










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkMedicina (Buenos Aires)
Print version ISSN 0025-7680
Medicina (B. Aires) vol.73 no.6 Ciudad Autónoma de Buenos Aires Dec. 2013
ARTÍCULO ORIGINAL
Sexual hormones modulate compensatory renal growth and function
Pablo J. Azurmendi1, Elisabet M. Oddo1, Jorge E. Toledo1, Rodolfo S. Martin1, 2, Fernando R. Ibarra1, Elvira E. Arrizurieta1, 2
1Laboratorio de Nefrología Experimental, Instituto de Investigaciones Médicas Alfredo Lanari, Universidad de Buenos Aires,
2Consejo Nacional de Investigaciones Científicas y Técnicas
Postal address: Dra. Elvira E. Arrizurieta, Av. Combatientes de Malvinas 3150, 1427 Buenos Aires, Argentina
Fax: (54-11) 4523-4094 e-mail: ilanari@pinos.com
Abstract
The role played by sexual hormones and vasoactive substances in the compensatory renal growth (CRG) that follows uninephrectomy (uNx) is still controversial. Intact and gonadectomized adult Wistar rats of both sexes, with and without uNx, performed at 90 days age, were studied at age 150 days. Daily urine volume, electrolyte excretion and kallikrein activity (UKa) were determined. Afterwards, glomerular filtration rate and blood pressure were measured, the kidneys weighed and DNA, protein and RNA studied to determine nuclei content and cell size. When the remnant kidney weight at age 150 days was compared with the weight of the kidney removed at the time of uNx, male uNx rats showed the greatest CRG (50%) while growth in the other uNx groups was 25%, 15% and 19% in orchidectomized, female and ovariectomized rats, respectively. The small CRG observed in the uNx female rats was accompanied by the lowest glomerular filtration value, 0.56 ± 0.02 ml/min/g kwt compared, with the other uNx groups, p < 0.05. Cell size (protein or RNA/DNA) was similar for all the groups except for uNx orchidectomized rats. In this group the cytoplasmatic protein or RNA content was lower than in the other groups while DNA (nuclei content) was similar. Some degree of hyperplasia was determined by DNA content in the uNx groups. Male sexual hormones positively influenced CRG and its absence modulated cell size. Female sexual hormones, instead, did not appear to stimulate CRG. The kallikrein kinin system may not be involved in CRG.
Key words: Cell volume regulation; Compensatory renal hypertrophy; Kallikrein kinin system; Renal size; Uninephrectomy
Resumen
Las hormonas sexuales modulan el crecimiento renal compensador (CRC) y la función renal en la uninefrectomía (uNx). La importancia que pueden tener las hormonas sexuales y sustancias vasoactivas sobre el crecimiento renal compensador (CRC) que sigue a la uninefrectomía es aún materia de debate. Se estudiaron ratas Wistar de ambos sexos, a los 150 días de vida, intactas y gonadectomizadas con y sin uNx, realizada a los 90 días de vida. Se midió volumen urinario diario y excreción de electrolitos y actividad de kalikreína urinaria. Se midió filtrado glomerular y presión arterial media extrayéndose luego los riñones que fueron pesados y preparados para estudios histológicos y determinación de ADN, ARN y proteínas para estimar contenido nuclear y tamaño celular. El CRC fue calculado comparando el peso del riñón al momento de las uNx (90 dias de vida) con aquel obtenido a los 150 días de vida. En las ratas macho uNx se observó el mayor CRC (50%) mientras que, en los otros grupos uNx solo alcanzó un 25%, 15% y 19%. El filtrado glomerular acompañó los cambios morfológicos observándose el menor filtrado en las ratas hembras uNx respecto al resto de los grupos 0.56 ± 0.02, p < 0.05. El tamaño celular (proteína o ARN/ ADN) fue similar para todos los grupos excepto para los orquidectomizados uNx, cuyo contenido citoplasmático fue menor. El contenido nuclear (ADN) fue semejante en todos los grupos. Se observó que el CRC está influenciado positivamente por las hormonas sexuales masculinas y su ausencia modula el tamaño celular. La falta de hormonas sexuales femeninas, en cambio, afecta negativamente el CRC. El sistema kalikreína kinina no parecería estar involucrado en el CRC.
