










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEl hornero
versión impresa ISSN 0073-3407versión On-line ISSN 1850-4884
Hornero v.18 n.1 Buenos Aires ene./ago. 2003
Diet of the Red-backed Hawk (Buteo polyosoma) in a forested area of the Chilean Patagonia and its relation to the abundance of rodent prey
Ricardo A. Figueroa Rojas 1,2, E. Soraya Corales Stappung 1 and Sergio Alvarado O. 1
1 Estudios para la Conservación y Manejo de la Vida Silvestre Consultores. Blanco Encalada 350, Chillán, Chile.
2 asio@tie.cl
Abstract
The diet of Red-backed Hawk (Buteo polyosoma) was quantified by analyzing 40 pellets collected during winter and spring 1999 and winter 2000 in a forested area of Aysén, Chilean Patagonia. Rodents were the most numerous prey (82% of all prey), and accounted for the highest contribution to total biomass (83%). Among rodents, Oligoryzomys longicaudatus was the most consumed prey species, accounting for 70% of total number and 64% of total biomass. Birds accounted for less than 6% in number, but their biomass contribution was significantly higher (more than 15%). Insects constituted 12% in number, but less than 0.5% of total biomass. The Red-backed Hawk seemed to consume Oligoryzomys longicaudatus according to their field abundance. Also, it seemed to prey disproportionately upon young adult rodents. Relating these results with previous studies on the same species, we conclude that this hawk mostly predates upon vertebrate species throughout its distribution range, with rodents being the most important prey.
Key words: Buteo polyosoma, Chilean Patagonia, diet, Oligoryzomys longicaudatus, prey selection, Red-backed Hawk.
Resumen
Dieta del Aguilucho Común (Buteo polyosoma) en un área boscosa de la Patagonia Chilena y su relación con la abundancia de roedores presa
La dieta del Aguilucho Común (Buteo polyosoma) fue cuantificada sobre la base de 40 regurgitados colectados durante el invierno y la primavera de 1999 y el invierno de 2000 en un área boscosa de Aysén, en la Patagonia chilena. Los roedores fueron las presas más numerosas (82% del total de presas) y constituyeron el mayor aporte de biomasa (83%). El roedor más consumido fue Oligoryzomys longicaudatus, que alcanzó el 70% del número total de presas y el 64% en biomasa. La incidencia numérica de las aves fue baja (menor a 6%), pero la contribución en biomasa fue mayor (más del 15%). Los insectos constituyeron el 12% del total de presas, pero menos del 0.5% en biomasa. El Aguilucho Común pareció consumir a Oligoryzomys longicaudatus de acuerdo a su abundancia en el ambiente. El Aguilucho Común también pareció depredar de manera desproporcionada sobre roedores adultos jóvenes. Una revisión de estudios anteriores indica que las especies de vertebrados son el núcleo de la dieta de este aguilucho a lo largo de su rango de distribución, siendo los roedores las presas más relevantes.
Palabras clave: Aguilucho Común, Buteo polyosoma, dieta, Oligoryzomys longicaudatus, Patagonia chilena, selección de presas.
Received 13 Mayo 2003, accepted 29 August 2003
The Red-backed Hawk (Buteo polyosoma) is a South American hawk distributed from the central Andes in Colombia to the Cape Horn Archipelago (Vaurie 1962, Brown and Amadon 1968, Hilty and Brown 1986) and Staten Island in Chile (Couve and Vidal-Ojeda 2000). This species inhabits arid lowlands, arid and humid montane shrub, tropical deciduous and southern temperate forests, shrub-grasslands, and farmlands in rugged or mountainous terrain (Brown and Amadon 1968, Jiménez 1995).
Jiménez (1995) has compiled the natural history of the Red-backed Hawk and, more recently, Jaksic et al. (2001, 2002) gave a species account for the Metropolitan and Magellan regions in Chile. Breeding biology, habitat use and selection, and diet of the Red-backed Hawk are all poorly known aspects. Literature indicates that the Red-backed Hawk preys on a wide spectrum of prey such as mammals, birds, reptiles, amphibians, and invertebrates (del Hoyo 1994, Jiménez 1995 and references therein). Island's population also consumes carrion (Brown and Amadon 1968, Woods 1975, Fuentes et al. 1993). Jiménez's (1995) revision, however, notes rodents as their main prey.
