










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkDarwiniana, nueva serie
Print version ISSN 0011-6793On-line version ISSN 1850-1699
Darwiniana, nueva serie vol.4 no.2 San Isidro Dec. 2016
SISTEMÁTICA Y TAXONOMÍA DE PLANTAS
Revisión del género megalastrum (dryopteridaceae) para la Argentina
Marcelo D. Arana1, Jefferson Prado2 & Mónica Ponce3
1 Orientación Plantas Vasculares, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, Universidad Nacional de Río Cuarto, Ruta 36 km 601, X5804ZAB Río Cuarto, Córdoba, Argentina; marana@exa.unrc.edu.ar (autor corresponsal)
2 Instituto de Botânica, Av. Miguel Stéfano 3687, Agua Funda, C.P. 68041, CEP 04045-972, São Paulo, Brasil; jprado.01@uol.com.br
3 Instituto de Botánica Darwinion, (ANCEF-CONICET), Labardén 200, C.C. 22, B1624HYD San Isidro, Buenos Aires, Argentina; mponce@darwin.edu.ar
Abstract
So far the information about the genus Megalastrum (Dryopteridaceae) for Argentina is fragmented and incomplete. A revision of Megalastrum in Argentina is here presented; it is represented by six species: M. adenopteris, M. ciliatum, M. connexum, M. crenulans, M. fugaceum, and M. spectabile. A key to the species is provided, plus descriptions, notes on distribution and habitat, as well as illustrations of diagnostic characters for each taxon. Megalastrum crenulans and M. spectabile are lectotypified. The species occur primarily in the Argentinean forests, with a disjunct distribution, three species grow in the north-western Yungas, two species grow in the Parana forest, and one species grows in the Valdivian temperate rain forest. Megalastrum pulverulentum and M. subincisum are excluded from the Argentinean flora.
Keywords: Dryopteridaceae; Eupolypods I; Ferns; Flora; Southern Cone; Taxonomy.
Revision of the genus Megalastrum (Dryopteridaceae) for Argentina
Resumen
La información referente al género Megalastrum (Dryopteridaceae) para la Argentina es hasta ahora fragmentaria e incompleta. Se presenta aquí una revisión de Megalastrum en Argentina, el cual está representado por seis especies: M. adenopteris, M. ciliatum, M. connexum, M. crenulans, M. fugaceum y M. spectabile. Se provee una clave para las especies, junto con descripciones, notas sobre la distribución y el hábitat, y se incluyen ilustraciones de los caracteres diagnósticos para cada taxón. Se lectotipifican Megalastrum crenulans y M. spectabile. Las especies habitan primariamente en las selvas de Argentina, con una distribución disyunta, tres especies crecen en las Yungas del noroeste, dos especies crecen en la selva Paranaense y una especie crece en la selva Valdiviana templada. Megalastrum pulverulentum y M. subincisum son excluidas de la flora Argentina.
Palabras clave: Cono Sur; Dryopteridaceae; Eupolipoideas I; Flora; Helechos; Taxonomía.
Original recibido el 29 de abril de 2016,
aceptado el 8 de junio de 2016
Editora: Nataly O’Leary
INTRODUCCIÓN
Megalastrum Holttum es un género de helechos que pertenece a la familia Dryopteridaceae, linaje que incluye alrededor de 1700 especies de distribución subcosmopolita (ausente en Europa y norte de Asia) y cuyo ancestro data de aproximadamente 100 millones de años (Schuettpelz & Pryer, 2009). El género comprende helechos de tamaño mediano a grande con frondes hasta de 4 metros, generalmente con lámina muy dividida (1-4-pinnado-pinnatífidas). También se caracteriza por poseer rizomas erectos a decumbentes, pinnas basales prolongadas en el lado basiscópico, raquis de las pinnas y cóstulas adaxialmente pubescentes y nervaduras con ápice claviforme, engrosado. Además, se puede distinguir por la combinación de los siguientes caracteres: la pilosidad blanquecina de los ejes foliares, con pelos estrigosos multiseptados de ápice agudo, y la característica de la venación de los segmentos en las porciones distales del raquis, en donde los segmentos se vuelven progresivamente más adnatos y decurrentes sobre el raquis; en estos segmentos la vena que lo irriga se origina en el raquis de la pinna y no de la cóstula (Holttum, 1986; Smith & Moran, 1987; Moran & Prado, 2010). Entre las Dryopteridaceae, este tipo de venación de los segmentos distales sólo está presente en algunas especies de Ctenitis (C. Chr.) C. Chr. y Stigmatopteris C. Chr. (Moran et al., 2014a). Sin embargo, la combinación del origen de la vena del segmento basiscópico de las pínnulas y la pilosidad adaxial de los ejes en Megalastrum, con pelos estrigosos, multiseptados, blanquecinos, de ápice agudo, a veces antrorsos, esúnica y define al género (Smith & Moran, 1987). Estas características fueron descriptas primeramente por Christensen (1913) como particulares de uno de los 11 subgéneros (subg. Subincisae) en que dividió a Dryopteris Adans., grupo que luego reconoció como sección Subincisae C. Chr. (Christensen, 1934). Posteriormente, Tindale (1965) trasladó la sección Subincisae al género Ctenitis (C. Chr.) C. Chr., criterio seguido por la mayoría de los autores sucesivos (Stolze, 1981; Mickel & Beitel, 1988; Proctor, 1989), hasta que Holttum (1986) elevó el rango de la sección a nivel genérico como Megalastrum, criterio seguido por Smith & Moran (1987). Estudios moleculares muestran que Megalastrum es monofilético y forma parte de un clado denominado ‘‘helechos lastreopsidos’’ (Labiak et al., 2014). Este clado es de distribución pantropical, extendiéndose hacia las zonas templadas del hemisferio sur, e incluye a los géneros Lastreopsis Ching, Megalastrum, Parapolystichum (Keyserl.) Ching y Rumohra Raddi (Schuettpelz & Pryer, 2007; Labiak et al., 2014, 2015a, 2015b).
Megalastrum está compuesto por alrededor de 91 especies de distribución principalmente selvática, estando la mayor diversidad en la región Neotropical, con solo tres especies en África y Madagascar (Rohuan & Moran, 2011; Moran et al., 2014a; 2014b). El rango altitudinal del hábitat varía entre los 250–3500 m s.m., con la mayoría de las especies entre los 800–2000 m s.m.
No existe un tratamiento integral del género para la Argentina, sólo ha sido tratado en algunas floras regionales (de la Sota, 1977; de la Sota et al., 1998; Ponce, 2001; Ponce & Martínez, 2012). Estudios recientes (Moran et al., 2014a, 2014b) muestran que estos tratamientos están desactualizados, por lo tanto se presenta esta revisión, que incluye la descripción de todos los taxones del género que habitan en la Argentina, una clave, ilustraciones y comentarios sobre la distribución y características del hábitat.
MATERIALES Y MÉTODOS
El estudio se llevó a cabo utilizando la consulta bibliográfica y revisión de los ejemplares depositados en los herbarios: BA, CTES, JUA, MCNS, MERL, LIL, LP, RB, RCVC, SI y SP (Thiers, 2016). Además se realizaron viajes de estudio a varias provincias de Argentina y el material coleccionado fue depositado en JUA, LP, RCVC y SI.
Megalastrum en general comprende plantas que alcanzan un gran tamaño, lo que implica que en general los especímenes de herbario se encuentren en estado fragmentario y que consistan en la mayoría de los casos en pinnas o fragmentos de ellas y, en pocos casos, anexada la base del pecíolo. Debido a esto, priorizamos el análisis de caracteres observables en estos sectores del cuerpo de la planta, tales como la pubescencia y las escamas, así como la presencia y pubescencia del indusio. Las características de las esporas fueron observadas utilizando microscopio óptico, y para la descripción de las mismas se siguió el criterio de Moran et al. (2014a).
TRATAMIENTO TAXONÓMICO
Megalastrum Holttum, Gard. Bull. Singapore 39(2): 161. 1987 [1986 publ. 1987]. ESPECIE TIPO: Megalastrum villosum (L.) Holttum (= Polypodium villosum L.).
