










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkDarwiniana, nueva serie
versión impresa ISSN 0011-6793versión On-line ISSN 1850-1699
Darwiniana, nueva serie vol.2 no.1 San Isidro jun. 2014
ARTÍCULO
Microfungal Composition in an Astelia-Donatia Cushion Peatland in Tierra Del Fuego, Argentina
Noelia I. Paredes*1, 2, Verónica F. Consolo*1, Verónica A. Pancotto2,6, Christian Fritz3,7, Marcelo D. Barrera4, Angélica M. Arambarri5 & Graciela L. Salerno1
1 Instituto de Investigaciones en Biodiversidad y Biotecnología (CONICET), Fundación para Investigaciones Biológicas Aplicadas (FIBA), Vieytes 3103, 7600 Mar del Plata, Buenos Aires, Argentina; faconsolo@fba.org.ar (author for correspondence).
2 Centro Austral de Investigaciones Científcas (CONICET), Houssay 200, 9410 Ushuaia, Tierra del Fuego, Argentina.
3 Experimental Plant Ecology, Radboud University, Heyendaalseweg 135, 6525AJ, Nijmegen, Netherlands.
4 Laboratorio de Investigación en Sistemas Ecológicos y Ambientales, Universidad Nacional de La Plata, Diagonal 113 Nº 469, 1900 La Plata, Buenos Aires, Argentina.
5 Instituto de Botánica Carlos Spegazzini, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Calle 53 Nº 477, 1900 La Plata, Argentina.
6 Cátedra de Botánica, Universidad de Tierra del Fuego, Onas 450, 9410 Ushuaia, Tierra del Fuego, Argentina.
7Aquatic Ecology, Radboud University, Heyendaalseweg 135, 6525AJ, Nijmegen, Netherlands
*Both authors contributed equally to this work
Editor Asociado: Fernando Biganzoli
Original recibido el 21 de diciembre de 2013,
aceptado el 28 de mayo de 2014
Abstract
Southern Hemisphere peatlands store substantial amounts of soil carbon. Despite their importance in the global carbon cycle, little is known about decomposition processes and the associated fungal diversity. The present study describes the composition of fungal assemblage in two depths from a cushion peatland of predominating Astelia (Asteliaceae) and Donatia (Donatiaceae) species in Moat, Tierra del Fuego. From 48 samples processed, we obtained 338 isolates. Using different culturing methodologies, through direct and microscopic observation and using molecular methods we identified 38 fungal species and 18 genera of Ascomycetes and Zygomycetes. Isolates belonging to Ascomycetes were the most abundant, with dominance of Penicillium and Trichoderma. We described fungal composition and compared species diversity and evenness across two dephts. No differences in the diversity index and evenness were found between depths. The studied peat is an ecosystem that has a great diversity of fila-mentous fungal species, some of which are described in other peatlands worldwide.
Key words: Astelia-Donatia; Diversity; Fungal community; PCR- RFLP; Peatland
Resumen
Composición de microhongos en una turbera compacta de Astelia-Donatia en Tierra del Fuego, Argentina. En el Hemisferio Sur las turberas almacenan grandes cantidades de carbono en el suelo. A pesar de su importancia en el ciclo global del carbono, se sabe poco acerca de los procesos de descomposición y su diversidad fúngica. El presente estudio se realizó para describir la composición de hongos filamentosos en dos profundidades de una turbera compacta donde las especies vegetales predominantes son Astelia (Asteliaceae) y Donatia (Donatiaceae) en Moat, Tierra del Fuego, Argentina. De 48 muestras procesadas, se obtuvieron 338 aislamientos. Mediante el uso de diferentes metodologías de cultivo, a través de observación microscópica y por métodos moleculares identificamos 38 especies fúngicas y 18 géneros de Ascomycetes y Zygomycetes. Los aislamientos de Ascomycetes fueron los más abundantes, con dominancia de Penicillium y Trichoderma. Se describe la composición fúngica y se comparó la diversidad y equitatividad de las especies a dos profundidades de muestreo. No se encontraron diferencias significativas entre la composición de especies, la diversidad y equitatividad. La turbera estudiada es un ecosistema que presenta alta diversidad de especies fúngicas filamentosas, algunas de ellas descritas en otras turberas en todo el mundo.
Palabras clave: Astelia-Donatia; Comunidad fúngica; Diversidad; PCR- RFLP; Turbera
INTRODUCTION
Peatlands are wetlands with a permanently high water level which accumulate organic matter resulting from incomplete decomposition of plant material (Koesnandar et al., 2006). These environments represent the major important store of terrestrial carbon due to a long-term imbalance between rates of net primary production by plants and microbial decomposition (Clymo, 1984). Primary production rates in peatlands are generally slow (Frokling et al., 1998) and decomposition rates are even slower due to environmental factors, including cold temperatures, partial anoxia, soil chemical and microbiological factors (Moore & Basiliko, 2006). Although peatlands sequester organic matter into the anoxic layer, most of the carbon flux takes place in the oxic layer above the water level (Jaatinen et al., 2007).
The microflora in peatlands consists of prokaryotes (bacteria and Archaea) and fungi (Kamal & Varma, 2008). These microbial decomposers are of particular interest, because they are involved in decomposition of plant tissues, producing both peat accumulation and the release of CO2 and CH4 (Moore & Basiliko, 2006). All of these organisms play important roles in carbon cycling, interact with plants via exchange of organic and inorganic compounds and they can also alter the physicochemical environment of the interacting organisms (Kamal & Varma, 2008). Several reports had showed that microbial biomass decreases with depth, explained by the presence of more labile organic matter and higher redox states near the surface of peat profiles (Blodau et al., 2004; Basiliko et al., 2007). Despite their importance in the global carbon cycle, microbial communities in these sites are still poorly understood (Peltoniemi et al., 2009; Lin et al., 2012). The most studied microorganisms in peatlands are bacteria, specially methanotrophs (Chen et al., 2008; Kip et al., 2012) and methanogenic archaea (Basiliko et al., 2003; Sizova et al., 2003).