Quantitative information on the diet of the Red-backed Hawk has been documented separately for northern and central Chile (e.g., Schlatter et al. 1980, Jiménez 1995). Coincidentally, both studies found that rodents were the core of the diet. To our knowledge, no study has reported the food of this hawk in southern Chile. Here, we reported for the first time the diet of the Red-backed Hawk in a forested area of Patagonia, in southernmost Chile.
Methods
Observations on the diet of the Red-backed Hawk were taken on the north-western edge of General Carrera Lake, near Puerto Tranquilo village (46°37'S, 72°34'W) in Aysén, southernmost Chile. The Aysén region lies roughly between 44–49°S. The Andes mountains decline in height towards the south, and in this area the average elevation of passes is almost 1000 m. Because this region lies within the zone of the southern tradewinds, winds are predominantly from the West, depositing heavy precipitation on the western slope of the Andes and producing a rainshadow effect to the East. Precipitation declines from 4300 mm at Puerto Cisnes to roughly 240 mm at Chile Chico, an east-west distance of roughly 100 km (Cruces et al. 1999). A marked vegetation gradient parallels the climate trend, with Valdivian temperate rainforest on the western slope being replaced on the eastern slope by deciduous forests dominated by southern beeches (Nothofagus spp.), park-like woods, shrubs and finally the steppe and bunch-grass that is characteristic of Patagonia (Gajardo 1994). The entire Aysén region has been impacted heavily by human activities. Widespread clearing of forests with fire in the 1930s and 1940s led to the creation of extensive areas of pasture (Grosse 1974).
The landscape around Puerto Tranquilo is rugged and composed of fairly high hills (>500 m in elevation) and extensive flat areas in the valley. The highest hilltops and slopes (1000–1500 m in elevation) are covered by old deciduous Nothofagus forest dominated by lenga (Nothofagus pumilio). The lower slopes and the valleys are composed of wide pastures with scattered old trees, small meadows, and small patches and peninsulas of evergreen forest in varying degrees of degradation. The evergreen forests are constituted mainly of coihue (Nothofagus dombeyi), arrayán (Luma apiculata), arrayán macho (Rhaphithamnus spinosus), luma (Amomyrtus luma), michay (Berberis darwini), chilco (Fuchsia magellanica), calafate (Berberis buxifolia), maqui (Aristotelia chilensis) and aromo (Azara spp.). The main human activities are extensive livestock raising (e.g., cows, sheeps), firewood extraction, and agricultural crops (e.g., potatoes) in small areas.
From July 1999 to July 2000, we searched for pellets under tall trees located in a small evergreen forest remnant (approximately 3 ha) where at least a pair of Red-backed Hawks was observed perching and hunting. Fresh pellets were successfully collected during winter and spring 1999, and winter 2000. Although we made efforts to collect more pellets in the surrounding areas where hawks were sighted, none were found. Due to the abundant rainfall and the movement of domestic cattle inside the forest patch, most of the pellets were broken. Entire pellets were measured for length and diameter to the nearest 0.1 mm using a caliper and weighed on a digital balance to the nearest 0.01 g. Pellets were dissected to separate all prey remains. Because of their deformation, some pellets were not measurable for length and width, but we considered that they were complete in order to estimate weight. Mammal prey were identified and quantified on the basis of skulls or dental pairs (whichever gave the highest count) following Pearson's (1995) key. Bird prey were identified on the basis of feathers using two complementary methods: microscopic analysis of feather structures such as nodes and barbules (Reyes 1992), and a comparison of feather coloration patterns with voucher specimens deposited in the Zoology Department of the Austral University of Chile in Valdivia. Presence of feathers of a given species in one pellet was deemed to represent only one individual. Insects were recognized and counted by head capsules or mandibles and elytra pairs, following Peña (1986). We identified prey items down to the finest possible taxonomic category. The biomass percentage of each prey species was calculated following the method of Marti (1987), as Bi = 100[(Spi Ni)/Σ(Spi Ni)], where Spi is the mass of the ith species, Ni is the number of individuals of ith species consumed, and Bi is the percentage of the total prey mass contributed by the ith species. Masses of small mammals were obtained from individuals captured in the same study site, and masses of birds were obtained from the literature (Morgado et al. 1987, Egli 1996). Masses of insects correspond to unpublished data of the authors. Masses of unidentified prey items were assumed to be similar to the mean of the most closely related identified taxon.
