mayormente en posición central o subcentral (77%) con menor proporción de casos con hilios excéntricos (23%). Se presentaron 10% de casos con anillos de crecimiento y un 7% tuvo estrías radiales. Los bordes dobles representaron el 35% y los simples el 65% de los casos. En cuanto a los rasgos de la cruz de polarización, un 64% presentó brazos rectos, 18% curvos y otro 18% quebrados, mayormente posicionados en ángulo recto (91%) y de espesor intermedio (58%) o delgado (36%). El brillo de la cruz polarización fue mayormente moderado (64%) a suave (21%) mientras que un (15%) presentó brillo fuerte.
Complejo tropical. El conjunto de almidones de la raza complejo tropical presentó un tamaño promedio de 11,36 mm y el rango entre 9-12 mm fue el más poblado. Las morfologías fueron mayormente circulares (56%) con menor incidencia de gránulos de almidón con caras rectas (25%), ovalados (15%), poliédricos (2%), irregulares (1%) y campanuliformes (1%). Dominaron los hilios puntiformes (79%), seguidos por los filiformes (12%) y los no visibles (9%) y la ausencia total de trífidos; adquirieron mayormente posición central o subcentral (63%) con menor proporción de casos con hilios excéntricos (37%). Se presentaron 4% de casos con anillos de crecimiento y un 11% tuvo estrías radiales. Los bordes dobles representaron el 63% y los simples el 37% de los casos. En cuanto a los rasgos de la cruz de polarización, un 79% presentó brazos rectos, 12% curvos y 9% quebrados, mayormente posicionados en ángulo recto (93%) y de espesor intermedio (74%) o delgado (17%). El brillo de la cruz polarización fue mayormente moderado (74%) y en menor proporción suave (23%).
Dentado amarillo-colorado. El conjunto de almidones de la raza dentado amarillo-colorado presentó un tamaño promedio de 14,66 mm, estableciéndose en los rangos 9-12 mm (26%), 12-15 (22%) y 15-18 (19%) los más representados. Las morfologías más abundantes fueron circulares (48%) con menor incidencia de gránulos de almidón ovalados (19%), con caras rectas (14%), irregulares (14%) y poliédricos (5%). Los hilios que adquirieron el estado puntiforme mostraron un 52%, seguidos por los no visibles (25%), filiformes (22%) y trífidos (1%). Todos se encontraron mayormente en posición central o subcentral (74%) con menor proporción de casos con hilios excéntricos (26%). Se presentaron 3% de casos con anillos de crecimiento y un 5% tuvo estrías radiales. Los bordes dobles representaron el 51% y los simples el 49% de los casos. En cuanto a los rasgos de la cruz de polarización, un 76% presentó brazos rectos, 14% curvos y 9% quebrados 1% de curvos quebrados y mayormente posicionados en ángulo recto (97%) y de espesores intermedios (52%) o delgados (36%). El brillo de la cruz polarización fue mayormente moderado (57%) a fuerte (27%).
Chullpi. El conjunto de almidones de la raza chullpi presentó un tamaño promedio de 16,47 mm y los rangos rango de tamaño más poblados fueron los de 15-18 mm (33%) y 18-21 mm (24%). Las morfologías fueron mayormente circulares (86%) con menor incidencia de gránulos almidón ovalados (5%), con caras rectas (5%), campanuliformes (2%), irregulares (1%) y poliédricos (1%). Los hilios más frecuentes fueron los filiformes (46%), no visibles (33%), puntiformes (27%) y trífidos (6%). Se encontraron mayormente en posición central o subcentral (78%) con menor proporción de casos con hilios excéntricos (22%). Se presentaron 6% de casos con anillos de crecimiento y ninguno tuvo estrías radiales. Los bordes dobles representaron el 54% y los simples el 46% de los casos. En cuanto a los rasgos de la cruz de polarización, 65% presentó brazos rectos, 28% curvos, 6% quebrados y 1% curvos y quebrados, mayormente posicionados en ángulo recto (84%) y de espesor intermedio (51%) o delgado (46%). El brillo de la cruz polarización fue moderado (47%) a fuerte (47%).
Capia. El conjunto de almidones de la raza capia presentó un tamaño promedio de 16,47 mm y los rangos de tamaño más importantes fueron los de 15-18 mm (44%) y 18-21 mm (23%). Se destacaron las morfologías circulares (81%) con menor incidencia de gránulos de almidón con caras rectas (9%), poliédricos (4%), irregulares (3%), campanuliformes (2%) y ovalados (1%). Dominaron los hilios que adquirieron los estados puntiforme (53%), no visible (25%), filiforme (21%) y trífido (1%), especialmente en posición central o subcentral (85%) y en menor proporción hilios excéntricos (15%). Se presentaron 9% de casos con anillos de crecimiento y un 74% de estrías radiales. Los bordes dobles representaron el 39% y los simples el 61% de los casos. En cuanto a los rasgos de la cruz de polarización, 81% presentó brazos rectos, 17% curvos y 2% quebrados, mayormente posicionados en ángulo recto (92%) y de espesor intermedio (68%) o delgado (31%). El brillo de la cruz polarización fue mayormente moderado (62%) a fuerte (32%).
