










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkMedicina (Buenos Aires)
Print version ISSN 0025-7680
Medicina (B. Aires) vol.71 no.3 Ciudad Autónoma de Buenos Aires June 2011
COMUNICACIÓN BREVE
La depleción de las células T regulatorias aumenta el número de las células CD8 durante la infección con el virus del tumor mamario murino
Gabriel Cabrera, Juliana Mundiñano, Gabriela Camicia, Héctor Costa, Irene Nepomnaschy, Isabel Piazzon
Instituto de Leucemia Experimental, Consejo Nacional de Investigaciones Científicas y Técnicas, División Medicina Experimental, Instituto de Investigaciones Hematológicas, Academia Nacional de Medicina, Buenos Aires, Argentina
Dirección postal: Dra. Isabel Piazzon, ILEX- CONICET, División Medicina Experimental, Instituto de Investigaciones Hematológicas, Academia Nacional de Medicina, Pacheco de Melo 3081, 1425 Buenos Aires, Argentina.
Fax: (54-11) 4803-9475 e-mail: ipiazzon@hematologia.anm.edu.ar
Resumen
El virus del tumor mamario murino (MMTV) es un retrovirus que se transmite durante la lactancia y que ha desarrollado estrategias para explotar y subvertir el sistema inmune. En un modelo de infección natural con MMTV hemos mostrado previamente que la infección causa incrementos tempranos y progresivos de células T regulatorias (Treg) CD4+CD25+Foxp3+ específicas para el superantígeno (Sag) viral en las placas de Peyer (PP). En este trabajo se evaluó si la depleción de las células Treg influencia la población de células CD8+ durante la infección con MMTV a través del amamantamiento. La depleción de las células Treg al día 6 de infección causó incrementos en el porcentaje y número absoluto de las células CD8+ en los ganglios y provocó un incremento en la intensidad de fluorescencia media del marcador de activación CD44 en esas células. Los incrementos en el número absoluto de las células CD8 se observaron en células con cadenas variables Vβ del receptor de las células T (TCR) tanto reactivas como no reactivas al Sag. Previamente habíamos demostrado que la depleción de las células Treg al día 6 de infección disminuye la carga viral. Los resultados presentados en este trabajo sugieren que, al menos a partir del día 6 de infección con MMTV, las células Treg podrían tener un rol inhibiendo la generación de una respuesta CD8 antiviral.
Palabras clave: MMTV; Células T regulatorias; Células CD8
Abstract
Regulatory T cell depletion increases the number of CD8 cells during mouse mammary tumor virus infection. Mouse mammary tumor virus (MMTV) is a milk-borne betaretrovirus that has developed strategies to exploit and subvert the host immune system. We have shown in a natural model of MMTV infection that the virus causes early and progressive increases in superantigen (Sag)-specific CD4+ CD25+ Foxp3+ regulatory T cells (Treg) in Peyer's patches. Herein, we evaluated whether the depletion of Treg cells affects the CD8+ population during milk-borne MMTV infection. At day 6 of infection, the depletion of Treg cells increased the percentage and absolute number of CD8+ cells in lymph nodes as well as the mean intensity fluorescence of the CD44 activation marker. The absolute number of CD8+ cells was increased in cells bearing both Sag reactive and non-reactive TCR Vβ chains. We have previously shown that regulatory T cell depletion at day 6 of infection decrease viral load. Results reported herein suggest that at least after day 6 of MMTV infection Treg cells play an inhibiting role on CD8 antiviral response.
Key words: MMTV; Regulatory T cells; CD8 cells
El virus del tumor mamario murino (MMTV) es un retrovirus que se transmite durante la lactancia. Debido a que el MMTV es un patógeno que se caracteriza por utilizar al sistema inmune del huésped para establecer la infección, ha sido empleado en el desarrollo de modelos que permiten estudiar qué mecanismos pueden ser usados por los patógenos, y en este caso en particular por los retrovirus, con el fin de tomar ventaja de la respuesta inmune que ellos mismos generan1, 6. El MMTV codifica para un superantígeno (Sag) en su genoma, y los mecanismos utilizados por el MMTV para establecer la infección dependen en gran medida de la fuerte respuesta inmune que genera la expresión del Sag viral1, 6.
