










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEl hornero
versión impresa ISSN 0073-3407versión On-line ISSN 1850-4884
Hornero v.18 n.1 Buenos Aires ene./ago. 2003
Emplumando dinosaurios: la transición evolutiva de terópodos a aves
Luis M. Chiappe 1 y Alexander Vargas 2,3
1 Department of Vertebrate Paleontology, Natural History Museum of Los Angeles County. 900 Exposition Boulevard, Los Angeles, CA 90007, EEUU. chiappe@nhm.org
2 Departamento de Morfología, Facultad de Medicina, Universidad de Chile. Independencia 1027, Casilla 70079, Santiago 7, Santiago, Chile.
3 Sección Paleontología, Museo Nacional de Historia Natural. Casilla 787, Santiago, Chile.
Resumen
Desde el inicio del pensamiento evolutivo, el origen de las aves ha constituido uno de los problemas más polémicos de la biología evolutiva. En la actualidad, un gran cúmulo de evidencia indica que dicho problema se encuentra de gran manera resuelto. La evidencia osteológica, oológica, etológica y tegumentaria sustenta fuertemente la hipótesis que las aves descienden de dinosaurios terópodos, en particular a partir de formas clasificadas dentro de Maniraptora. Los contados investigadores que hoy en día se oponen a dicha hipótesis han sustentado sus críticas en observaciones erróneas o dentro de un marco teórico obsoleto. Este trabajo ofrece una breve discusión tanto de las principales líneas de evidencia en sustento de esta hipótesis filogenética como de los argumentos utilizados en su contra.
Palabras clave: Aves, origen, evolución, Mesozoico.
Abstract
Feathering dinosaurs: the transition from theropods to birds
Since the beginning of evolutionary thinking, the origin of birds has constituted one of the most debated problems in evolutionary biology. Today, a large body of evidence indicates that such a problem is to a great extent solved. Osteological, oological, ethological, and integumentary evidence strongly supports the hypothesis that birds evolved from theropod dinosaurs, in particular from dinosaurs classified within Maniraptora. Today's handful of researchers opposing this hypothesis have either based their criticisms on mistaken observations or they have framed them within dated methodologies. This paper provides a brief discussion of both the evidence in support to this hypothesis and the main arguments against it.
Key words: Aves, origin, evolution, Mesozoic.
El origen de las aves ha sido el foco de gran controversia científica durante toda la historia de la taxonomía y la biología evolutiva. Ya en el siglo XVIII, las aves eran con frecuencia listadas a continuación de los peces voladores en las "cadenas de la vida" que los naturalistas de ese tiempo solían construir. Con el establecimiento del pensamiento evolutivo, especialmente luego de que Charles Darwin formulara sus ideas de la evolución mediante la selección natural, fueron propuestas numerosas y diferentes hipótesis sobre el origen de las aves. Desde entonces, las aves han sido consideradas como los parientes cercanos de diversos linajes de reptiles, incluidas las tortugas, los lagartos, los cocodrilomorfos (cocodrilos modernos y sus parientes triásicos), diversos arcosaurios y arcosauromorfos basales, los pterosaurios o reptiles voladores del Mesozoico, y los dinosaurios, tanto ornitisquios como terópodos (Fig. 1).
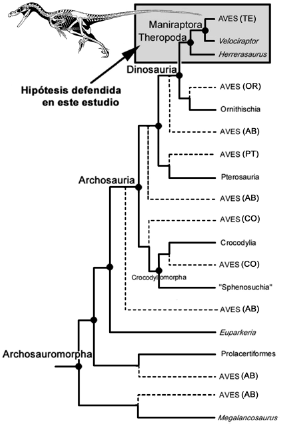
Figura 1. Hipótesis que enmarcan al origen de las aves dentro de los reptiles arcosauromorfos. AB: hipótesis que relacionan a las aves con ciertos arcosaurios basales o arcosauromorfos basales; CO: hipótesis que sustentan el origen de las aves a partir de cocodrilos o cocodrilomorfos; PT: hipótesis en defensa de un origen común entre aves y pterosaurios; OR: hipótesis que relacionan a las aves con los dinosaurios ornitísquios; TE: hipótesis de un origen a partir de los dinosaurios terópodos (la hipótesis defendida en este trabajo).
La historia de este fascinante debate científico ha sido sintetizada en varias otras publicaciones, entre ellas los trabajos de Witmer 1,2, Padian y Chiappe 3, y Chiappe 4. Las discusiones actuales sobre el origen de las aves tienen un claro punto de partida en el trabajo de John Ostrom, quien a principios de la década de 1970 revitalizara la teoría del origen de las aves a partir de los dinosaurios terópodos, una propuesta originalmente formulada poco después de que Darwin escribiera el "Origen de las especies". En los 30 años transcurridos desde los estudios iniciales de Ostrom, un gran cuerpo de evidencia ha sido acumulado en favor de esta idea 5,6. Hoy en día, varias líneas diferentes de evidencia convergen proveyendo sustento a esta hipótesis, si bien el linaje preciso de dinosaurios terópodos más cercanamente emparentado a las aves aún se encuentra bajo debate. Este enorme cuerpo de evidencia ha dado lugar a la idea que las aves modernas no son otra cosa que dinosaurios terópodos emplumados, con cola corta. En este trabajo, nosotros brindamos una muy breve síntesis de dicha evidencia y discutimos las críticas más frecuentes de la hipótesis que argumenta que las aves son los descendientes de los dinosaurios terópodos.