Análisis estadístico de los datos
En la Tabla 4 se presenta la estadística descriptiva de la variable diámetro máximo del gránulo para cada una de las cariopsis, consignando la cantidad de gránulos de almidón, la media, la desviación estándar, el máximo y el mínimo, la asimetría, la curtosis y el coeficiente de variación. Se observó que en general los coeficientes de variación fueron altos dando cuenta de una alta variabilidad de tamaños de gránulos de almidón al interior de cada cariopsis. Por lo tanto, optamos por considerar que la distribución de los datos no es normal y analizamos su comportamiento mediante la aplicación de test no paramétricos, al igual que en los casos de las variables cualitativas.
Tabla 4. Estadística descriptiva del diámetro máximo del gránulo de almidón de las cariopsis de las razas de maíz. Medidas en µm. Referencias de siglas en la Tabla 1. CV: coeficiente de variación; s: desvío estándar.

Se realizó la prueba no paramétrica de K-W considerando como variable a contrastar a cada uno de los estados de las variables (A-AK en Tabla 3) y la variable de agrupación fue seleccionada tanto al nivel de raza (Tabla 5) como de la textura del endosperma (Tabla 6). Pudo observarse que la mayoría de los estados no rechazaron la hipótesis nula del test K-W a ninguno de los dos niveles de agrupación considerados. La excepción a lo dicho fueron los estados que se expresan en la Tabla 7.
Tabla 5. Resultados del test Kruskal-Wallis de los estados registrados para las 32 cariopsis de maíces analizados. Nivel de agrupamiento por razas de maíces (7 grados de libertad). Se resaltan en color los estados que rechazan la H0. En la columna estado de contraste se consignan los estados de las variables (Tabla 3) en función de las variables transformadas (Tabla 2).
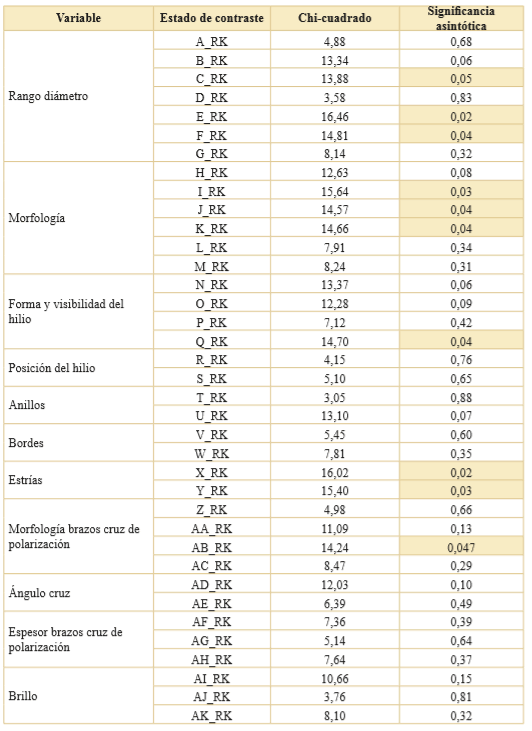
Tabla 6. Resultados del test de Kruskal-Wallis de los estados registrados en las 32 cariopsis de maíces estudiados. Nivel de agrupamiento por textura del endosperma (4 grados de libertad). Se resaltan en color los estados que rechazan la H0. En la columna estado de contraste se consignan los estados de las variables (Tabla 3) en función de las variables transformadas (Tabla 2).
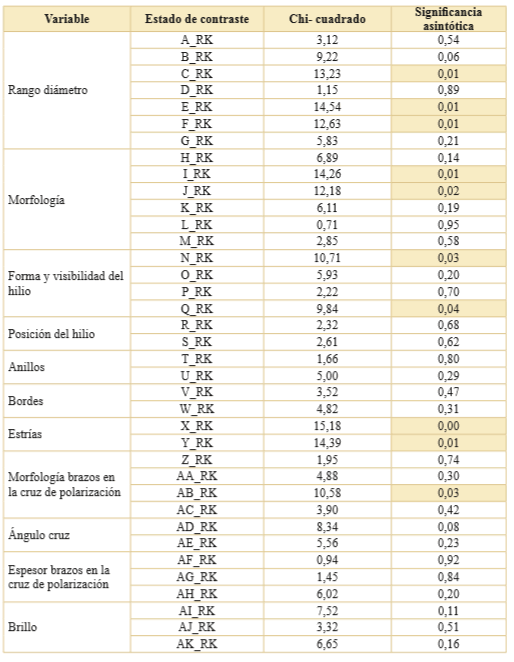
Tabla 7. Resumen de los estados de las variables que rechazan la H0 del test de Kruskal-Wallis e indican que las cariopsis de maíz (N = 32) pertenecen a poblaciones diferentes.
