










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEl hornero
versión impresa ISSN 0073-3407versión On-line ISSN 1850-4884
Hornero v.18 n.2 Buenos Aires sept./dic. 2003
Comunidades de aves de bosques fluviales habitados y no habitados por el hombre en el río Paraná medio, Argentina
María Alejandra Rossetti 1,2,3 y Alejandro R. Giraudo 1,2
1 Instituto Nacional de Limnología, CONICET-UNL. José Maciá 1933, 3016 Santo Tomé, Santa Fe, Argentina.
2 Facultad de Humanidades y Ciencias, Universidad Nacional del Litoral. Paraje El Pozo, 3000 Santa Fe, Santa Fe, Argentina.
3 marialerossetti@yahoo.com.ar
Resumen
Los bosques riparios del río Paraná medio son sitios frecuentemente utilizados para la construcción de viviendas humanas. No obstante, no se conocen los efectos de estas modificaciones sobre las comunidades de aves de estos bosques. Se comparó la avifauna de bosques fluviales habitados y no habitados por el hombre, para evaluar posibles cambios en las comunidades. Se realizaron 60 conteos en puntos, 30 en bosques habitados y 30 en bosques no habitados. Se compararon la riqueza, la abundancia, la equitatividad y la diversidad de aves entre las dos situaciones. Se registraron 60 especies y 591 individuos en los bosques no habitados, y 53 especies y 646 individuos en los habitados. No se detectaron diferencias significativas en la riqueza ni en la diversidad. La abundancia de aves fue significativamente mayor en los bosques habitados. Esto puede haber sido debido a que las comunidades de aves tuvieron pocas especies de bosque con respecto a especies de hábitats no boscosos o generalistas. Las variables estudiadas podrían no haber reflejado los cambios ambientales, o bien el tamaño de la muestra puede haber sido insuficiente para detectarlos. El relativo bajo impacto que producen las viviendas aisladas podría ser poco significativo para las aves en relación con otras perturbaciones locales. Otros factores, tales como las inundaciones y la estacionalidad, también podrían haber influido en estos resultados. Las diferencias registradas en la abundancia de algunas especies de hábitats humanos y boscosos muestran que hay cambios a nivel específico que deberían ser estudiados.
Palabras clave: abundancia, aves, bosques fluviales, diversidad, perturbación humana, riqueza.
Abstract
Bird communities of human inhabited and uninhabited riparian forests in the Middle Paraná River, Argentina
The riparian forests in the Middle Paraná River are places frequently used for the construction of human housings. However, the effects of these modifications on forest bird communities are unknown. We compared bird communities of human inhabited and uninhabited riparian forests in order to assess possible changes. We carried out 60 point counts, 30 in inhabited forests and 30 in uninhabited forests. We evaluated species richness, abundance, evenness and diversity between both types of forests. We recorded 60 species and 591 individuals in uninhabited forests, and 53 species and 646 individuals in inhabited forests. We did not detect significant differences in richness and diversity. Bird abundance was significantly higher in inhabited forests. These results could be related with the composition of the communities: they have few forest species with respect to non-forest or generalist species. The variables we studied could not be reflecting environmental changes, or sample size could be insufficient to detect the differences. The relative low impact that isolated housings produce could be not significant for bird communities with respect to other local disturbances. Other factors, such as floods and seasonality could have also influence on our results. Recorded differences in the abundance of some species of forest and human habitats showed that changes at the specific level should be studied.
Key words: abundance, birds, diversity, human disturbance, richness, riparian forests.
Recibido 5 abril 2003, aceptado 15 diciembre 2003
La utilización de los ambientes naturales para actividades económicas y urbanización ha dado como resultado un paisaje altamente modificado. Solo en aquellos lugares de difícil acceso o donde los suelos no son aptos para la explotación agropecuaria se pueden observar relictos de un paisaje natural. Esta reestructuración del hábitat no es reciente y es un proceso con una tasa alarmante en el mundo (Wiens 1994). En América Latina, entre 1850 y 1985, aproximadamente 370000000 ha de bosques (el 28% del área de bosque en 1850) fueron reemplazadas por algún otro tipo de ecosistema (Houghton et al. 1991), resultando de esta modificación áreas degradadas y fragmentadas.
