










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEl hornero
Print version ISSN 0073-3407On-line version ISSN 1850-4884
Hornero vol.18 no.1 Buenos Aires Jan./Aug. 2003
Comportamiento de alimentación de la Gaviota Cocinera (Larus dominicanus) durante la temporada no reproductiva en playas arenosas de Península Valdés, Argentina
Marcelo Bertellotti 1,2, Gustavo Pagnoni 2 y Pablo Yorio 1,3,4
1 Centro Nacional Patagónico, CONICET. Brown 3500, U9120ACV Puerto Madryn, Chubut, Argentina.
2 Facultad de Ciencias Naturales, Universidad Nacional de la Patagonia San Juan Bosco. Brown 3700, U9120ACX Puerto Madryn, Chubut, Argentina.
3 Wildlife Conservation Society. 2300 Southern Boulevard, Bronx, New York, NY 10460, EEUU.
4 yorio@cenpat.edu.ar
Resumen
Las aves que se alimentan en la zona intermareal explotan recursos que frecuentemente están distribuidos en parches disponibles durante intervalos regulares que dependen del ciclo de la marea. La calidad de los parches de alimento depende del tipo de presas que los componen y de la densidad, distribución de tallas y disponibilidad de las mismas para los depredadores. En este trabajo analizamos el comportamiento de alimentación de la Gaviota Cocinera (Larus dominicanus) en sectores del intermareal con diferentes densidades y tamaños de presas en una playa arenosa de Península Valdés, Chubut, Argentina. Las gaviotas se alimentaron a lo largo de la línea de la marea desenterrando presas con movimientos acompasados de sus patas y su dieta estuvo principalmente compuesta por las almejas Tellina petitiana y Darina solenoides. Los tamaños promedio de las almejas de ambas especies identificadas en las egagrópilas fueron mayores que aquellos encontrados en sedimentos de la playa. La actividad de alimentación se concentró en áreas con alta densidad de almejas y donde se encontraron las mayores tallas de ambas almejas. Aunque la Gaviota Cocinera es una especie generalista que usa un amplio espectro de alimentos, este trabajo muestra que también puede ser selectiva en su elección de presas.
Palabras clave: alimentación, Argentina, dieta, Gaviota Cocinera, Larus dominicanus, temporada no reproductiva.
Abstract
Feeding behavior of the Kelp Gull (Larus dominicanus) during the non-breeding season at sandy beaches of Península Valdés, Argentina
Birds that feed in intertidal areas exploit resources that often are distributed in patches, and which become available during regular intervals that depend on the tidal cycle. The quality of foraging patches depends on prey species composition, prey density, prey size distribution, and prey availability to predators. We analyzed the feeding behaviour of Kelp Gulls (Larus dominicanus) on intertidal sectors with different densities and sizes of prey on a sandy beach at Península Valdés, Chubut Province, Argentina. Gulls fed along the tide line by foot paddling, and their diet was mostly composed of sand burrowing clams, Tellina petitiana and Darina solenoides. Average sizes of both clams identified in Kelp Gull pellets were larger than those found in beach sediments. Feeding activity was concentrated in areas of high clam density and with larger sizes of both clam species. Even though Kelp Gulls are foraging generalists that use a wide spectrum of food items, these results show that they can also be selective in their choice of prey.
Key words: Argentina, diet, foraging, Kelp Gull, Larus dominicanus, non-breeding season.
Recibido 15 mayo 2003, aceptado 11 agosto 2003
Las aves marinas generalmente utilizan para su alimentación recursos distribuidos en parches, temporal y espacialmente impredecibles (Croxall 1987). En contraste, en la zona intermareal los recursos son relativamente predecibles ya que se encuentran disponibles en intervalos regulares dependiendo del ciclo de marea. En estos intermareales, la calidad de los parches de alimento está generalmente determinada por el tipo de presas que los componen y por la densidad, distribución de tallas y disponibilidad de las mismas para los depredadores (Leopold et al. 1989).
Las gaviotas son un importante componente de la comunidad de aves que se alimenta en los intermareales (Moreira 1995, Burger y Gochfeld 1996). En la costa patagónica, la Gaviota Cocinera (Larus dominicanus) es una de las especies de aves marinas más abundante y de más amplia distribución (Yorio et al. 1998). Esta especie se alimenta de una amplia variedad de presas, principalmente invertebrados marinos y peces (Bertellotti y Yorio 1999, Yorio y Bertellotti 2002). En Puerto San José (Península Valdés, Chubut, Argentina), la Gaviota Cocinera se alimenta en extensos intermareales arenosos durante el otoño y el invierno (Pagnoni 1997). En esta área, la comunidad de invertebrados en los sedimentos blandos del intermareal está dominada tanto en densidad como en biomasa por las almejas Tellina petitiana y Darina solenoides (Mollusca, Bivalvia) (Pagnoni 1997). En este trabajo presentamos información sobre el comportamiento de alimentación de la Gaviota Cocinera en estas playas arenosas durante la estación no reproductiva.