Palabras clave: Regulación del volumen celular; Hipertrofia renal compensadora; Sistema kalikreína kinina; Tamaño celular; Uninefrectomía
It is well known that after uninephrectomy (uNx) the remaining kidney undergoes morphological and functional adaptation to compensate for the organ absence. In fact, in male rats, after three weeks uNx, glomerular filtration rate and kidney weight increased around 70% to 85%. Proximal tubular diameter increased by 20%, as well1. No new nephrons were formed and the increment in glomerular filtration rate resulted, according to micropunture studies, from the enhancement of single nephron glomerular filtration rate by hemodynamic adjustments2.
The factors that are sensed after renal mass ablation and serve as signals to initiate the adjustments in glomerular hemodynamics responsible for the increase in the remnant kidney glomerular filtration rate remain to be identified. Several vasoactive substances, including angiotensin II2, aldosterone3, natriuretic peptides4, 5, endothelins6, 7, eicosanoids8, 9, and bradykinin2 have been implicated.
Bradykinin is increased in the intact portion of the remnant kidney and in its peri-infarct portion until, at least, the seventh week following 5/6 nephrectomy10. Bradykinin is a potent vasodilator peptide that may contribute to hemodynamic adaptations after partial renal mass removal. Unfortunately, data about bradykinin and/or kallikrein-kinin system on this subject are scarce.
On the other hand, we have recently found a strong influence of sexual hormones upon kallikrein-kinin system while studying intact and gonadectomized SHR with both kidneys and, also, a kallikrein-kinin system gender difference at baseline11.
As a consequence, it was our interest to study both, the compensatory renal growth and function and the kallikrein-kinin system response after two months uNx in intact and gonadectomized adult Wistar rats of both sexes.
Materials and Methods
Forty Wistar rats of either sex were studied at age 150 days. Fifty percent of these animals underwent a unilateral nephrectomy at age 90 days. Previously, at age 60 days, half of the animals were gonadectomized. Thus, eight groups of rats with five rats in each resulted: -intact male; -uNx male; -orchidectomized; -orchidectomized uNx; -intact female; -uNx female; -ovariectomized and -ovariectomized uNx. The animals were fed ad libitum throughout the study with Cargill (Alimentos Balanceados, Buenos Aires, Argentina) and had free access to tap water. They were housed at 24 Cº with a dark:light cycle 12:12 h.
Gonads removal and right unilateral nephrectomy were performed under anesthesia at the time indicated above. Ovariectomy and orchidectomy were performed through a bilateral flank or a scrotum incision and after vessel ligation. The right kidney was also removed through a flank incision, isolated and decapsulated, ligated with 4-0 silk suture, excised and weighed. To assess CRG this kidney weight was compared with the weight of kidney measured at age 150 days. Sham operated animals were prepared in the same manner and the right kidney was manipulated but not removed. The area was cleansed with an antimicrobial agent and the incisions were sutured closed.