La evidencia
El cuerpo de evidencia que sustenta la hipótesis que las aves se encuentran filogenéticamente dentro de los dinosaurios terópodos está basado en comparaciones de su osteología, su oología (el estudio de los huevos), su comportamiento y su integumento, con aquellos de una variedad de terópodos no-avianos. Fundamentalmente, estas comparaciones utilizan linajes que se clasifican dentro de los Maniraptora, el grupo de terópodos que incluye a los dromaeosáuridos, oviraptóridos, troodóntidos y therizinosáuridos. A continuación se discute brevemente cada una de estas líneas de evidencia.
Osteología
Un gran número de caracteres osteológicos derivados o sinapomorfías son compartidos entre las aves y todos, o algunos, maniraptores no-avianos (Fig. 2). Las comparaciones entre estos taxa han sido asistidas de gran manera gracias al descubrimiento de una enorme cantidad de fósiles de aves primitivas 7, los que poseen morfologías levemente modificadas con respecto a la condición ancestral exhibida por dichos maniraptores no-avianos. Estos caracteres incluyen la presencia de recesos timpánicos rostrales, dorsales y caudales (i.e., áreas pneumatizadas contenidas dentro de los huesos que rodean la cavidad ótica y que se encuentran conectadas a esta última). También incluyen la presencia de pronunciados procesos ventrales en las vertebras cervico-dorsales, costillas ventrales osificadas (las costillas que se articulan al esternón), miembros anteriores cuya longitud es superior a la mitad de la longitud de los miembros posteriores, un hueso carpal con forma de medialuna (una estructura que permite desarrollar los movimientos de "plegamiento" que caracterizan al carpo de las aves), clavículas fusionadas formando una fúrcula o "hueso de la suerte" (este carácter corresponde a una sinapomorfía de un grupo de dinosaurios terópodos aún más grande), la presencia en el ilion de un pedúnculo para el pubis más largo que el pedúnculo para el isquion (estos pedúnculos forman los márgenes anterior y posterior del acetábulo), un pubis que se orienta verticalmente o caudo-ventralmente y cuyo extremo distal forma una expansión en forma de bota que se proyecta posteriormente, un isquión que tiene dos tercios o menos de la longitud del pubis, un fémur con solo un débil cuarto trocánter (el punto de inserción de la musculatura caudo-femoral) y muchos otros caracteres distribuidos a lo largo del esqueleto 8-11.
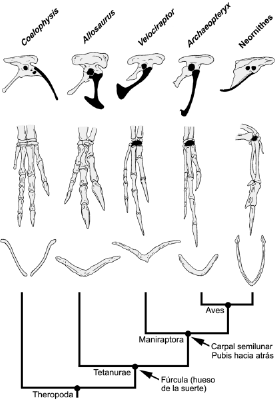
Figura 2. Algunas de las sinapomorfías osteológicas que sustentan el origen de las aves a partir de los terópodos maniraptores.
Oología
En general, la combinación del aspecto externo y la microestructura de los huevos de cáscara calcificada de distintos grupos de reptiles, tanto vivientes como extinguidos, es específica para cada uno de éstos, y dicha combinación permite diferenciar unos huevos de otros 12,13. Hasta hace poco, las características típicas de los huevos de los terópodos no-avianos eran desconocidas, ya que no se habían encontrado huevos fósiles que pudieran asignarse con exactitud a estos dinosaurios; en ninguna instancia se había encontrado un huevo conteniendo restos embrionarios que pudieran ser identificados como aquellos de algún linaje de terópodos no-avianos. El descubrimiento de un embrión de oviraptórido dentro de su propio huevo en rocas del Cretácico tardío del desierto de Gobi produjo la primera evidencia de la morfología de los huevos de estos dinosaurios 14,15. Luego de ése, fueron descubiertos otros embriones de maniraptores no-avianos. Estos incluyeron tanto a otras especies de oviraptóridos como también a troodóntidos 16. Estudios comparativos de la forma general y la microestructura de la cáscara de estos huevos y los de las aves modernas han revelado una serie de rasgos comunes y exclusivos de estos animales (Fig. 3). Una de estas sinapomorfías es la presencia de más de una capa diferenciada en la microestructura de la cáscara de huevo 17. Típicamente, la cáscara de huevo de terópodos no-avianos y aves se caracteriza por poseer unidades cuya porción interna está formada por una estructura cristalina de calcita que se dispone radialmente a partir de un núcleo, a veces llamado el núcleo orgánico. Dichas unidades también poseen una zona más externa que, en cortes delgados, tiene un aspecto laminar. Hasta la fecha, las cáscaras de huevo de maniraptores no-avianos conocidas poseen solo dos capas estructurales, mientras que los huevos de aves presentan tres o cuatro capas. No obstante, ningún otro reptil cuyos huevos tienen cáscaras calcificadas posee una zonación semejante 12,17.
