










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEl hornero
Print version ISSN 0073-3407On-line version ISSN 1850-4884
Hornero vol.18 no.2 Buenos Aires Sept./Dec. 2003
Impacto del parasitismo de cría del Tordo Renegrido (Molothrus bonariensis) sobre el éxito reproductivo de sus hospedadores
Juan C. Reboreda 1,2, Myriam E. Mermoz 1, Viviana Massoni 1, Andrea A. Astié 1 y Fabián L. Rabuffetti 1
1 Departamento de Ecología, Genética y Evolución, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Piso 4, Pabellón 2, Ciudad Universitaria, C1428EHA Buenos Aires, Argentina.
2 reboreda@bg.fcen.uba.ar
Resumen
Se presenta una revisión del impacto que produce el parasitismo de cría del Tordo Renegrido (Molothrus bonariensis) sobre el éxito reproductivo de sus hospedadores. Se analizó el efecto del parasitismo sobre la supervivencia de los nidos y sobre la supervivencia de huevos, el éxito de eclosión y la supervivencia de pichones en nidos exitosos. También se analizaron el efecto de la intensidad de parasitismo y del comportamiento de picadura de huevos en nidos no parasitados. El principal impacto del parasitismo sobre el éxito reproductivo de los hospedadores estudiados fue producido por la picadura de huevos. Este comportamiento también redujo el éxito reproductivo del hospedador en nidos no parasitados. El parasitismo redujo el éxito de eclosión y la supervivencia de los pichones solo en hospedadores mucho más pequeños que el parásito. En estos, el parasitismo también aumentó la frecuencia de abandono de nidos, generalmente como consecuencia de la destrucción de la mayoría de los huevos del hospedador. El costo producido por la picadura de huevos aumentó con la intensidad de parasitismo. Considerando que la intensidad y la frecuencia de parasitismo están estrechamente asociadas, esta última sería un buen indicador del impacto que el parasitismo de Molothrus bonariensis podría producir sobre el éxito reproductivo de sus hospedadores. Se mencionan algunos aspectos de la biología del parásito y de sus hospedadores que es necesario estudiar con mayor detalle para poder estimar el impacto que produce el parasitismo sobre la viabilidad de las poblaciones de los hospedadores.
Palabras clave: éxito reproductivo, Molothrus bonariensis, parasitismo de cría, picadura de huevos, Tordo Renegrido.
Abstract
Impact of brood parasitism by Shiny Cowbirds (Molothrus bonariensis) on the reproductive success of their hosts
We present a review of the impact of brood parasitism by Shiny Cowbirds (Molothrus bonariensis) on the reproductive success of their hosts. We analysed the effect of parasitism on nest survival and on egg survival, hatching success and chick survival in successful nests. We also analysed the effect of the intensity of parasitism and egg pecking behaviour in unparasitized nests. Egg pecking produced the main impact of parasitism on host's reproductive success. This behaviour also reduced host's reproductive success in unparasitized nests. Parasitism decreased hatching success and chick survival only in hosts much smaller than the parasite. In these hosts, parasitism also increased the frequency of nest abandonment, generally as a result of the destruction of the majority of host's eggs. The cost produced by egg pecking increased with intensity of parasitism. Considering that intensity and frequency of parasitism are tightly associated, the latter would be a good predictor of the impact that Shiny Cowbird's parasitism might produce on the reproductive success of their hosts. We mention some aspects of the biology of the parasite and their hosts that we need to study in more detail to estimate the impact that brood parasitism produces on the viability of hosts' populations.
Key words: brood parasitism, egg pecking, Molothrus bonariensis, reproductive success, Shiny Cowbird.
El parasitismo interespecífico de cría es una estrategia reproductiva en la que algunos individuos, los parásitos, depositan sus huevos en nidos de individuos de otras especies, los hospedadores, que proveen la totalidad del cuidado parental. En las aves, esta estrategia ha evolucionado en forma independiente en al menos siete oportunidades y en la actualidad está presente en aproximadamente 90 especies 1,2. Uno de los grupos en los cuales el parasitismo ha evolucionado en forma independiente es el de los tordos (familia Icteridae, géneros Molothrus y Scaphidura). Este grupo incluye a cinco especies que, de acuerdo a estudios filogenéticos hechos con secuencias de ADN mitocondrial, tendrían un ancestro común 3,4.
Los parásitos de cría interespecíficos dependen enteramente del hospedador para su reproducción y éste ve disminuido su éxito reproductivo como consecuencia del parasitismo (revisiones recientes del tema se encuentran en Ortega 5, Rothstein y Robinson 6 y Davies 2). En el caso de los tordos parásitos, éstos suelen disminuir el éxito reproductivo de sus hospedadores de distintas formas. Los tordos suelen picar o remover los huevos del hospedador cuando visitan sus nidos 7-10, sus huevos o pichones pueden reducir el éxito de eclosión de los huevos del hospedador 11,12, y sus pichones pueden reducir la tasa de crecimiento y la supervivencia de los pichones del hospedador 13-15, la supervivencia de las crías del hospedador luego de que éstas abandonan el nido 15 o el éxito reproductivo futuro de la pareja que los ha criado 6. Además, el parasitismo suele aumentar la probabilidad de que el hospedador abandone su nido 16,17 y, en algunos casos, la presencia de pichones del parásito puede aumentar la probabilidad de predación del nido 16.