As part of a study to characterize the hantavirus rodent hosts in the area, we live-trapped rodents from winter of 1998 to spring of 2002. During 1999 and 2000, when pellets were collected, seasonal trappings were carried out in three habitat types: evergreen forest remnants, Nothofagus park-like wood, and shrub-pasture lands (Table 1). This permitted us to obtain the clearest data about prey abundance with relation to the existing habitats. Trapping sites were separated 2–7 km among them. A trapping grid (5×10, 8–12 m apart) was established for each site using medium Sherman live-traps. Traps were active for one night for each season (seasonal effort = 200 traps-night, total effort = 1200 traps-night). The relative abundance of rodents was calculated using the number percentage, as Ni = 100(nSpi/NSpi), where nSpi is the number of individuals of the ith species, NSpi is the sum of rodents of all species captured, and Ni is the number percentage contributed by the ith species. In the field, rodents were aged by their body mass (Santana 1981, Meserve et al. 1991, Henríquez 1993, González and Mardones 2000, Figueroa et al. 2001; Table 2). Various studies have found that body mass confidently discriminates the relative age of small rodents in southern Chile (e.g., Santana 1981, Henríquez 1993, González and Mardones 2000). For ages of rodents found in the pellets we follow Feito et al. (1981), classifying prey as juveniles, young adults, or old adults. The individuals with unworn down teeth were assumed to be juveniles, those with scarce or medium wearing of teeth were assumed to be young adults, and those with well-worn teeth were assumed to be old adults.
Table 1. Sites where trapping of rodents was carried out during 1999 and 2000 in Puerto Tranquilo, Aysén, Chilean Patagonia. Habitat type (shrub-pasture lands, evergreen forest remnants, and Nothofagus park-like wood) and trapping season are indicated. A plus sign indicates functional trapping, whereas a minus sign indicates non-functional trapping.
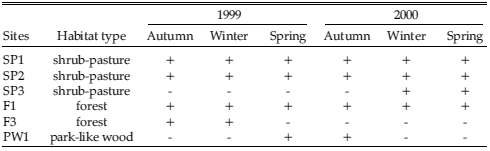
Table 2. Relative age classes estimated on the basis of body mass (g) for four rodent species captured in Puerto Tranquilo, Aysén, Chilean Patagonia.

For data analysis, we pooled results from all individual pellets due to the small seasonal sample size. To test statistically whether hawks took prey selectively or opportunistically, we compared the prey number in pellets with rodent abundance in the field using the Spearman's Rank Correlation Coefficient (rS) as recommended for coarse comparisons (Jaksic 1979, Fowler and Cohen 1986). This test is recommendable when small sample sizes are utilized, and when variables are not normally distributed (Zar 1984, Fowler and Cohen 1986). The pooling of samples could force inferences about prey selection. Thus, to examine possible temporal biases in the resulting rS values, we made comparisons on the basis of different time periods. In addition, to detect potential biases influenced by habitat type, we correlated separately results in diet with rodent captures in shrub-pasture land, forest and park-like wood. Due to the different technique of estimation, we avoided to make statistical comparisons among proportions of rodent's ages obtained from diet and field. The scientific names follow Araya et al. (1995) for birds, Musser and Carleton (1993) for rodents, and Gajardo (1994) for vegetation.
Results
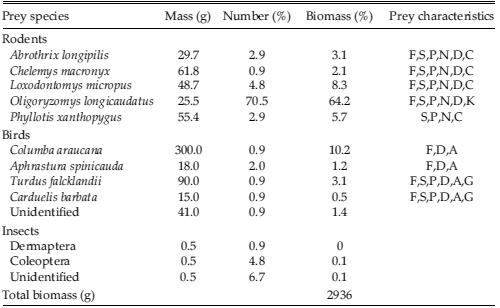
A total of 40 fresh pellets were collected, but only a few were complete. The complete pellets averaged 42.0 ± 0.4 mm length × 27.1 ± 0.1 mm width (n = 17; mean ± SE) and had a mean dry weight of 40.4 ± 0.4 g (n = 26). We found remains of 105 prey items in 12 prey categories (Table 3). Rodents were the most numerous prey and accounted for the highest contribution of biomass. The most consumed prey was Oligoryzomys longicaudatus (70% of all individual prey, 64% total biomass), followed by Loxodontomys micropus, Phyllotis xanthopygus, Abrothrix longipilis, and Chelemys macronyx. Birds were less numerous (<6%), but their biomass contribution was higher (>15%), mostly due to the contribution of Columba araucana (10% in biomass). Insects constituted 12% in number, but only <0.5% in biomass.