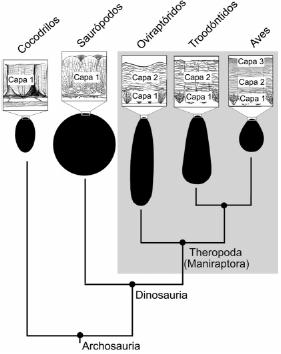
Figura 3. Cladograma ilustrando parte de la evidencia oológica en sustento de un origen de las aves a partir de dinosaurios terópodos. Nótese la presencia de dos o más capas en la cáscara de huevo de oviraptóridos, troodóntidos y aves, y la asimetría de la forma del huevo en los últimos dos grupos.
Etología
Muy ocasionalmente el registro fósil preserva algún tipo de evidencia sobre el comportamiento de organismos extinguidos. Sin embargo, unos pocos y espectaculares descubrimientos en rocas del Cretácico tardío de Mongolia y Estados Unidos han brindado información muy significativa sobre la conducta de nidificación de ciertos linajes de maniraptores no-avianos. Varios esqueletos de oviraptóridos del desierto de Gobi, aparentemente pertenecientes a Citipati osmolskae 18, fueron descubiertos encima de sus nidadas de huevos, con las patas plegadas dentro de un espacio sin huevos en el centro de la puesta y los brazos extendidos alrededor de la periferia de la misma 15 (Fig. 4). La morfología de los huevos encontrados con estos ejemplares es idéntica a la del huevo conteniendo al embrión de oviraptórido mencionado más arriba, hecho que, por un lado, sustenta la idea que los ejemplares adultos perecieron durante el período de empollamiento de sus huevos y, por otro lado, sugiere que el embrión podría pertenecer al oviraptórido Citipati osmolskae. Estos hallazgos han hecho reevaluar inferencias basadas en el holotipo de Oviraptor philoceratops 19, la especie tipo de los oviraptóridos, ejemplar que fuera descubierto a principios de la década de 1920 en estratos contemporáneos a los de Citipati osmolskae. El holotipo de Oviraptor philoceratops también fue encontrado encima de una puesta de huevos pero, en este caso, dicha asociación fue erróneamente interpretada como evidencia de depredación 19,20. Los oviraptóridos del desierto de Gobi no han sido los únicos dinosaurios que se encontraran "sentados" encima de puestas de huevos. Rocas del Cretácico tardío de Montana han brindado un esqueleto adulto del troodóntido Troodon formosus, también encontrado en contacto con una puesta de huevos cuya morfología es idéntica a huevos de los mismos estratos conteniendo embriones aparentemente pertenecientes a esta misma especie 21.
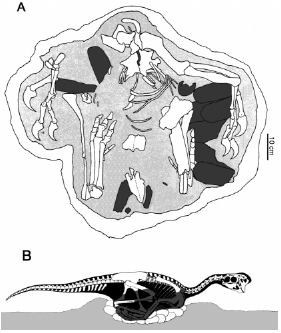
Figura 4. Esqueleto parcial de un oviraptórido del Cretácico tardío de Mongolia empollando su puesta de huevos (A) y reconstrucción de la posición de este animal en vida (B). Modificado de Clark et al. 15.
Todos estos raros y significativos hallazgos han documentado que, independientemente de su función específica (sea protección, incubación u otra), el típico comportamiento aviano de empollamiento (el comportamiento en el cual los adultos se sientan sobre sus propias puestas por prolongados períodos) era común entre los maniraptores no-avianos.
Tegumento
Las plumas siempre han sido la característica aviana por excelencia y un rasgo supuestamente exclusivo de las aves. Sin embargo, en los últimos años se ha descubierto un gran número de dinosaurios emplumados en los depósitos del Cretácico temprano del noreste de China 22 (Fig. 5). Estos dinosaurios incluyen el coelurosaurio basal Sinosauropteryx prima 24, el terizinosáurido Beipiaosaurus inexpectus 25, los oviraptorosaurios Caudipteryx zoui 26 y Caudipteryx dongi 27, los dromaeosáuridos Sinornithosaurus millenii 25, Microraptor zhaoianus 28 y Microraptor gui 29, y el enigmático Protarchaeopteryx robusta 26, cuya relación filogenética es más incierta. Todas estas especies pueden ser claramente clasificadas como dinosaurios terópodos y, con excepción de Sinosauropteryx prima, todos corresponden a maniraptores no-avianos. Mientras que las plumas de Sinosauropteryx prima son simples filamentos que exhiben un incipiente grado de ramificación, se encuentran preservadas plumas con raquis y vanos (pennáceas) en ejemplares de Caudipteryx zoui, Caudipteryx dongi, Protarchaeopteryx robusta y Microraptor gui. También se han encontrado plumas semejantes rodeando el esqueleto de un dromaeosáurido juvenil 30, que posiblemente corresponda a Sinornithosaurus millenii. En general, estos dos tipos de plumas se encuentran cubriendo gran parte del cuerpo de estos terópodos. En ambas especies de Caudipteryx, una serie de plumas remigias se inserta en el extremo distal del miembro anterior y, al final de la cola, un penacho de plumas rectrices forma un extenso abanico. En el pequeño Microraptor gui, las plumas del miembro anterior forman una amplia superficie alar, lo que hace pensar que esta forma también podría haber sido capaz de volar. Además, a diferencia de todos los restantes dinosaurios emplumados, las plumas pennáceas de Microraptor gui tienen vanos asimétricos 29, las que usualmente son interpretadas como evidencia de habilidades aerodinámicas 31. Curiosamente, plumas semejantes también se insertan en el miembro posterior de esta especie, lo que ha llevado a imaginar que Microraptor gui podría haber utilizado los miembros posteriores como un segundo par de "alas" 29, si bien en nuestra opinión esta es una propuesta dudosa desde un punto de vista funcional. El profundo significado de estos hallazgos debe verse a la luz de las interpretaciones filogenéticas propuestas para estos taxa (Fig. 6), ya que todas estas especies han sido clasificadas fuera de las aves y, como hemos visto, en diversos linajes de terópodos no-avianos (con excepción de Sinosauropteryx prima, que es considerado un terópodo aún más primitivo). Estos hallazgos no solo han documentado la presencia de plumas en animales que no son aves, sino que también han sugerido que algunos terópodos no-avianos (e.g., Microraptor gui) eran capaces de volar.