El parasitismo de cría está estrechamente relacionado con la dinámica poblacional y la conservación de las poblaciones de especies hospedadoras 18-20. Un ejemplo de ello es Molothrus ater, especie que recientemente expandió su distribución en América del Norte favorecido por la fragmentación de los bosques y el desarrollo de las actividades agropecuarias 21. Esta expansión le permitió incorporar como hospedadores a especies o poblaciones que no tuvieron una historia de simpatría previa con él y que, por lo tanto, no desarrollaron defensas antiparasitarias efectivas. De esta forma, el parasitismo de Molothrus ater se transformó en una de las principales amenazas para las poblaciones de varias especies de aves canoras migrantes neotropicales 22.
Los parásitos de cría generalistas (i.e., aquellos que utilizan varias especies de hospedadores) pueden constituir una seria amenaza para sus hospedadores preferidos, ya que la población del parásito depende principalmente de las especies más abundantes y no es afectada por la reducción de la población del hospedador preferido. Por lo tanto, estos hospedadores pueden experimentar aumentos en la frecuencia de parasitismo aún cuando sus poblaciones estén disminuyendo 18,20. Por otra parte, el efecto del parasitismo de cría puede ser en algunos casos más importante que el de la predación de nidos, ya que muchos hospedadores no vuelven a nidificar luego de haber criado exitosamente pichones, aún tratándose de pichones parásitos, mientras que sí lo intentan si su nido fue predado 1.
El Tordo Renegrido (Molothrus bonariensis) es un parásito de cría extremadamente generalista en cuanto a la selección de hospedadores 23. Sus huevos han sido encontrados en nidos de 232 especies y se presume que al menos 68 de ellas crían exitosamente a sus pichones 5,24,25. Esta especie estuvo históricamente restringida a los pastizales y bosques abiertos de América del Sur, desde el río Colorado en la Patagonia hasta la isla de Trinidad en el Caribe 26. Durante los últimos 100 años, Molothrus bonariensis expandió su distribución hacia el sur del río Colorado en la Patagonia y hacia las Antillas en América Central 27, donde es responsable de la casi extinción de Agelaius xanthomus 28. En la actualidad, Molothrus bonariensis se distribuye desde la provincia de Chubut en Argentina hasta la costa atlántica del sur de Estados Unidos 5,29,30.
Si bien existen numerosos trabajos que reportan la presencia de parasitismo de Molothrus bonariensis en distintos hospedadores, pocos de estos estudios han analizado los costos que produce el parasitismo sobre los distintos componentes del éxito reproductivo del hospedador. Evaluar el costo del parasitismo sobre el éxito reproductivo del hospedador resulta de particular importancia en algunos casos (e.g., Xanthopsar flavus o Curaeus forbesi) ya que se trata de especies globalmente amenazadas y se presume que el parasitismo podría ser una de las causas de su declinación 31,32.
En este trabajo se presenta una revisión de los costos que produce el parasitismo sobre el éxito reproductivo de sus hospedadores. En particular, se analiza la relación entre la frecuencia y la intensidad de parasitismo y el efecto que tiene el parasitismo sobre la supervivencia de los nidos del hospedador y sobre la supervivencia de huevos, éxito de eclosión y supervivencia de pichones del hospedador en nidos exitosos (i.e., nidos que produjeron volantones). También se analiza el efecto de la intensidad de parasitismo sobre el éxito reproductivo y los costos producidos por la picadura de huevos en nidos no parasitados. Por último, se mencionan algunos aspectos de la biología del parásito y de sus hospedadores que aún es necesario estudiar para poder predecir el impacto que produce el parasitismo sobre la viabilidad de las poblaciones de los hospedadores.
Métodos
Frecuencia e intensidad de parasitismo
Para evaluar la relación entre la frecuencia (proporción de nidos parasitados) y la intensidad de parasitismo (número de huevos parásitos por nido parasitado) se utilizaron datos provenientes de 28 estudios realizados en 15 especies de hospedadores (Tabla 1). Para este análisis sólo se tuvieron en cuenta estudios que tenían datos de al menos 25 nidos y que fueron realizados en el área de distribución histórica de Molothrus bonariensis. Se calculó la frecuencia de nidos no parasitados esperada según la distribución de Poisson para distintos valores del número promedio de huevos por nido (no parasitados y parasitados). A partir de esos datos se calcularon los valores de intensidad y frecuencia esperada de parasitismo.
Tabla 1. Estudios utilizados para el análisis de la relación entre intensidad y frecuencia de parasitismo de Molothrus bonariensis. Para cada estudio se indica la frecuencia e intensidad de parasitismo, el número de nidos reportados, la localidad y la fuente bibliográfica utilizada. La subespecie de Molothrus bonariensis analizada fue siempre Molothrus bonariensis bonariensis excepto donde se indica.

Efecto del parasitismo sobre distintos componentes del éxito reproductivo del hospedador
Para estimar los costos producidos por el parasitismo se compararon los siguientes componentes del éxito reproductivo del hospedador entre nidos no parasitados y parasitados: (1) supervivencia de nidos (proporción de nidos que produjeron al menos un volantón del hospedador o del parásito) y (2) eficiencia reproductiva en nidos exitosos (proporción de los huevos puestos por el hospedador que produjeron volantones). Además, en los casos en que se dispuso de los datos necesarios, se estimó el costo del parasitismo sobre tres componentes de la eficiencia reproductiva: (1) supervivencia de huevos (proporción de los huevos puestos por el hospedador que estaban presentes en el nido al final del período de incubación), (2) éxito de eclosión (proporción de huevos puestos por el hospedador presentes en el nido al final del período de incubación que eclosionaron) y (3) supervivencia de pichones (proporción de pichones del hospedador nacidos que abandonaron exitosamente el nido).