Se ha descrito que el MMTV activa e infecta linfocitos B y células dendríticas (CDs) de las Placas de Peyer (PP), y que luego las células infectadas expresan y presentan el Sag a células T portadoras de cadenas Vβ reactivas al mismo, causando su proliferación2, 7, 12. En un modelo de infección con MMTV en la almohadilla plantar de ratones se ha determinado que la interacción entre células T que reconocen al Sag y las células B causa la proliferación de los linfocitos B infectados y amplifica el reservorio viral1, 6. Aunque no ha sido probado formalmente, se asume que eventos similares tienen lugar durante la infección por la vía natural a través de la leche. Se sabe además, que los eventos que tienen lugar como consecuencia de la respuesta inmune que causa el Sag luego de la cooperación T-B son críticos, ya que si no se producen el virus infecta con muy baja eficiencia. Nuestro grupo de trabajo ha demostrado que dentro de estos eventos dependientes de la respuesta al Sag, se producen incrementos en células B IgA+ que podrían ser importantes en el posterior traslado del virus a la glándula mamaria5. Por último, una vez que el epitelio mamario es infectado, el virus es secretado a la leche y puede pasar a la próxima generación completando su ciclo de vida1, 6.
Estudios recientes han postulado que las células T regulatorias Foxp3+ (Treg) podrían ser capaces de impedir una adecuada respuesta inmune contra una variedad de patógenos foráneos13.
Nuestro grupo de trabajo ha mostrado que durante las dos primeras semanas de infección con MMTV(BALB6) se producen en las PP incrementos tempranos y progresivos de células T regulatorias (Treg) específicas para el Sag Vβ6+ CD4+ CD25+ Foxp3+4. Las células CD4+ CD25+ Foxp3+ Vβ10+ no específicas para el Sag no sufren alteraciones significativas4. Además, se determinó que las células Treg purificadas al día 6 de infección poseen capacidad inmunosupresora en cultivos mixtos linfocitarios y se observó que la depleción de las células Treg con anticuerpo anti-CD25 luego de los primeros 6 días de infección determina menores niveles de carga viral en el ganglio mesentérico y en el bazo, sugiriendo que las células Treg favorecerían la persistencia de la infección con MMTV4.
Se ha comunicado que durante la infección con MMTV el huésped es incapaz de generar una respuesta de células CD89. Las células Treg son capaces de inhibir la actividad de las células CD811 y en algunos modelos de infección se ha descrito que la depleción de las células Treg permite recuperar en cierta medida la respuesta inmune contra determinados patógenos13.
Considerando que la depleción de las células Treg afecta los niveles de infección del MMTV, el objetivo de este trabajo fue evaluar mediante experimentos de depleción de las células Treg, la posible influencia de dichas células en la población de células CD8 durante la infección con MMTV a través del amamantamiento.
Las depleciones con el anticuerpo anti-CD25 se llevaron a cabo luego del día 6 de infección, debido a que a partir de ese día aproximadamente el 85-90% de las células CD25+ expresan el factor de transcripción Foxp34. Con el protocolo utilizado se eliminaron aproximadamente el 90% de las células CD4+ CD25+ en todos los grupos de animales que tuvieron depleción4.
Se amamantaron ratones BALB/c de 8 días de edad con nodrizas infectadas con MMTV(BALB6) durante 6 días y luego fueron inoculados con el anticuerpo anti-CD25 durante 3 días consecutivos4. A los 23 días de infección los ratones fueron sacrificados, y se analizó por citofluorometría de flujo (FACS) la expresión de CD8 y CD44 en las células de los ganglios axilares e inguinales.
Los ratones fueron mantenidos de acuerdo a las normas de la guía para el cuidado y uso de animales de laboratorio utilizada en el ILEX-CONICET10.