It has long been suggested that heterotrophic fungi are the dominant microbes and principal decomposer organisms in many acidic ecosystems, such as peatlands, which in oxic conditions are considered to be the main agents of litter decomposition (Killham, 1994; Andersen et al., 2006). Fungi produce a wide range of enzymes that degrade complex polymers such as starch, cellulose, proteins, chitin, keratin, and even the most complex lignified materials such as wood.
Although there are reports from fungal communities, to date peatlands have not been extensively investigated. Studies including cultivation of fungal isolates or direct identification by microscopy were done several decades ago (Dooley & Dickinson, 1970). Other studies were focused on particular plant species or decomposition of fresh plant litter (Thormann et al., 2003, 2004; Thormann, 2006). Differences in microbial community structure have been identified between peatland types (eg. bogs and fens) as well as among depths down the peat profile (Jaatinen et al., 2007; Peltoniemi et al., 2009; Preston et al., 2012). Different approaches on fungal studies based on molecular methods or phylogenetic information obtained from 18S rRNA gene or ITS sequences have been focused on the classification and community ecology of mycorrhizal fungi (Horton & Bruns, 2001; Anderson & Cairney, 2004). Also, the use of denaturing gradient gel electrophoresis (DGGE) and clone libraries of fragments of the fungal rRNA internal transcribed spacer (ITS) region allowed the characterization of the fungal communities across a gradient of naturally regenerating peatland (Artz et al., 2007).
The last reviews on fungal diversity in a variety of peatlands in Europe, North and South America (Thormann, 2006; Thormann & Rice, 2007) report 601 species of fungi identified globally, being Ascomycetes the largest group with 276 species (46%), followed by Basidiomycetes (243 species, 40%), Zygomycetes (55 species, 9%), and Chytridiomycetes (26 species, 4%). In Argentina, peatlands are only found in the southern region of the country (Patagonia), and they are predominant in the south-western region, where they are widely distributed (Malvárez et al., 2004). Peatlands of Tierra del Fuego occur where the annual rainfall rises from 400mm, and are widespread through the deciduous and evergreen forest zone (Moore, 1983; Pisano, 1983). Most of raised bogs in Tierra del Fuego only have Sphagnum species, predominantly S. magellanicum (Bridel), which forms hummocks, hollows and all the other complex structures that are typical of bogs (Baumann, 2006). The uninhabited Eastern part, known as Península Mitre, is exposed to hard oceanic conditions; in this area mires are particularly frequent and cover a total area of approximately 2,400 km2. The forest and mires are dominated by Nothofagus betuloides (Mirb.) Oerst. and cushion plants (e.g. species of Astelia). There are numerous studies describing expeditions and fungal diversity from different sites in Tierra del Fuego, for example, those associated to Gunnera magellanica Lam. (Pancotto et al., 2003) or Nothofagus (Gamundí & Amos, 2007; Godeas, 1983; Godeas & Arambarri, 2007). Other reports have shown the influence of solar UV-B on the fungal communities in Sphagnum peatlands (Searles et al., 2001; Robson et al., 2004). However, there are no descriptions of the fungal composition on cushion peatlands of Tierra del Fuego composed by Astelia pumila (G. Forst.) Gaudich. (Asteliaceae) and Donatia fascicularis J.R. Forst. & G. Forst. (Donatiaceae).
The objective of this work is to describe depht-dependent microfungal community composition in a pristine bog located at the southeastern region of Tierra del Fuego (Moat) using different culture isolation methodologies and assisted by microscopic observation or molecular techniques.
MATERIAL AND METHODS
Site description
Collection of samples was carried out during the austral summer 2010 in Moat Valley, Isla Grande, Tierra del Fuego, Argentina. This area is part of the Subantartic phytogeographic Province, District Magellan, where characteristic vegetation consists of steppes, forests and peatlands (Moore, 1983). The study site (Moat Valley, 54° 58' 32.6'' S, 66° 44' 04.1'' W, 40 m a. s. l.) is a pristine cushion peatland near the southeastern coast of Tierra del Fuego at Bahia Moat, North of the Beagle Channel, 100 km East of Ushuaia (Fig. 1). The Moat Valley peatland is exposed to the sea, without protecting mountains, where precipitation conditions consist of 500-600 mm of rain and more than 150 mm of snow per year (Fritz, 2012), with average annual temperatures close to 6° C and low amplitude due to strong oceanic influence (Roig & Collado, 2004). This mire landscape in Moat Valley is characterized by abundant peat-forming cushion plants such as Astelia pumila, which is dominant and forms rigid ‘blankets' or sometimes real hummocks that gives the mire a green appearance, and Donatia fascicularis, which is less frequent (Gebser, 2008). The cushion plants are found in this bog system and form dense root systems consisting of shallow tap and aerenchymatous roots of > 100 cm in length (Fritz et al., 2011), although they are also found on blanket bogs on the hills close to Moat, where the peat layer is approximately 0.6 to 3 m thick.

Fig. 1. Fotomicrografías al MEB de antecios superiores de especies de Paspalum grupo Gardneriana. P. delicatum de Gutiérrez et al. 1292 (SI). A, vista dorsal de la lema. B, detalle de las papilas verrugosas de la lema. P. gardnerianum de Hassler 11904 (SI). C, detalle de las papilas verrugosas de la lema. P. pictum de Chase 11795 (US). D, vista dorsal de la lema. E, detalle de las papilas verrugosas de la lema. P. subsesquiglume de Swallen 4329½ (US). F, detalle de las papilas verrugosas de la lema.
Low nutrient concentrations (below 15 mg N/g and 0.3 mg P/g of peat dry weight) have been reported in the 2-m uppermost peat substrate (Fritz et al., 2011). The peatland sampled in this study preserved its pristine character remaining unaffected by anthropogenic alteration such as drainage, agriculture or elevated atmospheric nutrient deposition.
Sample collection and processing
Sample collection were done in triplicate peat cores along three parallel transects randomly traced. Along each transect, 8 sub-sampling points were placed 1 m apart and intact soil cores (20 cm depth, 12 cm diameter) were taken using a steel soil corer, knives and saws, due to the characteristic compacted peat formed by Astelia sp.