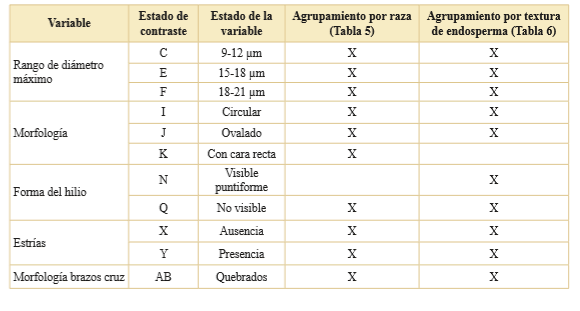
En resumen, el test de K-W mediante el cruce de un estado con los criterios de agrupación seleccionados indicó que en el caso del agrupamiento por raza de maíz solo 10 sobre los 37 estados registrados dieron cuenta de la diferenciación entre razas nativas. Cuando usamos el segundo agrupamiento, clasificación general por textura del endosperma, otros diez estados cumplieron esa condición. Además, se observó que los estados morfología con cara recta, hilio visible puntiforme quebrados difieren según el tipo de agrupamiento elegido (Tabla 7). Por lo tanto, los 11 estados que dan cuenta de diferencias en la población de maíces, independiente del nivel de agrupamiento seleccionado, fueron considerados para la realización de la estadística multivariada siguiendo el procedimiento expresado en Materiales y Métodos.
El ACM generó 10 ejes dimensionales que explican el 81,2 % de la varianza total, ubicándose las variables en forma diferencial dentro del espacio dimensional (Fig. 3). El porcentaje de la varianza explicada se distribuyó de la siguiente manera: Eje 1 17,206%, Eje 2 14,723%, Eje 3 13,667%, Eje 4 12,917%, Eje 5 12,651%, Eje 6 10,028%, Eje 7 9,878%, Eje 8 5,446%, Eje 9 2,514%, Eje 10 0,969. En la Fig. 3 se graficaron los primeros dos ejes, cuyas puntuaciones para cada variable fueron: C -0,61856, 0,010742; E 0,92675, 0,49513; F 0,42183, -1,1997; I 0,29663, -0,27099; J -1,0238, -0,67563; K -0,42083, 1,9483; N -0,26007, 0,28347; Q 0,39179, -0,85461; X -0,32911, -0,06752; Y 1,9672, 0,4424; AB -0,25596, -0,25596. Luego, sobre la base de los ejes dimensionales del ACM se realizó un AD para corregir la conformación de los grupos mediante la distancia de Mahalanobis (Fig. 4).
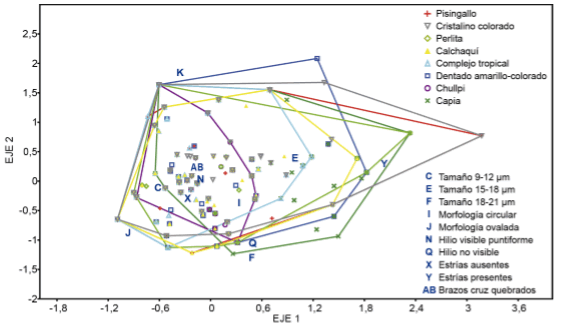
Fig. 3. Ubicación de las variables analizadas dentro del espacio dimensional. Tamaño de muestra N=800. Figura en color en la versión en línea http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/551/585.
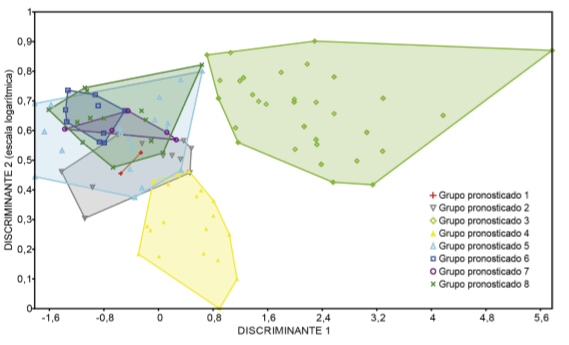
Fig. 4. Grupos pronosticados por el análisis discriminante realizado a nivel de agrupación por raza (ocho grupos). Figura en color en la versión en línea http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/551/585.