At age 150 days, the animals were housed in individual metabolic cages and a twenty-four-hour urine collection was made. The body weight was recorded and, later on, the animals were anesthetized with pentobarbital 50 mg/kg body weight, i.p. to perform clearance studies. Glomerular filtration rate was measured using Inutest (Laevosan Gesselschaft m.b.H, Linz, Austria), as previously described12. A tracheostomy was performed and the carotid artery, jugular vein and urinary bladder were catheterized with polyethylene tubes. Enough inulin to provide plasma concentrations of 0.2 mg/ml was given as a prime and sustaining infusion diluted in saline. The sustaining infusion was delivered at 0.036 ml/min by a Harvard syringe pump #944. Blank samples of blood and urine were collected and, after 45 minutes equilibrium, three thirty-minute urine collections were made and three blood samples were taken in the middle of the urine collection period. Inulin assay was made by the anthrone method13. Mean blood pressure was registered through the catheter placed in the carotid artery by a blood pressure transducer. At the end of the inulin clearance study the kidneys were removed, weighed, and fixed for histopathological studies. Thereafter, the kidneys were prepared for protein, DNA, and RNA measurements. Total protein and RNA content were used as cytoplasmatic markers and DNA as nuclei index14, 15. Protein/DNA and RNA/DNA ratios were used to measure cell size16, 17. To this end, the kidney cortex was separated from the medulla and divided into two parts. The tissues were homogenized and kept frozen until assayed. Urinary electrolyte, osmolality and urinary kallikrein activity were measured in the 24-h urine collection. Urine electrolyte and osmolality were measured by flame photometry and by the freezing point method, respectively. Urinary Kallikrein activity was measured in the 24-h urine samples by the amidolytic method18 using the chromogenic tripeptide substrate H-D-Val-Leu-Arg-p-nitroanilide (S2266, Kabi Diagnostica, Sweden) at pH 8.2. The p-nitroanilide release was measured at 405 nm. Kallikrein activity was expressed in nanokatal (nkat) units, 1 katal being the enzyme activity that splits 1 mol of substrate per second of incubation at 37 °C. For urine kallikrein determinations the samples were previously diluted in distilled water, 1:250.
Protein determination: Renal cortex was homogeneized at high velocity for quick dissolution with buffer (Tris 20 mM, EGTA 2 mM, EDTA 2mM, PMSF 1 mM, β-mercaptoethanol 10 mM, Trasylol 100 KIE/ml) and corrected for a final pH of 7.4. Proteins were measured by the Bradford colorimetric method19 using 1/100 dilution at 570 nm and expressed as mg/g renal cortex. Serum albumin (0.125 to 4 μg/ml) was used as standard.
Measurement of nucleic acids: DNA and RNA were isolated from the renal cortex homogenates using the SV/Total RNA Isolation System (Promega), according to manufacturer's instructions20. This system allows sequential separation of nucleic acids utilizing different ethanol solution concentrations in nuclease free water for elution. DNA and RNA samples were stored at -70 °C until assayed. A ratio between optical density at 260 and 280 nm greater than 1.8 was considered a good index of the quality of the sample. DNA and RNA were measured by UV spectrophotometry at 260 nm, considering that one optical density represents 50 and 40 μg/ml, respectively. Results were expressed as mg/g renal cortex.
Statistics: Results were expressed as mean ± SEM. A p value lower than 0.05 was considered significant. The least square method for correlation was used when indicated.
This study involved a 3 × 2 factorial design, with uninephrectomy, gender and gonadectomy representing the independent variables. Results were analyzed using a factorial MANOVA, with mean variables (kidney weight, glomerular filtration rate, cell size, urinary kallikrein excretion and urinary volume and electrolytes renal excretion) representing dependent measures to be investigated. After the overall MANOVA was found to be significant for the mean variables and/or for their respective interactions, differences between groups were explored according to dependent measure.
All procedures were in conformance with the rules for Care and Use of Laboratory Animals approved by a Local Ethics Committee in accordance with the National Institute of Health (NIH) Guide for the care and use of laboratory animals and with the Helsinki Declaration of 1975, as revised in 2000.
Results
Gonadectomy and uNx effect on kidney and body weight
The factorial MANOVA analysis showed that uNx, gender or gonadectomy by themselves, as well as their combinations, modified the remnant kidney weight (p < 0.005 in all cases). As shown in Table 1 and Fig. 1, remnant kidney weight increased after uNx in male, orchidectomized and ovariectomized and female rats (by 50%, 25%,19% and 15%, respectively) when compared with the kidney weight measured at the time of uNx (age 90 days), while gonadectomy increased the remnant kidney weight in a similar way, independently of gender. uNx female kidney growth was of less magnitude.
TABLE 1.- Kidney weight, body weight in gonadectomized (oVx or oRx) and intact rats undergoing uNx (at 90 days) and studied at 150 days.

Values are for one kidney weight only.
n = 5 in each experimental group
The factorial MANOVA analysis showed that gender, gonadectomy, uninephrectomy and its combinations modified kidney weight (p < 0.005 in all cases) whereas body weight gain was modified by gender alone (p < 0.05).