La degradación de un ambiente afecta la asociación entre la comunidad de plantas y animales silvestres, pudiéndose detectar diferencias en la reacción de los organismos a los cambios ambientales (Croonquist y Brooks 1991). Frecuentemente, las perturbaciones tienen efectos perjudiciales sobre la fauna (Hume 1976, citado en Gill et al. 1996), aunque esto depende de las características de la comunidad. En algunos casos, existe un claro vínculo entre la magnitud de la perturbación y la supervivencia y reproducción de los individuos, pero en muchos otros las perturbaciones actúan de una forma más sutil, reduciendo el acceso a los recursos alimenticios o a los sitios para nidificar. Estudios sobre tales efectos muestran que la redistribución de los organismos después de una perturbación no necesariamente refleja un efecto negativo, ya que el nuevo patrón de distribución puede ser temporario; después de un tiempo los animales podrían retornar a su distribución original para explotar el remanente de los recursos (Gill et al. 1996).
Los bosques riparios y las áreas cercanas a los cursos de aguas están entre los sistemas naturales más productivos y valiosos del planeta (Hunt 1985), pero han sido degradados en su mayoría (Croonquist y Brooks 1991, 1993). Estos bosques son importantes unidades de los sistemas fluviales, que actúan como zonas ecotonales entre el río y tierras adentro, albergando una alta diversidad de aves que responde a la habilidad adaptativa de esta comunidad (Dobrowolski 1997).
El valle aluvial del río Paraná es un extenso humedal que posee bosques fluviales, entre otros hábitats. En estos ambientes situados entre climas subtropicales y templados, la disponibilidad de recursos varía estacionalmente y año tras año (Beltzer y Neiff 1992). Durante crecientes excepcionales, cuando el valle del Paraná está cubierto por una única masa de agua, las copas de los árboles son las únicas unidades emergentes, ofreciendo sitios de refugio temporario a la fauna (Bonetto 1986). De esta manera, de las características del régimen hidrosedimentológico pulsátil dependen la oferta de hábitat (alimentación, refugio, nidificación) y la complejidad específica y las características de las poblaciones animales (Beltzer y Neiff 1992). En el río Paraná estos bosques se sitúan en las partes más altas o albardones, sitios frecuentemente utilizados para la construcción de viviendas humanas con el objeto de disminuir el riesgo de inundaciones. No existen antecedentes sobre el efecto de este tipo de asentamientos sobre las comunidades de aves de estos bosques riparios. Por esta razón, se realizó un análisis exploratorio con el objetivo de comparar la avifauna en bosques fluviales habitados y no habitados por el hombre.
Métodos
Área de estudio
El área de muestreo se ubica entre las localidades de Alto Verde (31°40'S, 60°41'O) y Cayastá (31°12'S, 60°10'O), provincia de Santa Fe (Fig. 1). Los bosques fluviales del río Paraná y sus afluentes son incluidos fitogeográficamente en la Provincia Paranaense (Cabrera 1976). Otros autores consideran a las sabanas inundables del río Paraná medio e inferior como una ecoregión con identidad propia (Dinerstein et al. 1995, Burkart et al. 1999). El valle de inundación de este río es un mosaico muy complejo de hábitats con una dinámica fuertemente modelada por los ciclos de crecientes y bajantes (Neiff 1990, Burkart et al. 1999). Las alturas máximas del río Paraná en la provincia de Santa Fe generalmente ocurren a fines de verano y a principios de otoño, como resultado de las precipitaciones en la cuenca superior (D'Angelo 1991). El clima es subhúmedo–húmedo mesotermal, con temperaturas promedio anuales de 18 °C, ocurriendo las dos terceras partes de las precipitaciones entre noviembre y abril, con mínimas entre junio y agosto (Panigatti et al. 1981).
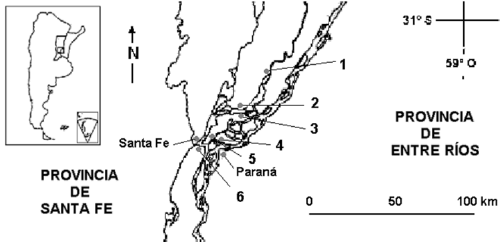
Figura 1. Área de estudio en el río Paraná medio, Argentina, mostrando la ubicación de los sitios de muestreo. 1: Cayastá, 2: Arroyo Leyes, 3: Arroyo Potrero, 4: isla Peruhil, 5: San José del Rincón, 6: Alto Verde.