Dryopteris sect. Subincisae C. Chr., Index Filic., Suppl. 3: 7. 1934. Ctenitis C. Chr. sect. Subincisae (C. Chr.) Tindale, Contr. New South Wales Natl. Herb. 3(5): 252. 1965. ESPECIE TIPO: Dryopteris subincisa (Willd.) Urb. (= Polypodium subincisum Willd.).
Plantas terrestres. Rizomas erectos a decumbentes. Pecíolos escamosos en la base, con 4-10 haces vasculares, los dos adaxiales de mayor tamaño; láminas hasta de 4 x 1,4 m, 1-4-pinnado-pinnatífidas, pinnas basales más divididas que las restantes, no equiláteras, mayores del lado basiscópico o más raramente equiláteras, catádromas por encima de las pinnas basales, tejido laminar sin glándulas; raquis, costas, y cóstulas escamosas abaxialmente, adaxialmente con tricomas pluricelulares, blanquecinos, con ápice agudo, pelos glandulares ausentes o, si presentes, de ca. 0,1 mm de ancho, esféricos, amarillentos o naranja pálido, sésiles o pedicelados; segmentos basiscópicos de las pínnulas basales decurrentes y adnatos al raquis, segmentos basales de las pínnulas distales con vena basal basiscópica, surgiendo de la costa (no de la cóstula), venas en general con ápice claviforme, submarginales. Soros generalmente sin indusio o, menos frecuentemente con indusios circulares desarrollados o reducidos, caducos o no. Esporas monoletes, equinadas o crestadas, con alas cortas, semiparalelas. Número básico de cromosomas x = 41 (Moran et al., 2014a).
Género de 91 especies de distribución mundial, 46 de las cuales crecen en la región andina de América del Sur (Moran et al., 2014a). En Argentina se encuentran seis especies nativas, de distribución disyunta en las áreas selváticas (Ponce et al., 2002).
Observaciones. Megalastrum puede distinguirse de otros helechos de gran porte por sus pinnas basales no equiláteras, basiscópicamente más desarrolladas, cara adaxial con tricomas blanquecinos, antrorsamente estrigosos, pluricelulares y con ápice agudo no retorcido; además en las porciones distales del raquis los segmentos se vuelven sucesivamente más adnatos y decurrentes sobre el raquis, en estos segmentos la vena que irriga al segmento se origina en el raquis de la pinna y no de la cóstula (Fig. 1).
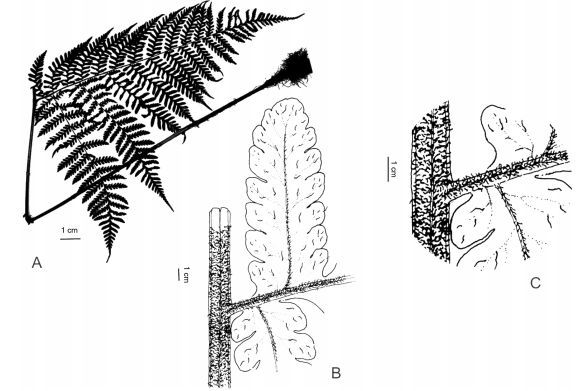
Fig. 1. Características morfológicas de Megalastrum. A, Megalastrum crenulans, pinna basal inequilátera, con mayor desarrollo de los segmentos basiscópicos. B y C, Megalastrum fugaceum, últimos segmentos adnatos y decurrentes sobre el raquis, mostrando el origen, en el raquis de la pinna, de la vena que lo irriga. A, de Handro 2224 (US); B-C, de Moran 5911 (UC).
Clave para las especies de Megalastrum en Argentina
1. Indusio notorio y persistente, o inconspicuo y caduco .............................................................................................. 2
1. Indusio ausente .......................................................................................................................................................... 4
2(1). Indusio notorio, circular, de 0,5-0,7 mm de diám. ............................................................................. M. crenulans
2. Indusio inconspicuo y caduco, de 0,1-0,2 mm de diám. ............................................................................................ 3
3(2). Lámina entre las venas densamente pubescente en ambas caras y con pelos glandulares amarillos, capitados;
indusio con pelos blanquecinos ca. 0,1 mm en la base o centro del soro ................................................ M. adenopteris
3. Lámina entre las venas en general glabra, a veces pubescente pero eglandular en ambas caras; indusio aparentando
un grupo de protoescamas rojizas, ca. 0,1 mm, que sobresalen desde el centro de los soros (no se observa fácilmente en
soros maduros) ............................................................................................................................................. M. fugaceum
4(1). Frondes fértiles de 0,5-0,8 m; raquis de las pinnas abaxialmente escamosos, con escamas fuertemente denticula-
das; adaxialmente pubescentes, sin escamas. Yungas .................................................................................... M. ciliatum
4. Frondes fértiles de 2-2,5 m; raquis de las pinnas glabro, pubescente o escamoso en ambas caras, escamas de margen
entero o eroso. Selva Paranaense o Bosques Andino-patagónicos ................................................................................ 5
5(4). Raquis y costas con pubescencia glandular, glándulas sésiles, hialinas a amarillentas y escamas; cóstulas con
escamas lanceoladas, de margen eroso. Bosques Andino-patagónicos ...................................................... M. spectabile
5. Raquis y costas con pubescencia no glandular, pelos simples de 0,1-0,5 mm; cóstulas con escamas filiformes a
linear-lanceoladas, de margen entero. Selva Paranaense .......................................................................... M. connexum
Megalastrum adenopteris (C. Chr.) A.R. Sm. & R.C. Moran, Amer. Fern J. 77(4): 127. 1987. Dryopteris adenopteris C. Chr., Kongel. Danske Vidensk. Selsk. Skr., Naturvidensk. Math. Afd., ser. 8, 6: 85. 1920. Ctenitis adenopteris (C. Chr.) Ching, Sunyatsenia 5: 250. 1940. TIPO: Brasil, Rio Grande do Sul, Silveira Martins, Val Veneta, ad terram silvae primaevae, 10-III-1893, C. A. M. Lindman s.n. (Regnell A 1313) (lectotipo, BM-000907710! designado por R. C. Moran et al., Amer. Fern J. 99: 8. 2009; isolectotipos C, L, MO-255620! fragmento ex US, S06-454!, S06-546!, S-R-1693!, U 0007363! fragmento ex US, UPS!, US 00067050!, fragmento ex MO!). Fig. 2.
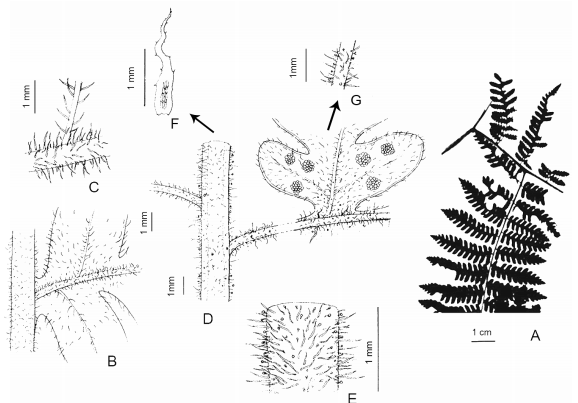
Fig. 2. Megalastrum adenopteris. A, disección de la pinna basal. B, superficie adaxial del raquis de la pinna y pínnula. C, detalle de los pelos de la cóstula. D, superficie abaxial del raquis de la pinna, cóstula y pínnula. E, raquis de la pinna. F, escama del raquis de la pinna. G, detalle de los pelos de la cóstula. A, de Venturi 886 (US); B-G, de Jürgens s.n. (S).
Dryopteris villosa (L.) Kuntze var. tomentosa Rosenst., Hedwigia 46: 130. 1906. TIPO: Brasil, Rio Grande do Sul, Rio Pardo, Fazenda Soledade, 1906, L. C. Jürgens s.n. (Herb. Rosenstock Filices Austrobrasilienses no. 207) (lectotipo, MICH! designado por R. C. Moran et al., Amer. Fern J. 99: 9. 2009; isolectotipos, B 20 0051906!, B 20 0051907!, S 06-547!).