Table 3. Prey consumption by Red-backed Hawk (Buteo polyosoma) in Puerto Tranquilo, Aysén, Chilean Patagonia. Total number of pellets = 40. Prey characteristics are taken from Murúa (1996). F: forest, S: shrubland, P: prairie, N: nocturnal, D: diurnal, C: cursorial, K: scansorial, A: arboreal, G: ground.
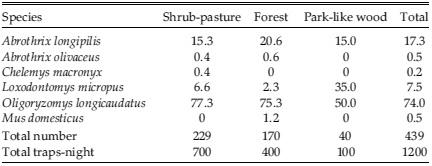
Throughout 1999-2000, we trapped 439 rodents of 6 species, being Oligoryzomys longicaudatus and Abrothrix longipilis the most captured (Table 4). Loxodontomys micropus accounted for only 7.5% in abundance, while the remaining species counted for less than 1% (Table 4). Phyllotis xanthopygus was not captured in the trapping. A similar trend was found for each season and habitat type, being Oligoryzomys longicaudatus and Abrothrix longipilis the most numerous (Tables 4 and 5). Loxodontomys micropus, however, constituted an important proportion of captures during autumn and spring, and in park-like wood (Tables 4 and 5).
Table 4. Percentage of rodents captured by season during 1999 and 2000 in Puerto Tranquilo, Aysén, Chilean Patagonia.

Table 5. Percentage of rodents captured in three habitat types during 1999 and 2000 in Puerto Tranquilo, Aysén, Chilean Patagonia.
Although positive correlation coefficients were found in all comparisons between consumption and field abundance of rodents, only in three cases there were significant correlations (Table 6). In all comparisons, Oligoryzomys longicaudatus seemed to be consistently eaten according to the field abundance (Table 7). Species that prevented a complete correlation were Abrothrix longipilis, Abrothrix olivaceus and Phyllotis xanthopygus (Table 7). Red-backed Hawk also seemed to consume disproportionately more young adult and less old adult individuals, either when considering all rodent prey species or Oligoryzomys longicaudatus alone (Fig. 1).
Table 6. Spearman's Rank Correlation (rS) for the comparison between number of rodents in Red-backed Hawk's diet and number of rodents in the field in Puerto Tranquilo, Aysén, Chilean Patagonia, evaluated for different time periods and habitat types. *: P < 0.05; ns: non significant; n = 7.

Table 7. Arithmetic difference between the Spearman's ranks of the rodent proportion in Red-backed Hawk's diet and the rodent number in the field in Puerto Tranquilo, Aysén, Chilean Patagonia, evaluated for different time periods and habitat types.
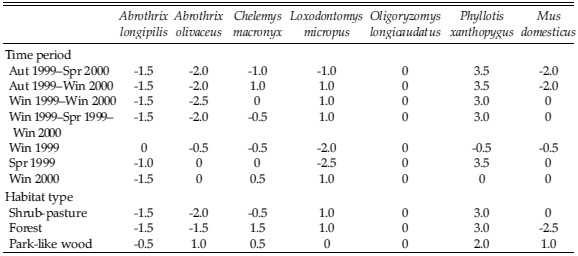

Figure 1. Relative abundance of individuals from each age class in Red-backed Hawk's diet (black bars) and in the field (white bars) in Puerto Tranquilo, Aysén, Chilean Patagonia. A = all rodent prey; B = Oligoryzomys longicaudatus.
Discussion
The average dimensions of pellets from Aysén were similar to those given by Schlatter et al. (1980) for central Chile (39.3 ± 1.4 mm length × 19.4 ± 0.6 mm width; mean ± SE). Pellets from Aysén, however, were evidently heavier than those from central Chile (2.6 ± 0.2 g). The highest weight of pellets from Aysén could be an artefact of our small sample size (40 vs 203 pellets for Aysén and central Chile, respectively) or a result of heavier remains composition of them.
Our results confirm Jiménez's findings (1995) about rodents as the main prey item of the Red-backed Hawk. Results obtained in Puerto Tranquilo suggest that the Red-backed Hawk would be a generalist predator with relation to rodent prey, but indicate also that some rodent prey were not taken according to its proportion in the field. When comparisons were made on the basis of time periods, most of the correlations were not statistically significant; they indicate also that Oligoryzomys longicaudatus seems to be consistently eaten according to field abundance. Significant positive correlations were only found during the winter 1999 and winter 2000, but in both cases the number of pellets was very small and results should be taken with caution. Studies carried out in other sites of Chile suggest also that the Red-backed Hawk is an opportunistic predator consuming the most abundant prey available. For example, degú (Octodon degu), the most abundant and stable rodent in xerophyllous shrublands of central Chile (Jaksic et al. 1981) was the most frequent prey of the Red-backed Hawk in La Dehesa (33°21'S, 70°32'W; Schlatter et al. 1980).