La depleción de las células Treg causó un incremento en el porcentaje y número absoluto de las células CD8+ en los ganglios de los ratones infectados (Fig. 1, A y B). Además, se observó que las células CD8+ de los ratones infectados y que tuvieron depleción de células Treg presentan una mayor intensidad de fluorescencia media del marcador de activación CD44 (Fig. 1, C y D).
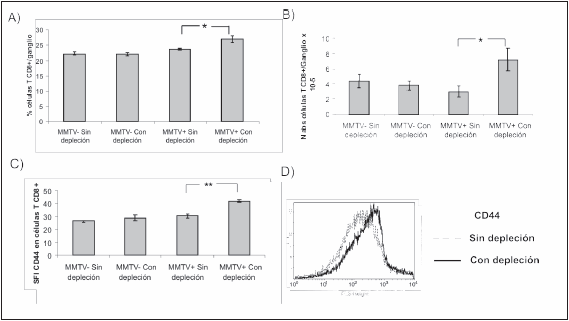
Fig. 1.- Expresión de CD8 y CD44 en los ganglios luego de la depleción de las células Treg al día 6 de infección. Ratones BALB/c de 8 días de edad fueron amamantados con nodrizas infectadas o no infectadas por el MMTV(BALB6). A los 6 días de infección la mitad de los ratones amamantados con cada nodriza sufrió depleción de células Treg con anticuerpo anti-CD25. Quince días después de la última inoculación los ratones fueron sacrificados y se midió por citometría de flujo la expresión de CD8 y CD44 en los ganglios axilares e inguinales de los ratones infectados o no infectados y que tuvieron o no depleción de células Treg. A) Porcentaje de células CD8+; B) Número de células CD8+; C) Indice de fluorescencia específica (SFI) de CD44 en las células CD8+. La SFI se calculó de acuerdo a la siguiente fórmula: Media de la intensidad de fluorescencia del anticuerpo anti-CD44/ Media de la intensidad de fluorescencia del control de isotipo; D) Histograma representativo de la expresión de CD44 en las células CD8+ de los ratones infectados que tuvieron o no depleción de células Treg. Los datos se presentan como la media ± Error estándar; n = 3. En A) *:p = 0.046, en B) *p = 0.034, en C) **p = 0.0017. Se muestra un experimento representativo de dos ensayos realizados de manera independiente y con resultados similares.
Se ha comunicado que la infección con MMTV causa una disminución en el porcentaje y en el número de las células CD8+ que portan cadenas Vβ reactivas al Sag en la periferia. Esta disminución sería una consecuencia de la deleción clonal intratímica producida por la expresión del Sag en el timo infectado8. Para evaluar si la depleción de las células Treg influencia tanto las células CD8 con cadenas Vβ en el TCR reactivas como no reactivas al Sag, se analizó por FACs la expresión de Vβ6 (reactivas al Sag) y Vβ10 (no reactivas al SAg) en las células CD8+.
Se observó que en los ganglios de los ratones infectados y con depleción de células Treg no se produce la disminución de las células CD8+Vβ6+ que tiene lugar en los ratones infectados que no sufrieron depleción de células Treg (Fig. 2A).
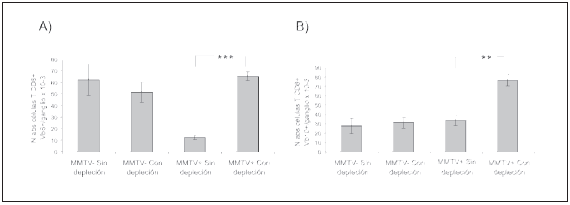
Fig. 2.- Especificidad de las células CD8+ en los ganglios luego de la depleción de las células Treg al día 6 de infección. Ratones BALB/c de 8 días de edad fueron amamantados con nodrizas infectadas o no infectadas por el MMTV(BALB6). A los 6 días de infección la mitad de los ratones amamantados con cada nodriza sufrió depleción de células Treg. Quince días después de la última inoculación los ratones fueron sacrificados y se midió por citometría de flujo la expresión de CD8, Vß6 y Vß10 en los ganglios axilares e inguinales. A) Número absoluto de las células CD8+ Vß6+ en los ratones infectados o no infectados y con o sin depleción de células Treg; B) Número absoluto de las células CD8+ Vß10+ en los ratones infectados o no infectados y con o sin depleción de células Treg. Los datos se presentan como la media ± error estándar; n = 3. En A) ***p = 0.0008, en B) **p = 0.0074. Se muestra un experimento representativo de dos ensayos realizados de manera independiente y con resultados similares.