Dryopteris oreocharis Sehnem var. canescens Sehnem, Fl. Ilustr. Catarin. 1(ASPID): 177. 1979. TIPO: Brasil, Santa Catarina, Lauro Müller, Novo Horizonte, 400 m s.m., 24-X-1958, R. Reitz & R. M. Klein 7516 (lectotipo, PACA-68744! designado por R. C. Moran et al., Amer. Fern J. 99: 9. 2009; isolectotipo, HBR!).
Rizomas erectos hasta de 50 cm de alto, con escamas linear-lanceoladas, de 10-20 × 1-2 mm, adpresas o ascendentes, levemente tortuosas apicalmente, margen denticulado, anaranjadas, opacas o brillantes. Frondes fértiles hasta de 4 m; pecíolos con la base escamosa, las escamas iguales a las del rizoma pero de disposición más espaciada; láminas de 1-2 m, base 4-pinnada-pinnatífida, en la parte media 3-pinnada-pinnatisecta; raquis de la lámina pubescente adaxialmente; pinnas basales hasta de 1 m, no equiláteras, el lado basiscópico más desarrollado; raquis y costas de las pinnas piloso-glandulares en ambas caras, con escamas esparcidas, las glándulas sésiles o pediceladas, amarillas, los pelos 1-3(-4) celulares, aciculares, erectos; las escamas ovadas a lanceoladas, de 0,5-2,5 × 0,2-0,5 mm, adpresas, denticuladas, castañas, lustrosas, no clatradas, no buladas; pínnulas basal-basiscopicas de las pinnas medias no alargadas ni superpuestas sobre el raquis de la lámina; cóstulas en ambas superficies densamente pubescentes, abaxialmente con pelos glandulares y poco escamosas, pelos 0,3-0,4 mm, 3-4 celulares; venas y lámina entre las venas densamente pubescentes en ambas caras, pelos de 0,1-0,3(-0,5) mm, 1-4 celulares, a menudo capitados, amarillentos o anaranjados, márgenes de la lámina con pelos simples, 0,2-0,3 mm. Indusios inconspicuos y caducos, de 0,1-0,2 mm de diám., con la apariencia de un grupo de pelos blanquecinos ca. 0,1 mm, no glandulares, en la base o centro del soro. Esporas equinadas.
Distribución y hábitat. Esta especie habita en Perú, Bolivia, sudeste de Brasil y noroeste de la Argentina, desde los 750 hasta los 2100 m s.m. (Fig. 8). Crece en quebradas o laderas húmedas en el sotobosque de las selvas y bosques montanos.
Observaciónes. Megalastrum adenopteris es cercana a M. pulverulentum (Poir.) A.R. Sm. & R.C. Moran, especie que no se encuentra en la Argentina y se distribuye desde México y las Antillas, América Central y norte de América del Sur hasta Perú. Se diferencia de M. adenopteris por tener la lámina glabra entre venas en ambas superficies y por la ausencia de indusios. Megalastrum adenopteris se caracteriza por sus frondes de gran tamaño, muy divididas, con la lámina densamente pubescente en ambas caras, a menudo con pelos glandulares de célula apical globosa, amarillenta o anaranjada, indusio fugaz (aparentemente ausente), evidente por un mechón de pelos blanquecinos en el centro del soro (Moran et al., 2014a).
Material representativo examinado
ARGENTINA. Jujuy. Depto. Ledesma, Parque Nacional Calilegua, camino a Mesada de las Colmenas, 24-V-2011, Ganem et al. 25 (JUA, LP, RCVC). Salta. Depto. Anta, Parque Nacional El Rey, 21-III-1981, Brown 1339 (MO, NY). Tucumán. Depto. Tafí, en quebrada húmeda y con agua terrestre, 850 m, 15-VIII-1920, Venturi 886 (SI). Quebrada de Cañizo, 900 m, 18-XI-1951, Sleumer 2097 (BM). Quebrada de las Piedras, Bosques subtropicales, 600 m, 08-I-1899, Lillo 2207 (SI). Ídem, 750 m, 27-IV-1890, Lillo 1692 (CTES, LIL).
Megalastrum ciliatum M. Kessler & A.R. Sm., Amer. Fern J. 96(1): 38. 2006. TIPO: Bolivia, La Paz, Sud Yungas, camino Chulumani–Ocobaya, 2.5 km después de Chulumani, 1600 m, 6-I-1992, J. P. Schmit, T. Mione & W. Zuazo 422 (holotipo UC 1616956!; isotipo LPB). Fig. 3.
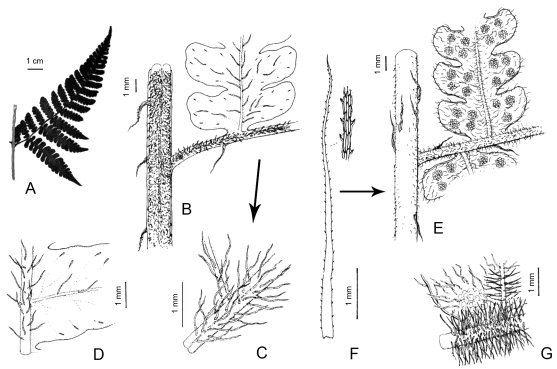
Fig. 3. Megalastrum ciliatum. A, disección de la pinna basal. B, superficie adaxial del raquis de la fronde, de la pinna y pínnula. C, detalle de los pelos del raquis y la cóstula. D, superficie adaxial de la cóstula y último segmento. E, superficie abaxial del raquis de la fronde, raquis de la pinna y pínnulas. F, escama del raquis, con detalle del margen. G, detalle de los pelos del raquis. De Fuentes et al. 9013 (MO, UC).
Rizomas erectos hasta 20 cm alto, con escamas linear-lanceoladas, de 6-10 x 1-1,5 mm, castañas con márgenes total o parcialmente negros, fuertemente denticulados, brillantes. Frondes fértiles, relativamente pequeñas, de 0,5-0,8 m; pecíolos con la base escamosa, las escamas iguales a las del rizoma pero de disposición más espaciada y levemente ascendentes; láminas de 0,4-0,7 m, base 3-pinado-pinnatífidas, parte media 2-pinado-pinnatisectas; raquis de la lámina glabros abaxialmente; pinnas basales de 10-30 cm, no equilaterales, raquis y costas de las pinnas pubescentes, abaxialmente escamosos, pelos de 0,8-1,5 mm, 4-7(-8) celulares, aciculares, escamas 1-2 mm, castaño oscuras, a veces más oscuras distalmente, linear-lanceoladas, fuertemente denticuladas, no clatradas, no buladas, adaxialmente con escasas glándulas, de ca. 0,1 mm, esféricas, sésiles, de color naranja amarillento, pínnulas basal-basiscopicas de las pinnas medias no alargadas ni superpuestas sobre el raquis de la lámina; cóstulas sin glándulas, en ambas superficies densamente pubescentes, poco escamosas, pelos 0,7-1 mm, 3-6 celulares, adpresos a ascendentes, escamas de 1,5 mm, linear-lanceoladas, brillantes, fuertemente denticuladas; venas y lámina entre las venas abaxialmente con pocas glándulas globosas, sésiles, anaranjado-amarillentas, moderado a densamente puberulentas, pelos de 0,6-0,8(-1) mm, adpresos a erguidos, aciculares, adaxialmente glabro; venas visibles en ambas caras o sólo abaxialmente, márgenes de la lámina con los pelos 0,4-1 mm. Indusios ausentes. Esporas crestadas.
Distribución y hábitat. Especie endémica de los Andes del sur de Bolivia (Kessler & Smith, 2006) y del noroeste de la Argentina, crece entre los 1600-1830 m s.m.; en lugares muy húmedos del sotobosque de las Yungas en la provincia de Jujuy (Fig. 8).
Observaciones. Megalastrum ciliatum se caracteriza por sus frondes fértiles relativamente pequeñas, que no alcanzan el metro de longitud, venas adaxialmente con pelos estrigosos de 0,5-1 mm, tejido laminar abaxialmente pubescente, con pelos de 0,8-1 mm y raquis de las pinnas y cóstulas con escamas fuertemente denticuladas.
Material representativo examinado
ARGENTINA. Jujuy. Depto. Ledesma, Parque Nacional Calilegua, Aguada del Tigre, 1600 m, 7-IV-2010, Martínez & Prado 1893 (MCSN, NY, SP).