Los bosques insulares varían desde una sola hilera de árboles a, raramente, más de 100 m de ancho, siendo el sauce (Salix humboldtiana) y el aliso de río (Tessaria integrifolia) especies pioneras que ocupan en general sectores más bajos e inundables. En sitios más altos, otras especies como el seibo (Erythrina crista-galli), el laurel (Nectandra microcarpa), el timbó blanco (Albizia inundata) y el curupí (Sapium haematospermum) enriquecen la composición de estos bosques. El estrato herbáceo está compuesto por especies como Baccharis sp., Sida rhombifolia, Aspilia silphioides, y son frecuentes las enredaderas como Ipomoea spp., Muehlenbeckia sagittifolia y Mikania micrantha (Beltzer y Neiff 1992).
La accesibilidad y la autorización de los pobladores de los bosques ribereños influyeron en la elección de los sitios de muestreo. Las viviendas consideradas correspondieron a construcciones pequeñas, sencillas, utilizadas algunas como casas de fin de semana, y, en algunos casos, a construcciones precarias de pobladores de escasos recursos económicos, principalmente pescadores. La principal y más evidente perturbación que produce este tipo de viviendas localmente consiste en la generación de un pequeño claro en el bosque fluvial, generalmente no mayor a 20–30 m de diámetro. Esta perturbación es claramente diferenciable de otras que existen en todos los bosques (habitados y no habitados), como la tala de árboles, la presencia de especies introducidas de plantas, la presencia de ganado y actividades de cacería. En un 63% de las viviendas se observó actividad permanente.
Muestreo
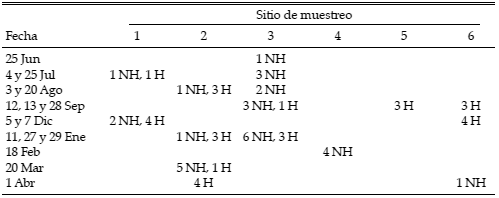
Entre el 25 de junio de 1999 y el 1 de abril de 2000 se realizaron 16 campañas en 6 localidades donde se completaron 60 conteos de aves en puntos, 30 en bosques habitados y 30 en bosques no habitados (Tabla 1). Los conteos fueron distribuidos en cada sitio de muestreo con un diseño sistemático con arranque al azar, separando cada uno de ellos por al menos 250 m (Ralph et al. 1996). Durante 20 min se registraron todas las aves vistas u oídas en un radio máximo de 25 m (Ordano 1999). Los conteos se realizaron siempre en buenas condiciones climáticas, entre el amanecer y las 5 h siguientes (Ralph et al. 1996).
Tabla 1. Distribución temporal de los muestreos de aves en el río Paraná medio, Argentina, entre junio de 1999 y abril de 2000. Se indica el número de puntos de conteo por sitio de muestreo para bosques no habitados (NH) y habitados por el hombre (H). Los códigos de los sitios son los mismos que en la figura 1.
Las variables analizadas fueron riqueza, abundancia, equitatividad (índice de Shannon) y diversidad (índice de Shannon) en cada situación. Estas variables fueron comparadas entre bosques habitados y no habitados mediante pruebas paramétricas (Prueba de t de Student) y no paramétricas (Prueba de Mann-Whitney), según el ajuste de las variables a la distribución normal (Prueba de Kolmogorov-Smirnov) y a su homogeneidad de varianzas (Prueba de Bartlett) (Sokal y Rohlf 1969).
