











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
Las estrategias de conservación ex situ constituyen el conjunto de acciones que se desarrollan fuera de los hábitats naturales y permiten preservar un amplio número de especies, en un espacio relativamente reducido (Mounce et al., 2017). Estas acciones tienen como objetivo resguardar la diversidad genética, principalmente de especies amenazadas, endémicas y con valor de uso, a corto, mediano y largo plazo. Entre ellas, los bancos de semillas se destacan como la estrategia más comúnmente usada para este propósito (Hay, 2021).
Para conservar una especie en bancos de semillas, es necesario conocer su tolerancia a la desecación y su comportamiento durante el almacenamiento a diferentes condiciones de humedad y temperatura. Con base en su comportamiento, las especies pueden ser clasificadas en ortodoxas, intermedias y recalcitrantes. Las semillas ortodoxas toleran el secado a bajos niveles de humedad (3-7 %) y el almacenamiento a bajas temperaturas (< 0 °C). Las intermedias pueden desecarse, aunque no a tan bajos niveles como las ortodoxas y con frecuencia son sensibles al frío. Por último, las semillas recalcitrantes son muy sensibles a la desecación y al frío (Hong & Ellis, 1996; Pritchard et al., 2014). El tipo de comportamiento también puede predecirse de algunas características de las especies, tales como hábitat, linaje al que pertenecen y características morfo-fisiológicas de la semilla (contenido de humedad, tamaño, forma y espesor de la cubierta, entre otros; Pelissari et al., 2018; Junio-Ramos et al., 2019). Si bien, esto último puede ser de utilidad ante la ausencia total de información, es necesario evaluar la tolerancia a la desecación en cada especie objeto de conservación, ya que, de ser clasificadas erróneamente, fracasará su preservación ex situ (especialmente en especies recalcitrantes; Pritchard et al., 2014).
Una vez establecido que una especie puede conservarse en bancos de semillas (comportamiento ortodoxo), se realizan monitoreos periódicos (por ejemplo, cada 10 años) para garantizar que la viabilidad de las semillas se mantenga durante el almacenamiento, y de ser necesario, poder regenerarlas a tiempo (especies anuales) o renovar las colecciones con nuevas muestras (especies perennes; Walters, 2015). Si bien la frecuencia de monitoreo se desarrolla con base en el amplio conocimiento que se tiene de especies cultivadas, se considera que dichos monitoreos son dependientes de la longevidad de la semilla, es decir el tiempo que la misma puede mantenerse viva. Se han propuesto distintos métodos para evaluar la longevidad potencial de las semillas (Hay & Probert, 2013; Walters, 2015). Entre ellos, el envejecimiento acelerado es un método relativamente sencillo que permite estimar el tiempo requerido para que la viabilidad disminuya al 50 % (P50). Este valor permite categorizar la esperanza de vida de las semillas, comparándolas con especies de longevidad conocida (Newton et al., 2014). Se ha indicado que las familias con mayor longevidad son Anacardiaceae, Fabaceae, Cistaceae y Malvaceae, asociando dicha característica con la presencia de dormición física (Pritchard & Dickie, 2003).
Numerosos autores han destacado que, a pesar de su importancia, las especies nativas con valor de uso están poco representadas en las colecciones ex situ (Walters, 2015; Curti et al., 2017; Hanson & Ellis, 2020), por lo que revertir esta situación es de suma prioridad. Para ello, es necesario, entre otros aspectos, conocer su comportamiento durante el almacenamiento y los determinantes de su germinación (respuesta a la luz, tipo de dormición, entre otras), teniendo en cuenta que puede existir variación intraespecífica en la respuesta a las condiciones a las que son sometidas las semillas (Galíndez et al., 2019; Hanson & Ellis, 2020).