Megalastrum connexum (Kaulf.) A.R. Sm. & R.C. Moran, Amer. Fern J. 77(4): 127 1987. Polypodium connexum Kaulf., Enum. Filic.: 120. 1824. Phegopteris connexa (Kaulf.) Christ, Prim. Fl. Costaric. 3(1): 36. 1901. Dryopteris connexa (Kaulf.) C. Chr., Index Filic.: 258. 1905. Ctenitis connexa (Kaulf.) Copel., Ann. Crypt. Phytopath. [Gen. Fil.] 5: 124. 1947. TIPO: Brasil, Santa Catharina, Ilha de Santa Catarina (Florianópolis), Sertao da Lagoa, 26-VI-1948, J. A. Rohr 1071 (neotipo US [3 hojas] US 01105667!, US 1105668! & US 01105669! designado por R. C. Moran et al., Amer. Fern J. 99(1): 13. 2009; isoneotipos, HB!, HBR!, NY 00678920!). Fig. 4.
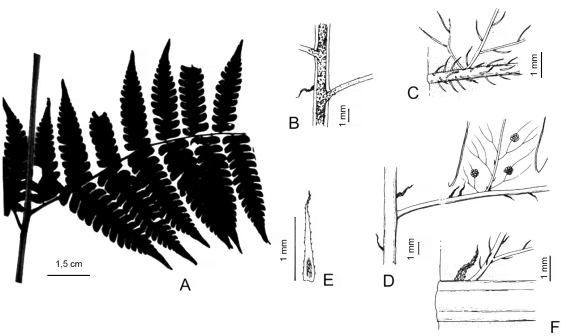
Fig. 4. Megalastrum connexum. A, disección de la pinna basal. B, superficie adaxial de la cóstula. C, detalle de los pelos en la superficie adaxial. D, superficie abaxial de la pínnula. E, escama de la cóstula. F, detalle de la cóstula mostrando escamas y pelos. A, de Matos et al. 1104 (NY); B-F, de Rohr 1071 (US).
Phegopteris brevinervis Fée, Crypt. Vasc. Brésil 1: 243, t. 77, fig. 2. 1869. Stigmatopteris brevinervis (Fée) R. C. Moran, Ann. Missouri Bot. Gard. 78(4): 871. 1991. TIPO: Brasil, sine data, P. Claussen s.n. (holotipo P00610577!).
Phegopteris eriopodia Fée, Crypt. Vasc. Brésil 1: 102, t. 31, fig. 1. 1869. TIPO: Brasil, Rio de Janeiro, Rio de Janeiro, Gavia sur les bords du Chemin, 12-III-1868, A. F. M. Glaziou 2397 (lectotipo P-00610813! designado por R. C. Moran et al., Amer. Fern J. 99: 13. 2009).
Phegopteris adnata Fée, Crypt. Vasc. Brésil 1: 103. 1869. TIPO: Brasil, Rio de Janeiro, Rio de Janeiro, Alto Macaé, 21-V-1868, A. F. M. Glaziou 2398 (lectotipo P-00610860! designado por R. C. Moran et al., Amer. Fern J. 99: 13. 2009).
Phegopteris propinqua Fée, Crypt. Vasc. Brésil 1: 103, t. 32, fig. 3. 1869. TIPO: Brasil, Rio de Janeiro, Rio de Janeiro, Fazenda do Ariro, 28-VI-1868, A. F. M. Glaziou 2399 (holotipo P00170108!; isotipo P00610856!).
Polypodium willsii Baker, Ann. Bot. (Oxford) 5: 458. 1891. Dryopteris willsii (Baker) C. Chr., Index Filic.: 301. 1905. TIPO: Brasil, Rio de Janeiro, Rio de Janeiro, VI-1881, A. Wills s.n. (holotipo K000512181!).
Phegopteris lateadnata Christ, Annuaire Conserv. Jard. Bot. Genève 3: 36 1899. Dryopteris lateadnata (Christ) C. Chr., Index Filic. 274. 1905 [como ‘‘D. latealata’’, error tipográfico]. Dryopteris connexa var. lateadnata (Christ) C. Chr., Kongel. Danske Vidensk. Selsk. Skr., Naturvidensk. Math. Afd., ser. 8, 6: 80. 1920. Megalastrum connexum var. lateadnatum (Christ) A.R. Sm. & R.C. Moran, Amer. Fern J. 77(4): 127. 1987. TIPO: Paraguay, Caaguazú, dans les forêts, 1-IV-1885, B. Balansa 313a (holotipo P-00610679!; isotipos B, K, L, P).
Rizomas erectos o decumbentes, densamente escamosos, con escamas castañas clatradas, lineares, de margen dentado. Frondes de 1,5-2,5 m; pecíolos gruesos castaño claros, con escamas en la base hasta de 2 cm, linear a lanceoladas, escasamente denticuladas, pardo-amarillentas o pardo-doradas, retorcidas o crispadas, formando una masa densa; láminas de 1-2 m, 3-pinnadopinnatífidas en la base, 2-pinnado-pinnatífidas en la parte media; raquis abaxialmente, glabro o subglabro, no glandular con pocas escamas (por lo general en la base de las pinnas) de 2,5 mm, lineares, denticuladas, adaxialmente densamente pubescentes, no glandulares, pelos de 0,1-0,5 mm, 1-4 celulares; pinnas basales de 30-50 cm, peciólulos de hasta 2 cm, fuertemente no equilaterales, pínnulas acroscópicas ligeramente reducidas hacia la base de las pinnas; costas y cóstulas abaxialmente no glandulares, con pelos de 0,2-0,3 mm, 2-4 celulares o ausentes, escasamente escamosas, las escamas de 1-2 mm, filiformes a linear-lanceoladas, no buladas, adaxialmente con pelos de 0,1-0,5 mm, 3-5 celulares; lámina entre las venas abaxialmente no glandular, glabras, a veces con escamas uniseriadas de 0,2 mm, adpresas, pardas, adaxialmente glabras; venas adaxialmente glabras o con pelos dispersos, visibles, abaxialmente glabras o pubescentes, pelos ca. 0,2 mm, con escasas escamas filiformes, uniseriadas, de 0,2 mm adpresas, pardas; márgenes de la lámina no glandular, escasamente ciliados o aparentemente no ciliados. Soros circulares sin indusios.Esporas crestadas.
Distribución y hábitat. Esta especie se distribuye en Brasil, Paraguay, Uruguay y la Argentina, desde el nivel del mar hasta los 1000 m s.m. En Argentina se encuentra en la provincia de Misiones (Fig. 8), donde es una especie relativamente común. Aunque ha sido citada para la provincia de Corrientes (Hicken, 1908, Ponce, 1996), no se ha podido encontrar material que documente la presencia en la provincia.
Observaciones. Megalastrum connexum se caracteriza por los raquis de las pinnas glabros abaxialmente, por la presencia de escamas filiformes a linear-lanceoladas en la cara abaxial de las cóstulas y venas, y el tejido laminar siempre glabro entre venas. Puede ser confundido con Megalastrum brevipubens R.C. Moran, J. Prado & Labiak, que habita en Paraguay y Brasil (Moran et al., 2009), ambas entidades se diferencian porque esta última posee el tejido laminar pubescente entre las venas en la cara abaxial, con pelos aciculares, erectos, cortos, de aproximadamente 0,1 mm.
Nephrodium connexum (Kaulf.) Hicken, Revista Mus. La Plata, Secc. Bot. 2(15): 231. 1908, es un nombre illegítimo [non Kuhn ex Baker (1870)].
Material representativo examinado
ARGENTINA. Misiones. Depto. Cainguás, 08-I-2003, Márquez 32 (LP, SI). Depto. Candelaria, VII-1927, Burkart 1537 (SI). Depto. General Manuel Belgrano, 260 m, 22-IX-2004, Múlgura 3892 (SI). Depto. Guaraní, interior de selva con helechos arborescentes, hierba, ca. 1 m alt, 15-X-2002, Tressens 6733 (SI). Depto. Iguazú, selva con Euterpe, Meliáceas, Erythrina. Rizoma erecto, hojas fasciculadas, base del pecíolo con escamas, 11-X-1996, Morrone 1184 (SI). Depto. Leandro N. Alem, borde de arroyo, lugar sombría. Hierba terrestre. Frondes inferiores mayores, 27-I-1994, Guillén 310 (SI). Depto. Oberá, VII-1927, Burkart 1585 (SI). Depto. San Pedro, 430 m, 20-XI-2005 Zuloaga 8809 (SI).