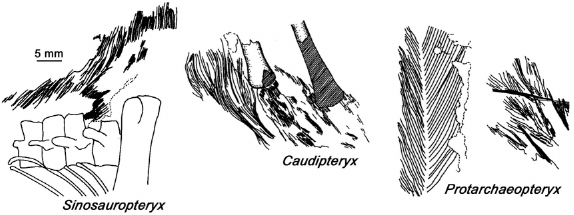
Figura 5. Esquemas del tegumento de varios dinosaurios terópodos del Cretácico temprano de China. Nótese la presencia de plumas con vanos en Protarchaeopteryx. Modificado de Padian et al. 23.
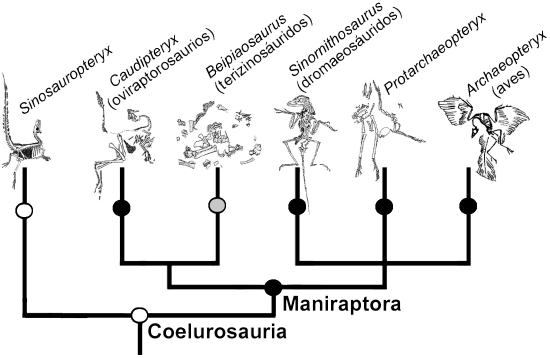
Figura 6. Cladograma ilustrando las relaciones filogenéticas entre varios grupos de dinosaurios emplumados. Los círculos blancos indican la presencia de plumas filamentosas, los negros la presencia de plumas con vanos y los grises la presencia inferida de plumas con vanos. Mientras que la presencia de plumas filamentosas es una sinapomorfía de terópodos coelurosaurios, la presencia de vanos es una sinapomorfía de terópodos maniraptores. Modificado de Chiappe y Dyke 7.
Las críticas
La "paradoja temporal"
Ciertos críticos del origen de las aves a partir de maniraptores no-avianos han enfatizado la aparente discrepancia entre el registro fósil de estos dinosaurios y el de las aves, y básicamente el hecho que los maniraptores frecuentemente utilizados en las discusiones del origen de las aves (Deinonychus antirhopus, Velociraptor mongoliensis, Oviraptor philoceratops) son muchos millones de años más jóvenes que Archaeopteryx lithographica, el ave más antigua conocida 32. Dicha crítica es en general conocida como la "paradoja temporal", ya que realza la supuesta inconsistencia de argumentar que las aves descendieron de animales que vivieron mucho después que el origen de las mismas. No obstante, esta crítica es insustancial, ya que nunca nadie ha defendido la posición que se critica (i.e., la idea de que especies como Velociraptor mongoliensis u Oviraptor philoceratops fueron los antecesores directos de las aves). Como la hipótesis del origen de las aves a partir de maniraptores no-avianos se encuentra enmarcada en la metodología cladista, las especies recién mencionadas son interpretadas como grupos externos a las aves y sus relaciones con éstas son de grupo-hermano y no de ancestro-descendiente 3. Además, los partidarios de la "paradoja temporal" frecuentemente pasan por alto que varias especies de coelurosaurios (el grupo de terópodos que incluye a los maniraptores) provienen de capas jurásicas más antiguas que aquellas que contienen a los ejemplares de Archaeopteryx lithographica 11 e, incluso, que algunos de estos coelurosaurios han sido clasificados como maniraptores 33,34. Si bien la evidencia de coelurosaurios más antiguos que Archaeopteryx lithographica es aún limitada, es importante tener en cuenta que el volumen de depósitos continentales (con dinosaurios) del Jurásico medio y temprano es sustancialmente menor que el correspondiente al Jurásico tardío o al Cretácico (Fig. 7).

Figura 7. Distribución estratigráfica de varios linajes de terópodos coelurosaurios (abajo) y cantidad de localidades (loc./ma.) con dinosaurios por cada serie estratigráfica (arriba). Nótese cómo algunos de los terópodos coelurosaurios se encuentran registrados en depósitos más antiguos que los que contienen a Archaeopteryx lithographica (círculo) y cuántas menos localidades con restos de dinosaurios se conocen para el Jurásico medio y temprano. Modificado de Clark et al. 11.
