










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEl hornero
versión impresa ISSN 0073-3407versión On-line ISSN 1850-4884
Hornero v.18 n.2 Buenos Aires sept./dic. 2003
Biología reproductiva de los cormoranes Imperial (Phalacrocorax atriceps) y Cuello Negro (P. magellanicus) en el Golfo San Jorge, Chubut, Argentina
Gabriel Punta 1, Pablo Yorio 2,3, Gonzalo Herrera 4 y José Saravia 4
1 Secretaría de Turismo y Áreas Protegidas de la Prov. del Chubut. 9 de Julio 280, 9103 Rawson, Chubut, Argentina.
2 Centro Nacional Patagónico (CONICET). Boulevard Brown 3500, U9120ACV Puerto Madryn, Chubut, Argentina. yorio@cenpat.edu.ar.
3 Wildlife Conservation Society. 2300 Southern Boulevard, Bronx, New York, NY 10460, EEUU.
4 Dirección General de Intereses Marítimos y Pesca Continental de la Provincia del Chubut. 9 de Julio 280, 9103 Rawson, Chubut, Argentina.
Resumen
Estudiamos la biología reproductiva de los cormoranes Imperial (Phalacrocorax atriceps) y Cuello Negro (Phalacrocorax magellanicus) en el Golfo San Jorge, Argentina, durante las temporadas reproductivas de 1991–1993. Los fechas en que fueron observados los primeros huevos variaron entre años, siendo entre mediados de octubre y mediados de noviembre para el Cormorán Imperial, y entre fines de octubre y principio de diciembre para el Cormorán Cuello Negro. Durante 1993, el 87% y el 86% de las parejas de Cormorán Imperial y Cormorán Cuello Negro, respectivamente, completaron su nidada en las cuatro semanas siguientes al inicio de la puesta. Para ambas especies, el número de huevos por nido al mes de iniciada la puesta presentó diferencias significativas entre años. El número promedio de huevos por nido fue similar entre los cormoranes Imperial y Cuello Negro en 1993 (2.4 vs. 2.1, respectivamente) pero significativamente diferente en 1991 (1.9 vs. 0.8). En dos de las temporadas de cría estudiadas, el volumen del tercer huevo fue significativamente menor comparado con los volúmenes de los primeros dos huevos. El período de incubación promedio se estimó en 27.9 días para el Cormorán Imperial. El éxito de eclosión para el Cormorán Imperial fue de 38.2% y 52.5% en las temporadas 1991 y 1992, respectivamente. El incremento en peso y el crecimiento del culmen registraron valores máximos entre la segunda y la tercera semana de vida, y valores próximos al asintótico a los 60 días. El crecimiento del tarso alcanzó valores máximos en la primera semana y valores asintóticos a los 30 días. Durante 1993, el número promedio de pichones sobrevivientes a la independencia por nido fue de 1.13 para el Cormorán Imperial y de 0.83 para el Cormorán Cuello Negro.
Palabras clave: Argentina, biología reproductiva, Cormorán Cuello Negro, Cormorán Imperial, Phalacrocorax atriceps, Phalacrocorax magellanicus.
Abstract
Breeding biology of Imperial Cormorant (Phalacrocorax atriceps) and Rock Shag (P. magellanicus) at Golfo San Jorge, Chubut, Argentina
We studied the breeding biology of Imperial Cormorant Phalacrocorax atriceps and Rock Shag Phalacrocorax magellanicus at Golfo San Jorge, Argentina, from 1991 to 1993. Dates in which first eggs were observed varied among seasons, being between mid October and mid November for Imperial Cormorants, and between late October and early December for Rock Shags. During 1993, 87% and 86% Imperial Cormorant and Rock Shag pairs, respectively, completed their clutches within four weeks after egg-laying started. The number of eggs per nest recorded one month after the start of egg laying for both species was statistically different among years. The average number of eggs per nest was similar between Imperial Cormorants and Rock Shags during 1993 (2.4 vs. 2.1, respectively) but significantly different during 1991 (1.9 vs. 0.8). During two of the study seasons the volume of the third egg was significantly smaller than the first and second eggs. The average length of the incubation period for Imperial Cormorants was estimated at 27.9 days. Hatching success of Imperial Cormorants was 38.2% (1991) and 52.5% (1992). Chick weight increase and culmen growth showed maximum values during the second and third weeks after hatching, reaching asymptotic values at 60 days. Chick tarsus growth reached maximum values during the first week and asymptotic values at 30 days. During 1993 mean number of chicks fledged per nest was 1.13 for Imperial Cormorants and 0.83 for Rock Shags.