Limitaciones del estudio
Debido a que existen unidades de muestreo en bosques habitados y no habitados en una misma localidad, estas unidades podrían constituir pseudoréplicas, con las limitaciones que esto implica en la interpretación y generalización de los resultados (ver Hurlbert 1984, Eberhardt y Thomas 1991). Los muestreos no estuvieron distribuidos de manera uniforme a lo largo del ciclo anual, por lo que el efecto de la estacionalidad, que puede aportar a la variación observada en la comunidad de aves, no ha sido evaluado. Debido a que es una región con una densidad de población relativamente alta que utiliza el río y los bosques para diversas actividades, todos los bosques presentan otras perturbaciones además de las provocadas por las viviendas. No fue posible encontrar bosques sin perturbación humana (controles) en el área; por lo tanto, los resultados obtenidos deben ser considerados exploratorios y no estrictamente relacionados de manera directa con la perturbación de interés. Existen otras variables (pastoreo, extracción de leña, uso recreativo, inundaciones) que se supone que actúan, en mayor o menor grado, en todos los bosques, aunque se desconocen los efectos que producen.
Resultados
Se registraron 68 especies pertenecientes a 25 familias en las dos situaciones estudiadas. La riqueza total de especies fue levemente superior en los bosques no habitados, con 60 especies, respecto a los bosques habitados, con 53 (Tabla 2). Un total de 45 especies (66% del total) fueron observadas en las dos situaciones, 15 especies (22%) fueron registradas solo en los bosques no habitados y 8 especies (12%) fueron registradas solo en los bosques habitados, aunque en este caso se trata de especies representadas por una escasa cantidad de individuos (8 o menos individuos).
Tabla 2. Abundancia de aves en bosques no habitados (NH) y habitados por el hombre (H) en el río Paraná medio, Argentina. Los datos se expresan como promedio ± EE (n = 30). El signo + indica las especies para las cuales se registró un solo individuo. Las especies están ordenadas en forma decreciente en relación con su abundancia en bosques no habitados. a: Especies propias de bosque o arbustal, estricta o facultativamente (Stotz et al. 1996, Rossetti y Giraudo, datos no publicados).

La riqueza promedio de especies y la diversidad promedio no presentaron diferencias estadísticamente significativas entre bosques no habitados y habitados (Tabla 3). No obstante, la diversidad total fue levemente mayor en los bosques no habitados (H = 3.349) respecto a los bosques habitados (H = 3.307), debido posiblemente a la mayor riqueza total de los primeros, ya que la equitatividad de la abundancia de especies fue levemente mayor en los bosques habitados (E = 0.833) respecto a los no habitados (E = 0.818). Las comunidades de aves, en general, mostraron una menor proporción de aves que dependen, facultativa o estrictamente, de bosques o arbustales (23 especies, 34% del total; véase Tabla 2) que de especies no dependientes de bosque (45 especies, 66%). Éstas incluyen a especies generalistas (e.g., Furnarius rufus, Zonotrichia capensis, Pitangus sulphuratus), aves propias de pastizales (e.g., Nothura maculosa, Vanellus chilensis) o de ambientes acuáticos (e.g., Aramus guarauna).
Tabla 3. Diferencias en la riqueza de especies, la abundancia y la diversidad de aves en bosques no habitados (NH) y habitados por el hombre (H) en el río Paraná medio, Argentina. Los datos se expresan como promedio ± EE (n = 30). Se muestran los valores del estadístico t de Student y U de Mann-Whitney y su nivel de significación.
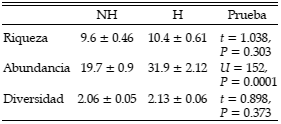
La abundancia promedio de aves fue significativamente más alta en los bosques habitados; fue un 62% más alto en los bosques habitados respecto a los no habitados (Tabla 3). En los bosques habitados se registraron 646 individuos, de los cuales el 53% perteneció a solo ocho especies (15% de las 53 especies registradas en estos bosques): Furnarius rufus, Pitangus sulphuratus, Phacellodomus ruber, Saltator coerulescens, Troglodytes aedon, Zonotrichia capensis, Polioptila dumicola y Molothrus bonariensis (Tabla 2). Los bosques no habitados presentaron 591 individuos, de los cuales el 55% estuvo concentrado en siete especies (el 12% de las allí presentes): Phacellodomus ruber, Furnarius rufus, Polioptila dumicola, Progne tapera, Pitangus sulphuratus, Troglodytes aedon y Saltator coerulescens. Passer domesticus y Molothrus bonariensis, especies propias de o favorecidas por hábitats humanos, presentaron una clara tendencia con mayor abundancia en los bosques habitados respecto a los no habitados (Tabla 2). Si se observan las abundancias de las aves propias de bosque (23 especies; Tabla 2), no hubo diferencias estadísticamente significativas entre los bosques habitados y no habitados en la riqueza de este subconjunto de especies ni en su abundancia, aunque algunas de ellas, como Taraba major, mostraron una tendencia de mayor abundancia en los bosques no habitados (Tabla 2).