Fig. 1.- Kidney weight recorded at the time of uninephrectomy (uNx, 90 days of age) relative to that registered sixty days later at 150 days age.
Broken line indicates identity line. Female uNx (○), female gonadectomized uNx (Δ) and male gonadectomized uNx (◊) symbols, more close to the identity line, show lower CRG than male uNx (□) whose hypertrophy is greater an their symbols play far away of the identity line.
Male body weight gain, during the time elapsed between uNx and the final study, was greater than in the other groups 35% vs. 13-16%.
It is important to indicate that gonads removal had an effect upon body and kidney weight. Body and kidney weight in male non nephrectomized rats at age 150 days was higher than in orchidectomized rats (433 ± 15 and 1.38 ± 0.05 vs. 350 ± 9 and 1.04±0.02 g, respectively, p < 0.001). Instead, body and kidney weight did not change after ovariectomy in relation to female rats (317 ± 13 and 0.87 ± 0.04 vs. 280 ± 9 and 0.97 ± 0.04, NS, respectively)
Cell size, protein and nuclei content
After uNx, protein content was similar in all the groups except in orchidectomized one: 233.3 ± 4.3, 207.0 ± 18.3 and 192.2 ± 4.9 mg/g kwt for male uNx, female uNx and ovariectomized uNx rats, respectively vs. 132.1 ± 8.9 for orchidectomized uNx , p < 0.05. RNA content, was also smaller for this group: 3.34 ± 0.19, 3.24 ± 0.13 and 4.02 ± 0.033 mg/g kwt for male uNx, female uNx and ovariectomized uNx rats, respectively vs. 1.24 ± 0.02, p < 0.05 for orchidectomized uNx.
The nuclear marker DNA was similar in all uNx groups (1.38 ± 0.09; 1.33 ± 0.08; 1.32 ± 0.05; and 1.33 ± 0.08 mg/g kwt for male uNx, orchidectomized uNx, female uNx and ovariectomized uNx rats).
Final cell size, at age150 days, measured by protein/DNA (Fig. 2) or RNA/DNA ratio (2.47 ± 0.12, 2.47 ± 0.09 and 3.33 ± 0.42) was homogeneous in male uNx, female uNx and ovariectomized uNx groups, since protein and RNA increase was also paralleled by the increase in the DNA content. This increase in DNA in parallel to protein (Fig. 2) or RNA content (0.98 ± 0.04) was absent in orchidectomized uNx group. Accordingly, the factorial MANOVA analysis showed that only in this group, uNx and gonadectomy and gender combination modified both Protein/DNA and RNA/DNA ratios (p < 0.05 and p < 0.001, respectively).

Fig. 2.- Renal cell size (protein/DNA) at age 150 days in the different groups.
Data are means ± SEM. Black columns indicate non-nephrectomized male (left panel) and female (right panel) rats; striped columns, male and female uNx animals; white columns, orchidectomized (oRx) and ovariectomized (oVx) groups and dotted columns oRx and oVx uNx rats.
No statistical difference was observed among non nephrectomized and uNx groups with and whitout gonads removal except for oRx uNx (* p < 0.05) with respect to the other groups.
In fact, number of cells, indicative of hyperplasia, measured by DNA content was about 36% higher in all uNx groups in comparison with the non-nephrectomized ones, 1.32 ± 0.04 vs. 0.96 ± 0.03 mg/g renal cortex, p < 0.001, respectively.
Protein and RNA methods yielded similar cytoplasmatic content estimations (y = 32.9x + 77, r = 0.75, p < 0.05).
Is important to note that histopathological studies did not show abnormal findings -such as infiltrating cells or cellular fibrosis- in any group.
Gonadectomy and uNx effect on blood pressure and renal function
Mean blood pressure ranged from 101 to 114 mmHg for all groups, except for the ovariectomized group. The ovariectomized rats showed the lowest mean blood pressure level, 92.5 ± 8.3 (Table 2).
TABLE 2.- Mean blood pressure (MBP) and GFR at age 150 days in non- and uni- nephrectomized (uNx) rats with and without gonadectomy (oVx or oRx).