Para evaluar si el parasitismo afectó la supervivencia de los nidos se compararon las proporciones de nidos no parasitados y parasitados exitosos mediante una Prueba de Independencia 45. En los casos en que se dispuso de información sobre las tasas de supervivencia diaria estimadas mediante el método de Mayfield 46, se realizó una comparación entre nidos no parasitados y parasitados con el programa CONTRAST 47. Por otra parte, para evaluar si el parasitismo afectó la eficiencia reproductiva en nidos exitosos se compararon las proporciones de volantones por huevo puesto en nidos no parasitados y parasitados mediante la Prueba de Independencia o, cuando los datos disponibles lo permitieron, la Prueba de Mann-Whitney 45. Finalmente, para evaluar si el parasitismo afectó la supervivencia de huevos, el éxito de eclosión y la supervivencia de pichones en nidos exitosos se compararon estas variables entre nidos no parasitados y parasitados mediante la Prueba de Mann-Whitney.
Efecto de la intensidad del parasitismo sobre la supervivencia de huevos del hospedador
Para evaluar el efecto de la intensidad de parasitismo sobre la supervivencia de huevos durante los estadíos de puesta e incubación se realizó un Análisis de Correlación de Spearman entre la proporción de huevos puestos por el hospedador que permanecían en el nido al final de la incubación y el número de huevos parásitos que recibió el nido. Este análisis se realizó en forma independiente para tres hospedadores: Pseudoleistes virescens, Mimus saturninus y Turdus amaurochalinus.
Costos producidos por la picadura de huevos en nidos no parasitados
Para evaluar los costos asociados al comportamiento de picadura de huevos del hospedador por parte de Molothrus bonariensis se comparó la supervivencia de huevos en cuatro grupos de nidos: (1) nidos no parasitados y sin indicios de picadura de huevos, (2) nidos no parasitados en los que hubo picadura de huevos, (3) nidos parasitados sin picadura de huevos, y (4) nidos parasitados con picadura de huevos. Este análisis se realizó en forma independiente para dos especies de hospedadores: Agelaius thilius y Turdus amaurochalinus. Las comparaciones entre grupos se realizaron mediante la Prueba de Kruskal-Wallis y comparaciones múltiples 45.
Uno de los problemas para analizar el efecto de la picadura de huevos en nidos no parasitados es poder determinar si efectivamente no existió parasitismo en esos nidos. En una parte de su distribución (este de Argentina, Uruguay y sudeste de Brasil), Molothrus bonariensis presenta un polimorfismo en la coloración de sus huevos, que pueden ser blanco-inmaculados o manchados. Algunos hospedadores, como Agelaius thilius, aceptan huevos de ambos morfos 57, mientras que otros, como Mimus saturninus, Turdus rufiventris o Pseudoleistes virescens, aceptan los huevos manchados pero rechazan los del morfo blanco 36,38,48,49. Para estos últimos hospedadores no es posible distinguir entre nidos no parasitados y nidos parasitados con huevos del morfo blanco que fueron rechazados por el hospedador antes de que el investigador visitara el nido. La elección de Agelaius thilius y Turdus amaurochalinus para estimar los costos asociados a picaduras en nidos parasitados y no parasitados se debió a que el primer hospedador no rechaza huevos parásitos del morfo manchado o blanco 16, mientras que el segundo fue estudiado en una zona donde no hay huevos de dicho morfo (Astié y Reboreda, datos no publicados).
Resultados
Frecuencia e intensidad de parasitismo
La información recopilada en los 28 estudios analizados (Tabla 1) muestra que, si bien el rango de frecuencias de parasitismo es muy amplio (3–94%), en la mayoría de los casos éstas son mayores de 50%. Estos estudios muestran una asociación positiva entre la frecuencia y la intensidad de parasitismo (Correlación de Rangos de Spearman, rS = 0.86, Z = 4.4, P < 0.001; Fig. 1). Esta asociación continúa siendo altamente significativa aún cuando se excluyan del análisis los estudios que presentan frecuencias extremas de parasitismo (i.e., Sicalis flaveola 37 y Troglodytes aedon 35; rS = 0.82, Z = 4.1, P < 0.001). La relación entre intensidad y frecuencia de parasitismo observada se ajusta a la relación esperada en el caso en que la puesta de huevos de Molothrus bonariensis en los nidos del hospedador fuese al azar y, por lo tanto, siga una distribución de Poisson (Fig. 1).
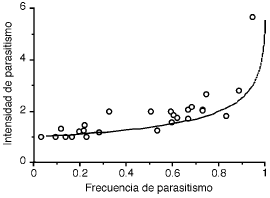
Figura 1. Intensidad de parasitismo de Molothrus bonariensis en función de la frecuencia de parasitismo. Los puntos corresponden a 28 estudios realizados en 15 especies de hospedadores (ver Tabla 1). La línea continua indica la relación intensidad-frecuencia esperada según una distribución de Poisson (ver Resultados).