Samples were placed separately in sterile plastic zipper bags and they were immediately stored at 4 °C in the laboratory. Two days after collection, they were transported to the laboratories of IN-BIOTEC (Mar del Plata, Buenos Aires Province). Processing the 24 sub-samples included removal of the first 3 cm (usually containing live tissue of Astelia, Donatia or other plant species) and the remaining peat block was divided according two depths: (A) 3-10 cm and (B) 11-20 cm. Stones and macroscopic dead plants were removed.
Soil pH, temperature and distance from deep water were determined for each sample. Average values of pH and temperature of the soil were 4.23 and 12.4 ºC respectively. Water levels were automatically recorded (Odyssey capacitance probes ®, Dataflow). During the sampling period we observed a 3-week period with water level 10 cm below the surface (data not shown). Peat sampled showed a high degree of decomposition while oxygen loss by roots may stimulate oxidation processes down processes below the upper 200 cm of the peat profile (Fritz et al., 2011).
Microfungal isolation
A combination of isolation methods was applied in order to obtain more information about the composition of peat fungal communities. Each sub-sample was homogenized and one gram of wet soil was processed with each of the three isolation techniques: direct sowing soil, soil diluted methodology and moist chamber incubation. Fungal growth was macro- and microscopically observed under stereoscope on culture plates and fungal structures were examined by optical microscope (40×).
Direct sowing soil
Each sub-sample was divided into five small portions and placed in Petri dishes containing solid culture media Potato Dextrose Agar (PDA: 200 g of potato infusion, 20 g of dextrose, 15 g of agar and 1L of distilled water) or Czapeck Dox (CD: 1 g of KH2PO4, 30 g of sucrose, 2 g of NaNO3, 1 g of K2HPO4, 0.5 g of MgSO4, 0.5 g of KCl, 0.01g of FeSO4, 15 g of agar, and 1 L of distilled water) with the addition of 1 g/L of chloramphenicol and adjusted to pH 5. A total of 240 particles, [5 particles of each sub-samples at depth A (24) and depth B (24)] were seeded in 48 Petri dishes with PDA and 240 particles in 48 Petri dishes with CD. The plates were incubated at room temperature (18±2 ºC), since colonies developed faster than 10±2 ºC (temperature of collected samples), until the appearance of fungal colonies. The number of colonies per species was registered by Petri dish.
Diluted soil
Each sub-sample were placed in a test tube with 10 ml of sterile distilled water. The suspension was stirred with a Vortex mixer for 1 min and two dilutions (1:100 and 1:1000) were done. A 600 μl aliquot of 10-3 dilution was seeded on Petri dishes on PDA and CD media. A total of 96 dishes were seeded and incubated at room temperature (18±2 ºC), until the appearance of fungal colonies.
Moist incubation chamber
Moist chambers (24 for each sub-sample set) were prepared using a sterile plate containing filter paper, two plastic rods and a slide with the corresponding sample of peat. They were regularly moistened with sterile water and maintained at room temperature until the appearance of fungal colonies. To isolate fungal species, a portion of fungal mycelium or spores was taken and seeded in a dish with PDA medium.
Identification of peat microfungi
Pure cultures of isolated fungi were characterized on the basis of their micro- and macromorphology according to previous reports (Ellis, 1971, 1976; Carmichael et al., 1980; Domsch et al., 1980; Ramírez & Martínez, 1982). Some isolates of Trichoderma together with a group of sterile mycelia were identified with molecular methodologies.
DNA extraction and PCR analysis
Each isolate was grown for 5 days at room temperature in 50 ml of PDA medium under orbital shaking. Mycelia were harvested under vacuum and lyophilized. About 100 mg of powered mycelia were used for DNA extraction following a modified versión of the cetyltrimethylammonium bromide (CTAB) method (Murray & Thompson, 1980). DNA quality was determined by electrophoresis on 1% agarose gel. PCR amplification of fungal the ITS región (including ITS1 and ITS2, and 5.8S gene of rDNA) was carried out with primers ITS5 (5'-GGAAGTAAAAGTCGTAACAAGG-3') and ITS4 (5'-TCCTCCGCTTATTGATATGC-3'; White et al., 1990). PCR was conducted in a 25 μL final volume with 10 ng of témplate DNA, 20 mM Tris- HC1 (pH 8.4), 50 mM KC1, 1.5 mM MgCL2, 0.2 mM of dNTPs, 0.5 μM of each primer, and 1 unit of Taq polymerase (Invitrogen, Carlsbad, CA, USA). Thermal cycling conditions involved an initial denaturation step at 94° C for 2.5 min, followed by 30 cycles of 94° C for 15 s, 48° C for 1 min and 72 °C for 1.5 min, and a final extensión at 72 °C for 10 min PCR producís were separated by electrophoresis in a gel containing 1% agarose (Invitrogen Carlsbad, CA, USA) in TAE buffer (40 mM Tris-HCl, pH 8, and 2 mM EDTA). Gels were run for 1 h at 120 V, stained with ethidium bromide and photographed with a Fotodyne system (Hartland, WI, USA).
PCR-RFLP analysis
Amplification producís of ITS regions from 40 Mycelia sterilia were analyzed through PCR-RFLP (PCR-restriction fragment length polymorphism). The restriction reactions were performed in a 20 μL final volume using 10 μl of PCR amplification producís and 1 U of Haelll or Hinfl in 2 μl of appropriate buffer (Promega). After incubation for 1 h at 37° C, the digestión producís were run out on 2% agarose gels.
DNA sequencing and data analysis
DNA sequencing and PCR amplificalion producís were purified and sequenced by Macrogen Inc. (Soulh Korea) wilh íhe ITS4 primer. Sequences were idenlified using íhe BLAST program (hllp://blasl.ncbi.nlm.nih.gov), and deposiled in the European Molecular Biology Laboralory (EMBL) dalabase.
Conservation of fungal isolates
One representative of each species of sporulating fungi was stored on filter paper at -20 °C (Nakasone et al., 2004). In the case of species of Mycelia sterilia, they were preserved in Petri dishes containing PDA culture media.