La controversia embriológica
Otro argumento en contra del origen de las aves a partir de dinosaurios terópodos se enfoca en la embriología de la mano de las aves vivientes. La mano de estas aves presenta inicialmente cinco condensaciones precartilaginosas, de las cuales solo tres (las correspondientes a las condensaciones 2, 3 y 4) se osifican y llegan a constituir la mano tridáctila de las aves adultas 35,36. En terópodos basales como Herrerasaurus o Coelophysis, del Triásico tardío de Argentina y Estados Unidos, respectivamente, es posible observar la presencia de cinco osificaciones digitales, de las cuales las correspondientes a los dígitos 4 y 5 se encuentran notablemente reducidas. Los tres dígitos más anteriores (1, 2 y 3) de estos dinosaurios tienen la misma fórmula falangeal, y (en el caso de Coelophysis) las mismas proporciones relativas, que los tres dígitos de la mano tridáctila presente en terópodos maniraptores no-avianos y en Archaeopteryx lithographica 3. Sobre la base de esta evidencia paleontológica, se ha inferido que, a diferencia de las aves vivientes, la mano de los terópodos más basales debió desarrollar sus tres dígitos principales (los dígitos funcionales) a partir de las condensaciones digitales 1, 2 y 3. Esta importante diferencia, el desarrollo de los dígitos 2, 3 y 4 en aves vivientes, tal como es indicado por los estudios embriológicos más modernos 35,36, y el desarrollo de los dígitos 1, 2 y 3, inferido a partir de la evidencia paleontológica, ha sido fuertemente esgrimida por los críticos de la hipótesis de un origen de las aves a partir de dinosaurios terópodos. Estos críticos han frecuentemente extrapolado el desarrollo de los dígitos de las aves vivientes a todas las aves extinguidas, incluso a aquellas que, como Archaeopteryx lithographica, vivieron hace más de 100 millones de años 35,37,38. Utilizando esta extrapolación, dichos críticos han aducido que, dada la inconsistencia entre el desarrollo de la mano de "todas" las aves y el inferido para los dinosaurios terópodos, las primeras no pueden haber descendido a partir de los últimos 35,37,38.
Si bien existe un interesante problema que resolver, la lógica utilizada por estos críticos 35,37,38 es inconsistente, ya que no se dispone de ninguna evidencia embriológica que pueda identificar los dígitos de la mano de aves primitivas como Archaeopteryx lithographica, cuya morfología general es prácticamente idéntica a la de terópodos dromaeosáuridos y otros maniraptores no-avianos 39. A su vez, este argumento carece de sustento metodológico, dado que el rechazo de la hipótesis de un origen a partir de maniraptores sobre la base de las diferencias aparentes en el desarrollo de la mano contradice los postulados de la metodología cladista (de hecho, el paradigma de la inferencia filogenética), la cual enfatiza la importancia de la parsimonia a la hora de optar entre hipótesis filogenéticas alternativas. En otras palabras, si esta aparente diferencia ontogenética se utilizara para argumentar que las aves se han originado a partir de alguna forma que no es un dinosaurio terópodo, el gran número de similitudes osteológicas, tegumentarias, oológicas y etológicas entre las aves y los maniraptores no-avianos tendría que ser explicado como el resultado de numerosos eventos de convergencia evolutiva.
Si las diversas líneas sintetizadas más arriba proveen evidencias muy sólidas a favor del origen de las aves a partir de los dinosaurios terópodos, ¿es posible que la mano de las aves tenga un desarrollo significativamente diferente al de estos dinosaurios? Una forma en la cual esta diferencia embriológica se ha explicado se centra en un patrón ontogenético conocido como "corrimiento de marco". Dentro de este marco teórico, se ha hipotetizado que el desarrollo de la mano de la aves vivientes se ha transformado durante su evolución, de manera que los tres dígitos funcionales que previamente se desarrollaban a partir de las condensaciones 1, 2 y 3, en las aves vivientes se desarrollan a partir de las condensaciones 2, 3 y 4 40. Estudios sobre el desarrollo de los miembros en tetrápodos indican que este tipo de transformación es posible. Por ejemplo, en Hemiergis quadrilineata, una especie de lagarto de la familia Scincidae, los dígitos cuya morfología corresponde a la de los dígitos 2 y 3 de las formas ancestrales se desarrollan a partir de las condensaciones 3 y 4 41 (Fig. 8), constituyendo un claro antecedente de un cambio muy similar al postulado durante la evolución de las aves. La experimentación en el laboratorio también ha demostrado que la identidad de las condensaciones digitales en las extremidades de embriones de pollos domésticos (Gallus gallus) no es una propiedad fija y autónoma de éstos, sino que las condensaciones pueden ser alteradas para adoptar la identidad de otra condensación a través de manipulaciones embriológicas o moleculares 42. Sin embargo, es justo decir que la hipótesis del "corrimiento de marco" no se encuentra libre de críticas. Por ejemplo, Galis et al. 43 creen que dicho corrimiento es imposible. Estos autores han argumentado que la principal dificultad en aceptar un corrimiento de marco reside en que dicho cambio carece de un valor adaptativo aparente. No obstante, no es claro por qué un cambio ontogenético de este tipo podría ocurrir sólo si fuese adaptativo; después de todo, uno podría replicar con el hecho de que un proceso de cuello de botella o de efecto fundador podría ser capaz de imponer una variación neutra en una población (i.e., una que no contenga ningún valor adaptativo). Más allá de estas disquisiciones teóricas, lo cierto es que existen incontables ejemplos que documentan la evolución de desarrollos ontogenéticos y de cómo estas transformaciones pueden ocurrir sin alterar la morfología o función de la estructura en cuestión 44,45. Estos ejemplos demuestran que, filogenéticamente, dichas estructuras son homólogas y no el resultado de una convergencia evolutiva. El mecanismo de neurulación de la especie modelo Brachydanio rerio (pez cebra) difiere del de otros vertebrados y del de los cefalocordados, ya que su cavidad interna se desarrolla por cavitación y no por el descenso de la placa neural. Sin embargo, esta diferencia jamás ha sido utilizada para cuestionar que la neurula del pez cebra no sea homóloga a las neurulas de otros cordados, o que Brachydanio rerio deba ser clasificado como un cordado. De la misma manera, la similitud morfológica de los dígitos de Deinonychus antirhopus y Archaeopteryx lithographica 39, enmarcada dentro de las muchas otras similitudes entre estos organismos, sugiere que las estructuras de la mano de estos animales representan homologías, más allá de que hubieran tenido o no (ya que no lo sabemos) un desarrollo ontogenético diferente.