En Argentina, el noroeste del país es una de las regiones más diversas, con una gran heterogeneidad de paisajes (Juárez et al., 2007). En la región, se observan ocho unidades de vegetación entre las cuales están representados dos bosques subtropicales estacionales con precipitaciones contrastantes. Por un lado, las selvas y bosques montanos pertenecientes a la Provincia fitogeográfica de las Yungas, de clima cálido, húmedo a subhúmedo, con una precipitación media anual entre 600 y 3000 mm. Por otro lado, los bosques xerófitos de la Provincia Fitogeográfica Chaqueña, de clima continental, cálido subtropical seco con una precipitación media anual entre 400 y 700 mm (Cabrera, 1976; Oyarzabal et al., 2018). En estos ambientes, muchas especies tienen valor de uso como alimenticias, forrajeras, medicinales, maderables, entre otras aplicaciones. Sin embargo, el cambio en el uso del suelo por expansión de la frontera agropecuaria, en detrimento de la superficie ocupada por el bosque nativo (Volante et al., 2016), sumada a la sobreexplotación con fines maderables, han llevado a que muchas especies de árboles nativos se encuentren amenazados. Tal es el caso de especies del género Amburana, Cedrela, Gonopterodendron, Jacaranda, entre otras (Americas Regional Workshop, 1998; Barstow, 2018; Hills, 2020; 2021; IUCN, 2023).
En este contexto, el propósito del presente trabajo fue evaluar si las semillas de cuatro especies nativas de importancia forestal, Handroanthus impetiginosus (Mart. ex DC.) Mattos, Jacaranda mimosifolia D. Don, Neltuma alba (Griseb.) C.E. Hughes & G.P. Lewis y Pterogyne nitens Tul., que crecen en el noroeste de Argentina, pueden ser incorporadas a bancos de semillas como una estrategia de conservación ex situ. Estas poblaciones no han sido evaluadas previamente y habitan una región donde ya se han registrado otras especies de árboles nativos con variación inter poblacional en el comportamiento de sus semillas durante el almacenamiento. Para cada especie, se estudió la tolerancia a la desecación y el comportamiento durante el almacenamiento. En adición, se evaluó la longevidad de las semillas a través de un ensayo de envejecimiento acelerado. Con base en estudios previos y lo informado por las bases de datos SID KEW (SER, INSR, RBGK, 2023) y GENESYS (USDA, 2023), esperamos que las cuatro especies se comporten como ortodoxas y, con base en el linaje al que pertenecen las especies, esperamos que la familia Fabaceae presente mayor longevidad debido a la presencia de dormición física.
MATERIALES Y MÉTODOS
Especies estudiadas
Se estudiaron cuatro especies de importancia forestal que habitan en el noroeste de Argentina: Handroanthus impetiginosus y Jacaranda mimosifolia, pertenecientes a la familia Bignoniaceae y Neltuma alba (= Prosopis alba Griseb. var. alba) y Pterogyne nitens, pertenecientes a la familia Fabaceae. La fructificación de N. alba ocurre entre los meses de diciembre y enero, mientras que en P. nitens se observa entre septiembre y noviembre, aunque la cosecha de frutos maduros se realiza a partir de diciembre. La dispersión natural de H. impetiginosus se observa entre octubre y diciembre, mientras que la de J. mimosifolia entre julio y septiembre (Martínez, 2006).
Recolección de los frutos y las semillas
Durante el período de dispersión natural de cada especie, se cosecharon frutos (y/o semillas) maduros de entre cinco y diez individuos, dependiendo del número de individuos sanos por sitio (Tabla 1). Los frutos maduros de N. alba y P. nitens se colectaron directamente de los árboles, mientras que en el caso de H. impetiginosus y J. mimosifolia, se emplearon trampas de semillas. El material colectado se llevó al laboratorio para su limpieza y posterior caracterización de las semillas.