Community description and analysis
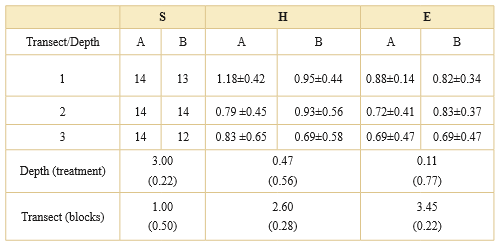
Constancy of species was determined as (number of the sub-samples in which a species was recorded/total number of sub-samples) × 100. Community of fungal species across two depths was analyzed by (i) species richness (S), (ii) Shannon's diversity index, (iii) evenness E = H´/H´max, where H´max is the maximum value of the diversity for the number of species that are present (Magurran, 1988). Species richness, means of Shannon´s diversity Index (H) and evenness (E) between depths were examined with one-way blocked ANOVA, were transects were blocks and depth treatment.
RESULTS
Isolation and identification of fungal microfungi
Using different culturing methodologies, through direct and microscopic observation and assisted by molecular methods we identified 38 fungal species and 18 genus comprising ana-morph genera of Ascomycetes and Zygomycetes.
Eighteen genera were identified as Acremonium, Alternaria, Aphanocladium, Aspergillus, Beauveria, Cladosporium, Giberella, Metacordyceps, Mortierella, Mucor, Paecilomyces, Penicillium, Phoma, Physcia, Scopulariopsis, Trichoderma, Ulocladium, and Verticillium. Penicillium and Trichoderma were the most representative genera (Tables 1 and 2).
Table 1. Taxonomic summary and number of isolates of fungi from Astelia-Donatia cushion peatland, Moat (Argentina).
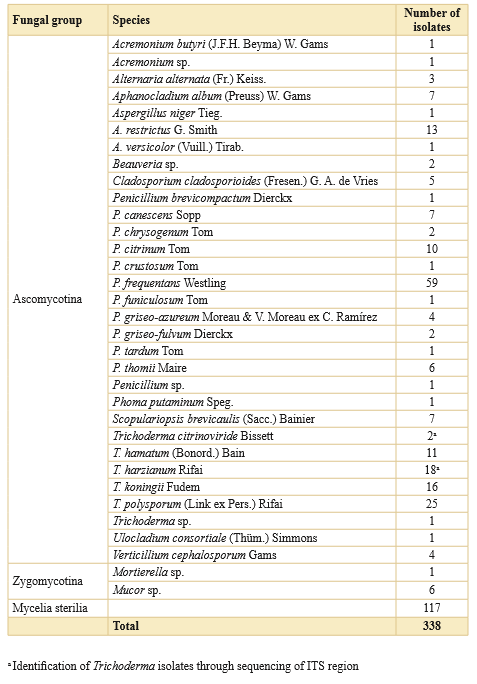
Table 2. Representative members of each of the fve PCR-RFLP groups. Identity was corroborated by comparing the ITS region of fungal species in the BLAST program of NCBI database with a similarity percentage greater than 90%. ITS sequences of fungal isolates were deposited on EMBL.
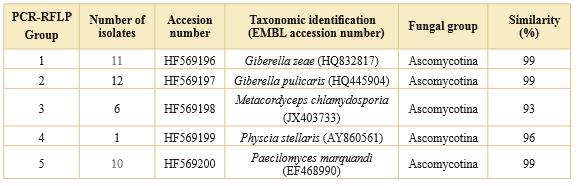
From three different isolation methodologies, direct sowing was the most efficient technique, since 299 isolates were obtained. From these isolates, 182 were identified at species or genus level according to the presence of characteristic reproductive structures and 117 isolates could not be classified because they presented only a hyaline sterile mycelium. From these results, a group of 40 Mycelia sterilia colonies, which presented macroscopic differences in colony topography, color, texture or exudates production, were selected for identification through PCR-RFLP analysis of the ITS region. Five restriction patterns were obtained from all colonies analyzed. The amplification product of the ITS region from one representative isolate of each PCR-RFLP group was sequenced. Five fungal species were identified different to the sporulating fungi taxonomically determined through microscopic analysis (Table 2).
Using the dilution method of soil, a total of 39 colonies were isolated and identified as Trichoderma. Nineteen isolates could be directly identified by microscopy and a group of twenty isolates, were identified mostly as T. harzianum after sequencing the ITS region (Table 1).
Finally, using the humid chamber method, a predominant development of opportunistic saprophytic fungi (mainly yeasts) were only observed and consequently, no fungal isolation was considered.
Community structure
Species richness, Shannon´s diversity and evenness did not show a significant difference among dephts (Table 3). Species richness was similar between depths with 12 to 14 species by transect. Mean of Shannon´s diversity index ranged from 0.69 to 1.18 and evenness from 0.69 to 0.88.
Table 3. Total number of microfungal species (S), mean and standard deviation of Shannon´s diversity Index (H), and evenness (E) for each transect, at two sampling depths (A: 3-10 cm depth; B: 11-20 cm depth) in Astelia-Donatia cushion peatland, Moat, Tierra del Fuego. F and p-value (between brackets) from one-way blocked ANOVA for depth (treatment) and transect (blocks) are shown for each index. There were no signifcant differences between depths (p > 0.05).
Fungal species found in sub-samples A and B and constancy values (%) are shown in Table 4. We observed exclusive species in depht A (8) or in B (10) while 14 species were found in both sub-samples. Fungal isolates considered as Mycelia sterilia presented a high constancy (79.1%) meanwhile microfungal identified species showed that P. frequentans (54.1%), T. harzianum (33.3%), T. koningii (29.1%) T. polysporum (22.9%) T. hamatum (20.8%), and A. restrictus (16.6%) were the taxa with a greater constancy.
Table 4. Fungal species identifed at different sampling depths (A: 3-10 cm depth; B: 11-20 cm depth) and constancy (%) from Astelia-Donatia cushion peatland.