Discusión
Los resultados obtenidos deben ser interpretados entendiendo las limitaciones de este estudio (véase Métodos). El estudio no mostró cambios significativos en las variables analizadas al nivel de la comunidad entre bosques habitados y no habitados, con excepción de la abundancia. Esto podría relacionarse con diversos factores. En principio, el elenco de especies observado está constituido en elevada proporción (66%) de aves que no dependen estrictamente de la presencia de bosques. Solo 23 especies (34%) fueron aves características de bosques o arbustales, aunque muchas de ellas son también especies que habitan principalmente en los bordes de bosques, como Tyrannus melancholicus (Lopez de Casenave et al. 1998), e incluso en bosques exóticos, como Colaptes melanochloros (Giraudo, obs. pers.). Tal composición de especies podría deberse a que los bosques fluviales se distribuyen linealmente a lo largo de cursos de agua y son relativamente estrechos, lo que posiblemente provoque un elevado efecto de borde. Esto favorecería a especies de borde y generalistas, así como diversas modificaciones en la abundancia y composición de la avifauna como las observadas y discutidas por Lopez de Casenave et al. (1998) para el Chaco semiárido de Argentina. Las diferencias en abundancia podrían también tener relación con el aumento de la disponibilidad de alimento para muchas especies en los bordes (Lopez de Casenave et al. 1998). Estos resultados coinciden con los de Hennings y Edge (2003), quienes registraron una mayor abundancia de aves nativas en bosques estrechos rodeados por terrenos rurales en Oregon, EEUU.
Las perturbaciones que generan las viviendas pueden aumentar la oferta de hábitats adicionales dentro de los bosques, facilitando la presencia de especies propias de otros hábitats y la mayor abundancia de algunas especies (e.g., aquellas que frecuentan ambientes humanos). El aporte de recursos antes inexistentes para las aves alrededor de las casas, tal como alimentos adicionales (semillas y otros alimentos utilizados para aves de corral y otros animales domésticos, plantas ornamentales o frutales que brindan flores y frutos), podría influir sobre el aumento de la abundancia de aves. Algunas especies, como Passer domesticus, mostraron una mayor abundancia en los bosques habitados, en coincidencia con lo indicado por Sodhi (1992) para hábitats urbanos respecto a rurales. Este podría ser el caso de Molothrus bonariensis, una especie más abundante en las áreas habitadas que generalmente es favorecida por perturbaciones provocadas por el hombre. Varios autores han registrado un aumento en la densidad de varias especies de bosques templados en áreas perturbadas (Karr 1990), y otros estudios reportan una alta abundancia de aves en hábitats urbanos cuando se los compara con ambientes nativos (Aldrich y Coffin 1980, Bessinger y Osborne 1982, Rosenberg et al. 1987, Mills et al. 1989), documentándose en algunos casos que, con el desarrollo urbano, las especies nativas son reemplazadas por especies introducidas (Dance 1986).