Megalastrum crenulans (Fée) A.R. Sm. & R.C. Moran, Amer. Fern J. 77(4): 127. 1987. Aspidium crenulans Feé, Crypt. Vasc. Brésil 1: 139, tab. 47, fig. 1. 1869. Dryopteris crenulans (Feé) C. Chr., Kongel. Danske Vidensk. Selsk. Skr., Naturvidensk. Math. Afd., ser. 8, 6: 90. 1920. Ctenitis crenulata (Feé) Ching, Sunyatsenia 5(4): 250. 1940. TIPO: Brasil, Rio de Janeiro, Rio de Janeiro, A. F. M. Glaziou 1781 (lectotipo P 01631468! aquí designado; isolectotipos BM 000788102!, K, P 00170085!, P 00610618!, RB). Fig. 5.

Fig. 5. Megalastrum crenulans. A, disección de la pinna basal. B, superficie adaxial del raquis y pínnulas. C, superficie adaxial del raquis con detalle de las glándulas estipitadas. D, superficie abaxial del raquis y pínnulas. E, detalle de la superficie abaxial del raquis, con pelos y glándulas estipitadas. F, escama bulada del raquis de la pinna. G, soro, con detalle de las glándulas sobre el indusio. A, de Handro 2224 (US); B-G, de Jürgens 206 (UC).
Dryopteris villosa var. glandulosa Rosenst., Hedwigia 46: 129. 1906. Dryopteris crenulans fo. glandulosa (Rosenst.) C. Chr., Kongel. Danske Vidensk. Selsk. Skr., Naturvidensk. Math. Afd., ser. 8, 6: 91. 1920. TIPO: Brasil, Rio Grande do Sul, Mun. Rio Pardo, banks of Rio Cyriaco, 1906, C. Jürgens s.n. (Herb. Rosenstock Filices Austrobrasilienses no. 206) (lectotipo BM 000907729! designado por R. C. Moran et al., Amer. Fern J. 99: 16. 2009; isolectotipos GH, HB, K, MICH, MO, NY 00678937!, P 00610623!, P 00610624!, P 00610625!, S 06-524!, S 06-525!, S 06-526!, UC 1193512!, UC 1194373!, US 00067057!, UV 001519!).
Rizomas erectos o decumbentes, densamente escamosos, con escamas castañas, linear-lanceoladas, ca. de 2 cm, retorcidas. Frondes hasta de 2,5 m; pecíolos con escamas de la base lineares, escasamente denticuladas, de color castaño claro, retorcidas, formando una densa masa lanosa; láminas de 1 m, 4-pinnado-pinnatífidas en la base, 2-pinado-pinnatífidas en el medio; pinnas basales 30-50 cm, fuertemente inequilaterales, peciólulos hasta de 2,5 cm de largo, pínnulas acroscópicas levemente reducidas hacia la base de la pinna; raquis abaxialmente pubescentes, pelos 0,8-1,2 mm, 4 o 5 celular, densamente glandular, glándulas ca. 0.1 mm, de 2 células, con escamas de 1 mm, lanceoladas, subenteras, planas (no-buladas), densamente pubescente adaxialmente, pelos de 1 mm, 3-6 celulares, no glandulares; cóstulas abaxialmente glandulares, pubescentes, pelos de 0,3-0,4 mm, 1-3 celulares, glándulas ca. 0,1 mm, 2-celulares, con escamas ca. 1 mm, buladas, subenteras, pubescentes adaxialmente, pelos de 0,4-0,8 mm, 1-3 celulares, escasamente glandulares, lámina entre las venas glandular en ambas caras, glándulas más densas abaxialmente, pelos ca. 0,1 mm, unicelulares, solo en la cara abaxial; venas visibles en ambas superficies, glandulares y escasamente pubescentes, márgenes de la lámina densamente ciliados, pelos ca. 0,3-0,4 mm, 1 o 2 celulares. Indusios circulares, notorios, desarrollados, de 0,5-0,7 mm, castaño-oscuros, glandulares y/o pubescentes. Esporas equinadas.
Distribución y hábitat. Esta especie habita entre los 600 a 1760 m s.m desde Venezuela, Paraguay, hasta Brasil y la Argentina, en donde se encuentra en la provincia de Misiones (Fig. 8), creciendo en el sotobosque de la selva paranaense y bosques de Araucaria Juss. (Ponce, 2001).
Observaciones. Megalastrum crenulans se caracteriza por poseer un indusio bien desarrollado, notorio, con pubescencia glandular; también se caracteriza por poseer pelos glandulares en ambas superficies de la lámina.
De acuerdo a Moran et al. (2009), el lectotipo de Megalastrum crenulans fue designado por Christensen (1920) con el espécimen A. F. M. Glaziou 1781. Esta lectotipificación no es efectiva ya que Christensen citó dos herbarios (H, RB); por lo que en este trabajo se designa como lectotipo al ejemplar depositado en P (P 01631468!) que es el espécimen más completo.
Material representativo examinado
ARGENTINA. Misiones. Depto. San Pedro, Parque Provincial Cruce Caballero, 690 m, 15-IV-1996, Zuloaga 5551 (CTES, SI).
Megalastrum fugaceum R.C. Moran, J. Prado & Sundue, Amer. Fern J. 104(3): 158. 2014. TIPO: Bolivia, Santa Cruz, Florida, Cantón Mairana, Parque Nacional Amboró, 7 km NNE by air from Mairana, 900–2200 m, 23-VII-1994, R.C. Moran 5911 (holotipo LPB!; isotipos AAU!, NY!, UC-1604363!, UC-1604364!). Fig. 6.

Fig. 6. Megalastrum fugaceum. A, superficie adaxial de pínnulas y ejes. B, superficie abaxial de pínnulas y ejes. C, escama de la cóstula con detalle del ápice más oscuro. D, superficie abaxial de un último segmento. E, superficie adaxial de las pínulas y ejes. F, superficie abaxial de pínulas y ejes. G, escamas de la cóstula. A-D, de Moran 5911 (UC); E-G, de Øllgaard et al. 99556 (AAU).
Rizomas erectos, con escamas de 20-35 mm, lineares, ascendentes, castañas, ligeramente brillantes, planas o algo retorcidas, no tortuosas, escasamente a retrorso-denticuladas; frondes hasta de 4 m; pecíolos con escamas en la base como las del rizoma, a veces formando una masa lanosa; láminas 1-3 m, basalmente 3-4-pinnado-pinnatisectas, medias 3-pinnado-pinnatisectas; raquis abaxialmente eglandular, glabro a escasamente pubescente, pelos (cuando presente) 0,4-0,8 mm, 3-5 celulares, con pocas escamas de 0,1-1,8 mm, ovadas a lanceoladas, adpresas o erguidas, castaño claras a oscuras, fuertemente denticuladas, no clatradas, no buladas, adaxialmente eglandular, densamente pubescente, pelos de 0,3-0,5 mm, 3-6 celulares, con escamas dispersas, de 0,5-3 mm, linear-lanceoladas, oscurecidas hacia el ápice, fuertemente denticuladas; pinnas basales ca. 1,4 m, fuertemente no equilaterales; pínnulas basiscópicas basales de las pinnas medias no elongadas ni superpuestas sobre el raquis; cóstulas con indumento similar al del raquis, pero las escamas más grandes, de 0,5-2,5 mm, ovadas a lineares, oscurecidas hacia el ápice, denticuladas, ascendentes; lámina entre las venas en general glabra, a veces pubescente pero eglandular en ambas caras, los pelos (cuando presentes) 0,2-0,4 mm, patentes; venas visibles en ambas superficies de lámina, eglandulares, abaxialmente subglabras a moderadamente pubescentes, los pelos (cuando presentes) 0,2-0,4 mm, patentes, adaxialmente cuando pubescentes los pelos 0,5-0,7 mm, estrigosos a laxamente ascendentes; márgenes de la lámina ciliados. Indusio inconspicuo y caduco, de 0,1-0,2 mm de diám, aparentando un grupo de protoescamas rojizas, ca. 0,1 mm, que sobresalen desde el centro de los soros (no se observa fácilmente en soros maduros). Esporas equinadas.