Key words: Argentina, breeding biology, Imperial Cormorant, Phalacrocorax atriceps, Phalacrocorax magellanicus, Rock Shag.
Recibido 29 mayo 2003, aceptado 20 diciembre 2003
Los cormoranes Imperial (Phalacrocorax atriceps) y Cuello Negro (Phalacrocorax magellanicus) están ampliamente distribuidos a lo largo de la costa de la provincia de Chubut (Punta y Saravia 1993, Yorio et al. 1998). El Cormorán Imperial es la cuarta especie en orden de importancia numérica en la Patagonia, con colonias de hasta varios miles de individuos, mientras que el Cormorán Cuello Negro posee un bajo tamaño poblacional con colonias de tamaño relativamente pequeño (Yorio et al. 1999). El Cormorán Imperial es el principal productor del recurso guanero, siendo de importancia económica a nivel regional (Punta 1996). En la mayor parte de su distribución las dos especies se reproducen en simpatría, nidificando en colonias adyacentes (Punta 1989, Yorio et al. 1998).
El norte del Golfo San Jorge es uno de los sectores de mayor importancia del litoral patagónico por su diversidad de aves marinas. En este sector costero se reproduce aproximadamente el 35% de los Cormoranes Imperiales y el 45% de los Cormoranes Cuello Negro de la Patagonia. A pesar de la importancia de las poblaciones de cormoranes en el Golfo San Jorge, es poco el conocimiento acerca de los patrones de reproducción en esta zona. La biología reproductiva del Cormorán Imperial en la costa atlántica de la Patagonia ha sido escasamente estudiada, disponiéndose únicamente de datos obtenidos en colonias continentales del norte de Chubut (Malacalza 1984, Yorio et al. 1994, Malacalza y Navas 1996) o en sitios bajo influencia humana en el sector central del Golfo San Jorge, Chubut (Libenson 1997) y en el sur de Santa Cruz (Arrighi y Navarro 1998). Los aspectos de la biología reproductiva del Cormorán Cuello Negro son mucho menos conocidos, disponiéndose sólo de información fragmentaria o descriptiva, mayormente para colonias en la provincia de Chubut (Murphy 1936, Humphrey et al. 1970, Boswall y Prytherch 1972, de la Peña 1980, Siegel-Causey 1986, Punta y Saravia 1993, Malacalza 1995, Libenson 1997, Woods y Woods 1997).
En este trabajo se describe la biología reproductiva de los cormoranes Imperial y Cuello Negro en Bahía Bustamante, al norte del Golfo San Jorge, siendo la primera información proveniente de un área de importancia relevante para ambas especies. Teniendo en cuenta que las poblaciones de esta zona costera pueden estar sometidas a la extracción comercial de guano (Punta 1996) y que están comenzando a recibir visitas turísticas, el conocimiento de la biología reproductiva de ambas especies permitirá elaborar lineamientos de manejo. Además, el presente estudio comunica por primera vez información comparativa sobre la biología reproductiva de este par de especies congenéricas que comparten los mismos sitios de nidificación.
Métodos
Los cormoranes Imperial y Cuello Negro presentan características morfológicas similares, aunque el Cormorán Imperial es de mayor tamaño (peso promedio: 2.2 vs. 1.5 kg; G Punta, datos no publicados). Las observaciones sobre los ciclos reproductivos se efectuaron en las colonias de Isla Isabel (45°07'S, 66°30'O) durante la temporada 1991 y de Isla Galiano (45°06'S, 66°25'O) durante las temporadas 1992 y 1993. La frecuencia de visita a las colonias fue distinta entre años. Durante las temporadas 1991 en Isla Isabel y 1992 en Isla Galiano, se efectuaron visitas desde el mes de octubre con una frecuencia variable (1–7 días), aunque durante las épocas de máxima ocurrencia de posturas y eclosión las visitas tuvieron frecuencias diarias. Para la temporada 1993 en Isla Galiano, en cambio, la frecuencia de visita fue semanal.
El número de nidos activos se estimó quincenalmente entre fines de octubre y fines de enero, considerando como nido activo a todo aquel en el que se observó la presencia de material de nidificación fresco, al menos un individuo de la pareja, huevos o pichones. En las colonias de Cormorán Cuello Negro y en la de Cormorán Imperial de Isla Isabel, el número de nidos se estimó mediante conteo directo. En la colonia de Cormorán Imperial de Isla Galiano se estimó utilizando el método de parcelas (Frere y Gandini 1996). Para la estimación del número de parejas que se reprodujeron en cada año se utilizaron los censos efectuados durante la máxima ocurrencia de posturas.