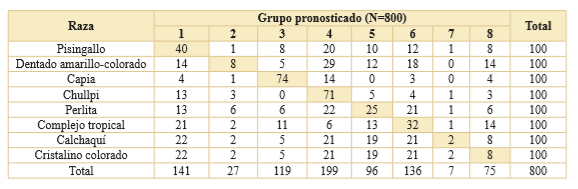
El análisis multivariado indica que, a nivel de clasificación por raza, ninguna presenta características distintivas que permitan discriminarlas en grupos exclusivos (Tabla 8; Fig. 4). La situación ideal de una identificación excluyente es que el 100% de los casos de una raza sean clasificados dentro del grupo discriminante correspondiente a la misma. Esta situación no se encontró en nuestro estudio, pero existieron algunas tendencias que resultan relevantes. El caso más auspicioso fue el de la raza capia cuya probabilidad de correcta clasificación fue del 74%. Para chullpi esta probabilidad descendió al 71% y para el resto de las razas osciló entre el 40% y el 2%.
Tabla 8. Asignación de grupos pronosticados para cada raza de maíz nativo. Los valores sombreados corresponden a la probabilidad de correcta clasifcación de los gránulos.
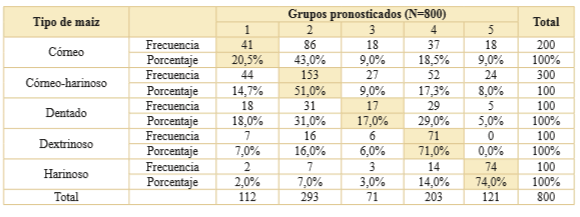
Sobre la base de los mismos ejes del ACM, se repitió el procedimiento estadístico para el AD utilizando el nivel de agrupamiento de la clasificación por textura del endosperma. Se obtuvieron los resultados expresados en la Fig. 5 y la Tabla 9. Se observó que la situación no cambió ya que el maíz harinoso se siguió diferenciando de los otros grupos pero los centroides del maíz córneo y del maíz córneo-harinoso continúan estando muy próximos, al igual que el maíz dentado con el maíz dextrinoso. En este caso, las probabilidades de correcta clasificación fueron de 74% para los harinosos, 71% para los dextrinosos, 51% para los córneo-harinosos, 20,5% para los córneos y 17% para los dentados.

Fig. 5. Grupos pronosticados por el análisis discriminante realizado a nivel de agrupación por textura del endosperma (cinco grupos). Figura en color en la versión en línea http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/551/585.
Tabla 9. Asignación de grupos pronosticados en función de la clasifcación general por textura del endosperma. Los valores sombreados corresponden a la probabilidad de correcta clasifcación de los gránulos.
DISCUSIÓN
En este trabajo consideramos las clasificaciones de los maíces a dos niveles. Por un lado, partimos de la clasificación por raza siguiendo la implementada por el Banco de Germoplasma y por otro, clasificamos las razas por la textura de su endosperma (Abiusso & Cámara Hernández, 1974; Miante Alzogaray & Cámara Hernández, 1996; Oliszewski, 2008). Las razas analizadas fueron tratados numéricamente tanto al nivel de la cariopsis (N = 32) como del gránulo (N = 800) implementando diferentes métodos estadísticos acordes con la naturaleza de los datos donde predominaron las variables cualitativas. Esto obligó a la transformación de variables cualitativas nominales a ordinales por un “ranking” en función de su abundancia relativa.
La prueba de K-W aplicada sobre los 37 estados que conforman las 11 variables analizadas dio cuenta de que solo en 11 estados existen diferencias significativas en la población de maíces. El resultado del análisis multivariado indicó que ninguna de las razas de maíz presenta características que permita discriminarlas en grupos distintivos. Ningún estado de variable es característico de una única raza. Solo dos casos - capia y chullpi- presentaron una razonable probabilidad de correcta clasificación, mayor al 70%, para los dos niveles de agrupación implementados. En el resto, los mayores errores de clasificación se dan al nivel del agrupamiento por raza (Tabla 8). Un caso extremo es la raza calchaquí donde solo el 2% de los 100 gránulos de almidón analizados fueron correctamente clasificados. Por otra parte, observamos que cuando el agrupamiento fue al nivel de la textura del endosperma se obtuvieron resultados levemente más auspiciosos (Tabla 9). Al respecto, para la clasificación córneo-harinoso (razas cristalino colorado, perlita, calchaquí y complejo tropical) se obtuvo un 51% de registros correctamente clasificados, cuando con el agrupamiento por raza osciló entre el 2% y el 32% de casos bien asignados.
Estos resultados le otorgan especial importancia a la clasificación por textura del endosperma en relación con su aplicación en arqueobotánica. Los maíces con distintos endospermas fueron utilizados para diferentes comidas tanto en tiempos históricos como actuales (Cámara Hernández & Arancibia de Cabezas, 2007). Por ejemplo: (a) los maíces reventadores (incluye a los córneos y los córneo-harinosos) comúnmente son utilizados para la preparación de palomitas de maíz o pochoclo; (b) los maíces dentados son utilizados generalmente para preparar guisados; (c) los dextrinosos se utilizan en la elaboración de recetas dulces como la mazamorra, y (d) los harinosos son usados para la obtención de harina y la fabricación de bebidas alcohólicas fermentadas. Es posible que una discriminación a este nivel tenga una relevancia arqueológica significativa, dado que las inferencias sobre las prácticas culinarias, tales como los modos de preparación y consumo de alimentos, pueden basarse sobre este nivel de clasificación.