Además, se observó que los ratones infectados y con depleción de células Treg presentan un mayor número absoluto de células CD8+Vβ10+ con respecto a los ratones infectados pero sin depleción de células Treg (Fig. 2B)
Discusión
Se ha descrito que el MMTV es capaz de subvertir de tal manera el sistema inmune del huésped que la respuesta inmune de células CD8 no puede generarse9. Nuestro grupo de trabajo ha comunicado que la infección con MMTV por la vía natural a través de la leche genera incrementos tempranos en células Treg Foxp3+ específicas para el Sag viral4. Considerando que en un número creciente de publicaciones se postula que las células Treg pueden ser inducidas por diferentes patógenos como parte de una estrategia para atenuar o impedir una respuesta CD8 citotóxica por parte del huésped13, en este trabajo se investigó si la depleción de las células Treg podría influenciar a la población de células CD8 durante la infección con MMTV.
Los resultados mostraron que la depleción de las células Treg luego de los primeros días de infección causa un incremento en las células CD8+ en los ganglios, sugiriendo que durante la infección con MMTV las células Treg podrían tener el rol de impedir que se produzca la expansión de las células CD8, que suele ser necesaria para que se genere una adecuada respuesta citotóxica. En correspondencia con esta observación, sólo en los ratones infectados que sufrieron depleción de las células Treg se produjeron incrementos en la intensidad de fluorescencia media del marcador de activación CD44 en las células CD8+.
Se ha comunicado que la infección con MMTV causa en la periferia un descenso progresivo del número de células CD8+ portadoras de cadenas Vβ reactivas al Sag. Esta disminución no se produciría como consecuencia de la interacción de las células CD8+ con el Sag viral en la periferia, ya que éste se presenta exclusivamente en el contexto de moléculas de histocompatibilidad de clase II, sino debido a que el MMTV es capaz de infectar el timo, causando la deleción de todos los timocitos inmaduros reactivos al Sag8. Como consecuencia de la infección del timo dejarían de generarse y exportarse células CD8 con receptores Vβ con especificidad para reconocer al Sag viral, y el porcentaje y número de esas células disminuiría en la periferia por un efecto de dilución generado por la exportación continua de células CD8+ con receptores Vβ con otras especificidades8.
Se observó que cuando las células Treg son eliminadas, no se produce la disminución de las células CD8+ Vβ6+ que tiene lugar en los ratones infectados con MMTV que no fueron deplecionados, lo cual sugiere que en ausencia de las células Treg, se produciría una menor infección del timo, o bien que la infección sería más lenta.
Por otra parte, se observó que los ratones infectados y que tuvieron depleción de células Treg tienen mayor número absoluto de células CD8+ con receptores con cadenas Vβ10 que no son reactivas al Sag. Este resultado sugiere que las células CD8+ que reconocen otros antígenos virales podrían estar incrementadas en los ratones que tuvieron depleción de células Treg.
Resultados preliminares obtenidos en nuestro laboratorio sugieren que la citotoxicidad in vivo de las células CD8 está aumentada en los ratones que tuvieron depleción de células Treg.
Los datos presentados correlacionan con resultados previos que muestran que la depleción de las células Treg al día 6 de infección determina menores niveles de carga viral, y en su conjunto sugieren que la falta de respuesta CD8 frente al MMTV9 puede ser, al menos en parte, una consecuencia de la actividad de las células Treg.
Se ha descrito que el MMTV impide la respuesta de células CD8+ mediante un mecanismo que depende del TLR49. En este sentido, es interesante señalar que resultados obtenidos en nuestro laboratorio muestran que en ratones deficientes en TLR4 se producen menores incrementos en células Treg3.