Hennings y Edge (2003) observaron en Oregon que el desarrollo de las poblaciones humanas y la urbanización afectan a las comunidades de aves riparias, registrando que la riqueza y diversidad de especies fueron mayores en áreas menos desarrolladas. En este estudio se observó que la riqueza y la diversidad fueron más altas en bosques no habitados, aunque tales diferencias no fueron estadísticamente significativas, lo que podría tener relación con limitaciones metodológicas relacionadas con la potencia de las pruebas. Otro factor que podría haber influido es el relativamente bajo y localizado impacto que ocasionan este tipo de viviendas aisladas, que puede ser menor en relación con los procesos de urbanización. Una perturbación no es siempre necesariamente negativa (Gill et al. 1996). Las viviendas podrían aumentar localmente la heterogeneidad espacial y el efecto de borde, lo que podría influir en la riqueza y abundancia de la comunidad de aves (Karr 1990, Wiens 1994). La influencia de otros factores de disturbio a los que se ven sometidos los bosques fluviales podría tener una magnitud más importante en el modelado de las comunidades de aves y, por lo tanto, enmascarar el efecto de este tipo de viviendas. Los ciclos de crecientes y bajantes del río Paraná han sido considerados una importante perturbación para los bosques fluviales de la región, debido a que determinan modificaciones acentuadas de parámetros ambientales como la estructura de la comunidad de plantas, las cuales representan cambios directos en la oferta de hábitat para las aves, produciéndose con las crecientes una disminución de la heterogeneidad espacial de la vegetación, con una menor oferta de recursos (Beltzer y Neiff 1992, Giraudo 1992). Esto pudo haber influenciado los resultados obtenidos, debido a que, si bien los puntos de conteo no se encontraban en zonas inundables, el agua en algunos casos estuvo muy cerca de ellos. La altura del río Paraná dentro del período de muestreo varió entre 4.27 m en julio de 1999 y 2.10 m en noviembre de 1999 (promedios mensuales), sin que se registraran crecientes o bajantes extremas. Las variaciones estacionales, no analizadas en este trabajo, también podrían ser un factor importante actuando sobre la riqueza y la abundancia de la comunidad de aves (Karr 1976).
La respuesta individual de las especies a las modificaciones humanas puede ser diferente, como se observó para Taraba major, Passer domesticus y Molothrus bonariensis. Banchs y Fraga (1994) mostraron que Paroaria coronata tiene una mayor supervivencia de nidos en bosques chaqueños perturbados por el hombre, lo que podría deberse a numerosos factores que incluyen la disminución de la predación. Las diferencias registradas en la abundancia de las especies antes mencionadas indican la posibilidad de impactos diferenciales a nivel específico, que deberían ser analizados mediante el estudio detallado de la demografía de las especies.
Agradecimientos. A Mariano Ordano, Soledad López Severín y Adolfo H. Beltzer por sus aportes y colaboraciones para llevar adelante este proyecto. A CONICET, organismo donde trabaja el segundo autor. A dos revisores anónimos que aportaron con sus acertadas críticas al mejoramiento de este trabajo.
Bibliografía Citada
Aldrich JW y Coffin RW (1980) Breeding bird populations from forest to suburbia after thirty–seven years. American Birds 34:3–7 [ Links ]
Banchs A y Fraga R (1994) Supervivencia de nidos de cardenal de cresta roja (Paroaria coronata) en ambientes modificados del Chaco Argentino. Vida Silvestre Neotropical 3:91–95 [ Links ]
Beltzer A y Neiff JJ (1992) Distribución de las aves en el valle del río Paraná. Relación con el régimen pulsátil y la vegetación. Ambiente Subtropical 2:77–102 [ Links ]
Bessinger SR y Osborne DA (1982) Effects of urbanization on avian community organization. Condor 84:75–83 [ Links ]
Bonetto AA (1986) The Paraná river system. Pp. 541–555 en: Davies BR y Walker KF (eds) The ecology of river system. W Junk Publishers, Dordrecht [ Links ]
Burkart R, Bárbaro NO, Sánchez RO y Gómez DA (1999) Eco–regiones de la Argentina. Administración de Parques Nacionales, Buenos Aires [ Links ]
Cabrera AL (1976) Regiones fitogeográficas argentinas. Pp. 1–85 en: Enciclopedia argentina de agricultura y jardinería. Tomo II. Fascículo 1. Acme, Buenos Aires [ Links ]