Distribución y hábitat. Esta especie se distribuye desde Ecuador hasta Bolivia y Argentina en las provincias de Jujuy, Salta y Tucumán (Fig. 8), donde habita lugares muy húmedos del sotobosque de las Yungas y orillas de curso de agua entre los 1060-2300(-3500) m s.m.
Observaciones. Megalastrum fugaceum es una de las especies de helechos no arborescentes que alcanzan el mayor tamaño en la Argentina. Es similar a M. pulverulentum (Poir.) A.R. Sm. & R.C. Moran, especie excluida de la flora Argentina en este trabajo (ver sección de especies excluidas); con la cual ha sido confundida porque ambas especies poseen hojas grandes hasta de 4 m y escamas de los raquis de las pinnas y cóstulas, generalmente oscuras apicalmente y de margen fuertemente denticulado; sin embargo M. fugaceum es mucho menos pubescente y variable, con los pelos más cortos (Fig. 6 A), o con la lámina normalmente glabra entre venas en la cara abaxial (Fig. 6 E). Esta especie fue tratada para Bolivia como M. pulverulentum en la flora del Parque Nacional Amboró (Sundue, 2011), lugar de donde proviene el ejemplar tipo de M. fugaceum.
Material representativo examinado
ARGENTINA. Jujuy. Depto. Ledesma, Iudica 352 (SI). Parque Nacional Calilegua, hierba de 1.5–2 m de alt., 20-II-1998, Morrone et al. 2863 (SI). Parque Nacional Calilegua, Ruta Provincial 83, Aguada del Tigre, 1600 m, 7-IV-2010, Martínez & Prado 1894 (MCSN, NY, SP). Ídem, Larsen & Arana s.n. (RCVC 5730, SI). Depto. Dr. Manuel Belgrano, 1710 m.s.m., 19-XI-2002, Zuloaga 7844 (SI). Salta. Depto. La Caldera, Ruta Nac. Nº 9, Km 1500, 1400 m, 16-XII-1995, Martínez 149 (MCSN). Tucumán. Depto. Famaillá, quebrada del Caspichango, 11-VII-1911, Castillón s.n. (CTES 150721, LIL 40958). Depto. Monteros, Quebrada Pueblo Viejo, I-1965, de la Sota 4069 (LP, US). Depto. Tafí, Frondes de 2-2,5 m de longitud, en quebradas húmedas, 100 m, 02-IX-1957, de la Sota 1662 (LP).
Megalastrum spectabile (Kaulf.) A.R. Sm. & R.C. Moran, Amer. Fern J. 77(4): 129. 1987. Polypodium spectabile Kaulf., Enum. Fil.: 121. 1824. Phegopteris spectabilis (Kaulf.) Fée, Mém. Foug. 5, Gen. Filic. 243. 1852. Dryopteris spectabilis (Kaulf.) Macloskie & Dusén, Rep. Princeton Univ. Exped. Patagonia, Botany 8, Suppl.: 10. 1915. Ctenitis spectabilis (Kaulf.) G. Kunkel, Nova Hedwigia 13(3-4): 333. 1967. TIPO: Chile, sine data, 1827, L. A. von Chamisso s.n. (lectotipo LE 00000057! aquí designado; isolectotipo P 00610839!). Fig. 7.
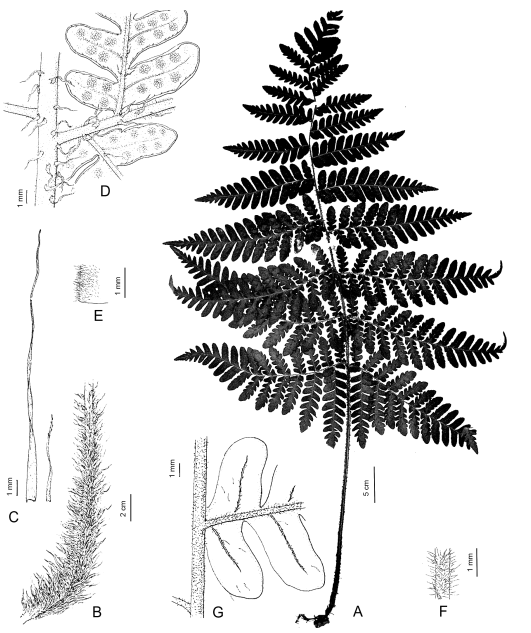
Fig. 7. Megalastrum spectabile. A, fronde. B, porción basal del pecíolo. C, escamas de porción basal del pecíolo. D, superficie abaxial del raquis y pinnas, con detalle de la inserción en el raquis de la lámina. E, detalle de los pelos en la superficie abaxial del raquis de la lámina. F, detalle de los pelos y escamas en la superficie abaxial de la costa de la pinna. G, superficie adaxial del raquis de la pinna y pínnula. A, de Skottsberg 921 (S); B-C, de Dusén s.n. (NY); D-G, de Solomon & Solomon 4559 (MO, NY).
Polypodium contractum Desv., Mém. Soc. Linn. Paris 6: 242. 1827. TIPO: Chile, habitat in regno Chilense, sin colector s.n., Herb. A. N. Desvaux s.n. (holotipo P 00610840!).
Dryopteris spectabilis var. philippiana C. Chr., Kongel. Danske Vidensk. Selsk. Naturvidensk. Math. Afd., ser. 8, 6: 70. 1920. Ctenitis spectabilis var. philippiana (C. Chr.) R.A. Rodr., Bol. Soc. Biol. Concepción 45: 148. 1972. Megalastrum spectabile var. philippianum (C. Chr.) A.R. Sm. & R.C. Moran, Amer. Fern J. 77(4): 129. 1987. TIPO: Chile, Valdivia, H. Krause s.n. (lectotipo, NY 01185562!, designado por M. Sundue et al., Syst. Bot. 35: 472. 2010).
Rizomas erectos aunque sin formar troncos, con escamas subuladas, de 5-20 mm, castaño o castaño verdosas, enteras. Frondes de 0,5-2 m, fasciculadas; pecíolos con escamas de 1-2,2 cm, linear-lanceoladas a triangulares, clatradas, castaño doradas, diminutamente denticuladas, ápices filiformes, algo tortuosos; láminas 2-pinado-pinnatisectas a 3-pinado-pinnatífidas en la base, 35-60 cm; pinnas basales (12-)22-28(-52) cm, no equilaterales; pínnulas cortamente pecioluladas a sésiles, las pínnulas mayores de las pinnas basales de 9-11 cm; raquis y costas escasamente glandulares, densamente pubescentes y moderadamente escamosos tanto abaxial como adaxialmente, las glándulas ca. 0,1 mm, sésiles, hialinas, amarillentas, los pelos de 0,3-0,5 mm, (1-)2-5 células, aciculares, hialinos, esparcidos, escamas 0,8-3,5 × 0,2-0,6 mm, lanceoladas, castañas, lustrosas, margen eroso, adaxialmente pubescentes; pínnulas largamente triangulares lobuladas o levemente crenadas, lámina entre venas abaxialmente glabra a puberulenta, con pelos erectos de 0,3 mm, 3-4 celulares y protoescamas uniseriadas, adpresas, rojizas, adaxialmente glabra a moderadamente pubescente, con pelos de 0,3 mm, 3-4 celulares; venas visibles en el lado abaxial de la lámina, abaxialmente pubescente y con pelos glandulares, pelos de 0,2-0,7 mm, glándulas sésiles, amarillas, hialinas, adaxialmente pubescentes con pelos similares a los de la cara abaxial, margen de la lámina ciliado. Soros circulares, sin indusio, medianos o submarginales. Esporas equinadas.
Distribución y hábitat. Esta especie es endémica del sur de Chile y Argentina en las provincias de Chubut, Neuquén y Río Negro (Fig. 8), restringida a la selva templada Valdiviana y bosques Andino-patagónicos, entre los 10-350(-500) m s.m., donde se encuentran ejemplares solitarios a la orilla de arroyos y lugares muy húmedos.