Caracterización de las semillas
Se determinó, para cada especie, peso de la semilla, contenido de humedad (CH), viabilidad de las semillas y capacidad germinativa. El peso promedio de la semilla se obtuvo pesando, individualmente, 25 semillas en una balanza analítica de precisión 0,1 mg. El CH se determinó gravimétricamente, secando tres réplicas de diez semillas cada una, en estufa a 103 °C ± 2 °C, durante 17 hs (ISTA, 2016). La viabilidad inicial de las semillas (V ) se evaluó con cuatro réplicas de 25 semillas cada una, empleando la prueba de tetrazolio (ISTA, 2016). La capacidad germinativa se evaluó sembrando cuatro réplicas de 25 semillas cada una, en cajas de Petri sobre papel de filtro humedecido con agua destilada, garantizando la disponibilidad de agua durante todo el experimento (Davies et al., 2015). Las semillas sembradas se incubaron en cámaras de germinación a 25 °C, bajo luz blanca (30 μmol m-2 s-1; 400-700 nm de luz blanca fluorescente fría) con un fotoperiodo de 8/16 hs luz-oscuridad. El número de semillas germinadas se registró diariamente por un período de 30 días, considerando como criterio de germinación la emergencia de la radícula (> 1 mm; Ortega-Baes et al., 2011; Galíndez et al., 2019). Al finalizar el experimento, las semillas no germinadas se cortaron longitudinalmente con un bisturí y sus embriones se observaron en microscopio óptico. La presencia de embriones blancos y turgentes fueron el criterio para considerar a una semilla viable (Ortega-Baes et al., 2010). En las semillas de N. alba y P. nitens se determinó la presencia de dormición física mediante curvas de imbibición (datos no publicados), por lo que, siempre que se evaluó su capacidad germinativa se sembraron semillas intactas y escarificadas mecánicamente (con un corte en la región opuesta al micrópilo). La variable de respuesta fue el porcentaje de semillas germinadas.
Comportamiento durante el almacenamiento
El comportamiento de las semillas durante el almacenamiento (ortodoxas, intermedias o recalcitrantes) se determinó siguiendo la metodología propuesta por Hong & Ellis (1996). Primeramente, una muestra de 130 semillas por especie se colocó en un desecador que simulaba una humedad relativa en el ambiente (HR) de 60 %, empleando la sal CaNO3. Al cabo de tres semanas, cuando las semillas equilibraron su CH (entre un 7 y 9 % CH, dependiendo de la especie), se evaluó su capacidad germinativa, como fue anteriormente descripto. Una vez corroborado que las semillas sobreviven al secado a 60 % HR (ver resultados), nuevas muestras de 130 semillas (12 para las especies de Bignoniaceae y 24 para las especies de Fabaceae) se colocaron en desecadores en los que se generó una HR de 15 %, empleando la sal LiCl. Al cabo de tres semanas, cuando las semillas equilibraron su CH (entre un 4 y 5 % CH, dependiendo de la especie), se evaluó su capacidad germinativa. Las semillas desecadas a 15% HR se preservaron en bolsas de aluminio trilaminadas y se almacenaron a -20 °C, 5 °C y 20 °C, durante 3, 6 y 12 meses, tiempos en los que se evaluó la capacidad germinativa.
Longevidad potencial de las semillas
Dado que todas las semillas se comportaron como ortodoxas (ver resultados), la longevidad potencial se determinó para las cuatro especies, siguiendo la metodología de envejecimiento acelerado propuesta por Newton et al. (2014). Para ello, 12 muestras de 100 semillas por especie se colocaron en desecadores con sal de CaNO3 para simular 60 % HR, a una temperatura constante de 45 °C. Estas condiciones fueron controladas, durante todo el experimento, mediante un registrador de datos (“data logger” HOBO UX 100). En intervalos de tiempo, cuatro réplicas de 25 semillas se extrajeron de los desecadores para evaluar su capacidad germinativa, hasta que ninguna semilla germinó. En primera instancia, los intervalos de tiempo se planificaron siguiendo el protocolo estándar de Probert et al. (2009) en el que las semillas se removerían después de 0, 1, 2, 5, 9, 15, 20, 30, 50, 75, 100 y 125 días. Sin embargo, estos intervalos de tiempo se fueron modificando, dependiendo del último registro de germinación. Así, especies que disminuían