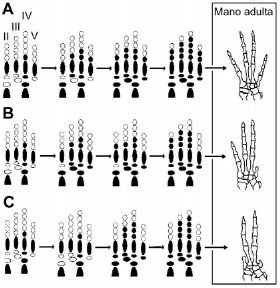
Figura 8. Patrones de condrificación de los elementos de la extremidad anterior de lagartos del género Hemiergis y homeosis de la fórmula falangeal en Hemiergis quadrilineata. En Hemiergis perioni existen morfos de 3 y 4 dígitos (A, B), en los cuales las condensaciones digitales 2 y 3 dan origen a los dos dígitos más anteriores, los que se componen de 3 y 4 falanges, respectivamente. En Hemiergis quadrilineata (C), las condensaciones digitales que dan origen a los dígitos de 3 y 4 falanges (los únicos dígitos de esta especie) corresponden a las condensaciones 3 y 4. Por lo tanto, mientras que en Hemiergis perioni los dígitos más anteriores, con 3 y 4 falanges, se osifican de las condensaciones 2 y 3, en Hemiergis quadrilineata lo hacen de las condensaciones 3 y 4. Modificado de Shapiro 41.
En suma, la aparente diferencia en la embriogénesis de la mano de las aves vivientes y la de los terópodos no-avianos requiere ser explicada, pero el vínculo evolutivo entre estos organismos no debería ser cuestionado sobre la base de especulaciones impregnadas de teoría e inconsistentes con el paradigma metodológico actual.
Estructura y ventilación pulmonar
Sobre la base de estructuras interpretadas como indicadores de compartimentalización visceral en el coelurosaurio basal Sinosauropteryx prima y de la morfología general del pubis de los dinosaurios terópodos, Ruben et al. 46 reconstruyeron el mecanismo de ventilación pulmonar de estos dinosaurios de forma semejante al de los cocodrilos actuales (i.e., uno en el cual los pulmones se inflan a consecuencia de la presión diferencial creada por el desplazamiento del hígado bajo la acción de una serie de músculos que se originan en el pubis). Ruben et al. 46 argumentaron que su interpretación implicaba serios impedimentos para la hipótesis de un origen de las aves a partir de dinosaurios terópodos, ya que la transición desde un sistema pulmonar semejante al de los cocodrilos al sistema de flujo unidireccional característico de las aves habría requerido la evolución de una apertura en la supuesta separación entre las cavidades abdominal y torácica de las formas intermedias, y dicha apertura (o hernia diafragmática) habría comprometido la eficiencia del sistema pulmonar de dichas formas intermedias.
Al igual que las discutidas previamente, esta crítica se fundamenta en nociones erróneas. En primer lugar, estudios detallados del esqueleto de Sinosauropteryx prima utilizado por Ruben et al. 46 han indicado que la estructura interpretada como el septo (diafragma) que separa las cavidades abdominal y torácica es un artificio de la preservación del ejemplar 47. Además, la evidencia osteológica indica que varias de las estructuras óseas correlacionables con el sistema respiratorio y el mecanismo de ventilación de los pulmones de las aves se encontraban ya presentes en maniraptores no-avianos (y algunas, incluso, en terópodos más primitivos). Por ejemplo, los forámenes neumáticos para la entrada de divertículos, extensiones de los sacos aéreos pulmonares, son comunes en las vértebras de terópodos no-avianos. A su vez, la presencia de articulaciones intracostales que delimitan segmentos dorsales y ventrales en las costillas torácicas se encuentra bien documentada en dromaeosáuridos y oviraptóridos 15, y en éstos y en otros maniraptores las placas esternales son relativamente grandes. De esta manera, la morfología de las costillas y del esternón de estos terópodos sugiere que los movimientos costales y esternales coordinados que ventilan los pulmones de las aves actuales podrían ya haber estado presentes en sus ancestros maniraptores.