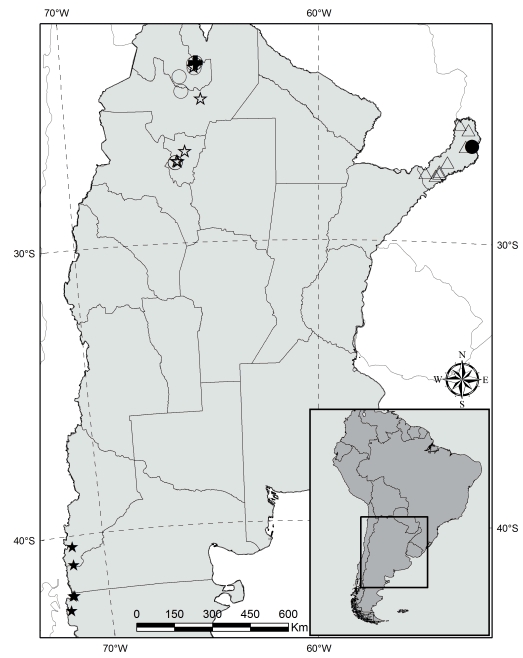
Fig. 8. Mapa de distribución de las especies de Megalastrum en Argentina, (estrella vacia) M. adenopteris, (cruz) M. ciliatum, (estrella rellena) M. spectabile, (círculo vacío) M. fugaceum, (círculo relleno) M. crenulans, (triángulo) M. connexum.
Observaciones. La pubescencia es muy variable en cuanto a la longitud y distribución de los pelos, con especímenes casi glabros hasta puberulentos en el tejido laminar entre venas de la cara abaxial. La longitud de los pelos en ambas caras de la hoja sobre las venas también es variable, desde 0,2 hasta 0,7 mm, las plantas que poseen los pelos más largos han sido reconocidas previamente como M. spectabile var. philippiana, pero este carácter se evidencia como continuo y no permite diferenciar taxones subespecíficos.
De acuerdo con Sundue et al. (2010) el lectotipo de Polypodium spectabile fue designado por Christensen (1920), aunque en el mencionado trabajo no se expresa específicamente en cual herbario estaba depositado el tipo (L. A. von Chamisso s.n.). Aquí se designa al ejemplar L. A. von Chamisso s.n. depositado en LE (LE 00000057!) como lectotipo de la especie, ya que representa mejor las características de la especie al poseer mayor cantidad de pinnas maduras queel ejemplar en P (P00610839!).
Dryopteris spectabilis (Kaulf.) C. Chr., Kongel. Dankse Vidensk. Selsk. Skr. Naturvidensk. Math. Afd., ser. 8, 6: 69. 1920, constituye una combinatio superflua debido a la combinación propuesta con anterioridad por Macloskie & Dusén.
Material representativo examinado
ARGENTINA. Chubut: Depto. Cushamen, Lago Puelo, 2-II-1945, Castellanos 114252 (NY). Depto Futaleufú, brazo norte Lago Menéndez, Castellanos s.n. (LIL 149440, SI); Arroyo del Fuinque, entre Gendarmería y Los Hitos, 360 m s. n. m. 28-IV-1999, Vidoz 133 (LP). Neuquén: Depto. Los Lagos, Lago Espejo, 40°37’S, 71°45’W, 20-III-1939, Cabrera 5037 (GH, LP); puerto Manzano, Lago Nahuel Huapi, 41°0’S, 71°30’W, 12-III-1963, de la Sota 2799 (LP, MICH). Río Negro: Depto. Bariloche, Bariloche, Puerto Blest Area, west end of Lago Nahuel Huapi, 41°0’S, 71°30’W, 5-XI-1982, Sage s.n. (UC).
ESPECIES EXCLUIDAS DE LA FLORA DE ARGENTINA
Megalastrum pulverulentum (Poir.) A.R. Sm. & R.C. Moran, Amer. Fern J. 77(4): 129. 1987. Polypodium pulverulentum Poir., Encycl. Méth. Bot. 5: 555. 1804. Aspidium pulverulentum (Poir.) Desv., Mém. Soc. Linn. Paris 6: 251. 1827. Dryopteris pulverulenta (Poir.) C. Chr. in Urb., Symb. Antill. 9: 305. 1925. Ctenitis pulverulenta (Poir.) Copel., Gen. Fil. (Copeland): 124. 1947. TIPO: La Española, “Insulam Sandominicanam”. “Filix hirsuta lutescente pulvisculo sordescens” en C. Plumier, Traité Foug. Amér. 27, pl. 34!. 1705 (lectotipo designado por G. R. Proctor, Ferns of Jamaica: 423. 1985).
Esta especie, citada para Argentina en las provincias de Salta y Jujuy (de la Sota, 1977; Smith & Moran, 1987; Ponce & Martínez, 2012, Ganem et al., 2013), es una de las especies de Megalastrum de mayor tamaño, con hojas de más de 4 m y pinnas basales de hasta 1,4 m. Se caracteriza por poseer la lámina densamente pubescente en ambas superficies, con pelos blanquecinos de 1-2 mm (más largos que los de M. fugaceum, que alcanzan 0,4-0,8 mm) y pelos glandulares amarillentos sésiles o cortamente pedicelados. Además M. pulverulentum es densamente pubescente en el tejido laminar entre venas abaxialmente, mientras que en M. fugaceum es glabro o con muy escasos pelos dispersos. Las escamas del raquis de las pinnas son similares a las de M. fugaceum, de margen dentado y ápice oscuro. Todo el material citado para la Argentina como M. pulverulentum corresponde a M. fugaceum. Megalastrum pulverulentum se distribuye por México, Guatemala, Honduras (Stolze, 1981), Costa Rica, Panamá, Jamaica, Haití, República Dominicana, Guadalupe, Martinica, Dominica, Venezuela, Colombia, Ecuador y Perú, entre los 1200-2500 (-3200) m s.m., siendo este último país el límite austral de distribución de la especie (Moran et al., 2014b).
Observaciones. Aspidium lutescens Willd., Sp. Pl. 5: 272. 1810, constituye un nomen superfluum dado por Willdenow a Polypodium pulverulentum.
Megalastrum subincisum (Willd.) A. R. Sm. & R. C. Moran, Amer. Fern J. 77(4): 129. 1987 [publicado el 3 de mayo de 1988]. Polypodium subincisum Willd., Sp. Pl., ed. 4, 5: 202. 1810. Phegopteris subincisa (Willd.) Fée, Mém. Foug., 5 Gen. Filic.: 243. 1852. Nephrodium villosum (L.) C.Presl var. subincisum (Willd.) Baker, Fl. Bras. (Martius) 1(2): 483. 1870. Dryopteris subincisa (Willd.) Urb., Symb. Antill. 4: 19. 1903. Aspidium subincisum (Willd.) Christ, Bull. Herb. Boissier, sér. 2, 6(1): 56. 1906. Ctenitis subincisa (Willd.) Ching, Sunyatsenia 5(4): 250. 1940. TIPO: Venezuela, Distrito Federal, Caracas, s.d., F. Bredemeyer s.n. (holotipo B-W-19701!).
De acuerdo con de la Sota et al. (1998), esta especie fue citada erróneamente para la Patagonia argentina por Hicken (1909), confundida con Megalastrum spectabile. Megalastrum subincisum se caracteriza por poseer el raquis de las pinnas sin pelos y con escamas linear-lanceoladas de color marrón dorado. Habita en México, Honduras, Cuba, Jamaica, Haití, República Dominicana, Guadalupe, Dominica, el norte de Venezuela y Colombia, entre los 450-3000 m s.m. (Moran et al., 2014b).
Observaciones. Nephrodium villosum (L.) C. Presl var. subincisum (Willd.) Hook. ex Sodiro, Rescens. Crypt. Vasc. Quit.: 54. 1883, y Nephrodium villosum (L.) C. Presl var. subincisum (Willd.) Jenman, Bull. Bot. Dept. Jamaica n.s. 3:114. 1896 constituyen combinatio superflua, posteriores a la propuesta por Baker.