Efecto del parasitismo sobre distintos componentes del éxito reproductivo del hospedador
En la tabla 2 se presenta un análisis cualitativo de los resultados de 12 estudios realizados en 8 hospedadores, a partir de los cuales se pudo estimar el efecto del parasitismo sobre la supervivencia de los nidos del hospedador y sobre la eficiencia reproductiva en nidos exitosos. Estos estudios incluyen hospedadores con un rango de tamaños que va desde los 15 g (Troglodytes aedon) hasta los 80 g (Pseudoleistes virescens). El parasitismo de cría disminuyó la supervivencia de los nidos del hospedador sólo en las especies de menor tamaño corporal (Troglodytes aedon, Fluvicola albiventer y Zonotrichia capensis). En estos casos, el abandono de los nidos se produjo como consecuencia de la destrucción de la mayoría de los huevos del hospedador. En relación con la eficiencia reproductiva del hospedador, ésta se redujo en todos los casos analizados.
Tabla 2. Impacto del parasitismo de Molothrus bonariensis sobre la supervivencia de los nidos del hospedador y su éxito reproductivo total. Para cada hospedador se indica la frecuencia e intensidad de parasitismo, el número de nidos reportados, si existen diferencias significativas (S), o no (NS) entre nidos no parasitados y parasitados en la supervivencia durante el período de puesta e incubación (P < 0.05, a dos colas), y si existen diferencias significativas entre nidos no parasitados y parasitados en la eficiencia reproductiva (P < 0.05, a dos colas). Los asteriscos indican estudios en los que se calculó la supervivencia diaria utilizando el método de Mayfield 46; en los restantes casos se compararon las proporciones de nidos exitosos y no exitosos (ver Métodos). Para cada estudio se indica la fuente bibliográfica utilizada. En todos los estudios la subespecie de Molothrus bonariensis analizada fue Molothrus bonariensis bonariensis excepto donde se indica. Los hospedadores están ordenados por tamaño corporal creciente.

Con el propósito de evaluar qué componentes de la eficiencia reproductiva del hospedador fueron mayormente afectados por el parasitismo se comparó la supervivencia de huevos, el éxito de eclosión y la supervivencia de pichones entre nidos no parasitados y parasitados. En la tabla 3 se presentan los resultados de siete estudios realizados en seis hospedadores en los que se recopiló dicha información. En casi todos los casos (seis de siete estudios), el parasitismo redujo la supervivencia de los huevos del hospedador en forma significativa (rango: 12–25%). El éxito de eclosión y la supervivencia de pichones solo se redujeron en dos de siete y en uno de siete casos, respectivamente. Si se tiene en cuenta el efecto del parasitismo sobre las tres variables en forma combinada, la reducción de la eficiencia reproductiva del hospedador fue de 30–55% para los hospedadores de tamaño similar o mayor a Molothrus bonariensis y de un 81% para el hospedador más pequeño (Troglodytes aedon).
Tabla 3. Impacto del parasitismo de Molothrus bonariensis sobre la supervivencia de huevos, el éxito de eclosión y la supervivencia de pichones del hospedador. Para cada hospedador se indica su peso corporal y la frecuencia e intensidad de parasitismo. Los datos se expresan como promedio ± EE, con el número de nidos utilizados entre paréntesis. Los valores de P corresponden al resultado de la comparación del promedio entre nidos no parasitados (NP) y parasitados (PA) mediante la Prueba de Mann-Whitney. Para cada estudio se indica la fuente bibliográfica utilizada. En todos los estudios la subespecie de Molothrus bonariensis analizada fue Molothrus bonariensis bonariensis (peso: 45–50 g).
Efecto de la intensidad del parasitismo sobre la supervivencia de huevos del hospedador
Como se mostró previamente (Fig. 1, Tablas 2 y 3), la mayoría de los hospedadores de Molothrus bonariensis tiene intensidades promedio de parasitismo de 1.5–2 huevos por nido. En estos hospedadores los nidos parasitados reciben entre 1 y 4–6 huevos de Molothrus bonariensis. Teniendo en cuenta que el principal efecto del parasitismo es la destrucción de huevos del hospedador como resultado de las picaduras de Molothrus bonariensis cuando visita sus nidos, se espera que este costo aumente con la intensidad de parasitismo. Para poner a prueba esta predicción se analizó el efecto de la intensidad de parasitismo sobre la supervivencia de huevos en tres hospedadores (Pseudoleistes virescens, Mimus saturninus y Turdus amaurochalinus). En estas especies la frecuencia de parasitismo es de 60–83%, mientras que su intensidad varía entre 1.6–2.2 huevos por nido (Tabla 3).
Los resultados de este análisis se presentan en forma gráfica en la figura 2. En los tres hospedadores se observó una asociación negativa significativa entre la intensidad de parasitismo y la supervivencia de sus huevos. La proporción de huevos perdidos durante los estadíos de puesta e incubación en nidos que recibieron cuatro huevos parásitos varió entre 42% (Pseudoleistes virescens) y 83% (Turdus amaurochalinus).
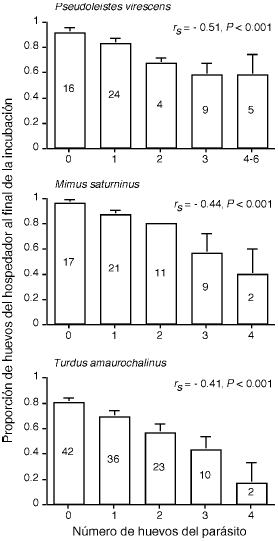
Figura 2. Promedio (+ EE) de la proporción de huevos del hospedador presentes en el nido al final del período de incubación en función del número de huevos parásitos puestos en el nido por Molothrus bonariensis. Los números indican el número de nidos. Se muestra el valor del Análisis de Correlación de Rangos de Spearman entre la proporción de huevos puestos que llegan al final de la incubación y la intensidad de parasitismo. Los datos de Pseudoleistes virescens fueron obtenidos de Mermoz 44, mientras que los de Mimus saturninus y Turdus amaurochalinus son de Rabuffetti y Reboreda (datos no publicados) y de Astié y Reboreda (datos no publicados), respectivamente.