CONCLUSIONES
El análisis realizado nos permitió discutir la identificación arqueobotánica de las razas de maíz a través del estudio de sus almidones. El análisis estadístico dio cuenta de la alta variabilidad interna de las distintas razas de maíz actuales analizadas. Esto conlleva a la necesidad de ser muy cautos al clasificar especímenes aislados de almidones arqueológicos al nivel de raza.
Asimismo, proponemos que la baja probabilidad de realizar una correcta clasificación de las razas puede deberse en parte a la misma heterogeneidad de cariopsis en un mismo marlo. En nuestro estudio hemos hallado ejemplos representativos de cómo este fenómeno puede afectar la homogeneidad de una muestra. Esta cuestión puede tener raíz en el carácter reproductivo característico del maíz que consiste en la polinización abierta. Es decir que es muy frecuente que el polen de una raza fecunde una planta de otra raza, resultando en marlos con caracteres combinados (Bannert & Stamp, 2007; Bannert et al., 2008; Vogler et al., 2009). La cercanía geográfica de las parcelas de cultivo de maíz en el Noroeste argentino registrada etnográficamente hace que este fenómeno tenga recurrencia (Cámara Hernández & Arancibia de Cabezas, 2007). Como consecuencia, las razas no serían clases discretas y por este motivo debemos tener en cuenta la dificultad significativa de reconocimiento racial en muestras arqueológicas. Considerando que en el presente las muestras fueron tomadas de campos de cultivo de productores locales donde el control de la polinización es prácticamente nulo, no sería sorpresivo encontrarnos con contextos de cultivo similares en el pasado prehispánico. Una explicación de la alta variabilidad en las características de los almidones de maíz podría encontrarse precisamente en las prácticas tradicionales de manejo agrícola donde el conocimiento es la base de la biodiversidad. Esto se debe a una serie de prácticas agrícolas tales como los usos de huertos y rastrojos familiares, la complementariedad zonal de los cultivos, la adaptación de plantas a diversas ecozonas, el intercambio de semillas a nivel comunitario y la interacción entre las variedades cultivadas (Hilgert & Gil, 2005; Pochettino & Lema, 2008; Lema, 2012).
En síntesis, la caracterización de una amplia muestra de almidones de razas del Noroeste argentino permitió conocer y cuantificar la variabilidad interna de cada raza y estimar la probabilidad de una correcta clasificación racial. Los resultados indicaron que esta probabilidad es relativamente alta solo para los maíces harinosos y dextrinosos, mientras que para los córneos, córneo-harinosos y dentados disminuyó notablemente, constituyendo un límite para la identificación. A futuro se ofrece la posibilidad de que los gránulos de almidón arqueológicos sean incluidos dentro de la base de referencia, utilizando los mismos criterios de clasificación y las mismas variables implementadas en este trabajo, para luego comprobar dentro de qué grupo pronosticado se posicionan y estimar el error de dicha identificación. Consideramos que nuestro trabajo constituye un aporte para la identificación de restos microscópicos arqueobotánicos ya que permite una construcción más robusta de los datos.
AGRADECIMIENTOS
Este trabajo fue realizado con el apoyo de la Agencia Nacional de Promoción Científica y Tecnológica (PICT-2010-1332, director Marco Giovannetti) y de la Universidad de Buenos Aires (UBACYT F-0357, directora Norma Ratto). Irene Lantos contó con una Beca Doctoral del Consejo Nacional de Investigación Científica y Tecnológica. Marco Giovannetti es Investigador del Consejo Nacional e Investigación Científica y Tecnológica. Agradecemos a Raquel Defacio (Banco de Germoplasma, INTA-Pergamino) por proveernos con los especímenes de maíces nativos utilizados en este estudio, a Científica Central por colaborar con los insumos de laboratorio y a María L. Pochettino por sus inestimables consejos. Los evaluadores y editores han hecho valiosos aportes a este trabajo.