AGRADECIMIENTOS
Los autores desean agradecer a los curadores y personal de los herbarios visitados por su amable atención y predisposición. Un agradecimiento muy especial para Robbin Moran (NY) y Michael Sundue (VT) por permitir la utilización de las imágenes reproducidas; a Nataly O’Leary y dos revisores anónimos, cuyos comentarios contribuyeron a mejorar este artículo y Cristian Larsen y Juan Angeli por el mapa de distribución. Este trabajo se realizó en el marco del proyecto Flora Argentina (http://www.floraargentina.edu.ar), financiado por PIP 537-CONICET.
BIBLIOGRAFÍA
1. Christensen, C. 1913. A monograph of the genus Dryopteris, Part I. The tropical American pinnatifid-bipinnatifid species. Kongelige Danske videnskabernes selskabs skrifter. Naturvidenskabelig og mathematisk afd; 7. raekke, X, 2. 10: 55-282. [ Links ]
2. Christensen, C. 1920. A monograph of the genus Dryopteris, Part II. The tropical American bipinnate-decompound species. Kongelige Danske Videnskabernes Sselskabs Skrifter. Naturvidenskabelige og Mathematiske Afdeling, ser. 8(6): 3-132. [ Links ]
3. Christensen, C. 1934. Index Filicum Supplementum Tertium pro annis 1917–1933. Copenhagen: H. Hagerup.
4. de la Sota, E. R. 1977. Pteridophyta, en A. L. Cabrera (ed.), Flora de la Provincia de Jujuy. Colección Científica del Instituto Nacional de Tecnología Agropecuaria 13(2): 1-275. [ Links ]
5. de la Sota, E. R., M. M. Ponce, M. A. Morbelli, & L. A. Cassá de Pazos. 1998. Pteridophyta, en M. N. Correa (ed.), Flora Patagónica. Colección Científica del Instituto Nacional de Tecnología Agropecuaria 8(1): 252-369. [ Links ]
6. Ganem, M. A.; J. P. Ramos Giacosa, M. L. Luna, M. D. Arana, A. Rotman, O. Ahumada, E. R. de la Sota & G. E. Giudice. 2013. Diversidad de helechos y licofitas del Parque Nacional Calilegua, Provincia de Jujuy, Argentina. Boletín de la Sociedad Argentina de Botánica 48: 567-584. [ Links ]
7. Hicken, C. M. 1908. Polypodiacearum Argentinarum Catalogus. Revista del Museo de La Plata, sección Botánica 15: 226-282. [ Links ]
8. Hicken, C. M. 1909. Clave artificial de las Aspidiéas argentinas. Apuntes Historia Natural 1 (10-11): 145-154. [ Links ]
9. Holttum, R. E. 1986. Studies in the fern-genera allied to Tectaria Cav. VI, a conspectus of genera in the Old World regarded as related to Tectaria, with descriptions of two genera. The Gardens’ Bulletin Singapore 39(2): 153-167.
10. Kessler, M. & A. R. Smith. 2006. Megalastrum (Dryopteridaceae: Pteridophyta) in Bolivia, with descriptions of six new species. American Fern Journal 96(1): 31-44. [ Links ]
11. Labiak, P.H.; M. Sundue, G. Rouhan, J. Garrison Hanks, J. T. Mickel & R. C. Moran. 2014. Phylogeny and historical biogeography of the lastreopsid ferns (Dryopteridaceae). American Journal of Botany 101(7): 1207-1228. DOI: 10.3732/ajb.1400071 [ Links ]
12. Labiak, P. H.; M. A. Sundue, G. Rouhan & R. C. Moran. 2015a. New combinations in Lastreopsis and Parapolystichum (Dryopteridaceae). Brittonia 67(1): 79-86. DOI: 10.1007/s12228-014-9531-3 [ Links ]
13. Labiak, P. H.; M. A. Sundue, G. Rouhan & R. C. Moran. 2015b. Rhopalotricha, a new subgenus of the fern genus Lastreopsis (Dryopteridaceae). American Fern Journal 105(1): 20-30. [ Links ]
14. Mickel, J. T. & J. M. Beitel. 1988. Pteridophyte Flora of Oaxaca, Mexico. Memoirs of the New York Botanical Garden 46: 128-135. [ Links ]
15. Moran, R. C. & J. Prado. 2010. Megalastrum (Dryopteridaceae) in Central America. Kew Bulletin 65(2): 137-188. [ Links ]
16. Moran, R. C.; J. Prado & P. H. Labiak. 2009. Megalastrum (Dryopteridaceae) in Brazil, Paraguay, and Uruguay. American Fern Journal 99(1): 1-44. [ Links ]
17. Moran R. C.; J. Prado & M. A. Sundue 2014a. Megalastrum (Dryopteridaceae) in Andean South America, Part I. American Fern Journal 104(3): 109-178. [ Links ]
18. Moran R. C.; J. Prado & M.A. Sundue 2014b. Megalastrum (Dryopteridaceae) in Andean South America, Part II. American Fern Journal 104(4): 181-236. [ Links ]
19. Ponce, M. M. 1996. Pteridophyta, en F. O. Zuloaga & O. Morrone (eds.), Catálogo de las plantas vasculares de la República Argentina I: Pteridophyta, Gymnospermae y Angiospermae (Monocotyledoneae). Monographs in Systematic Botany from the Missouri Botanical Garden 60: 1-79. [ Links ]
20. Ponce, M. M. 2001. Additions and Corrections to the Pteridophyte Flora of Northeastern Argentina. American Fern Journal 91(2): 70-72. [ Links ]
21. Ponce, M. M. & O. G. Martinez. 2012. Dryopteridaceae, en L. J. Novara (ed.), Flora del Valle de Lerma. Aportes Botánicos de Salta, serie Flora 11(8): 1-31. [ Links ]
22. Ponce, M. M.; K. Mehltreter & E. R. de la Sota. 2002. Análisis biogeográfico de la diversidad pteridofítica en Argentina y Chile continental. Revista Chilena de Historia Natural 75(4): 703-717. [ Links ]
23. Proctor, G. R. 1989. Ferns of the Puerto Rico and the Virgin Islands. Memoirs of the New York Botanical Garden 53: 248-249. [ Links ]
24. Rouhan, G. & R. C. Moran. 2011. Revision of Paleotropical Megalastrum (Dryopteridaceae). Annals of the Missouri Botanical Garden 98(1): 90-100. [ Links ]
25. Schuettpelz, E. H. & K. M. Pryer. 2007. Phylogeny of ferns inferred from 400 leptosporangiate species and three plastid genes. Taxon 56(4): 1037-1050. [ Links ]
26. Schuettpelz, E. H. & K. M. Pryer. 2009. Evidence for a Cenozoic radiation of ferns in an angiosperm-dominated canopy. Proceedings of the National Academy of Sciences of the United States of America 106(27): 11200-11205. [ Links ]
27. Smith, A. R. & R. C. Moran. 1987. New combinations in Megalastrum (Dryopteridaceae). American Fern Journal 77(4): 124-130. DOI: 10.2307/1547480 [ Links ]
28. Stolze, R. G. 1981. Ferns and fern allies of Guatemala. Part II. Polypodiaceae. Fieldiana Botany new series 6: 143-162. [ Links ]
29. Sundue, M. A. 2011. Licofitas y helechos de la región del Parque Nacional Amboró, Bolivia, en M. H. Nee, (ed.), Flora de la Región del Parque Nacional Amboró - Bolivia, vol. 1, pp. 37-368. Santa Cruz de la Sierra: Fundación Amigos de la Naturaleza. [ Links ]
30. Sundue, M. A.; G. Rouhan & R. C. Moran. 2010. Megalastrum (Dryopteridaceae) of the Circumaustral region: Chile, Argentina, and Southern Islands of the Atlantic, Pacific, and Indian Oceans. Systematic Botany 35(3): 461-475. [ Links ]
31. Thiers, B. [permanentemente actualizado, consulta 2016] Index Herbariorum: A global directory of public herbaria and associated staff. New York Botanical Garden’s Virtual Herbarium. http://sweetgum.nybg.org/ih/
32. Tindale, M. D. 1965. A monograph of the genus Lastreopsis Ching. Contributions of the New South Wales National Herbarium 3(5): 249-339. [ Links ]