DISCUSSION
This study is the first to analyze microfungal diversity in a southernmost pristine peatland of Tierra del Fuego by using a combination of classic and molecular methods for fungal identification. In the studied peatland the anamorphic Ascomycetes represents the largest group of fungi isolated as in other environments in the world. Additionally, fungi of the subdivision Zygomycetes and an important group of unidentified fungi represented by Mycelia sterilia were isolated. These results are in accordance with previous studies describing fungal diversity found in surveys conducted in Europe, North and South Hemisphere peatlands (Thormann, 2005; Gilbert & Mitchell, 2006; Thormann & Rice, 2007).
Fungi of genera Aspergillus, Cladosporium, Mortierella, Mucor, Ulocladium, Penicillium and Trichoderma were identified, and interestingly, they correspond to fast growing species with copious spore production. The high number and proportion of these fungal species on peat is a consequence of the acidic conditions, poor aeration and low content of mineral in these environments. Furthermore, these fungi have been reported representing the vast majority of isolates obtained from worldwide peat samples. Their growth habits and relative easiness of cultivation on standard media could explain this prevalence (Thormann, 2005; Artz et al., 2007; Thormann & Rice, 2007). The mentioned ñingal species, were also reported in fungal biodiversity studies associated with Sphagnum bogs at Tierra del Fuego (Searles et al., 2001; Robson et al., 2004).
Species of Aspergillus, Penicillium and Trichoderma were the most common found in the present study (Tables 1 and 4). These genera have broad enzymatic profile and, consequently, are able to degrade a wide variety of structural and storage polymers of peat and peatland plants, where they are dominant (Thormann, 2005). Particularly, Penicillium species were the most numerous at the studied site, being P. frequentans the species with the highest constancy (54.1%) (Table 4), probably due to their ability to produce a wide range of antibiotics and mycotoxins protecting them from other soil organisms, and hindering the growth of other fungal species. These results are similar to studies conducted by Khalid et al. (2006) in which they reported the high percentage of ocurrence of this genus as cellulose decomposing fungi in soils samples. Moreover, it has been shown that Penicillium is the dominant genus on peat samples, and is more frequent with increasing acidity of peats (Küster, 1963; Repečkiené et al., 2012).
Soil pH has been identified as a highly influential factor controlling microbial diversity and community composition across soil types (Fierer & Jackson, 2006; Allison & Treseder, 2008; Rousk et al., 2010) and differences in microbial communities structures have been identified between peatland types. The pH of our studied site averaged 4.23, which represents conducive environmental conditions for fungal development.
Trichoderma was another important genus found at the Moat cushion peatland. This is a soil-borne free-living, non-pathogenic fungus, which can also colonize roots of numerous plants, has a broad enzymatic profile and often prevenís the development of other fungi. In this study we identified five Trichoderma species, three of them through direct microscopic observation and two additional ones by a molecular approach (Table 1). All these species have been reported from peatlands in the Northern Hemisphere (Thormann & Rice, 2007).
PCR-RFLP was an additional and useful technique that allowed us to identify other fungal species from 40 Mycelia sterilia isolates. Thus, five different groups of fungal taxa were determined (Table 2). As reported in other studies, this method can be of great utility when is not possible to use the classical methods for identification or may be used as an efficient culture indepedent-molecular tool for ecological studies since more information about fungal diversity could be obtained (Viaud et al., 2000; Watrud, 2006).
Although species richness, diversity or eveness did not show a significant statistical difference between depths (Table 3), different fungal species was exclusively isolated from peat samples taken on the surface or in depht (Table 4). Also there were shared species isolated from two depths as Aspergillus, Cladosporium, Penicillium and Trichoderma corresponding to dominant fungal taxa with higher constancy (Table 4). These fungi are ubiquitous and in this study their isolation were not affected by depth Most common species of Penicillium and Trichoderma genera were present in both depths although certain species of these fungi with low constancy (2.0%) were present only in one depth (Table 4). As reported in other studies about richness and diversity of fungal species on different peatland, cellulose-decomposing fungi were the most representative group and their wide distribution, fast growing development and sopurulation suppressed the growth of other fungal species (Thormann, 2005; Repečkiené et al., 2012).
Other species with low constancy as Aphanocladium (4.1%), Scopulariopsis (4.1%) and Verticillium (4.1%) occurred exclusively at surface samples, whereas those of the genera Mortierella (2.0%) and Ulocladium (2.0%) were only identified at depth samples. In this study, as reported in other peat-lands the occurrence rate of these fungal taxa was rare or occasional (Repečkiené et al., 2012).
In contrast with several reports that showed a decrease of microbial biomass with depth, associated by the presence of more labile organic matter or higher redox states near the surface (Blodau et al., 2004; Basiliko et al., 2007), it seems that in the case of the site sampled at the Moat peatland, fungal species diversity were not affected by depth. Maybe similar organic matter quality (Paredes, pers. com.) or a similar relationship between oxygen and carbon dioxide levels (Giri et al., 2005; Bragazza et al., 2007; Preston et al., 2012) could contribute to this fungal distribution. Additionally, it was reported that fungal activity often decreases under anoxic conditions when anaerobic bacteria are the major players in the mineralization of organic matter via fermentation and respiration processes (Kamal & Varma, 2008; Thormann, 2011; Lin et al., 2012). In this study we recorded that the water table was close to the surface during 95% of the summer season, and samples at depth were taken from a partial water-logged condition. On the other hand, anoxia could be prevented by oxygen releasing from roots (Fritz et al., 2011) of cushion plants like A. pumila and D. fascicularis which have the ability to form long roots down to 2 m depth. Fritz et al. (2011) reported that in cushion plant lawns, high soil oxygen coincided with these root systems, and methane emissions were absent. We suggest that anoxia could be prevented by the oxygen released by roots which creates a potential microhabitat for aerobic fungi and allow much higher fungal diversity and activity compared to Sphagnum-bogs and fens where anaerobic processes dominate the anoxic zones (Clymo, 1984; Thormann, 2011). Further-more, the link between aerobic decomposition and fungal diversity in cushion bogs warrants further investigations.