BIBLIOGRAFÍA
1. Abiusso, N. & J. A. Cámara Hernández. 1974. Los maíces autóctonos de la Quebrada de Humahuaca (Jujuy, Argentina), sus niveles nitrogenados y su composición en aminoácidos. Revista de la Facultad de Agronomía 3: 1-25. [ Links ]
2. Babot, M. P. 2005. Plant resource processing by Argentinian Puna hunter-gatherers (ca. 7000-3200 B.P): microfossil record. Phytolitharien: Bulletin of the Society for Phytolith Research 17: 9-10. [ Links ]
3. Babot, M. P. 2007. Granos de almidón en contextos arqueológicos: posibilidades y perspectivas a partir de casos de Noroeste argentino, en M. B. Marconetto, M. P. Babot & N. Oliszewski (eds.), Paleoetnobotánica Del Cono Sur: Estudios de Casos y Propuestas Metodológicas, pp. 95-125. Córdoba: Ferreyra Editor. [ Links ]
4. Babot, M. P. 2009. La cocina, el taller y el ritual: explorando trayectorias del procesamiento vegetal en el noroeste argentino. Darwiniana 47: 7-30. [ Links ]
5. Babot, M. P. & M. C. Apella. 2003. Maize and Bone: Residues of Grinding in Northwestern Argentina. Archaeometry 45: 121-132. DOI: http://dx.doi.org/10.1111/1475-4754.00099 [ Links ]
6. Babot, M. P.; N. Oliszewski & A. Grau. 2007. Análisis de caracteres macroscópicos y microscópicos de Phaseolus vulgaris (Fabaceae, Faboideae) silvestres y cultivados del noroeste argetnino: una aplicación arqueobotánica. Darwiniana 45: 149-162. [ Links ]
7. Bannert, M. & P. Stamp. 2007. Cross-pollination of maize at long distance. European Journal of Agronomy 27: 44-51. DOI: http://dx.doi.org/10.1016/j.eja.2007.01.002 [ Links ]
8. Bannert, M.; A. Vogler & P. Stamp. 2008. Short-distance cross-pollination of maize in a small-feld landscape as monitored by grain color markers. European Journal of Agronomy 29: 29-32. DOI: http://dx.doi.org/10.1016/j.eja.2008.02.002 [ Links ]
9. Blalock, H. M. 1996. Estadística social. México D.F: Fondo de cultura económica. [ Links ]
10. Boyd, M.; T. Varney, C. Surette & J. Surette. 2008. Reassessing the northern limit of maize consumption in North America: stable isotope, plant microfossil, and trace element content of carbonized food residue. Journal of Archaeological Science 35: 2545-2556. DOI: http://dx.doi.org/10.1016/j.jas.2008.04.008 [ Links ]
11. Bracco, M.; V. V. Lia, J. C. Hernández, L. Poggio & A. M. Gottlieb. 2012. Genetic diversity of maize landraces from lowland and highland agro-ecosystems of Southern South America: implications for the conservation of native resources. Annals of Applied Biology 160: 308-321. DOI: http://dx.doi.org/10.1111/j.1744-7348.2012.00544.x [ Links ]
12. Cámara Hernández, J. A. & D. Arancibia de Cabezas. 2007. Maíces andinos y sus usos en la Quebrada de Humahuaca y regiones vecinas (Argentina). Buenos Aires: Facultad de Agronomía, Universidad de Buenos Aires. [ Links ]
13. Coil, J.; M. A. Korstanje, S. Archer & C. A. Hastorf. 2003. Laboratory goals and considerations for multiple microfossil extraction in archaeology. Journal of Archaeological Science 30: 991-1008. DOI: http://dx.doi.org/10.1016/S0305-4403(02)00285-6 [ Links ]
14. Cortella, A. R. & M. L. Pochettino. 1994. Starch grain analysis as a Microscopic Diagnostic Feature in the Identifcation of Plant Material. Economic Botany 48: 171-181. DOI: http://dx.doi.org/10.1007/BF02908212 [ Links ]
15. Dickau, R.; A. J. Ranere & R. G. Cooke. 2007. Starch grain evidence for the preceramic dispersals of maize and root crops into tropical dry and humid forests of Panama. Proceedings of the National Academy of Sciences 104: 3651-3656. DOI: http://dx.doi.org/10.1073/pnas.0611605104 [ Links ]
16. Drennan, R. D. 1996. Statistics for archaeologists. A Commonsense Approach. New York: Plenum Press. DOI: http://dx.doi.org/10.1007/978-1-4899-0165-1 [ Links ]
17. Ferrer, M. E.; L. Solari & M. J. Hourquescos. 1998. Maíz: Los Recursos fitogenéticos en Argentina. Pergamino: EEA Pergamino. [ Links ]
18. Galinat, W. C. 1988. The Origin of Maiz de Ocho. American Anthropologist 90: 682-683. DOI: http://dx.doi.org/10.1525/aa.1988.90.3.02a00110 [ Links ]
19. Giovannetti, M. A. 2009. Articulación entre el sistema agrícola, redes de interacción y áreas de molienda como medida del grado de ocupación Inka en El Shincal y Los Colorados (Prov. de Catamarca). Tesis Doctoral, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. [ Links ]