Costos producidos por Molothrus bonariensis en nidos no parasitados
En la sección anterior se mostró que existe una asociación entre la intensidad de parasitismo y la destrucción de huevos del hospedador. La figura 2 también indica que se produjeron pérdidas de huevos del hospedador en nidos no parasitados. Si bien este resultado (i.e., la destrucción de huevos del hospedador por picaduras producidas por los tordos en nidos no parasitados) ha sido mencionado en otros trabajos, su impacto sobre el éxito reproductivo del hospedador ha sido poco estudiado. Una de las dificultades para cuantificar dicho costo es que varios de los hospedadores que han sido estudiados rechazan los huevos parásitos del morfo blanco por lo que no resulta posible discriminar entre nidos no parasitados y nidos parasitados con huevos de este morfo. En esta sección se analizan los costos producidos por Molothrus bonariensis en nidos no parasitados en dos hospedadores, Agelaius thilius y Turdus amaurochalinus, en los que es posible discriminar sin ambigüedad entre nidos no parasitados y parasitados (ver Métodos).
En la figura 3 puede observarse que, si bien la frecuencia de nidos con picaduras es mayor en el grupo "parasitados" que en el "no parasitados" (80% vs. 38% en Agelaius thilius y 64% vs. 52% en Turdus amaurochalinus), la proporción de nidos no parasitados con picaduras es importante. Tanto en el grupo "no parasitados" como en el "parasitados" la pérdida de huevos fue mayor en los nidos en que se registraron picaduras que en los que éstas no ocurrieron (Prueba de Kruskal-Wallis; Agelaius thilius: H = 53.8, P < 0.001, comparaciones múltiples P < 0.01; Turdus amaurochalinus: H = 28.5, P < 0.001, comparaciones múltiples P < 0.01). Por otra parte, aunque el efecto de las picaduras fue mayor en el grupo de nidos parasitados que en el de no parasitados (Agelaius thilius: P < 0.05; Turdus amaurochalinus: P < 0.01), en este último grupo las picaduras produjeron la pérdida del 22% y del 24% de los huevos de Agelaius thilius y Turdus amaurochalinus, respectivamente.

Figura 3. Promedio (+ EE) de la proporción de huevos puestos por el hospedador presentes en el nido al final del período de incubación en nidos no parasitados y parasitados por Molothrus bonariensis en los que no se detectaron pérdidas de huevos por picaduras (barras blancas) o en los que sí se detectaron pérdidas por picaduras (barras negras). Los números indican el número de nidos. Los datos de Agelaius thilius fueron obtenidos de Massoni 51, mientras que los de Turdus amaurochalinus son de Astié y Reboreda (datos no publicados).
Discusión
Los resultados presentados muestran que el principal impacto del parasitismo sobre el éxito reproductivo de la mayoría de los hospedadores estudiados es la destrucción de sus huevos como resultado de las picaduras producidas por Molothrus bonariensis. El comportamiento de picadura de huevos también reduce el éxito reproductivo del hospedador en nidos en los que el parásito no pone huevos. El efecto negativo del parasitismo sobre el éxito de eclosión y la supervivencia de los pichones del hospedador es menos frecuente y se manifiesta principalmente en los hospedadores que tienen un tamaño corporal mucho menor al del parásito. En estos hospedadores, el parasitismo también aumenta la probabilidad de abandono del nido, generalmente como resultado de la destrucción de la mayoría de sus huevos. Nuestros resultados también muestran que el costo producido por la picadura de huevos aumenta con la intensidad de parasitismo. Si se tiene en cuenta que la intensidad y la frecuencia de parasitismo están estrechamente asociadas, esta última sería un buen indicador del impacto que el parasitismo de Molothrus bonariensis puede estar produciendo sobre el éxito reproductivo de los hospedadores.
Como ya fuera mencionado, muchas especies de paseriformes vuelven a nidificar si su nido es predado pero no lo hacen si éste fue exitoso. Los resultados presentados en este trabajo indican que la mayoría de los hospedadores de Molothrus bonariensis no abandonan los nidos parasitados, pero como resultado de la alta intensidad de parasitismo producen muy pocos volantones. En el caso en que estos hospedadores no vuelvan a nidificar luego de un intento reproductivo exitoso, el impacto del parasitismo sobre sus poblaciones podría ser de una importancia comparable al de predación de nidos, aún siendo menor su frecuencia 1.
Nuestros resultados muestran que el parasitismo de Molothrus bonariensis reduce en forma importante el éxito reproductivo de los hospedadores. Sin embargo, esta información resulta insuficiente para predecir el impacto del parasitismo sobre la viabilidad de las poblaciones de los hospedadores. En tal sentido, resulta importante estudiar algunos aspectos poco conocidos de la biología del parásito y de sus hospedadores y desarrollar modelos de su dinámica poblacional que incorporen esta información, así como otras variables que no han sido consideradas hasta el presente. A continuación se presentan brevemente algunos de estos aspectos.