Por otra parte, es importante destacar que, si bien los resultados obtenidos con la depleción de las células Treg a partir del día 6 de infección sugieren que esas células favorecerían al virus atenuando una respuesta CD8 antiviral, resultados previos obtenidos en nuestro laboratorio indican que, por el contrario, al inicio de la infección las células Treg favorecían al huésped al disminuir la respuesta inmune que genera el Sag, que es crítica para la amplificación del reservorio viral4. En su conjunto, los resultados obtenidos indicarían que durante el desarrollo de la infección por este retrovirus las células Treg podrían tener un rol dual y complejo.
El conocimiento de los mecanismos que utilizan los patógenos para evadir o impedir la respuesta inmune del huésped es imprescindible para diseñar e implementar estrategias terapéuticas contra diferentes agentes infecciosos. Los resultados preliminares presentados en este trabajo indicarían que, al menos a partir del día 6 de infección con MMTV, las células Treg podrían tener un rol inhibiendo la generación de una respuesta CD8 antiviral.
1. Acha-Orbea H, Shakhov AN, Finke D. Immune response to MMTV infection. Front Biosci 2007; 12: 1594-609. [ Links ]
2. Burzyn D, Rassa JC, Kim D, Nepomnaschy I, Ross SR, Piazzon I. Toll-like receptor 4-dependent activation of dendritic cells by a retrovirus. J Virol 2004; 78: 576-84. [ Links ]
3. Cabrera G, Burzyn D, Courreges MC, et al. Increases in Superantigen-specific Treg cells during MMTV infection are TLR4 dependent. In: 13th International Congress of Immunology - ICI, Rio de Janeiro, Brazil, 2007, Medimond - Monduzzi Editore International Proceedings Division; 523-528. [ Links ]
4. Cabrera G, Burzyn D, Mundiñano J, et al. Early increases in superantigen-specific Foxp3+ regulatory T cells during mouse mammary tumor virus infection. J Virol 2008; 82: 7422-31. [ Links ]
5. Cabrera G, Vercelli C, Burzyn D, et al. Increases in IgA+ B cells in the Peyer's patches during milk-borne Mouse Mammary Tumor Virus infection are influenced by TLR4 and are completely dependent on the superantigen response. J Gen Virol 2010; 91(Pt 11): 2814-20. [ Links ]
6. Czarneski J, Rassa JC, Ross SR. Mouse mammary tumor virus and the immune system. Immunol Res 2003; 27: 469-80. [ Links ]
7. Held W, Waanders GA, Shakhov AN, Scarpellino L, Acha-Orbea H, MacDonald HR. Superantigen-induced immune stimulation amplifies mouse mammary tumor virus infection and allows virus transmission. Cell 1993; 74: 529-40 [ Links ]
8. Ignatowicz L, Kappler J, Marrack P. The effects of chronic infection with a superantigen-producing virus. J Exp Med 1992; 175: 917-23. [ Links ]
9. Jude BA, Pobezinskaya Y, Bishop J, et al. Subversion of the innate immune system by a retrovirus. Nat Immunol 2003; 4: 573-8. [ Links ]
10. Institute of Laboratory Animal Resources Commision on Life Sciences. Guide for the care and use of laboratory animals. National Research Council. Washington, DC.: National Academy Press, 1996. [ Links ]
11. Piccirillo CA, Shevach EM. Cutting edge: control of CD8+ T cell activation by CD4+CD25+ immunoregulatory cells. J Immunol 2001; 167: 1137-40. [ Links ]
12. Rassa JC, Meyers JL, Zhang Y, Kudaravalli R, Ross SR. Murine retroviruses activate B cells via interaction with toll-like receptor 4. Proc Natl Acad Sci USA 2002; 99: 2281-6. [ Links ]
13. Rouse BT, Sarangi PP, Suvas S. Regulatory T cells in virus infections. Immunol 2006; 212: 272-86. [ Links ]
Recibido: 19-1-2011
Aceptado: 18-5-2011