Métodos
Puerto San José (42°25'S, 64°04'O), también conocido como Playa Fracasso, es una playa arenosa ubicada en el sudeste del golfo San José, Península Valdés, Chubut. La costa presenta una suave pendiente, quedando expuesta durante las mareas bajas una extensa superficie de aproximadamente 1.6 km2. La amplitud promedio de marea es de 5.6 m, alcanzando un máximo de 7 m. Para el análisis de la disponibilidad de presas, la playa fue dividida en seis niveles de altura de marea. Cada nivel fue marcado con una estaca de madera y numerado de acuerdo con la profundidad del agua durante la marea alta en intervalos de 1 m. Por ejemplo, cuando la línea de marea alcanzó la estaca del nivel 5, la profundidad del agua en el nivel 0 fue de 5 metros.
Durante mayo de 1996 se analizó la distribución y abundancia de almejas sobre una transecta perpendicular a la línea de marea. En cada nivel de marea se tomaron tres muestras en los primeros 15 cm del sedimento, utilizando un cilindro de 20 cm de diámetro. Los sedimentos fueron tamizados en zaranda de 1.5 mm de malla. En cada muestra se identificaron y contaron todas las almejas contenidas en el cilindro. Las valvas se midieron con calibre digital con una precisión de 0.1 mm para determinar la disponibilidad de las diferentes tallas de almeja presentes en cada nivel del intermareal.
Entre el 7 y el 10 de junio de 1996, de 08:00 a 20:00 h, se realizaron observaciones del comportamiento de alimentación de las Gaviotas Cocineras, utilizando un telescopio con zoom 15-40×. Se realizaron muestreos de barrido (Altman 1974) para determinar la actividad de las gaviotas a lo largo de la playa, registrándose si cada individuo se estaba alimentando o descansando. Los muestreos de barrido se efectuaron sobre grupos de un tamaño promedio de 44.6 gaviotas (DE = 8.9, n = 639 grupos). Se completó un total de 48 h de observación, distribuidas equitativamente en cada nivel de marea. Así, se determinó para cada nivel de marea el porcentaje del número de gaviotas en actividad de alimentación y de descanso. Los muestreos en cada nivel de marea fueron realizados tanto durante el ascenso como durante el descenso de la marea.
Complementariamente al registro del porcentaje de gaviotas con comportamiento de alimentación, y al final del período de observaciones, se recolectaron egagrópilas ("pellets"). Estas egagrópilas fueron recolectadas en una zona que las gaviotas utilizaban como área de descanso durante las mareas altas. La Gaviota Cocinera fue la única especie presente allí. Durante la primera visita se eliminaron todas las egagrópilas encontradas en esa área con el fin de descartar aquellas producidas en fechas anteriores a los muestreos. Las egagrópilas fueron analizadas bajo lupa binocular (aumento 5-20×). Se identificaron los restos de alimento hasta el menor nivel taxonómico posible, utilizando fragmentos de caparazón y quelas de crustáceos, fragmentos de valvas de moluscos, mandíbulas y quetas de poliquetos, otolitos y huesos de peces. Para la identificación de las presas se usaron claves publicadas (Castellanos 1967, Boschi et al. 1992, Gosztonyi y Kuba 1996) y material de referencia recolectado en la zona de estudio. En cada muestreo se estimó la frecuencia de ocurrencia de cada ítem (Ashmole y Ashmole 1967), expresada como el porcentaje de presencia de cada presa sobre el total de egagrópilas. Además se estimó el tamaño de las almejas consumidas por la Gaviota Cocinera usando la técnica descripta por Dekinga y Piersma (1993) y adaptada por Pagnoni (1997) para esta localidad. De este modo, se desarrollaron modelos de regresión (Fig. 1) entre la talla total y el tamaño de la charnela (condróforo) para las dos principales especies de almejas recolectadas en la playa de arena (Tellina petitiana y Darina solenoides).

Figura 1. Relación entre la longitud de la charnela y la longitud total de Tellina petitiana y Darina solenoides, utilizadas para predecir el tamaño de presa de la Gaviota Cocinera en Puerto San José, Chubut, a partir de los restos en las egagrópilas. Para Tellina petitiana: y = -0.09 + 28.54x; R2 = 0.95, P < 0.0001, n = 72. Para Darina solenoides: y = -1.90 + 12.21x; R2 = 0.93, P < 0.0001, n = 49.