Para la mayoría de los hospedadores de Molothrus bonariensis se desconoce (1) el número de intentos reproductivos que realiza cada pareja durante una temporada y si éste es modificado por el parasitismo, (2) las tasas de supervivencia anual de los juveniles y de los adultos y si éstas son reducidas por el parasitismo, y (3) el éxito reproductivo que tiene Molothrus bonariensis en sus nidos y el efecto de la competencia intraespecífica sobre esta variable 52. Esta información es fundamental para poder estimar el número de hospedadores que van a reproducirse en el futuro y la frecuencia e intensidad de parasitismo que van a experimentar. Esto, a su vez, permitirá predecir si los tamaños de las poblaciones del hospedador y del parásito van a mantenerse estables, aumentarán o disminuirán 18.
Si bien se sabe que Molothrus bonariensis es un parásito generalista, se desconoce si el generalismo se manifiesta a escala poblacional (i.e., una hembra utiliza solo un hospedador, pero distintas hembras parasitan distintos hospedadores) o si éste ocurre a escala individual (i.e., una hembra utiliza diferentes hospedadores), y, en este último caso, si existen preferencias por algún hospedador en particular. En el caso en que las hembras de Molothrus bonariensis utilicen durante su vida un solo hospedador (i.e., si las hembras forman razas hospedador-específicas), el tamaño poblacional de cada raza parásita va a estar asociado en forma denso-dependiente al del hospedador que utiliza, por lo que se espera que los tamaños poblacionales lleguen a valores de equilibrio. Por el contrario, si una misma hembra utiliza diferentes hospedadores, el parasitismo puede resultar una seria amenaza para hospedadores poco abundantes o preferidos, ya que la población del parásito va a depender mayormente de los hospedadores más abundantes y no se verá afectada en forma importante por una reducción del tamaño poblacional de los hospedadores poco abundantes o preferidos. Por lo tanto, estos hospedadores pueden experimentar aumentos en la frecuencia de parasitismo aun cuando sus poblaciones estén disminuyendo 18,20.
Los estudios realizados en Molothrus ater indican que una hembra puede utilizar más de un hospedador en una misma temporada reproductiva 53,54. Sin embargo, estos últimos autores también encontraron que aproximadamente el 50% de las hembras de esta especie utilizaron un único hospedador. A partir de estos resultados, ellos sugieren que la población de Molothrus ater podría estar integrada por una combinación de hembras hospedador-especialistas y hospedador-generalistas 54. Como se mencionó anteriormente, conocer cuál de estas estrategias presentan las hembras de Molothrus bonariensis tiene particular interés en aquellos casos en que el parasitismo reduce en forma significativa el éxito reproductivo de los hospedadores, ya que la viabilidad de sus poblaciones puede diferir según la estrategia utilizada.
Si bien algunos estudios han analizado el posible valor adaptativo de la picadura y destrucción de huevos en nidos no parasitados 55,56, su impacto sobre el éxito reproductivo del hospedador casi no ha sido tenido en cuenta. La mayoría de los trabajos han estimado el impacto de Molothrus bonariensis comparando el éxito reproductivo de los hospedadores en nidos no parasitados y parasitados. Como se mostró anteriormente, el costo producido por la picadura de huevos en nidos no parasitados es importante y en algunos hospedadores (e.g., Agelaius thilius, Turdus amaurochalinus) puede producir el abandono del nido (Massoni y Reboreda 16 y Astié y Reboreda, datos no publicados).
Por último, los modelos que describen la dinámica poblacional de los sistemas parásito de cría-hospedador (e.g., May y Robinson 18) no tienen en cuenta algunas características relevantes de la biología de los parásitos (como el comportamiento de picadura de huevos del hospedador) y de sus hospedadores (como el abandono de nidos con destrucción masiva de huevos o con alta intensidad de parasitismo, o la probabilidad de volver a nidificar). La incorporación de estas variable en nuevos modelos permitirá predecir en forma más precisa las consecuencias del parasitismo sobre la viabilidad de las poblaciones de los hospedadores.
Agradecimientos. Vanina D. Fiorini realizó valiosas sugerencias a una versión previa de este trabajo. Durante la realización de este estudio Andrea A. Astié y Viviana Massoni fueron becarias de postgrado y postdoctoral, respectivamente, del Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). Fabian L. Rabuffetti fue becario doctoral del FOMEC. Myriam E. Mermoz y Juan C. Reboreda son miembros de la Carrera del Investigador Científico del CONICET. Este trabajo fue financiado con subsidios del CONICET (PID 0798/98), de la Universidad de Buenos Aires (TW88) y de la Agencia Nacional de Promoción Científica y Tecnológica (01-09237).