Es justo decir que no sabemos si los terópodos no-avianos poseyeron un septo que dividía la cavidad visceral semejante al diafragma de los cocodrilos vivientes, pero la presencia en estos dinosaurios de características óseas relacionadas al sistema de ventilación y estructura pulmonar de las aves es innegable. Además, si uno reconstruyera el sistema de ventilación pulmonar de terópodos no-avianos como uno similar al de los cocodrilos, la supuesta imposibilidad de que este sistema haya evolucionado en el mecanismo de ventilación de las aves aún debe ser demostrada.
Conclusiones
En las últimas décadas, una serie de significativos descubrimientos ha demostrado que características por siglos interpretadas como únicas de las aves, desde la fúrcula y la estructura del carpo hasta el comportamiento de empollamiento y la presencia de plumas, se originaron en grupos de terópodos maniraptores no-avianos. El enorme cuerpo de evidencia acumulado en estas décadas indica que, dentro del ámbito hipotético que caracteriza la inferencia filogenética, el origen de las aves a partir de estos dinosaurios es indiscutible. El examen de los fundamentos que sustentan las críticas elevadas por los detractores de esta hipótesis sugiere que éstas se encuentran basadas en observaciones erróneas o que su marco teórico carece del rigor requerido por la escuela moderna de inferencia filogenética. Las principales transformaciones que llevaron a la evolución de las aves modernas a partir de maniraptores no-avianos se encuentran vívidamente preservadas en el registro fósil.
Bibliografía Citada
1 Witmer LM (1991) Perspectives on avian origins. Pp. 427-466 en: Schultze H-P y Trueb L (eds) Origins of the higher groups of tetrapods: controversy and consensus. Cornell University Press, Ithaca [ Links ]
2 Witmer LM (2002) The debate on avian ancestry: phylogeny, function, and fossils. Pp. 3-30 en: Chiappe LM y Witmer LM (eds) Mesozoic birds: above the heads of dinosaurs. University of California Press, Berkeley [ Links ]
3 Padian K y Chiappe LM (1998) The origin and early evolution of birds. Biological Reviews 73:1-42. [ Links ]
4 Chiappe LM (2001) The rise of birds. Pp. 102-106 en: Briggs DEC y Crowther PR (eds.) Paleobiology II. Blackwell Science, Oxford [ Links ]
5 Gauthier JA y Gall LF (2001) New perspectives on the origin and early evolution of birds: proceedings of the international symposium in honor of John H. Ostrom. Peabody Museum of Natural History, Yale University, New Haven [ Links ]
6 Chiappe LM y Witmer LM (2002) Mesozoic birds: above the heads of dinosaurs. University of California Press, Berkeley [ Links ]
7 Chiappe LM y Dyke G (2002) The mesozoic radiation of birds. Annual Review of Ecology and Systematics 33:91-124 [ Links ]
8 Novas FE y Puerta P (1997) New evidence concerning avian origins from the Late Cretaceous of Patagonia. Nature 387:390-392 [ Links ]
9 Holtz TR Jr (1998) A new phylogeny of the carnivorous dinosaurs. Gaia 15:5-61 [ Links ]
10 Sereno PC (1999) The evolution of dinosaurs. Science 284:2137-2147 [ Links ]
11 Clark JM, Norell MA y Makovicky PJ (2002) Cladistic approaches to bird origins. Pp. 31-61 en: Chiappe LM y Witmer LM (eds) Mesozoic birds: above the heads of dinosaurs. University of California Press, Berkeley [ Links ]
12 Mikhailov KE (1997) Fossil and recent eggshell in amniotic vertebrates: fine structure, comparative morphology and classification. Special Papers in Palaeontology 56:1-80 [ Links ]
13 Carpenter K (1999) Eggs, nests, and baby dinosaurs. Indiana University Press, Bloomington [ Links ]
14 Norell MA, Clark JM, Dashzeveg D, Barsbold R y Chiappe LM (1994) A theropod dinosaur embryo, and the affinities of the flaming cliffs dinosaur eggs. Science 266:779-782 [ Links ]
15 Clark JM, Norell MA y Chiappe LM (1999) An oviraptorid skeleton from the Late Cretaceous of Ukhaa Tolgod, Mongolia, preserved in an avian-like brooding position over an oviraptorid nest. American Museum Novitates 3265:1-36 [ Links ]
16 Varricchio DJ, Horner JR y Jackson FD (2002) Embryos and eggs for the Cretaceous theropod Troodon formosus. Journal of Vertebrate Paleontology 22:564-576 [ Links ]
17 Grellet-Tinner G y Chiappe LM (en prensa) Dinosaur eggs and nesting: implications for understanding the origin of birds. En: Bakker RT y Currie PJ (eds) Symposium on the origin of birds. Indiana University Press, Bloomington [ Links ]
18 Clark JM, Norell MA y Barsbold R (2001) Two new oviraptorids (Theropoda: Oviraptorosauria), Upper Cretaceous Djadokhta Formation, Ukhaa Tolgod, Mongolia. Journal of Vertebrate Paleontology 21:209-213 [ Links ]