n = 5 in each experimental group
The factorial MANOVA analysis showed that uninephrectomy and gender combination modified GFR (*p < 0.05).
Glomerular filtration rate (expressed in ml/min/g kwt) was similar in male and female intact rats (0.99 ± 0.05 and 0.93 ± 0.05, NS), respectively and slightly lower after gonadectomy, NS. After uNx, glomerular filtration rate remained in comparable levels to those described above in male uNx, orchidectomized uNx and ovariectomized uNx. On the contrary, female uNx rats were not able to increase glomerular filtration rate, thus demonstrating a lack of compensatory functional adaptation to unilateral kidney removal (female vs. female uNx, 0.93 ± 0.05 vs. 0.56 ± 0.02, (p < 0.05; Table 2). The uninephrectomy, as well as gonadectomy increased urinary volume, sodium and potassium excretion (p < 0.001 and p < 0.05, respectively), but the combination of both failed to shown an additive effect in factorial MANOVA analysis (Table 3).
TABLE 3.- Functional data at age 150 days in non-and uni-nephrectomized (uNx) rats with and without gonadectomy (oVx or oRx).
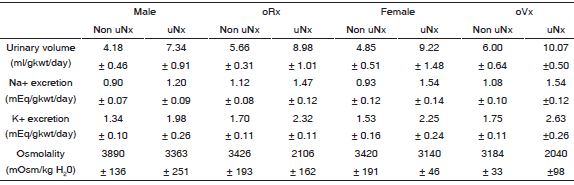
n = 5 in each experimental group
The factorial MANOVA analysis showed that uninephrectomy and gonadectomy modified urinary volume, Na+, K+ excretion and osmolality (p < 0.001 and p < 0.05, respectively) whereas uninephrectomy and gonadectomy combination modified osmolality (p < 0.05).
Both uninephrectomy and gonadectomy diminished urinary osmolality (p < 0.001 in both cases), while its combination showed an additive effect in factorial MANOVA analysis (p < 0.05; Table 3).
Gonadectomy and uNx effect on kallikrein kinin system
Urinary kallikrein activity, shown in Fig. 3, was higher in female than that in male rats independently of uninephrectomy status (p < 0.001), while gonadectomy increased kallikrein activity in male rats (p < 0.05). In this regard, urinary kallikrein activity in orchidectomized groups approached the ovariectomized levels, independently of uNx status.

Fig. 3.- Urinary kallikrein activity (UKa, nkat/g kwt/day) at age 150 days in the different groups.
Data are means ± SEM. Black columns indicate intact male (left panel) and female (right panel) rats; striped columns, male and female uNx animals; white columns, orchidectomized (oRx) and ovariectomized (oVx) groups and dotted columns oRx and oVx uNx rats.
Sexual dimorphism, observed in intact male and female rats is maintained in non-gonadectomized uNx male and female groups (* p < 0.001). Gonadectomy increased kallikrein activity in male rats (** p < 0.01).
Discussion
Present results show that compensatory renal growth is maximum in male rats after two months uNx when compared with uninephrectomized female or gonadectomized groups. Compensatory renal growth, instead, is lowest in uninephrectomized female rats. Protein/DNA and RNA/DNA ratios demonstrate that orchidectomized uNx rats are unable to attain the cell size reached by the other groups, since the nuclei marker increase is not paralleled by cytoplasmatic growth in them. A 36% DNA increase in all the uninephrectomized groups with respect to the non-nephrectomized ones clearly indicates the presence of hyperplasia in the CRG of this model.
Glomerular filtration rate in female uNx rats, as well as CRG, is lower than in the intact female rats. The compensatory renal growth in ovariectomized uNx rats, however, proceeds as in the rest of the groups. It may be that the lack of female sexual hormones allows the remnant kidney to grow normally.