Bibliografía Citada
1 Rothstein SI (1990) A model system for coevolution: avian brood parasitism. Annual Review of Ecology and Systematics 21:481–508 [ Links ]
2 Davies NB (2000) Cuckoos, cowbirds and other cheats. Oxford University Press, Oxford [ Links ]
3 Lanyon SM (1992) Interspecific brood parasitism in blackbirds (Icterinae): a phylogenetic perspective. Science 255:77–79 [ Links ]
4 Johnson KP y Lanyon SM (1999) Molecular systematics of the grackles and allies, and the effect of additional sequence (Cyt b and ND2). Auk 116:759-768 [ Links ]
5 Ortega C (1998) Cowbirds and other brood parasites. University of Arizona Press, Tucson [ Links ]
6 Rothstein SI y Robinson SK (1998) The evolution and ecology of avian brood parasitism. Pp. 3–56 en: Rothstein SI y Robinson SK (eds) Parasitic birds and their hosts: studies in coevolution. Oxford University Press, New York [ Links ]
7 Hudson WH (1874) Notes on the procreant instincts of the three species of Molothrus found in Buenos Aires. Proceedings of the Zoological Society 11:153–174 [ Links ]
8 Hoy G y Ottow J (1964) Biological and oological studies of the molothrine cowbirds (Icteridae) of Argentina. Auk 81:186–203 [ Links ]
9 Sealy SG (1992) Removal of Yellow Warbler eggs in association with cowbird parasitism. Condor 94:40–54 [ Links ]
10 Massoni V y Reboreda JC (2002) A neglected cost of brood parasitism: egg punctures by Shiny Cowbirds during inspection of potential host nests. Condor 104:407–411 [ Links ]
11 Carter MD (1986) The parasitic behavior of the Bronzed Cowbird (Molothrus aeneus) in south Texas, USA. Condor 88:11–25 [ Links ]
12 Petit LJ (1991) Adaptive tolerance of cowbird parasitism by Prothonotary Warblers. A consequence of nest-site limitation. Animal Behaviour 41:425–432 [ Links ]
13 King JR (1973) Reproductive relationships of the Rufous-collared Sparrow and the Shiny Cowbird. Auk 90:19–34 [ Links ]
14 Marvil RE y Cruz A (1989) Impact of Brown-headed Cowbird parasitism on the reproductive success of the Solitary Vireo. Auk 106:476–480 [ Links ]
15 Payne RB y Payne LL (1998) Brood parasitism by cowbirds: risks and effects on reproductive success and survival in Indigo Buntings. Behavioral Ecology 9:64–73 [ Links ]
16 Massoni V y Reboreda JC (1998) Costs of brood parasitism and the lack of defenses on the Yellow-winged Blackbird-Shiny Cowbird system. Behavioral Ecology and Sociobiology 42:273–280 [ Links ]
17 Clotfelter ED y Yasukawa K (1999) The effect of aggregated nesting on Red-winged Blackbird nest success and brood parasitism by Brown-headed Cowbirds. Condor 101:729–736 [ Links ]
18 May RM y Robinson SK (1985) Population dynamics of avian brood parasitism. American Naturalist 130:161–167 [ Links ]
19 Pease CM y Grzybowski JA (1995) Assessing the consequences of brood parasitism and nest predation on seasonal fecundity in passerine birds. Auk 112:343-363 [ Links ]
20 Trine ChL, Robinson WD y Robinson SK (1998) Consequences of Brown-headed Cowbird parasitism for host population dynamics. Pp. 273–295 en: Rothstein SI y Robinson SK (eds) Parasitic birds and their hosts: studies in coevolution. Oxford University Press, New York [ Links ]
21 Robinson SK, Rothstein SI, Brittingham MC, Petit LJ y Grzybowski JA (1995) Ecology and behaviour of cowbirds and their impact on host populations. Pp. 428–460 en: Martin TE y Finch DM (eds) Ecology and management of Neotropical migratory birds. Oxford University Press, New York [ Links ]
22 Robinson SK, Thompson III FR, Donovan TM, Whitehead DR y Faaborg J (1995) Regional forest fragmentation and the nesting success of migratory birds. Science 267:1987–1990 [ Links ]
23 Friedmann H y Kiff LF (1985) The parasitic cowbirds and their hosts. Proceedings of the Western Foundation of Vertebrate Zoology 2:226–304 [ Links ]
24 Lowther P y Post W (1999) Shiny Cowbird (Molothrus bonariensis). Pp. 1-24 en: Poole A y Gill F (eds) The birds of North America. Academy of Natural Sciences y American Ornithologist's Union, Philadelphia y Washington DC [ Links ]
25 Fraga RM (2002) Notes on new or rarely reported Shiny Cowbird hosts from Argentina. Journal of Field Ornithology 73:213–219 [ Links ]
26 Friedmann H (1929) The cowbirds: a study in the biology of social parasitism. C. C. Thomas, Springfield [ Links ]
27 Cruz A, Manolis T y Wiley JW (1985) The Shiny Cowbird: a brood parasite expanding its range in the caribbean region. Ornithological Monographs 36:607–620 [ Links ]
28 Wiley JW, Post W y Cruz A (1991) Conservation of the Yellow-shouldered Blackbird Agelaius xanthomus, an endagered West Indian Species. Biological Conservation 55:119–138 [ Links ]
29 Post W, Cruz A y McNair DB (1993) The North-American invasion pattern of the Shiny Cowbird. Journal of Field Ornithology 64:32–41 [ Links ]
30 Cruz A, Post W, Wiley JW, Ortega CP, Nakamura TK y Prather JW (1998) Potential impacts of cowbird range expansion in Florida. Pp. 313–336 en: Rothstein SI y Robinson SK (eds) Parasitic birds and their hosts: studies in coevolution. Oxford University Press, New York [ Links ]