su capacidad germinativa con mayor velocidad se evaluaron antes del intervalo planificado (H. impetiginosus y J. mimosifolia), mientras que especies en las que la capacidad germinativa se mantenía constante fueron ampliando sus intervalos de tiempo (N. alba y P. nitens). Estos ajustes se realizaron para obtener el mayor número posible de intervalos que incluyan la disminución significativa de la germinación (ver Newton et al., 2014). Las semillas de N. alba y P. nitens se envejecieron intactas y escarificadas mecánicamente para evaluar el efecto de la cubierta dura impermeable sobre la longevidad de las semillas. Los porcentajes de germinación a lo largo del tiempo se graficaron en curvas de supervivencia de las semillas, que se ajustaron mediante un análisis Probit (Hay et al., 2014), empleando la ecuación de viabilidad descripta por Ellis & Roberts (1980):
v = K i - (p/σ), (1)
donde ν es la viabilidad de las semillas (en desviaciones equivalentes normales; NED) en un tiempo p (días), K i es una medida de la viabilidad inicial (NED) y σ es la desviación estándar de la distribución normal que describe la frecuencia de semillas muertas en el tiempo p de envejecimiento. El P50 o tiempo requerido para que la viabilidad disminuya al 50 % se calculó mediante la ecuación:
P 50 = K i × σ (2)
Las especies se clasificaron en semillas de vida muy corta (P50 ≤ 2 días), vida corta (P50 entre 2 y 10 días), vida intermedia (P50 entre 10 y 100 días), vida larga (P50 entre 100 y 1000 días) y vida muy larga (P50 ≥ 1000 días). Las categorías se basaron en lo establecido por Probert et al. (2009), ajustado por Mondoni et al. (2011) y Davies et al. (2020), siguiendo una escala logarítmica.
Análisis estadístico
Modelos lineales generalizados (MLG) se emplearon para evaluar el efecto de la humedad relativa (HR), la temperatura y el tiempo de almacenamiento sobre el número de semillas germinadas, por su distribución binomial. En el modelo, el número de semillas germinadas fue usado como variable dependiente, mientras que el número de semillas sembradas se ingresó como covariable. La HR, la temperatura y el tiempo de almacenamiento fueron considerados como factores fijos. Cuando los factores tuvieron efecto significativo (P < 0,05), se utilizó la prueba de comparación de múltiples medias DGC (Di Rienzo et al., 2020) para establecer las diferencias entre ellas. Los tratamientos en los que la germinación fue igual a cero, en todas las réplicas, no fueron incluidos en el análisis. Todos los análisis estadísticos se realizaron con InfoStat (Di Rienzo et al., 2020).
RESULTADOS
Caracterización de las semillas
El peso y el contenido de humedad (CH) inicial de las semillas fueron variables entre las especies (Tabla 1). Jacaranda mimosifolia fue la especie con las semillas más livianas (10,7 ± 1,6 mg) y Pterogyne nitens las más pesadas (95,5 ± 22,6 mg). El menor CH inicial se registró en P. nitens (5,8 ± 0,02 %), mientras que el mayor en J. mimosifolia (8,1 ± 0,2 %). En todos los casos, la viabilidad inicial de las semillas (Vi) fue mayor o igual a 87%. El porcentaje de germinación inicial fue mayor o igual a 83 % en Handroanthus impetiginosus, J. mimosifolia, y en las semillas escarificadas de Neltuma alba y P. nitens. En estas especies, cuando las semillas se sembraron intactas, sin escarificar, germinaron un 13% y 36%, respectivamente (Tabla 1).
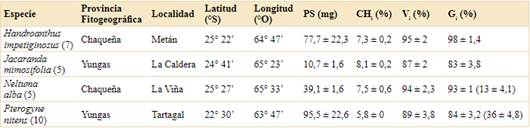
Tabla 1 Caracterización de las poblaciones evaluadas. Provincia Fitogeográfica, localidad, latitud y longitud de los sitios de colecta, peso de la semilla (PS; media ± desvío estándar), contenido de humedad inicial (CH ; media ± error estándar), viabilidad inicial determinada por la prueba de tetrazolio (Vi; media ± desvío estándar) y porcentaje de germinación inicial (Gi; media ± error estándar). En cada especie se indica entre paréntesis el número de individuos recolectados por sitio. La Provincia Fitogeográfica a la que pertenece cada sitio se estableció según Oyarzabal et al. (2018). En Gi de N. alba y P. nitens se indica entre paréntesis el porcentaje de germinación corresponde a las semillas intactas, sin escarificar.