We conclude that Astelia-Donatia peatland is a complex ecosystem which has a diversity of filamentous fungal species similar to other peatlands around the world. Future efforts and research will be carried out to understand and quantify the mechanisms that govern the diversity of fungal communities in Tierra del Fuego cushion peatlands.
ACKNOWLEDGMENTS
We thank Hernán Dieguez for editing the map presented in this paper. V. F. Consolo, V. A. Pancotto, A. M. Arambarri and G. L. Salerno are researchers of CONICET (Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina). C. Fritz is a Postdoctoral researcher of Nijmegen University, The Netherlands supported by a FP7 grant (289300 – EUroot) and the ERA foundation. MD Barrera is Professor of Universidad Nacional de La Plata. This research was supported by Universidad Nacional de Mar del Plata, CONICET and FIBA.
BIBLIOGRAPHY
1. Allison, S. & K. Treseder. 2008. Warming and drying suppress microbial activity and carbon cycling in boreal forest soils. Global Change Biology 14: 2898-2909. DOI: http://dx.doi.org/10.1111/j.1365-2486.2008.01716.x [ Links ]
2. Andersen, R.; A. J. Francez & L. Rochefort. 2006. The physicochemical and microbiological status of a restored bog in Québec: Identifcation of relevant criteria to monitor success. Soil Biology & Biochemistry 38: 1375-1387. DOI: http://dx.doi.org/10.1016/j.soilbio.2005.10.012 [ Links ]
3. Anderson, I. C. & J. W. G. Cairney. 2004. Diversity and ecology of soil fungal communities: increased understanding through the application of molecular techniques. Environmental Microbiology 6: 769-779. DOI: http://dx.doi.org/10.1111/j.1462-2920.2004.00675.x [ Links ]
4. Artz, R. R. E.; I. C. Anderson, S. J. Chapman, A. Hagn, M. Schloter, J. Potts & C. D. Campbell. 2007. Changes in fungal community composition in response to vegetational succession during natural regeneration of cutover peatlands. Microbial Ecology 54: 508-522. DOI: http://dx.doi.org/10.1007/s00248-007-9220-7 [ Links ]
5. Basiliko, N.; C. Blodau, C. Roehm, P. Bengston & T. R. Moore. 2007. Regulation of decomposition and methane dynamics across natural commercially mined and restored northern peatlands. Ecosystems 10: 1148-1165. DOI: http://dx.doi.org/10.1007/s10021-007-9083-2 [ Links ]
6. Basiliko, N.; J. B. Yavitt, P. Dees & S. M. Merkel. 2003. Methane biogeochemistry and methanogen communities in two northern peatland ecosystems, New York State. Geomicrobiology Journal 20: 563-577. DOI: http://dx.doi.org/10.1080/713851165 [ Links ]
7. Baumann, M. A. 2006. Water fow, spatial patterns and hydrological self-regulation of a raised bog in Tierra del Fuego (Argentina). Ph D diss. Ernst-Moritz-Arndt University Greifswald. [ Links ]
8. Blodau, C.; N. Basiliko & T. R. Moore. 2004. Carbon turnover in peatland mesocosms exposed to different water table levels. Biogeochemistry 67: 331-351. DOI: http://dx.doi.org/10.1023/B:BIOG.0000015788.30164.e2 [ Links ]
9. Bragazza, L.; C. Siff, P. Iacumin & R. Gerdol. 2007. Mass loss and nutrient release during litter decay in peatland: The role of microbial adaptability to litter chemistry. Soil Biology & Biochemistry 39: 257-267. DOI: http://dx.doi.org/10.1016/j.soilbio.2006.07.014 [ Links ]
10. Carmichael, J. W.; W. B. Kendrick, I. L. Conners & L. Sigler. 1980. Genera of Hyphomycetes. Edmonton: University of Alberta Press. [ Links ]
11. Chen, Y. ; M. G. Dumont, N. P. McNamra, P. M. Chamberlain, L. Bodrossy, N. Strails-Pavese & J. C. Murrel. 2008. Diversity of the active methanotrophic community in acidic peatlands as assessed by mRNA and SIP-PLFA analyses. Environmental Microbiology 10: 446-459. DOI: http://dx.doi.org/10.1111/j.1462-2920.2007.01466.x [ Links ]
12. Clymo, R. S. 1984. The limits to peat bog growth. Philosophical Transactions of the Royal Society Biological Sciences 303: 605-654. DOI: http://dx.doi.org/10.1098/rstb.1984.0002 [ Links ]
13. Domsch, K. H.; W. Gams & T. H. Anderson. 1980. Compendium of soil fungi. Vol. 1. London: Academic Press. [ Links ]
14. Dooley, M. J. & C. H. Dickinson. 1970. Microbiology of cut-away peat II. The ecology of fungi in certain habitats. Plant Soil 32: 454-446. DOI: http://dx.doi.org/10.1007/BF01372883 [ Links ]
15. Ellis, M. B. 1971. Dematiaceous Hyphomycetes. Kew: Commonwealth Mycological Institute. [ Links ]
16. Ellis, M. B. 1976. More Dematiaceous Hyphomycetes. Kew: Commonwealth Mycological Institute. [ Links ]
17. Fierer, N. & R. B. Jackson. 2006. The diversity and biogeography of soil bacterial communities. Proceedings of the Natural Academy of Sciences 103: 626-631. DOI: http://dx.doi.org/10.1073/pnas.0507535103 [ Links ]
18. Fritz, C. 2012. Limits of Sphagnum bog growth in the Southern Hemisphere. Diploma thesis, Radboud University. [ Links ]