Resultados
La almeja Tellina petitiana se distribuyó principalmente en el nivel 1 de altura de marea y, en menor proporción, en el nivel 2, estando ausente en el resto de los niveles en los que se subdividió la playa (Tabla 1). La talla promedio disponible fue de 23.2 ± 4.9 mm (rango: 10.7–36.0 mm, n = 83). La almeja Darina solenoides estuvo presente entre los niveles 3 y 6, con las mayores densidades en los niveles 5 y 6 (Tabla 1). El tamaño promedio de esta almeja fue de 12.1 ± 6.6 mm (rango: 1.9–30.0 mm, n = 193).
Tabla 1. Abundancia (individuos/m2) de Tellina petitiana y Darina solenoides de diferente tamaño en cada nivel de marea en playas arenosas de Puerto San José, Chubut, en junio de 1996.

La Gaviota Cocinera se alimentó en la zona tixotrópica (donde una delgada capa de agua cubre la arena) mediante chapoteo ("foot paddling"), haciendo un hoyo de unos 30 cm de diámetro con movimientos acompasados de sus patas y desenterrando así sus presas. El porcentaje de gaviotas en comportamiento de alimentación difirió significativamente entre niveles (H5 = 462.6, P < 0.0001, n = 639; Prueba de Kruskal-Wallis). La actividad de alimentación fue mayor en los niveles 1 y 5, mientras que la mayoría de las aves descansaron en los niveles 3 y 6 (Fig. 2). La actividad de alimentación no difirió entre los niveles 1 y 5 (Z = -2.5, no significativo; Prueba de Mann-Whitney), 2 y 4 (Z = -0.5, no significativo; Prueba de Mann-Whitney), ni 3 y 6 (Z = -1.2, no significativo; Prueba de Mann-Whitney), considerando 0.003 como valor crítico de la prueba (usando la corrección de Bonferroni) (Fig. 2).
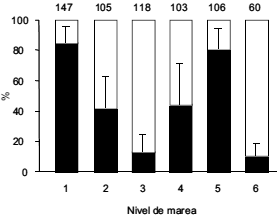
Figura 2. Porcentaje promedio (+ DE) de individuos de Gaviota Cocinera observados en actividad de alimentación (barras negras) y descanso (barras blancas) en cada nivel de marea en playas arenosas de Puerto San José, Chubut. El tamaño de muestra se indica sobre las barras.
La dieta de la Gaviota Cocinera estuvo compuesta por presas de 14 especies diferentes, sumadas a un número desconocido de especies de poliplacóforos, ofiuras, insectos y peces que no pudieron ser identificadas. Los moluscos fueron las presas dominantes. Las especies con mayor frecuencia de ocurrencia fueron las almejas Darina solenoides y Tellina petitiana (Tabla 2). La talla promedio de las almejas identificadas en las egagrópilas fue de 30.4 ± 4.9 mm (rango: 19.9–47.0 mm, n = 116) y 21.5 ± 5.9 mm (rango: 8.6–38.5 mm, n = 679) para Tellina petitiana y Darina solenoides, respectivamente. La talla de las almejas consumidas fue significativamente mayor a la de aquellas disponibles en la playa (t197 = -10.15, P < 0.001, para Tellina petitiana; t870 = -18.9, P < 0.001, para Darina solenoides; Prueba de t).
Tabla 2. Frecuencia de ocurrencia de presas presentes en la dieta de la Gaviota Cocinera en Puerto San José, Chubut, en junio de 1996 (n = 112 egagrópilas).

Discusión
La dieta de la Gaviota Cocinera en Puerto San José estuvo compuesta principalmente por moluscos bivalvos. Estudios recientes han demostrado que los invertebrados, particularmente los moluscos, constituyen una importante proporción de la dieta de la Gaviota Cocinera durante la temporada reproductiva en la costa patagónica (Bertellotti y Yorio 1999, Yorio y Bertellotti 2002). Esta especie también se alimenta predominantemente de moluscos en Sudáfrica (Brooke y Cooper 1979, Steele 1992), en las islas Shetland del Sur (Favero et al. 1997), en la Península Antártica (Fraser 1989), en la isla Marion (Blankley 1981) y en Chile (Bahamondes y Castilla 1986). Sin embargo, la información acerca de su dieta y comportamiento de alimentación durante la etapa invernal es aún muy escasa, aunque también muestra que los bivalvos son incorporados en su dieta en algunos sitios de la provincia de Buenos Aires (Silva et al. 2000). En el área de estudio, la Gaviota Cocinera se alimentó principalmente de Tellina petitiana y Darina solenoides, muy probablemente porque ambas especies son los invertebrados más abundantes del intermareal en ese sector de costa, constituyendo más del 75% de la biomasa infaunal (Pagnoni 1997). Las gaviotas que se alimentaron y descansaron en el área de estudio también se alimentaron en otros sectores de costa, tal como lo muestra la presencia en las egagrópilas de presas características de otros hábitats, como por ejemplo de costas rocosas (e.g., mejillones).