20. Giovannetti, M. A.; I. Lantos, R. Defacio & N. Ratto. 2012. Construcción de un banco de almidones de variedades nativas de Zea mays L. del noroeste argentino. Propuesta metodológica y primeros resultados, en M. P. Babot, M. Marschoff &F. Pazzarelli (eds.), Las manos en la masa. Arqueologías, antropologías e historias de la alimentación en Suramérica, pp. 361-385. Córdoba: Corintios 13. [ Links ]
21. Giovannetti, M. A.; V. S. Lema, C. G. Bartoli & A. Capparelli. 2008. Starch grain characterization of Prosopis chilensis (Mol.) Stuntz and P. fexuosa DC, and the analysis of their archaeological remains in Andean South America. Journal of Archaeological Science 35: 2973-2985. DOI: http://dx.doi.org/10.1016/j.jas.2008.06.009 [ Links ]
22. Hammer, O.; D. A.T. Harper & P. D. Ryan. 2001. PAST: Paleontological statistics software package for education and data analysis. Palaeontológica Electrónica 4: 1-19. [ Links ]
23. Haslam, M. 2004. The decomposition of starch grains in soils: implications for archaeological residue analyses. Journal of Archaeological Science 31: 1715-1734. DOI: http://dx.doi.org/10.1016/j.jas.2004.05.006 [ Links ]
24. Hilgert, N. & G. Gil. 2005. Traditional andean agriculture and changing processes in the Zenta river basin, Salta, Northwestern Argentina. Darwiniana 43: 30-43. [ Links ]
25. Holst, I.; J. E. Moreno & D. R. Piperno. 2007. Identifcation of teosinte, maize, and Tripsacum in Mesoamerica by using pollen, starch grains, and phytoliths. Procedings of the National Academy of Sciences 104: 17608-17613. DOI: http://dx.doi.org/10.1073/pnas.0708736104 [ Links ]
26. IBM. 2010. Statistical Product and Service Solutions: SPSS Statistics 19.0, http://www.spss.com [ Links ]
27. Itlis, H. H. & J. Doebley. 1980. Taxonomy of Zea (Gramineae). II. Subspecifc categories in the Zea mays complex with a generic synopsis. American Journal of Botany 67: 994-1004. DOI: http://dx.doi.org/10.2307/2442442 [ Links ]
28. Korstanje, M. A. & P. Cuenya. 2010. Ancient agriculture and domestic activities: a contextual approach studying silica phytoliths and other microfossils in soils. Environmental Archaeology 15: 43-63. DOI: http://dx.doi.org/10.1179/146141010X12640787648739 [ Links ]
29. Lavalle, M. C. & A. Mengascini. 2007. Almidón de reserva en especies neotropicales de Marattia (Marattiaceae-Pteridoftas). Anales del Jardín Botánico de Madrid 64: 79-85. DOI: http://dx.doi.org/10.3989/ajbm.2007.v64.i1.56 [ Links ]
30. Lema, V. S. 2012. Nuevas ideas sobre la domesticación ¿nuevas ideas sobre el Formativo? Aportes para una relectura crítica. Pre-circulares del Taller de Arqueología Del Periodo Formativo, 12 al 15 de abril de 2012, Tafí del Valle. [ Links ]
31. Lentfer, C. 2009. Building a comparative starch reference collection for Indonesia and its application to palaeoenvironmental and archaeological research. Terra Australis 30: 80-101. [ Links ]
32. Lia, V. V.; V. A. Confalonieri, N. Ratto, J. A. Cámara Hernández, M. A. Miante Alzogaray, L. Poggio & T. A. Browns. 2007. Microsatellite typing of ancient maize: insights into the history of agriculture in southern South America. Proceedings of the Royal Society of Botany 274: 545-554. DOI: http://dx.doi.org/10.1098/rspb.2006.3747 [ Links ]
33. Lia, V.V.; L. Poggio, V. A. Confalonieri, N. Ratto, J. A. Cámara Hernández, A.M. Miante Alzogaray, L. Poggio & T. A. Browns. 2009. Microsatellite variation in maize landraces from Northwestern Argentina: genetic diversity, population structure and racial affiations. Theoretical and Applied Genetics 119: 1053-1067. DOI: http://dx.doi.org/10.1007/s00122-009-1108-0 [ Links ]
34. Loy, T.H. 1992. Methods in the analysis of starch residues on prehistoric stone tools. New York: Routledge. [ Links ]
35. McBird, R. 1994. Manual for the measurement of maize cobs, en S. Johannessen & C. A. Hastorf, (eds.), Corn and Culture in the Prehistoric New World, pp. 5-22. Boulder: Westview Press. [ Links ]
36. Miante Alzogaray, M. A. & J. A. Cámara Hernández. 1996. Restos arqueológicos de maíz (Zea mays ssp. mays) de Pampa Grande, Provincia de Salta, Argentina. Relaciones de la Sociedad Argentina de Antropología XXI: 149-159. [ Links ]