Comportamiento durante el almacenamiento
Los porcentajes de germinación cuando las semillas permanecieron en un ambiente con 60 % HR y 15 % HR no presentaron diferencias significativas con las semillas frescas recién colectadas (P > 0,05; Fig. 1 y 2), al igual que las semillas desecadas a 15 % HR y almacenadas a -20 °C durante 12 meses. Durante el tiempo de almacenamiento, el menor porcentaje de germinación correspondió a semillas almacenadas a 5 °C para P. nitens (60 %, P < 0,0001; Fig. 2), y a 20 °C para H. impetiginosus, J. mimosifolia y N. alba (79 %, P = 0, 0008; 64%, P = 0, 0065 y 62 %, P < 0,0001; respectivamente; Fig. 1 y 2). Las semillas intactas de N. alba y P. nitens no superaron el 25 % y 37 % de germinación en los tratamientos evaluados, siendo significativamente diferentes de la germinación en semillas escarificadas (P < 0,05; Fig. 2).

Fig. 1 Porcentaje de germinación (media ± error estándar) de las semillas de Handroanthus impetiginosus y Jacaranda mimosifolia, almacenadas a -20 °C, 5 °C y 20 °C, durante 3, 6 y 12 meses, respectivamente. Abreviaturas: F, semillas frescas; 60%, semillas almacenadas a 60 % HR; 15%, semillas almacenadas a 15 % HR. Un asterisco (*) indica diferencia significativa con respecto a los demás tratamientos (P < 0,05).
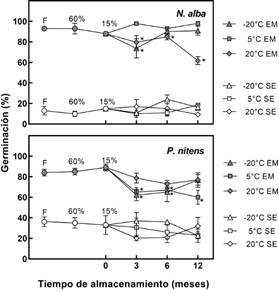
Fig. 2 Porcentaje de germinación (media ± error estándar) de las semillas de Neltuma alba y Pterogyne nitens, almacenadas a -20 °C, 5 °C y 20 °C, durante 3, 6 y 12 meses, respectivamente. Abreviaturas: F, semillas frescas; 60%, semillas almacenadas a 60 % HR; 15%, semillas almacenadas a 15 % HR; EM, semillas escarificadas mecánicamente; SE, semillas intactas, sin escarificado. Un asterisco (*) indica diferencia significativa con respecto a los demás tratamientos (P < 0,05).
Longevidad potencial de las semillas
Las curvas de supervivencia de las semillas, obtenidas empleando la ecuación (1), se observan en la Fig. 3. El tiempo en el que la germinación fue igual a 0 % fue variable entre especies. En H. impetiginosus las semillas perdieron la totalidad de su capacidad germinativa en 26 días, mientras que las semillas de J. mimosifolia alcanzaron el 0% de germinación a los 50 días. Los intervalos de tiempo en las semillas intactas de P. nitens se extendieron hasta los 200 días, momento en el que se registró 3% de germinación. Por el contrario, en N. alba, se registró un 70 % de germinación en el día 396, momento en el que se utilizó la última muestra de semillas envejecidas (de 12 muestras almacenadas inicialmente), por lo que no fue posible establecer el tiempo correspondiente a 0 % de germinación en esta especie. Para N. alba y P. nitens, las curvas de supervivencia fueron diferentes entre semillas envejecidas intactas y escarificadas. En las semillas escarificadas de N. alba y P. nitens, se alcanzó el 0% de germinación a los 160 y 75 días, respectivamente (Fig. 3).

Tabla 2 Tiempo requerido para que la viabilidad disminuya al 50% (P 50 ). En especies con dormición física (N. alba y P. nitens), las semillas se envejecieron bajo dos tratamientos: intactas sin escarifación (SE) y escarificadas mecánicamente (EM). El valor de R2 corresponde al análisis Probit.