Daily urinary volume, sodium and potassium excretions increase when gonadectomy or uninephrectomy is performed. Both, uninephrectomy and gonadectomy, instead, contribute to diminished urine osmolality. Several authors21-23, working in female experimental animals, in isolated perfused kidney or after estradiol supplementation, measuring diuresis or urine concentration capacity have found an increase in water excretion or a diminished response to an antidiuretic hormone. The same appears to be true in humans24: men excrete more solutes than women in the same urine volume. We found a significant increase in urinary volume excretion with respect to the intact animals as was already described. Spontaneous urine osmolality is lower in gonadectomized uNx animals, this may result from a lack of sexual hormone interaction. Our experimental design was not directed to explore electrolyte and urinary volume excretion, or to test antidiuretic hormone sensitivity. Others25 have also found changes in sodium and water excretion after uNx as we did, but not in glomerular filtration rate.
On the other hand, a low blood pressure sensitive to kallikrein kinin activity in ovariectomized rats has already been described by us26.
Schlondorff et al.27, demonstrated that exogenous testosterone administration increases the compensatory renal growth in female rats. However, male and orchidectomized rats were not affected by this treatment in their experiments. Zeier et al.28, on the contrary, reported that testosterone amplifies CRG in both, male and female uninephrectomized rats. Others29, have shown that the process of kidney hypertrophy after uNx is not accompanied by an increase in the number of cytoplasmatic androgen receptors.
Mulroney et al25, studying morphological and functional changes following 8-10 weeks uninephrectomy in both male and female adult rats, have already described a sexual dimorphism in the remnant kidney growth although they did not include orchidectomized animals in their experimental design. They concluded that testosterone is the driving force for the enhanced CRG since ovariectomized rats improved their response to uninephrectomy after gonadal male steroid supplementation. Besides, Jansson et al.30 reported that the secretory pattern of the growth hormone in the mature rat is sexually differentiated and that the growth promoting effect of androgens is indirect and depends on the specific growth hormone stimulation pattern. Other authors31, 32 have also stated that the secretory pattern of the growth hormone is sexually dimorphic in the adult rat and suggests that the steroid may influence the growth hormone secretory pattern through the hypothalamic neuropeptides. Our data are in agreement with the fact that sexual hormones may influence CRG since we found that CRG is maximum in the uninephrectomized male rats, that the kidney weight turns to be low after orchidectomy, even after uNx, and that CRG is poor in female rats. In order to determine whether these findings could be extended to humans, it appears to be of interest to explore CRG in female kidney donors.
Cell size, according to protein or RNA/ DNA ratios, attained a similar magnitude to their respective non-nephrectomized groups in all uninephrectomized groups, except in the orchidectomized uNx. These findings are in accordance with Liu & Preisig33 proposal that uninephrectomy induces CRG results through a cell cycle mediated mechanism. Cells of the orchidectomized uNx group, however, did not reach the other group cell size, cytoplasmatic protein content was small. It may be thought that the absence of male sexual hormones, which in turn diminishes growth hormone stimulus, contributes to alter the precise equilibrium between the rate of protein synthesis and degradation. Besides, since all uNx groups showed higher levels of DNA in comparison with the intact groups, we are allowed to suggest that some degree of hyperplasia may also contribute to CRG. Although Mulroney et al.34, in studies performed close to uNx, described that the presence of hyperplasia in female compensatory growth after uNx is probably associated to a different pattern in GH secretion and IGF-1 receptor expression than the one observed in male rats. Our studies, made late after uNx, do not allow us to disregard or confirm their results. Liu and Preisig33, working on a shorter experiment and using a different DNA synthesis marker, failed to demonstrate an increase in the number of nuclei in the remnant kidney.
Mackie et al.35 described an increase in intrarrenal bradykinin peptides in the intact portion of the 5/6 remnant kidney undergoing 2-7 week hypertrophy not entirely attributed to an inflammatory process. In our experimental model no relation between compensatory renal growth and kallikrein kinin system activity was found. A sex dimorphic pattern for urinary kallikrein activity was found in intact animals, as already described11. Urinary kallikrein activity was higher in female than male rats and this finding persisted after uninephrectomy. This sex dimorphism for kallikrein kinin system disappeared in the orchidectomized groups, uninephrectomized or not. In fact, urinary kallikrein activity in orchidectomized rats -with or without uNx- turns to be similar to female groups. The increase of urinary kallikrein activity that follows orchidectomy strongly suggests that a male sexual hormone represses kallikrein activity.