37. Molestina, C. J. 1987. Diálogo XVIII. Germoplasma de Maíz en el Cono Sur De América. Montevideo: IICA-BID-PRO-CISUR. [ Links ]
38. Moschetti, E.; S. Ferrero, G. Palacio & M. Ruiz. 2003. Introducción a la estadística para las ciencias de la vida. Río Cuarto: Universidad Nacional de Río Cuarto. [ Links ]
39. Muñóz Salas, E. 1998. La técnica de análisis discriminante: una aplicación para el área bancaria. San José: Banco Central de Costa Rica, División Económica, Departamento de Investigaciones Económicas. [ Links ]
40. National Institutes of Health. 2013. ImageJ 1.47, Image Processing and Analysis in Java. Bethesda: National Institutes of Health. [ Links ]
41. Oliszewski, N. 2008. Metodología para la identifcación subespecífca de maíces arqueológicos. Un caso de aplicación en el Noroeste de Argentina, en S. Archila, M. A. Giovannetti & V. S. Lema (eds.), Arqueobotánica y teoría arqueológica. Discusiones desde Suramérica, pp. 181-202. Bogotá: Uniandes-Ceso. [ Links ]
42. Oliszewski, N. & D. E. Olivera. 2009. Variabilidad racial de macrorrestos arqueológicos de Zea mays (Poaceae) y sus relaciones con el proceso agropastoril en la Puna Meridional Argentina (Antofagasta de la Sierra, Catamarca). Darwiniana 47: 76-91. [ Links ]
43. Pagán Jiménez, J. 2007. De antiguos pueblos y culturas botánicas en el Puerto Rico indígena. El archipiélago boricazo y la llegada de los primeros pobladores agroceramistas. Paris Monographs in American Archaeology. Oxford: BAR International Series. [ Links ]
44. Parodi, L. 1959. Enciclopedia argentina de agricultura y jardinería. Buenos Aires: ACME. [ Links ]
45. Pérez, C. 2001. Técnicas estadísticas con nultivariable con SPSS. Madrid: Prentice Hall. [ Links ]
46. Piperno, D. R. 2009. Identifying crop plants with phytoliths (and starch grains) in Central and South America: A review and an update of the evidence. Quaternary International 193: 146-159. DOI: http://dx.doi.org/10.1016/j.quaint.2007.11.011 [ Links ]
47. Piperno, D. R. & I. Holst. 1998. The presence of starch grains on prehistoric stone tools from the Humid Neotropics: Indications of early tuber use and agriculture in Panama. Journal of Archaeological Science 25: 765-776. DOI: http://dx.doi.org/10.1006/jasc.1997.0258. [ Links ]
48. Piperno, D. R., E. Weiss, I. Holst & D. Nadel. 2004. Processing of wild cereal grains in the Upper Palaeolithic revealed by starch grain analysis. Nature 430: 670-673. DOI: http://dx.doi.org/10.1038/nature02734 [ Links ]
49. Pochettino, M. L. & V. S. Lema. 2008. La variable tiempo en la caracterización del conocimiento botánico tradicional. Darwiniana 46: 227-239. [ Links ]
50. Portillo, M.T. & J. A. Salas. 2008. P. CH. Mahalanobis y las aplicaciones de su distancia estadística. Cultura Científca y Tecnológica, Universidad Autónoma de Ciudad Juárez 5:13. [ Links ]
51. Reichert, E. T. 1913. The differentiation and specifcity of starches in relation to genera, species, etc. Washington DC: Carnegie Institution. [ Links ]
52. Rodríguez, M. F. & C.A. Aschero. 2007. Archaeological Evidence of Zea mays L. (Poaceae) in the Southern Argentinean Puna (Antofagasta de la Sierra, Catamarca). Journal of Ethnobiology 27: 256-271. DOI: http://dx.doi.org/10.2993/0278-0771(2007)27[256:AEOZML]2.0.CO;2 [ Links ]
53. Shennan, S. 1992. Arqueología cuantitativa. Madrid: Crítica. [ Links ]
54. Solari, L. R. & S. G. Gómez. 1997. Catálogo de Germoplasma de Maíz. Argentina. Firenze: Instituto Agronomico per L'Oltremare. [ Links ]
55. Staller, J. E.; R. Tykot & B. Benz. 2006. Histories of maize: multidisciplinary approaches to the prehistory, linguistics, biogeography, domesticacion, and evolution of maize. Walnut Creek: Left Coast Press. [ Links ]
56. Torrence, R. & H. Barton (Eds.). 2006. Ancient Starch Research. Walnut Creek: Left Coast Press. [ Links ]
57. Torrence, R.; R. Wright & R. Conway. 2004. Identifcation of starch granules using image analysis and multivariate techniques. Journal of Archaeological Science 31: 519-532. DOI: http://dx.doi.org/10.1016/j.jas.2003.09.014 [ Links ]
58. Vogler, A.; H. Eisenbeiss, I. Aulinger-Leipner & P. Stamp. 2009. Impact of topography on cross-pollination in maize (Zea mays L.). European Journal of Agronomy 31: 99-102. DOI: http://dx.doi.org/10.1016/j.eja.2009.04.003 [ Links ]