Red-backed Hawk search for their prey mainly in open terrain (Jiménez 1995, Jaksic et al. 2002). Accordingly, in our study area this hawk was observed frequently searching for prey on open sites such as shrub-pasture land or park-like wood. Although we do not know how much area distinct habitat types covered, visually shrub-pasture land and park-like wood appeared the most extensive around our study site. If Red-backed Hawks in Puerto Tranquilo mostly hunted in these habitat types, then they ate the rodents inhabiting there according to field abundance, as was suggested by our analysis (see Table 6). It is difficult, however, to interpret results since we know not how much the Red-backed Hawks utilized forest to hunt or search for prey. In addition, it is possible that rodent abundance in shrub-pasture land or park-like wood was influenced by dispersion from adjacent forests.
All species consumed by the Red-backed Hawk were habitat-generalist (Table 3), being its rank of abundance similar either in shrub-pasture land, forest and park-like wood. Thus, the macrohabitat use by rodent prey would not explain clearly preference or avoidance. Although Loxodontomys micropus was proportionally more abundant in park-like wood than in shrub-pasture land and forest, captures were carried out in only one site and could not reflect the same abundance on other park-like wood sites.
The high consumption of Oligoryzomys longicaudatus by the Red-backed Hawk in Puerto Tranquilo could be explained by the following factors. First, Oligoryzomys longicaudatus had the higher local abundance in the field year-round, in spite of strong seasonal oscillations (Figueroa et al. 2001). During winter, this rodent reaches its greatest level of abundance ("winter peak") as a result of the recruitment of new individuals to the population after the intensive summer-autumn breeding activity (Figueroa et al. 2001). This has also been previously documented for the temperate rainforest in Valdivia (Murúa and González 1986, Murúa et al. 1986, 1987). During spring and summer in Puerto Tranquilo, this species strongly declines in number, but still remains as the most abundant rodent (Figueroa et al. 2001). The abundance of this rodent species is strongly associated with the seasonal availability of food, particularly fruits and seeds (Murúa and González 1986, Meserve et al. 1988, González et al. 1989, Figueroa et al. 2001). During the autumn, an increased amount of fruits of arrayán, luma, chaura (Pernetya spp.) and mosqueta (Rosa moschata) are generated in Puerto Tranquilo, which permits survival and assures an abundant stock of rodent prey year-round. Second, Oligoryzomys longicaudatus is a "seed-seeker" (Murúa et al. 1982, Murúa et al. 1986) that moves long distances, which would increase the opportunities of encounter with predators.
Although most of the Red-backed Hawk's prey species in Aysén were diurnal (see Table 3), the high consumption of Oligoryzomys longicaudatus, a nocturnal rodent (Murúa et al. 1982), does not seem surprising because this species may extend its activity period into the day (Muñoz et al. 1990). In contrast, the lower consumption of Abrothrix longipilis (also a crepuscular-nocturnal species) would be explained by a differential antipredatory behaviour and microhabitat selection. This rodent makes only short quadruped runs toward the nearest shelter (Mann 1978) or exhibit "freezing" behaviour when presented overhead raptor silhouettes in laboratory experiments (Simonetti 1986). In Puerto Tranquilo, Abrothrix longipilis prefers to use sites with dense shrubs or high abundance of fallen trunks (Figueroa et al. 2001), which would reduce their vulnerability in presence of aerial predators.
The preferential consumption of young adult rodents by the Red-backed Hawk would be explained by at least three complementary reasons. First, young adults constituted an important fraction of the rodent population in the autumn and winter seasons in Puerto Tranquilo. Second, during the late autumn and early winter, young adult individuals are dispersing, which would make them more vulnerable to predation due to the greater chance of being detected. Third, juveniles and young adults could have less experience in avoiding predators. In Lago Atravesado, Aysén region, 100-km north Puerto Tranquilo, young adult rodents also constituted the core of the diet of American Kestrel (Falco sparverius; Figueroa and Corales 2002). Several previous studies have also found a disproportionate predation upon juveniles, and it has been suggested that this is because of higher dispersal rates, displacements to open patches by adults, lack of experience or sensory skills to avoid predators (Kotler 1985, Longland and Jenkins 1987, Kotler et al. 1988, Dickman et al. 1991, Castro and Jaksic 1995).