A pesar de que las almejas se encontraban distribuidas en toda la playa, las gaviotas se alimentaron principalmente donde una fina capa de agua cubrió la arena durante el ascenso y descenso de la marea. Esto es debido seguramente a que las almejas permanecen cercanas a la superficie cuando están cubiertas por una capa de agua (Pagnoni 1997). En todos los casos, la Gaviota Cocinera utilizó el chapoteo ("foot paddling") para capturar las presas, un método de alimentación también registrado en Sudáfrica para esta misma especie (Steele 1992).
El tiempo utilizado por las gaviotas para alimentación y descanso difirió entre los niveles de altura de marea. Las gaviotas maximizaron su actividad de alimentación en áreas con mayor densidad de almejas (e.g., niveles 1 y 5). La selección de parches con altas densidades de presas ha sido reportada por Leopold et al. (1989), quien sugirió que esta selección podría ser ventajosa debido a una reducción en el tiempo de búsqueda. Sin embargo, observamos una excepción en el nivel 6, en el cual se registró una baja proporción de alimentación a pesar de las altas densidades de almejas. Esta inconsistencia podría deberse a la baja disponibilidad relativa de almejas de tallas grandes en el nivel 6 con relación al nivel 5. Por ejemplo, la densidad promedio de individuos de Darina solenoides de tallas mayores a los 20 mm fue más alta en el nivel 5 que en el 6 (137.9 vs. 21.2 ind/m2). Se requerirá de nuevos estudios y de experimentación para comprender la importancia relativa de las densidades y del tamaño de presas sobre la selección de parches de alimentación en la Gaviota Cocinera.
El tamaño promedio de las almejas identificadas en las egagrópilas de Gaviota Cocinera fue mayor al de las almejas encontradas en los sedimentos de la playa. Además, los niveles en los cuales las gaviotas se alimentaron más intensamente (niveles 1 y 5) presentaron una mayor densidad de las almejas de mayor tamaño. Esto sugiere una selección de presas de tallas más grandes por parte de la Gaviota Cocinera. Sin embargo, dado que los fragmentos de presas pequeñas podrían no ser detectados en la misma proporción que los de presas más grandes en las egagrópilas, y que las almejas de tallas mayores podrían por lo tanto estar siendo sobrestimadas en las egagrópilas, no es posible evaluar apropiadamente la selección de presas usando únicamente esta metodología. Otros estudios han mostrado también que la Gaviota Cocinera selecciona presas más grandes en el intermareal. Por ejemplo, Ward (1991) observó que durante la temporada no reproductiva las Gaviotas Cocineras seleccionaron solo individuos grandes de la almeja Donax serra, a pesar de la disponibilidad de diferentes clases de talla de esta almeja en el área de estudio. En forma similar, la Gaviota Cocinera seleccionó los mayores tamaños de lapas (Nacella concinna) en la Antártida (Fraser 1989, Silva et al. 1999).
Aunque la Gaviota Cocinera es una especie generalista que se alimenta de una amplia variedad de presas (Bertellotti y Yorio 1999, Yorio y Bertellotti 2002), este trabajo muestra que también puede ser selectiva en sus decisiones de alimentarse en determinados sitios y sobre determinados tamaños de presa, probablemente optimizando de esta forma su actividad de alimentación, como ha sido mostrado en otros estudios (e.g., Ward 1991).