Durante las tres temporadas y en ambas colonias estudiadas se obtuvo información sobre los patrones de asentamiento, las fechas de puesta y eclosión y el número de huevos por nido. Durante las temporadas 1991 en Isla Isabel y 1992 en Isla Galiano, se obtuvo, además, información sobre el tamaño y el peso de los huevos, el tiempo de incubación, el número de huevos eclosionados por nido y el número de pichones sobrevivientes a la quinta semana de edad (independencia del nido). Para la comparación de las medidas morfométricas y el peso a los 40 días de edad entre especies, los datos fueron obtenidos en ambas islas durante 1991.
Los huevos se marcaron con marcadores indelebles con un número indicativo del orden de puesta para aquellos casos en los cuales fue posible determinarlo. El peso de los huevos se obtuvo con balanza a resorte (precisión: 1 g), durante la semana en que fueron puestos. Las medidas se tomaron con Vernier (precisión: 0.1 mm). El volumen de los huevos, expresado en cm3, se calculó según la fórmula V = 0.0051 × longitud × ancho2 (Hoyt 1979).
Durante 1991 y 1992 se tomaron medidas del peso en una muestra de pichones dentro de las 48 hs de nacidos (identificados sobre la base del tamaño y la presencia de restos del huevo), con balanza a resorte (precisión: 1 g). Cada pichón fue identificado de acuerdo al orden de eclosión mediante la colocación de una cinta adhesiva de diferente color adherida al tarso. Durante 1992 en Isla Galiano, se obtuvo información semanal sobre el peso y tamaño de los primeros pichones nacidos en una muestra de 44 nidos, con balanza a resorte (precisión: 10 g) y Vernier (precisión: 0.1 mm). Las curvas de crecimiento promedio de pichones se ajustaron a las distribuciones teóricas de Gompertz y logística (Ricklefs 1968, 1971) utilizando el método de los cuadrados mínimos (Sokal y Rohlf 1981), considerando luego aquella que presentaba el mejor ajuste. Los ajustes obtenidos para cada ecuación fueron comparados utilizando la fórmula de cociente de varianzas.
Entre fines de febrero y comienzos de marzo de las temporadas 1990 y 1993 se marcaron con anillos metálicos numerados (provistos por el Instituto Miguel Lillo, Tucumán) un total de 456 adultos y 1171 volantones de Cormorán Imperial en las islas Isabel y Galiano. Únicamente 10 de estos adultos fueron capturados en sus nidos; el resto fue capturado en la periferia de las colonias. Durante la temporada no reproductiva de 1992 se efectuaron visitas mensuales para cuantificar el número de individuos presentes en las áreas de estudio. Los conteos se realizaron en horas cercanas al mediodía.
Resultados
Tamaño de colonias, asentamiento y nidificación
El tamaño de la colonia de Cormorán Imperial de Isla Isabel durante 1991 se estimó en 300 parejas reproductivas, mientras que el de la colonia de Isla Galiano se estimó en 900 y 1300 parejas durante 1992 y 1993, respectivamente. El tamaño de la colonia de Cormorán Cuello Negro de Isla Isabel se estimó en 22 parejas durante 1991, mientras que el de la Isla Galiano se estimó en 99 y 180 parejas reproductivas en 1992 y 1993, respectivamente. Durante el invierno del año 1992, por acción de un fuerte mar de fondo, el agua cubrió por completo a la Isla Isabel y afectó profundamente su topografía. La acción del mar produjo una gran deposición de rodados sobre la colonia de Cormorán Imperial. Durante la temporada 1992, 28 individuos de esta especie comenzaron a construir sus nidos en la zona ocupada durante las anteriores temporadas, pero abandonaron el intento. Durante dicha temporada solamente anidaron en Isla Isabel 32 parejas de Cormorán Imperial en un área de sustrato rocoso próxima a la ubicación anterior. A partir del año 1993, la colonia de Cormorán Imperial de Isla Galiano aumentó su número de nidos en una cantidad similar a la que tenía la colonia de Isla Isabel hasta el año 1991, desde alrededor de 900 nidos a aproximadamente 1300. Durante 1993 se observó que de 44 Cormoranes Imperiales anillados que estaban anidando en Isla Galiano, 8 (18.2%) habían sido anillados como reproductores en años previos en la colonia de Isla Isabel. Estas observaciones sugieren que parte de los individuos que formaban la colonia de Isla Isabel hasta la temporada 1991 se desplazaron a la Isla Galiano.
Durante 1992, los Cormoranes Imperiales comenzaron a arribar a las colonias de islas Isabel y Galiano durante la primera semana de septiembre. La mayor parte de los individuos comenzaron a construir nidos durante la primera semana de octubre. Algunos individuos de Cormorán Cuello Negro fueron observados llevando material para el nido a partir de julio, aunque la mayoría de los nidos comenzó a ser construida entre la cuarta semana de septiembre y la primera de octubre.