In the forested Patagonia in Aysén, the Oligoryzomys longicaudatus, Abrothrix longipilis and Abrothrix olivaceus triad dominate rodent communities (Kelt 1996, Figueroa et al. 2001). Occasionally, Loxodontomys micropus may be locally abundant too (Figueroa et al. 2001). Phyllotis xanthopygus, not captured in the trapping in Puerto Tranquilo, is an abundant species in the Patagonian steppe and shrub lands in Aysén (Tala et al. 1995, Kelt 1996). These habitat types are found as near as 50-70 km northeast and southeast of Puerto Tranquilo. Because hawks may move long distances, Red-backed Hawk probably take this prey species opportunistically along its displacements. It is curious that the European hare (Lepus europaeus) was not eaten in spite of its high local abundance (Cerda and Cruces 1998). Schlatter et al. (1980) argued that this hawk, being smaller, hunts the smallest rodents and is unable to kill and handle prey as large as lagomorphs (European hares's body mass range from 2.8–3.3 kg for adults in the Chilean Patagonia; Cerda and Cruces 1998).
We conclude that Red-backed Hawk in Puerto Tranquilo, (1) is essentially a rodent-eating predator, (2) consumes a fraction of their rodent prey according to field abundance, particularly Oligoryzomys longicaudatus, and (3) preys strongly upon young adult rodents. Further investigation is necessary to prove our assertions, since our sample size in Aysén was not large. Perhaps results reported here could be reverted if a greater number of pellets were collected and if other statistical tests were applied.
From an zoonotic perspective, the predatory impact of the Red-backed Hawk on Oligoryzomys longicaudatus could have a special importance locally because this rat has been recognized as the primary hantavirus rodent host in much of the Aysén region (Murúa 1998, Figueroa et al. 2001). In spite of the relevance for public health, role of raptors as regulators of rodents transmitting zoonotical diseases has been scarcely explored in southern South America (e.g., Bellocq 1987, 1990, Bellocq and Kravetz 1990).
Acknowledgements. This paper is part of the Monitoring Program of Hantavirus Reservoirs of Aysén, financed by the Regional Government of Aysén and the Livestock and Agricultural Service (Servicio Agrícola y Ganadero), in Aysén, and the Ministry of Agriculture of Chile. We thank Julio Cerda Cordero, chief of the Monitoring Program of Hantavirus Reservoirs, who provided support and advice, and Diane Haughney for helping us improve our English. Obtaining field data was made possible by the collaboration of Hernaldo Saldivia and Jorge Chavez. Part of the manuscript was written by the first author during a stay as invited professor in the Veterinary Medicine Faculty of the Concepción University in Chillán, by the initiative of Dr. Daniel González. Two anonymous reviewers made cogent suggestions that helped to improve this paper substantially.
Literature Cited
Araya B, Bernal M, Schlatter R and Sallaberry M (1995) Lista patrón de las aves chilenas. Editorial Universitaria, Santiago [ Links ]
Bellocq MI (1987) Selección de habitat de caza y depredación diferencial de Athene cunicularia sobre roedores en agroecosistemas agrarios. Revista Chilena de Historia Natural 60:81-86 [ Links ]
Bellocq MI (1990) Composición y variación temporal de la dieta de Tyto alba en ecosistemas agrarios pampeanos, argentina. Vida Silvestre Neotropical 2:32-35 [ Links ]
Bellocq MI and Kravetz FO (1990) Practical and theoretical implications of perch use for avian predator on rodent populations. Ecosur 16:61-67 [ Links ]
Brown LH and Amadon D (1968) Eagles, hawks and falcons of the world. Volume 1. Country Life Books, London [ Links ]
Castro SA and Jaksic FM (1995) Great Horned and Barn owls prey differentially according to the age/size of a rodent in northcentral Chile. Journal of Raptor Research 29:245-249 [ Links ]