19 Osborn HF (1924) Three new Theropoda, Protoceratops zone, central Mongolia. American Museum Novitates 144:1-12 [ Links ]
20 Norell MA, Clark JM, Chiappe LM y Dashzeveg D (1995) A nesting dinosaur. Nature 378:774-776 [ Links ]
21 Varricchio DJ, Jackson F, Borkowski J y Horner JR (1997) Nest and egg clutches of the dinosaur Troodon formosus and the evolution of avian reproductive traits. Nature 385:247-250 [ Links ]
22 Zhou Z, Barrett PM y Hilton J (2003) An exceptional preserved Lower Cretaceous ecosystem. Nature 421:807-814 [ Links ]
23 Padian K, de Ricqlès AJ y Horner JR (2001) Dinosaurian growth rates and bird origins. Nature 412:405-408 [ Links ]
24 Chen PJ, Dong Z-M y Zhen S-N (1998) An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China. Nature 391:147-152 [ Links ]
25 Xu X, Wang X y Wu X (1999) A dromaeosaur dinosaur with filamentous integument from the Yixian Formation of China. Nature 401:262-266 [ Links ]
26 Ji Q, Currie P, Norell MA y Ji S-A (1998) Two feathered dinosaurs from northeastern China. Nature 393:753-761 [ Links ]
27 Zhou Z y Wang X (2000) A new species of Caudipteryx from the Yixian Formation of Liaoning, northeast China. Vertebrata PalAsiatica 38:111-127 [ Links ]
28 Xu X, Zhou Z y Wang X (2000). The smallest known non-avian theropod dinosaur. Nature 408:705-708 [ Links ]
29 Xu X, Zhou Z, Wang X, Kuang X, Zhang F y Du X (2003) Four-winged dinosaurs from China. Nature 421:335-340 [ Links ]
30 Norell MA, Clark JM y Makovicky PJ (2001) Phylogenetic relationships among coelurosaurian theropods. Pp. 49-68 en: Gauthier JA y Gall LF (eds) New perspectives on the origin and early evolution of birds: proceedings of the international symposium in honor of John H. Ostrom. Peabody Museum of Natural History, Yale University, New Haven [ Links ]
31 Feduccia A y Tordoff HB (1979) Feathers of Archaeopteryx: asymetric vanes indicate aerodynamic function. Science 203:1021-1022 [ Links ]
32 Feduccia A (1996) The origin and evolution of birds. Yale University Press, New Haven [ Links ]
33 Jensen JA y Padian K (1989) Small pterosaurs and dinosaurs from the Uncompahgre fauna (Brushy Basin Member, Morrison Formation: ?Tithonian), Late Jurassic, Western Colorado. Journal of Paleontology 63:364-373 [ Links ]
34 Xu X, Zhao Z y Clark JM (2001) A new therizinosaur from the Lower Jurassic lower Lufeng Formation of Yunnan, China. Journal of Vertebrate Paleontology 21:477-483 [ Links ]
35 Feduccia A y Nowicki J (2002) The hand of birds revealed by early ostrich embryos. Naturwissenschaften 89:391-393 [ Links ]
36 Larsson HC y Wagner GP (2002) Pentadactyl ground state of the avian wing. Journal of Experimental Zoology 294:146-151 [ Links ]
37 Burke AC y Feduccia A (1997) Developmental patterns and the identification of homologies in the avian hand. Science 278:666-668 [ Links ]
38 Feduccia A (1999) The origin and evolution of birds. Second edition. Yale University Press, New Haven [ Links ]
39 Ostrom JH (1976) Archaeopteryx and the origin of birds. Biological Journal of the Linnean Society 8:91-182 [ Links ]
40 Wagner GP y Gauthier JA (1999) 1, 2, 3 = 2, 3, 4: a solution to the problem of homology of the digits of the avian hand. Proceedings of the National Academy of Sciences of the United States of America 96:5111-5116 [ Links ]
41 Shapiro MD (2002) Developmental morphology of limb reduction in Hemiergis (Squamata: Scincidae): chondrogenesis, osteogenesis, and heterochrony. Journal of Morphology 254:211-231 [ Links ]
42 Dahn RD y Fallon JF (2000) Interdigital regulation of digit identity and homeotic transformation by modulated BMP signaling. Science 289:438-441 [ Links ]
43 Galis F, Kundrát M y Sinervo B (2003) An old controversy solved: bird embryos have five fingers. Trends in Ecology and Evolution 18:7-9 [ Links ]
44 Wagner GP y Misof Y (1993) How can a character be developmentally constrained despite variation in developmental pathways? Journal of Evolutionary Biology 6:449-455 [ Links ]
45 Mabee PM (2000) The usefulness of ontogeny in interpreting morphological characters. Pp. 84-114 en: Wiens JJ (ed) Phylogenetic analysis of morphological data. Smithsonian Institution Press, Washington DC [ Links ]
46 Ruben JA, Jones TD, Geist NR y Hillenius WJ (1997) Lung structure and ventilation in theropod dinosaurs and early birds. Science 278:1267-1270 [ Links ]
47 Currie PJ y Chen P-J (2001) Anatomy of Sinosauropteryx prima from Liaoning, northeastern China. Canadian Journal of Earth Science 38:1705-1727 [ Links ]