31 Fraga RM, Casañas H y Pugnali G (1998) Natural history and conservation of the endangered Saffron-cowled Blackbird Xanthopsar flavus in Argentina. Bird Conservation International 8:255–267 [ Links ]
32 BirdLife International (2000) Threatened birds of the world. Lynx Edicions y BirdLife International, Barcelona y Cambridge [ Links ]
33 de la Peña MR (1995) Ciclo reproductivo de las aves argentinas. Primera parte. Universidad Nacional del Litoral, Santa Fe [ Links ]
34 Cruz A y Andrews RW (1997) The breeding biology of the Pied Water-Tyrant and its interactions with the Shiny Cowbird in Venezuela. Journal of Field Ornithology 68:91–97 [ Links ]
35 Kattan GH (1998) Impact of brood parasitism. Why do House Wrens accept Shiny Cowbird eggs? Pp. 212–220 en: Rothstein SI y Robinson SK (eds) Parasitic birds and their hosts: studies in coevolution. Oxford University Press, New York [ Links ]
36 Fraga RM (1985) Host-parasite interactions between Chalk-browed Mockingbirds and Shiny Cowbirds. Ornithological Monographs 36:829–844 [ Links ]
37 Mason P (1986) Brood parasitism in a host generalist, the Shiny Cowbird (Molothrus bonariensis): II. Host selection. Auk 103:61–69 [ Links ]
38 Sackmann P y Reboreda JC (2003) A comparative study of Shiny Cowbird parasitism in two large hosts: Chalk-browed Mockingbird and Rufous-bellied Thrush. Condor 105:728–736 [ Links ]
39 Salvador SA (1984) Estudio de parasitismo de cría del Renegrido (Molothrus bonariensis) en Calandria (Mimus saturninus) en Villa María, Córdoba. Hornero 12:141–149 [ Links ]
40 Llambías P (1999) Biología reproductiva del Zorzal Colorado, Turdus rufiventris: costos del parasitismo de cría del Tordo Renegrido, Molothrus bonariensis. Tesis de licenciatura, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires [ Links ]
41 Fraga RM (1978) The Rufous-collared Sparrow as a host of the Shiny Cowbird. Wilson Bulletin 90:271–284 [ Links ]
42 Sick H (1958) Notas biologicas sobre o gaudeiro, Molothrus bonariensis (Gmelin) (Icteridae, Aves). Revista Brasileira de Biologia 18:417–431 [ Links ]
43 Lyon BE (1997) Spatial patterns of Shiny Cowbird brood parasitism on Chestnut-capped Blackbirds. Animal Behaviour 54:927–939 [ Links ]
44 Mermoz ME (1996) Interacciones entre el Tordo Renegrido Molothrus bonariensis y el Pecho Amarillo Pseudoleistes virescens: estrategias del parásito de cría y mecanismos de defensa del hospedador. Tesis doctoral, Universidad de Buenos Aires, Buenos Aires [ Links ]
45 Siegel S y Castellan NJ (1988) Nonparametric statistics for the behavioral sciences. Segunda edición. McGraw-Hill, New York [ Links ]
46 Mayfield H (1975) Suggestions for calculating nest success. Wilson Bulletin 87:456–466 [ Links ]
47 Hines JE y Sauer R (1989) Program CONTRAST — A general program for the analysis of several survival or recovery rate estimates. US Fish and Wildlife Service Technical Report 24, Washington DC [ Links ]
48 Mermoz ME y Reboreda JC (1994) Brood parasitism of the Shiny Cowbird, Molothrus bonariensis on the Brown-and-yellow Marshbird, Pseudoleistes virescens. Condor 96:716-721 [ Links ]
49 Mermoz ME y Reboreda JC (1999) Egg laying behavior by Shiny Cowbirds parasitizing Brown-and-yellow Marshbirds. Animal Behaviour 58:873–882 [ Links ]
50 Mermoz ME y Reboreda JC (1998) Nesting success in Brown-and-yellow Marshbirds: effects of time of the breeding season, nest site and brood parasitism. Auk 115:871–878 [ Links ]
51 Massoni V (2002) Parasitismo de cría del Tordo Renegrido sobre el Varillero Ala Amarilla. Tesis doctoral, Universidad de Buenos Aires, Buenos Aires [ Links ]
52 Mermoz ME y Reboreda JC (2003) Reproductive success of Shiny Cowbird (Molothrus bonariensis) parasitizing the larger Brown-and-yellow Marshbird (Pseudoleistes virescens). Auk 120:1128–1139 [ Links ]
53 Fleischer RC (1985) A new technique to identify and assess the dispersion of eggs of individual brood parasites. Behavioural Ecology and Sociobiology 17:91–99 [ Links ]
54 Alderson GW, Gibbs HL y Sealy SG (1999) Determining the reproductive behaviour of individual brown-headed cowbirds using microsatellite DNA markers. Animal Behaviour 58:895–905 [ Links ]
55 Arcese P, Smith JNM y Hatch MI (1996) Nest predation by cowbirds and its consequences for passerine demography. Proceedings of the National Academy of Sciences 93:4608–4611 [ Links ]
56 Massoni V y Reboreda JC (1999) Egg puncture allows shiny cowbirds to assess host egg development, and suitability for parasitism. Proceedings of the Royal Society of London B 266:1871–1874 [ Links ]
57 Mason P (1986) Brood parasitism in a host generalist, the Shiny Cowbird (Molothrus bonariensis): I. The quality of different species as hosts. Auk 103:52–67 [ Links ]
58 Cruz A, Manolis TD y Andrews RW (1990) Reproductive interactions of the Shiny Cowbird Molothrus bonariensis and the Yellow-hooded Blackbird Agelaius icterocephalus in Trinidad, West Indies. Ibis 132:436-444 [ Links ]