Cerda J and Cruces P (1998) Investigación y estudio poblacional de la liebre en la XI región de Aysén. Gobierno Regional de Aysén-Servicio Agrícola y Ganadero, Ministerio de Agricultura, Chile [ Links ]
Couve E and Vidal-Ojeda C (2000) Birds of the Beagle Channel and Cape Horn. Fantástico Sur Birding, Punta Arenas [ Links ]
Cruces P, Cerda J and Ahumada M (1999) Guías de condición para los pastizales de la ecorregion templada húmeda de Aysén. Gobierno Regional de Aysén-Servicio Agrícola y Ganadero, Ministerio de Agricultura, Chile [ Links ]
Dickman CR, Prevadec M and Lynam AJ (1991) Differential predation of size and sex classes of mice by the barn owl, Tyto alba. Oikos 62:67-76 [ Links ]
Egli G (1996) Biomorfología de algunas aves de Chile central. Boletín Chileno de Ornitología 3:2-9 [ Links ]
Feito RS, González LA and Mondión R (1981) Determination of age and age classes of Akodon olivaceus in southern Chile. Studies on Neotropical Fauna and Environment 16:201-207 [ Links ]
Figueroa RA and Corales ES (2002) Winter diet of the American Kestrel (Falco sparverius) in the Forested Chilean Patagonia and its relation to the availability of prey. International Hawkwatcher 5:7-14 [ Links ]
Figueroa RA, Corales ES, Cerda J and Saldivia H (2001) Monitoreo de roedores silvestres en seis localidades rurales de la región de Aysén, Chile: Informe Final 1998-2000. Gobierno Regional de Aysén-Servicio Agrícola y Ganadero, Ministerio de Agricultura, Chile [ Links ]
Fowler J and Cohen L (1986) Statistics for ornithologists. British Trust for Ornithology, Norfolk [ Links ]
Fuentes MA, Simonetti JA, Sepúlveda MS and Acevedo PA (1993) Diet of the Red-backed Buzzard (Buteo polyosoma exsul) and the Short-eared Owl (Asio flammeus suinda) in the Juan Fernández Archipelago off Chile. Journal of Raptor Research 27:167-169 [ Links ]
Gajardo R (1994) La vegetación natural de Chile: clasificación y distribución geográfica. Editorial Universitaria, Santiago [ Links ]
González LA and Mardones M (2000) Determinación de edad en dos especies del género Abrothrix. Medio Ambiente 13:11-18 [ Links ]
González LA, Murúa RE and Jofré C (1989) The effect of seed availability on population density of Oryzomys in southern Chile. Journal of Mammalogy 70:401-403 [ Links ]
Grosse J (1974) Visión de Aysén. Second edition. Author's publication. Santiago [ Links ]
Henríquez C (1993) Estimación de edad a partir de caracteres dentarios, corporales y craneométricos en Oryzomys longicaudatus (Rodentia, Cricetidae). Biol. Chem. Sc. Teacher's Thesis, Universidad Austral de Chile, Valdivia [ Links ]
Hilty SL and Brown WL (1986) A guide to birds of Colombia. Princeton University Press, Princeton [ Links ]
del Hoyo J, Elliot A and Sargatal J (1994) Handbook of the birds of the world. Volume 2. New World vultures to guineafowl. Lynx Edicions, Barcelona [ Links ]
Jaksic FM (1979) Técnicas estadísticas simples para evaluar selectividad dietaria en Strigiformes. Medio Ambiente 4:114-118 [ Links ]
Jaksic FM, Iriarte JA and Jiménez JE (2002) The raptors of Torres del Paine National Park, Chile: biodiversity and conservation. Revista Chilena de Historia Natural 75:449-461 [ Links ]
Jaksic FM, Pavez EF, Jiménez JE and Torres-Mura JC (2001) The conservation status of raptors in the Metropolitan Region, Chile. Journal of Raptor Research 35:151-158 [ Links ]
Jaksic FM, Yañez JL and Fuentes ER (1981) Assessing a small mammal community in central Chile. Journal of Mammalogy 62:391-396 [ Links ]
Jiménez JE (1995) Historia natural del aguilucho Buteo polyosoma: una revisión. Hornero 14:1-9 [ Links ]
Kelt DA (1996) Ecology of small mammals across a strong environmental gradient in southern South America. Journal of Mammalogy 77:205-219 [ Links ]
Kotler BP (1985) Owl predation on desert rodents which differ in morphology and behavior. Journal of Mammalogy 66:824-828 [ Links ]
Kotler BP, Brown JS, Smith RJ and Wirtz WO (1988) The effects of morphology and body size on rates of owl predation on desert rodents. Oikos 53:145-152 [ Links ]