Agradecimientos. Agradecemos a Luis Bala y Julieta Rajlevsky por su ayuda durante la toma de datos y al Organismo Provincial de Turismo y Dirección de Fauna y Flora de Chubut, Argentina, por los permisos para trabajar en la Reserva Península Valdés. Agradecemos a D. Oro, G. Blanco y C. Alonso-Álvarez por sus comentarios sobre las primeras versiones del manuscrito. Durante la realización de este estudio, MB tuvo una beca del Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
Bibliografía Citada
Altman J (1974) Observational study of behavior: sampling methods. Behaviour 47:227-217 [ Links ]
Ashmole NP y Ashmole MJ (1967) Comparative feeding ecology of seabirds of a tropical oceanic island. Peabody Museum of Natural History Yale University Bulletin 24:1-131 [ Links ]
Bahamondes I y Castilla JC (1986) Predation of marine invertebrates by the kelp gull Larus dominicanus in an undisturbed intertidal rocky shore of central Chile. Revista Chilena de Historia Natural 59:65-72 [ Links ]
Bertellotti M y Yorio P (1999) Spatial and temporal patterns in the diet of Kelp Gull in Patagonia. Condor 101:790-798 [ Links ]
Blankley WO (1981) Marine food of Kelp Gulls, Lesser Sheathbills and Imperial Cormorants at Marion Island (Subantarctic). Cormorant 9:77-84 [ Links ]
Boschi E, Fischbach C e Iorio M (1992) Catálogo ilustrado de los crustáceos estomatópodos y decápodos marinos de Argentina. Volumen 10. Comisión Técnica Mixta del Frente Marítimo, Montevideo [ Links ]
Brooke RK y Cooper J (1979) What is the feeding niche of the Kelp Gull in South Africa? Cormorant 7:27-29 [ Links ]
Burger J y Gochfeld M (1996) Family Laridae (Gulls). Pp. 572-623 en: del Hoyo J, Elliott A y Sargatal J (eds) Handbook of the birds of the world. Volume 3. Hoatzin to auks. Lynx Edicions, Barcelona [ Links ]
Castellanos Z (1967) Catálogo de los moluscos bonaerenses. Anales de la Comisión de Investigaciones Científicas 8:1-365 [ Links ]
Croxall JP (1987) Seabirds: feeding ecology and role in marine ecosystems. Cambridge University Press, Cambridge [ Links ]
Dekinga A y Piersma T (1993) Reconstructing diet composition on the basis of faeces in a mollusc-eating wader, the Knot Calidris canutus. Bird Study 40:144-156 [ Links ]
Favero M, Silva P y Ferreyra G (1997) Trophic relationships between the Kelp Gull and the Antarctic Limpet at King George Island (South Shetland Islands, Antarctica) during the breeding season. Polar Biology 17:431-436 [ Links ]
Fraser WR (1989) Aspects of the ecology of Kelp Gull (Larus dominicanus) on Anvers Island, Antarctic Peninsula. PhD thesis, University of Minneapolis, Minneapolis [ Links ]
Gosztonyi AE y Kuba L (1996) Atlas de los huesos craneales y de la cintura escapular de peces costeros patagónicos. Informes Técnicos del Plan de Manejo Integrado de la Zona Costera Patagónica - Fundación Patagonia Natural 4:1-29 [ Links ]
Leopold MF, Swennen C y Bruijn LLM (1989) Experiments of selection of feeding site and food size in Oystercatchers, Haematopus ostralegus, of different social status. Netherlands Journal of Sea Research 23:333-346 [ Links ]
Moreira F (1995) Diet of Black-headed Gulls Larus ridibundus on emerged intertidal areas in the Tagus estuary (Portugal): predation or grazing? Journal of Avian Biology 26:277-282 [ Links ]
Pagnoni G (1997) Poblamiento de la infauna en la zona intermareal del golfo San José (Provincia del Chubut) y su importancia en la alimentación de aves migradoras. Tesis Doctoral, Universidad Nacional de La Plata, La Plata [ Links ]
Silva MP, Bastida R y Darrieu C (2000) Dieta de la Gaviota Cocinera (Larus dominicanus) en zonas costeras de la provincia de Buenos Aires, Argentina. Ornitología Neotropical 11:331-339 [ Links ]
Silva MP, Favero M y Martínez M (1999) Prey size selectivity by Kelp gulls on Antarctic limpets at King George Island, Antarctica. Polar Biology 21:397-400 [ Links ]
Steele WK (1992) Diet of Hartlaub's Gull Larus hartlaubii and the Kelp Gull L. dominicanus in the southwestern Cape Province, South Africa. Ostrich 63:68-82 [ Links ]
Ward D (1991) The size selection of clams by African Black Oystercatchers and Kelp Gulls. Ecology 72:513-522 [ Links ]
Yorio P y Bertellotti M (2002) Espectro trófico de la Gaviota Cocinera (Larus dominicanus) en tres áreas protegidas de Chubut, Argentina. Hornero 17:91-95 [ Links ]
Yorio P, Bertellotti M, Gandini P y Frere E (1998) Kelp Gulls (Larus dominicanus) breeding on the Argentine coast: population status and relationship with coastal management and conservation. Marine Ornithology 26:11-18 [ Links ]