Croonquist MJ y Brooks RP (1991) Use of avian and mammalian guilds as indicator of comulative impacts in riparian wetlands areas. Environmental Management 15:701–714 [ Links ]
Croonquist MJ y Brooks RP (1993) Effects of habitat disturbance on bird communities in riparian corridors. Journal of Soil and Water Conservation 48:65–70 [ Links ]
Dance KW (1986) Avifauna on an urbanizing environment in southern Ontario. 1921–1982. Ontario Birds 4:22–29 [ Links ]
D'Angelo ML (1991) Geografía. Pp. 33–136 en: Nueva enciclopedia de la provincia de Santa Fe. Tomo I. Ediciones Sudamérica, Santa Fe [ Links ]
Dinerstein E, Olson DM, Graham DJ, Webster AL, Primm SA, Bookbinder MP y Ledec G (1995) A conservation assessment of the terrestrial ecoregions of Latin América and Caribbean. World Bank y World Wildlife Fundation, Washington DC [ Links ]
Dobrowolski KA (1997) Bird diversity in ecotonal habitats. Pp. 205–227 en: Lachavanne JB y Juge R (eds) Biodiversity in land–inland water ecotones. Parthenon Pub. y UNESCO, Carnforth y París [ Links ]
Eberhardt LL y Thomas JL (1991) Designing environmental field studies. Ecological Monographs 61:53–73 [ Links ]
Gill J, Sutherland WJ y Watkinson AR (1996) A method to quantify the effects of human disturbance on animal populations. Journal of Applied Ecology 3:786–792 [ Links ]
Giraudo AR (1992) Avifauna de un ambiente lenítico del valle aluvial del Paraná Medio y consideraciones sobre la influencia del ciclo hidrológico en la variación estacional de sus poblaciones. Tesis de Licenciatura, Universidad Nacional de Córdoba, Córdoba [ Links ]
Hennings LA & Edge WD (2003) Riparian bird community structure in Portland, Oregon: habitat, urbanization, and spatial scale patterns. Condor 105:288–302 [ Links ]
Houghton RA, Lefkowitz DS y Skole DL (1991) Changes in the landscape of Latin America. Forest Ecology and Management 38:143–172 [ Links ]
Hume RA (1976) Reactions of goldeneyes to boating. British Birds 69:178–179 [ Links ]
Hunt C (1985) The need for riparian habitat protection. National Wetlands Newsletter 7:5–8 [ Links ]
Hurlbert SH (1984) Pseudoreplication and the design of ecological field experiments. Ecological Monographs 54:107–211 [ Links ]
Karr JR (1976) Seasonality, resource availability, and community diversity in tropical bird communities. American Naturalist 110:973–993 [ Links ]
Karr JR (1990) Interactions between forest birds and their habitats: a comparative synthesis. Pp. 379–386 en: Keast A (ed) Biogeography and ecology of forest bird communities. SPB Academic Publishing, The Hague [ Links ]
Lopez de Casenave J, Pelotto JP, Caziani SM, Mermoz M y Protomastro J (1998) Responses of avian assemblages to a natural edge in Chaco semiarid forest in Argentina. Auk 115:425–435 [ Links ]
Mills GS, Dunning JB Jr y Bates JM (1989) Effects of urbanization on breeding bird community structure in southwestern desert habitats. Condor 91:416–428 [ Links ]
Neiff JJ (1990) Ideas para la interpretación ecológica del Paraná. Interciencia 15:424–441 [ Links ]
Ordano M (1999) Aves de bosque de sauce del río Paraná medio (Argentina): evaluación de la técnica de conteos de puntos. Tesis de Posgrado, Instituto de Enseñanza Superior, Paraná [ Links ]
Panigatti J, Weber J y Pillati O (1981) Estado actual y futuro de los problemas de suelos de Santa Fe. INTA Rafaela, Santa Fe [ Links ]
Ralph CJ, Geupel GR, Pyle P, Martin TE, DeSante DF y Milá B (1996) Manual de métodos de campo para el monitoreo de aves terrestres. General Technical Report PSW–GTR–159, USDA Forest Service, Albany [ Links ]
Rosenberg KV, Terril SB y Rosenberg GH (1987) Value of suburban habitats to desert riparian birds. Wilson Bulletin 99:642–654 [ Links ]
Sodhi NS (1992) Comparison between urban and rural bird communities in prairie Saskatchewan: urbanization and short term population trends. Canadian Field Naturalist 106:210–215 [ Links ]
Sokal RR y Rohlf FJ (1969) Biometría. Principios y métodos estadísticos en la investigación biológica. Blume, Madrid [ Links ]
Stotz DF, Fitzpatrick JW, Parker III TA y Moskovits DF (1996) Neotropical birds. Ecology and conservation. University of Chicago Press, Chicago y Londres [ Links ]
Wiens JA (1994) Habitat fragmentation: island vs. landscape perspectives on bird conservation. Ibis 137:97–104 [ Links ]