19. Fritz, C.; V. A. Pancotto, J. T. Elzenga, E. J. W. Visser, A. P. Grootjans, A. Pol, R. Iturraspe, J. G. M. Roelofs & A. J. P. Smolders. 2011. Zero methane emission bogs: extreme rhizosphere oxygenation by cushion plants in Patagonia. New Phytology 190: 398-408. DOI: http://dx.doi.org/10.1111/j.1469-8137.2010.03604.x [ Links ]
20. Frokling, S. E.; J. L. Bubier, T. R. Moore, T. Ball, L. M. Bellisario, A. Bhardwaj, P. Carroll, P. M. Crill, P. M. Lafeur, J. H. McCaughey, N. T. Roulet, A. E. Suyker, S. B. Verma, J. M. Waddington & G. J. Whiting. 1998. Relationship between ecosystem productivity and photosyntetically active radiation for northern peatlands. Global Biogeochemimal Cycles 12: 115-126. DOI: http://dx.doi.org/10.1029/97GB03367 [ Links ]
21. Gamundí, I. J. & V. Amos. 2007. Exploraciones micológicas en Tierra del Fuego. Boletín de la Sociedad Argentina de Botánica 42: 131-148. [ Links ]
22. Gebser, R. 2008. Ecological studies on the vegetation of the Moat cushion peatland in Tierra del Fuego, Argentina. MSc Thesis, University of Greifswald. [ Links ]
23. Gilbert, D. & E. A. D. Mitchell. 2006. Microbial diversity in Sphagnum peatlands, en I. P. Martini, A. Martínez Cortizas & W. Chesworth (eds.), Peatlands: Evolution and records of environmental and climate change, pp. 287-318. Amsterdam-Oxford: Elsevier. [ Links ]
24. Giri, B.; P. Huong Giang, R. Kumari, R. Prassad & A. Varma. 2005. Microbial diversity in soils. Soil Biology 3: 19-55. DOI: http://dx.doi.org/10.1007/3-540-26609-7_2 [ Links ]
25. Godeas, A. M. 1983. Estudios cuali-cuantitativos de los hongos del suelo de Nothofagus dombeyi. Ciencia del suelo 1: 21-31. [ Links ]
26. Godeas, A. M. & A. M. Arambarri. 2007. Hifomicetes lignícolas de Tierra del Fuego (Fungi, Fungi Imperfecti, Hypho-mycetales). Boletín de la Sociedad Argentina de Botánica 42: 1-2. [ Links ]
27. Horton, T. R. & T. D. Bruns. 2001. The molecular revolution in ectomycorrhizal ecology: peeking into the black-box. Molecular Ecology 10: 1855-1871. DOI: http://dx.doi.org/10.1046/j.0962-1083.2001.01333.x [ Links ]
28. Jaatinen, K.; H. Fritze, J. Laine & R. Laiho. 2007. Effects of short- and long-term water level draw- down on the populations and activity of aerobic decomposers in a boreal peatland. Global Change Biology 13: 491-510. DOI: http://dx.doi.org/10.1111/j.1365-2486.2006.01312.x [ Links ]
29. Kamal, S. A. & A. Varma. 2008. Peatland Microbiology, en P. Dion & C. Nautiyal (eds.) Microbiology of Extreme Soils, pp. 177-203. Berlin-Heidelberg: Springer. [ Links ]
30. Khalid, M.; W. Yang, N. Kishear, Z. Rajput & A. Arizo. 2006. Study of cellulolytic soil fungi and two nova species and new medium. Journal of Zhejiang University Science 7: 459-466. DOI: http://dx.doi.org/10.1631/jzus.2006.B0459 [ Links ]
31. Killham, K. 1994. Soil Ecology. Cambridge: Cambridge University Press. [ Links ]
32. Kip, N.; C. Fritz, E. Langelaan, Y. Pan, L. Bodrossy, V. A. Pancotto, M. S. M. Jetten, A. J. M. Smolders & H. J. M den Camp. 2012. Methanotrophic activity and diversity in different Sphagnum magellanicum dominated peat bog habitats in Tierra del Fuego. Biogeosciences 9: 47-55. DOI: http://dx.doi.org/10.5194/bg-9-47-2012 [ Links ]
33. Koesnandar, P. S.; D. Nurani & E. Wahyono. 2006. Government role on research and application of technology for peatland utilization. National Seminar on Peatlands and their problems, Pontianak: University of Tnajungpura. [ Links ]
34. Kovach, W. L. 1999. MVSP-A Multivariate Statistical Package for Windows, Version 3.1. Pentraeth: Kovach Computing Services. [ Links ]
35. Küster, E. 1963. Infuence of peat substances on the metabolism of fungi. Pure and Applied Chemistry 4: 611-616. [ Links ]
36. Lin, X.; S. Green, M. M. Tfaily, O. Prakash, K. T. Konstantinidis, J. E. Corbett, J. P. Chanton, W. T. Cooper & J. E. Kostka. 2012. Microbial community structure and activity linked to contrasting biogeochemical gradients in bog and fen environments of the Glacial Lake Agassiz Peatland. Applied and Environmental Microbiology 78: 7023-7031. DOI: http://dx.doi.org/10.1128/AEM.01750-12 [ Links ]
37. Magurran, A. E. 1988. Ecological diversity and its measurement. New Jersey: Princeton University Press. DOI: http://dx.doi.org/10.1007/978-94-015-7358-0 [ Links ]
38. Malvárez, A. I.; P. Kandus & A. Carbajo. 2004. Distribución regional de los turbales en Patagonia (Argentina y Chile), en D. E. Blanco & V. M. de la Balze (eds.), Los turbales de la Patagonia. Bases para su inventario y la conservación de la biodiversidad, publicación n° 19, pp. 22-29. Buenos Aires: Wetlands International-América del Sur. [ Links ]
39. Moore, D. M. 1983. Flora of Tierra del Fuego. Nelson-Oswestry: Lubrecht & Cramer, Ltd. [ Links ]
40. Moore, T. R. & N. Basiliko. 2006. Decomposition in boreal peatlands, en R. K. Wieder & D. H. Vitt (eds.), Boreal peatland ecosystems, pp 125-143. Berlin: Springer [ Links ]
41. Murray, M. G. & W. F. Thompson. 1980. Rapid isolation of high molecular weight plant DNA. Nucleic Acid Research 8: 4321-4325. DOI: http://dx.doi.Org/10.1093/nar/8.19.4321 [ Links ]