Longland WS and Jenkins SH (1987) Sex and age affect vulnerability of desert rodents to owl predation. Journal of Mammalogy 68:746-754 [ Links ]
Mann G (1978) Los pequeños mamíferos de Chile: marsupiales, quirópteros, edentados y roedores. Gayana, Zoología 40:1-342 [ Links ]
Marti C (1987) Raptor food habits studies. Pp. 67-79 in: Pendleton BA, Millsap BA, Cline KW and Bird DM (eds) Raptor management techniques manual. National Wildlife Federation, Washington DC [ Links ]
Meserve PL, Lang BK, Murúa RE, Muñoz-Pedreros A and González LA (1991) Characteristics of a terrestrial small mammal assemblage in a temperate rainforest in Chile. Revista Chilena de Historia Natural 64:157-169 [ Links ]
Meserve PL, Lang BK and Patterson BD (1988) Trophic relationships of small mammals in a Chilean temperate rainforest. Journal of Mammalogy 69:721-730 [ Links ]
Morgado E, Gunther B and González U (1987) On the allometry of wings. Revista Chilena de Historia Natural 60:71-79 [ Links ]
Muñoz A, Murúa RE and González LA (1990) Nicho ecológico de la asociación de micromamíferos en un agroecosistema forestal de Chile. Revista Chilena de Historia Natural 63:267-277 [ Links ]
Murúa RE (1996) Comunidades de mamíferos del bosque templado de Chile. Pp. 113-133 in: Armesto JJ, Villagrán C and Arroyo MTK (eds) Ecología de los bosques nativos de Chile. Editorial Universitaria, Santiago [ Links ]
Murúa RE (1998) Ecología de los reservorios silvestres de hantavirus en Chile. Revista Chilena de Infectología 15:79-83 [ Links ]
Murúa RE and González LA (1986) Regulation of numbers in two Neotropical rodent species in southern Chile. Revista Chilena de Historia Natural 59:193-200 [ Links ]
Murúa RE, González LA and Jofré C (1982) Estudios ecológicos de roedores silvestres en los bosques templados fríos de Chile. Boletín del Museo Nacional de Historia Natural 38:105-116 [ Links ]
Murúa RE, González LA and Meserve PL (1986) Population Ecology of Oryzomys longicaudatus Phillipi (Rodentia: Cricetidae) in southern Chile. Journal of Animal Ecology 55:281-293 [ Links ]
Murúa RE, Rojas M and González LA (1987) Ciclo reproductivo anual de Akodon olivaceus y Oryzomys longicaudatus (Rodentia; Cricetidae). Medio Ambiente 8:4-51 [ Links ]
Musser GM and Carleton MD (1993) Family Muridae. Pp. 501-756 in: Wilson DE and Reeder DM (eds) Mammal species of the world: a taxonomic and geographic reference. Smithsonian Institution, Washington DC [ Links ]
Pearson O (1995) Annotated keys for identifying small mammals living in or near Nahuel Huapi National Park or Lanin National Park, southern Argentina. Mastozoología Neotropical 2:99-148 [ Links ]
Peña L (1986) Introducción a los insectos de Chile. Editorial Universitaria, Santiago [ Links ]
Reyes C (1992) Clave para la identificación de los órdenes de aves chilenas: microestructura de los nodos de las bárbulas. Biol. Teacher's Thesis, Universidad de los Lagos, Osorno [ Links ]
Santana AM (1981) Determinación de edad y clases de edad en Oryzomys longicaudatus, roedor silvestre del bosque higrófilo templado en el sur de Chile. Biol. Chem. Sc. Teacher's Thesis, Universidad Austral de Chile, Valdivia [ Links ]
Schlatter RP, Yañez JL and Jaksic FM (1980) Food-niche relationships between Chilean Eagles and Red-backed Buzzards in Central Chile. Auk 97:897-898 [ Links ]
Simonetti J (1986) Microhabitat use by small mammals in central Chile. PhD Dissertation, University of Washington, Seattle [ Links ]
Tala C, González B and Bonacic C (1995) Análisis de la dieta del tucúquere Bubo virginianus en el valle del Río Ibañez, Aysén. Boletín Chileno de Ornitología 2:34-35 [ Links ]
Vaurie C (1962) A systematic study of the Red-backed hawks of South America. Condor 64:277-290 [ Links ]
Woods RW (1975) The birds of the Falkland Islands. Compton Press, Wiltshire [ Links ]
Zar JH (1984) Biostatistical analysis. Second edition. Prentice-Hall, Englewood Cliffs [ Links ]