Our finding show that CRG strongly differs between sexes. In female and gonadectomized rats the compensatory growth and function is lower than in male uNx animals. In all groups, but not in the orchidectomized uNx the cell size is similar showing that the lack of male sexual hormones influences negatively renal hypertrophy. Hypertrophy is present in the rest of the groups accompanied by some degree of hyperplasia. The kallikrein kinin system does not appear to be linked to CRG at least at 2 months after uNx.
Acknowledgments: This work was supported by grants from Universidad de Buenos Aires, Ministerio de Ciencia y Técnica and Consejo Nacional de Investigaciones Científicas y Técnicas, Buenos Aires, Argentina. The authors want to thank Dr. Marina Khoury for the statistic assistance.
Conflicts of interest: The authors declare to have no conflicts of interest.
1. Arrizurieta-Muchnik EE, Lipham EM, Gottschalk CW. Form and function in normal and hypertrophied nephrons. In: Nowiski WW, Goss RJ, eds. Compensatory Renal Hypertrophy. New York: Academic Press, 1969, p 29-44. [ Links ]
2. Miller PL, Rennke HG, Meyer TW. Glomerular hypertrophy accelerates hypertensive glomerular injury in rats. Am J Physiol (Renal Fluid Electrolyte Physiol) 1991; 261: F459-65. [ Links ]
3. Arima S, Kohagura K, Xu HL, et al. Nongenomic vascular action of aldosterone in the glomerular microcirculation. J Am Soc of Nephrol 2003; 14: 2225-63. [ Links ]
4. Ortola FV, Ballerman BJ, Brenner BM. Endogenous ANP augments fractional excretion of Pi, Ca and Na in rats with reduced renal mass. Am J Physiol 1988; 255: F1091-7. [ Links ]
5. Zhang PL, Mackenzie HS, Troy JL, Brenner BM. Effect of natriuretic receptor inhibition on remnant kidney function in rats. Kidney Int 1994; 46: 414-20. [ Links ]
6. Benigni A, Perico N, Gaspari F, et al. Increased renal endothelium production in rats with reduced renal mass. Am J Physiol 1991; 260: F 331-9. [ Links ]
7. Orisio S, Benigni A, Bruzzi I, et al. Renal endothelium gene expression is increased in remnant kidney and correlates with disease progression. Kidney Int 1993; 43: 354-8. [ Links ]
8. Nath KA, Chmielewski DH, Hostetter TH. Regulatory role of prostanoids in glomerular microcirculation of remnant nephrons. Am J Physiol 1987; 252: F 829-37. [ Links ]
9. Pelayo JC, Shanley PF. Glomerular and tubular adaptive responses to acute nephron loss in the rat. Effect of prostaglandin synthesis inhibition. J Clin Invest 1990; 85: 1761-9. [ Links ]
10. Mackie FE, Campbell DJ, Meyer TW. Intrarenal angiotensin and bradykinin peptide levels in the remnant kidney model of renal insufficiency. Kidney Int 2001; 59: 1458-65. [ Links ]
11. Oddo EM, De Luca Sarobe V, Krmar R, et al. Effect of prepuberal gonadectomy upon aldosterone levels in female and male SHR: Interaction between blood pressure and kallikrein kinin system. Clin Exp Hypertens 2006; 28: 157-70. [ Links ]
12. Martín R, Nesse A, Arrizurieta de Muchnik E. Urinary kallikrein excretion, renal kallikrein content and renal plasma flow in the rat. Medicina (Buenos Aires) 1979, 39: 467-79. [ Links ]
13. Young MK, Raisz LG. The anthrone method. Proc Soc Exp Biol Med 1952, 80: 771. [ Links ]
14. Huang HC, Preisig PA. G1 kinases and transforming growth factor beta signalling are associated with a growth pattern switch in diabetes-induced renal growth. Kidney Int 2000; 58 : 162-72. [ Links ]