42. Nakasone, K.; S. Peterson & S. Jong. 2004. Preservation and distribution of fungal cultures, en G.M. Mueller, G. F. Bills & M. Foster (eds.), Biodiversity of fungi. Inventory and monitoring methods, pp. 37-47 Amsterdam: Elsevier Academic Press. [ Links ]
43. Pancotto, V. A.; O. E. Sala, M. Cabello, N. I. López, T. M. Robson, C. Bailaré, M. Caldwell & A. Scopel. 2003. Solar UV-B decreases decomposition in herbaceous plant litter in Tierra del Fuego, Argentina: potential role of an altered decomposer community. Global Change Biology 9: 1465-1474. DOI: http://dx.doi.Org/10.1046/j.1365-2486.2003.00667.x [ Links ]
44. Peltoniemi, K.; H. Fritze & R. Laiho. 2009. Response of fungal and actinobacterial communities to water-level drawdown in boreal peatland sites. Soil Biology & Biochemistry 41: 1902-1914. DOI: http://dx.doi.Org/10.1016/j.soilbio.2009.06.018 [ Links ]
45. Pisano, E. 1983. The Magellanic tundra complex, en A. J. P. Gore (ed.), Mires: swamp, bog, fen and moor, vol. 2, pp. 295-329. Amsterdam: Elsevier. [ Links ]
46. Preston, M. D.; K. A. Smemo, J. W. McLaughlin & N. Basiliko. 2012. Peatland microbial communities and decomposition processes in the James Bay Lowlands, Canadá. Frontiers in Microbiology 3: 1-15. DOI: http://dx.doi.org/10.3389/fmicb.2012.00070 [ Links ]
47. Ramírez, C. & T. Martínez. 1982. Manual and atlas of the Penicillia. Amsterdam-New York-Oxford: Elsevier Biomedical Press. [ Links ]
48. Repečkiené, J.; Jukoniené, I & O. Salina. 2012. Cellulosede-composing fungi in peatlands occupied by invasive moss Campylopus introfexus. Botánica Lithuanica 18:46-57. DOI: http://dx.doi.org/10.2478/vl0279-012-0007-5 [ Links ]
49. Robson, T. M.; V. A. Pancotto, C. L. Bailaré, O. E. Sala, A. L. Scopel & M. M. Caldwell. 2004. Reduction of solar UV-B mediates changes in the Sphagnum capitulum microenvironment and the peatland microfungal community. Oecologia 140: 480-490. DOI: http://dx.doi.org/10.1007/s00442-004-1600-9 [ Links ]
50. Roig, C. & L. Collado. 2004. Moat, provincia de Tierra del Fuego. Anexo II-G, en D. E. Blanco & V. M. de la Balze (eds.), Los Turbales de la Patagonia. Bases para su inventario y la conservación de su biodiversidad, publicación nº 19, pp. 66-78. Buenos Aires: Wetlands International-América del Sur. [ Links ]
51. Rousk, J.; E. Baath, P. C. Brookes, C. L. Lauber, C. Lozupone, J. G. Caporaso, R. Knight & N. Fierer. 2010. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME Journal 4: 1340-1351. DOI: http://dx.doi.org/10.1038/ismej.2010.58 [ Links ]
52. Searles, P. S.; B. R. Kropp, S. D. Flint & M. M. Caldwell. 2001. Infuence of solar UV-B radiation on peatland microbial communities of southern Argentina. New Phytologist 152: 213-221. DOI: http://dx.doi.org/10.1046/j.0028-646X.2001.00254.x [ Links ]
53. Sizova, M. V.; N. S. Panikov, T. P. Tourova & P. W. Flanagan. 2003. Isolation and characterization of oligotrophic acido-tolerant methanogenic consortia from a Sphagnum peat bog. FEMS Microbiology Ecology 45: 301-315. DOI: http://dx.doi.org/10.1016/S0168-6496(03)00165-X [ Links ]
54. Thormann, M. N. 2005. Diversity and function of fungi in peat-lands: a carbon cycling perspective. Canadian Journal of Soil Science 86: 281-293. DOI: http://dx.doi.org/10.4141/S05-082 [ Links ]
55. Thormann, M. N. 2006. The role of fungi in decomposition dynamics in peatlands, en R. K. Wieder & H. D. Vitt (eds.), Boreal Peatland Ecosystems, pp. 101-123. Berlin: Springer-Verlag. [ Links ]
56. Thormann, M. N. 2011. In vitro decomposition of Sphagnum-derived acrotelm and mesotelm peat by indigenous and alien basidiomycetous fungi. Mires and Peats 8: 1-12. [ Links ]
57. Thormann, M. N.; R. S. Currah & S. E. Bayley. 2003. Succession of microfungal assemblages in decomposing peat-land plants. Plant Soil 250: 323-333. DOI: http://dx.doi.org/10.1023/A:1022845604385 [ Links ]
58. Thormann, M. N.; R. S. Currah & S. E. Bayley. 2004. Patterns of distribution of microfungi in decomposing bogs and fen plants. Canadian Journal of Botany 82: 710-720. DOI: http://dx.doi.org/10.1139/b04-025 [ Links ]
59. Thormann, M. N. & A. V. Rice. 2007. Fungi from peatlands. Fungal Diversity 24: 241-299. [ Links ]
60. Viaud, M.; A. Pasquier & Y. Brygoo. 2000. Diversity of soil fungi studied by PCR-RFLP of ITS. Mycological Research 104: 1027-1032. DOI: http://dx.doi.org/10.1017/S0953756200002835 [ Links ]
61. Watrud, L. S; K. Martin, K. K. Donegan, J. K. Stone & C. G. Coleman. 2006. Comparison of taxonomic, colony morphotype and PCR-RFLP methods to characterize microfungal diversity. Mycologia 98: 384-392. DOI: http://dx.doi.org/10.3852/mycologia.98.3.384 [ Links ]
62. White, T. M.; T. Bruns, S. Lee & J. Taylor. 1990. Amplifcation and direct sequencing of fungal ribosomal RNA for phylogenetics, en M. A. Innis, D. H. Gelfand, J. J. Sninsky & T. J. White (eds), PCR protocols: a guide to methods and applications, pp 315-321. San Diego: Academic Press. [ Links ]