El P 50 obtenido a partir de la ecuación (2) fue variable entre las especies (Tabla 2). Las semillas de H. impetiginosus registraron el menor valor de P 50 (P 50 = 7,7 días), mientras que N. alba registró el valor más alto (P 50 = 700 días), siendo la única especie cuyo P 50 superó los 100 días. Las semillas envejecidas escarificadas de N. alba y P. nitens murieron antes que las semillas intactas y, en consecuencia, el P 50 fue menor. La mayor diferencia entre P 50 correspondió a N. alba, dónde las semillas escarificadas presentaron un P 50 = 55,3 días y las semillas intactas un P 50 = 700 días. En todos los casos, los valores estimados del análisis Probit se ajustaron con R2 > 0,8 (Tabla 2).

Fig. 3 Curvas de supervivencia de las semillas. Porcentaje de germinación (media ± error estándar) en función del tiempo de envejecimiento. Los puntos negros y las líneas entrecortadas indican que las semillas fueron escarificadas mecánicamente antes de iniciar el envejecimiento acelerado, a diferencia de las semillas intactas, sin escarificar, en puntos blancos y líneas completas.
DISCUSIÓN Y CONCLUSIONES
En este trabajo, las semillas de las cuatro especies toleraron la desecación (4-5 % CH) y el almacenamiento a bajas temperaturas (-20 °C) durante 12 meses, por lo que pueden clasificarse como ortodoxas y, por lo tanto, conservarse en bancos de semillas. Resultados similares se registraron en otras especies de árboles nativos de Argentina pertenecientes a los géneros Amburana, Aspidosperma, Cedrela, Cordia, Erithryna, Neltuma, Parkinsonia, Strombocarpa, Tecoma y Zizipus (De Viana et al., 2009; Giamminola et al., 2012; Morandini et al., 2013; Galíndez et al., 2015, 2019; Urtasun et al., 2015; Otegui et al., 2016). Si bien, estos resultados coinciden con la información presente en las bases de datos SID KEW (SER, INSR, RBGK, 2023) y GENESYS (USDA, 2023), otros estudios indicaron una disminución de la germinación por efecto de la desecación en poblaciones de Handroanthus impetiginosus provenientes de Brasil, distinto a lo observada en este trabajo (Ribeiro & Borghetti, 2014; Da Silva Araújo et al., 2021). Estas diferencias podrían estar asociadas a la variabilidad inter poblacional, a lo largo de gradientes ambientales, como se ha registrado en otras especies de árboles de la región (Carón et al., 2020; Galíndez et al., 2019). Futuros estudios deberán evaluar la variación inter poblacional en la respuesta germinativa de las cuatro especies estudiadas aquí y su implicancia sobre el comportamiento durante el almacenamiento. En las especies estudiadas, la sensibilidad al almacenamiento a 5 °C y 20 °C, denota la importancia de conservar las semillas en condiciones óptimas de banco de semillas (15 % HR y -20 °C) a fin de evitar el deterioro producido por temperaturas de almacenamiento inadecuadas (Galíndez et al., 2019; Solberg et al., 2020). Esto debe tenerse en cuenta en el diseño de protocolos de conservación específicos, los que deben considerar las posibles variaciones entre poblaciones de una misma especie, antes mencionadas.
De acuerdo con los resultados, la desecación y el almacenamiento a bajas temperaturas no afectó el estado de dormición física en las dos especies de leguminosas (Neltuma alba y Pterogyne nitens). Esto coincide con los resultados obtenidos por otros autores cuando se evaluó el efecto del almacenamiento sobre la dormición física (Galíndez et al., 2010; Godefroid et al., 2010; Ashworth et al., 2017). Morandini et al. (2013), registraron que las semillas frescas de P. nitens no presentaban dormición física, y, acorde a los bajos porcentajes de germinación obtenidos en semillas intactas durante el almacenamiento, ellos propusieron que las semillas adquieren un estado de dormición inducida cuando son desecadas. Contrario a esto, las semillas de P. nitens evaluadas en nuestro trabajo, presentaron dormición física cuando fueron evaluadas frescas y luego de ser sometidas a desecación. La presencia de dormición física en esta especie también fue reportada por Pece et al. (2010) y Nascimiento et al. (2021). Futuros estudios deberían reevaluar la presencia de dormición física en otras poblaciones, incluyendo la población estudiada por Morandini et al. (2013), analizando la variación espacial y temporal de esta característica a lo largo de su distribución geográfica en el noroeste de Argentina.