15. Winick M, Noble A. Quantitative change in DNA, RNA and protein during prenatal and postnatal growth in the rat. Development Biol 1965; 12: 451-66. [ Links ]
16. Widdowson EM, Crabb DE, Milner RDG. Cellular development of some human organs before birth. Arch Dis Child 1972; 47: 652-5 [ Links ]
17. Schmidt EE, Schibler U. Cell size regulation, a mechanism that controls cellular RNA accumulation: consequence on regulation of the ubiquitous transcription factor Oct1 and NF-Y and liver enriched transcription factor DBP. J Cell Biol 1995; 128: 467- 83 [ Links ]
18. Admunsen E, Putter J, Frieberger P, Knos M, Larsbraten M, Cleason G. Methods for of glandular kallikreins by means of a chromogenic tripeptide substrate. Adv Exp Med Biol 1979; 120: 83-5. [ Links ]
19. Bradford MM. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye. Anal Biochem 1976; 72: 248-54. [ Links ]
20. Otto P, Kephart D, Bitner R, Huber S, Volkerding K. Separate isolation of genomic DNA and total RNA from single samples using the SV total RNA isolation system. Promega Notes 1998; 69: 19-25. [ Links ]
21. Pellanda EB. Diuretic action of estradiol. Ann Endocrinol 1976; 37: 235-40. [ Links ]
22. Carlberg KA, Fregly MJ, Fahey M. Effects of chronic estrogen treatment on water exchange in rats. Am J Physiol 1984; 247: E101-10. [ Links ]
23. Wang YX, Crofton JT, Liu H, Sato K, Brooks DP, Share L. Estradiol attenuates the antidiuretic action of vasopressin in ovariectomized rats. Am J Physiol 1995; 268: R951-7. [ Links ]
24. Perucca J, Bouby N, Valeix P, Banker L. Sex difference in urine concentration across differing ages, sodium intake, and level of kidney diseases. Am J Physiol Regul Integr Comp Physiol 2007; 292: R700-5. [ Links ]
25. Mulroney SE, Woda C, Johnson M, Pesce C. Gender difference in renal growth and function after uninephrectomy in adult rats. Kidney Int 1999; 56: 944-53. [ Links ]
26. Azurmendi PJ, Oddo EM, Obika LF, et al. Gonadectomy influences blood pressure through the kallikrein-kinin system. Kidney Blood Press Res 2009; 32: 342-8. [ Links ]
27. Schlondorff D, Trizna W, De Rosis E, Korth-Schutz S. Effect of testosterone on compensatory renal hypertrophy in the rat. Endocrinol 1977; 101: 1670-5. [ Links ]
28. Zeier M, Schönherr R, Amann K, Ritz E. Effects of testosterone on glomerular growth after uninephrectomy. Nephrol Dial Transplant 1988; 13: 2234-40. [ Links ]
29. Tremblay RR, Ho-Kim MA, Dubé JY. Measurements of cytoplasmatic androgen receptor in compensatory renal hypertrophy in the rat. Ann Endocrinol (Paris) 1983; 44: 253-55. [ Links ]
30. Jansson JO, Edén S, Isaksson O. Sexual dimorphism in the control of growth hormone secretion. Endocr Rev 1985; 6: 128-50. [ Links ]
31. Chowen JA, García-Segura LM, González-Parra S, Argente J. Sex steroid effects on the development and funtioning of the growth hormone axis. Cell Mol Neurobiol 1996; 16: 297-310. [ Links ]
32. Chowen JA, Frago LM, Argente J. The regulation of GH secretion by sex steroids. Eur J Endocrinol 2004; 151 Suppl 3: U95-100. [ Links ]
33. Liu B, Preisig PA. Compensatory renal hypertrophy is mediated by a cell cycle-dependant mechanism. Kidney Int 2002; 62: 1650-8. [ Links ]
34. Mulroney SE, Pesce C. Early hyperplastic renal growth after uninephrectomy in adult female rats. Endocrinol 2000; 141: 932-7 [ Links ]
35. Mackie FE, Campbell DJ, Meyer TW. Intrarrenal angiotensin and bradykinin peptide levels in the remnant kidney model of renal insufficiency. Kidney Int 2001; 59: 1458-65. [ Links ]
Received 18-VI-2013
Accepted: 30-VII-2013