Las curvas de supervivencia de la semilla y la ecuación de viabilidad permitieron clasificar a las especies según su longevidad potencial comparativa. Teniendo en cuenta su valor de P 50 , H. impetiginosus se clasifica como especie de vida corta, Jacaranda mimosifolia y P. nitens como especies de vida intermedia y N. alba como especie de larga vida. Si bien las especies de la familia Fabaceae se han caracterizado como longevas, puede observarse un amplio rango de P 50 entre las especies ya evaluadas. De esta manera, especies de una misma familia pueden incluirse en distintas categorías de longevidad potencial (Walters et al., 2005; Probert et al., 2009; Solberg et al., 2020). Se ha indicado que la longevidad potencial puede variar entre especies dentro de una misma forma de vida (por ejemplo, árboles), linaje, o incluso entre poblaciones de una misma especie (Walters et al., 2005; Probert et, al. 2009; Mondoni et al., 2011; Daives et al., 2020; Pritchard et al., 2020). La importancia de esta prueba reside en que es una herramienta de suma utilidad, ya que permitiría saber cuándo debe regenerarse el material conservado si se trata de especies de ciclo de vida corto o cuándo colectar nuevo germoplasma en el caso de especies perennes (Walters, 2015). De acuerdo con los resultados, el monitoreo debe ser diferenciado entre especies de vida corta, intermedia y larga. Se ha propuesto que para especies clasificadas como longevas (como N. alba), el monitoreo debería realizarse a intervalos de tiempo más amplios que los estándares (por ejemplo, cada 15 años; Ellis et al., 2018; Hanson & Ellis, 2020). Mientras que, para especies de vida corta (como H. impetiginosus), será recomendable incorporar monitoreos antes de los 10 años (Probert et al., 2009).
Por último, las dos especies estudiadas de la familia Fabaceae (N. alba y P. nitens) presentaron longevidades diferentes cuando el envejecimiento acelerado se realizó en semillas escarificadas y sin escarificación. Sin embargo, en P. nitens, dicha diferencia no fue suficiente para alcanzar una categoría diferente, ya que en ambos casos se clasificó como de categoría intermedia. Independientemente de esto, la presencia de dormición física en las semillas de leguminosas (y posiblemente de otras familias con este tipo de dormición) les conferiría una gran longevidad durante el almacenamiento. Si bien se conoce poco sobre el impacto que la cubierta dura tiene sobre la longevidad de las semillas, se ha propuesto que la actividad metabólica sería extremadamente lenta, con un reducido intercambio con el medio (Chin et al., 2013). Estas características podrían ser muy valiosas para la propagación de las especies con fines restaurativos y de producción, principalmente para N. alba que, según informes, actualmente se propaga por técnicas de injerto y cultivo de tejidos in vitro (López Lauenstein et al., 2021). Nuestros resultados indican que, las semillas de N. alba, conservadas con escarificación previa (i.e. con ruptura de la dormición física; lista para la siembra), pueden sobrevivir por periodos similares a los de especies clasificadas como de vida intermedia, lo que facilitaría su conservación a corto-mediano plazo en bancos activos y su posterior uso con fines de producción, regeneración y restauración.
En conclusión, las semillas de las cuatro especies toleran la desecación, son ortodoxas y pueden almacenarse en bancos de semillas. Las semillas intactas de N. alba y P. nitens conservan su estado de dormición física incluso después de permanecer almacenadas 12 meses, en condiciones de 15% HR y -20 °C. Las semillas de H. impetiginosus se clasificaron como de vida corta, las de J. mimosifolia y P. nitens como de vida intermedia y las de N. alba como de larga vida, siendo estas últimas las únicas en las que la longevidad en semillas escarificadas correspondió a una categoría diferente (vida intermedia). La información obtenida en este trabajo permitirá incorporar nuevas poblaciones a la Red de Bancos de Germoplasma (RBG) del INTA y la Red Argentina de Bancos de Germoplasma de Plantas Nativas (Red-ARGENA), incrementando el conocimiento sobre la longevidad de las semillas en estas cuatro especies de árboles nativos.