Puesta e incubación
Las fechas en que fueron observados los primeros huevos variaron en los tres años estudiados en más de un mes (Tabla 1). Los primeros huevos del Cormorán Imperial fueron observados un mes antes en 1993 que en las dos temporadas anteriores. El período de puesta para ambas especies se extendió en algunas temporadas por más de dos meses a partir de la fecha señalada como de inicio de la misma. Sin embargo, durante la temporada 1993 en las colonias de Isla Galiano, el 87% y el 86% de las parejas de Cormorán Imperial (n = 33) y Cormorán Cuello Negro (n = 15), respectivamente, completaron su nidada en las cuatro semanas siguientes al inicio de la puesta. El número de nidos activos para ambas especies durante las temporadas reproductivas de 1991 y 1993 alcanzó valores máximos durante la quincena posterior al inicio de las puestas (Fig. 1).
Tabla 1. Fechas de observación de los primeros huevos de Cormorán Imperial y Cormorán Cuello Negro en las colonias de Isla Isabel e Isla Galiano, Golfo San Jorge, entre 1991 y 1993.
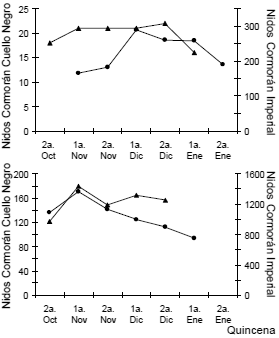
Figura 1. Variación del número de nidos activos de Cormorán Imperial (círculos) y Cormorán Cuello Negro (triángulos) a lo largo de la temporada reproductiva en las colonias de Isla Isabel durante 1991 (arriba) y de Isla Galiano durante 1993 (abajo).
Para el Cormorán Imperial, el intervalo transcurrido entre la puesta del primer y el segundo huevo, en nidadas de dos huevos, se estimó en 2.3 ± 0.6 días (n = 25). Por su parte, para las nidadas de tres huevos, los intervalos transcurridos entre la puesta del primer y el segundo huevo y entre la puesta del segundo y el tercero se estimaron en 2.8 ± 1.2 días (n = 35) y 3.5 ± 1.1 días (n = 15), respectivamente. El tamaño de muestra para el Cormorán Cuello Negro no fue adecuado como para efectuar este tipo de análisis.
El número de huevos por nido al mes de iniciada la puesta para el Cormorán Imperial fue distinto entre años (ANOVA, F = 3.2, P = 0.044, gl = 2; Tabla 2). No obstante, la Prueba de Rangos Múltiples indicó que la única diferencia significativa se observó entre los años 1991 y 1993. Del mismo modo, para el Cormorán Cuello Negro el número de huevos por nido al mes de iniciada la puesta presentó diferencias significativas entre años (ANOVA, F = 20.9, P < 0.0001, gl = 1; Tabla 2). Por su parte, el número de huevos por nido fue similar entre especies durante 1993 pero significativamente diferente durante 1991 (Tabla 2).
Tabla 2. Número de huevos por nido al mes de iniciada la puesta para Cormorán Imperial y Cormorán Cuello Negro en las colonias de Isla Isabel (1991) e Isla Galiano (1992 y 1993), Golfo San Jorge. Los datos se expresan como promedios (± DE), con el tamaño de muestra entre paréntesis. Se muestran los valores del estadístico H de Kruskal-Wallis y su nivel de significación. sd: sin datos.

Los huevos examinados tanto para el Cormorán Imperial como para el Cormorán Cuello Negro fueron de forma elipsoidal, de color azul-verdoso pálido, cubiertos con una capa calcárea blanca. Las dimensiones de los huevos fueron significativamente mayores para el Cormorán Imperial (Tabla 3), observándose la presencia en ambas especies de una baja proporción (menor al 0.3%) de huevos anormalmente pequeños. El peso de los huevos de las dos especies estuvo relacionado positivamente y en forma altamente significativa con el volumen (Cormorán Imperial: y = 0.82514x + 9.6232, n = 113, r = 0.89, P < 0.001; Cormorán Cuello Negro: y = 1.012x + 1.0174, n = 53, r = 0.83, P < 0.001).
Tabla 3. Longitud, ancho, volumen y peso de huevos de Cormorán Imperial y Cormorán Cuello Negro en las colonias de Isla Isabel (1991) e Isla Galiano (1992), Golfo San Jorge. Los datos se expresan como promedios (± DE), con el tamaño de muestra entre paréntesis.

El tamaño de los huevos de Cormorán Imperial en las nidadas de tres huevos dependió del orden de puesta. Para dos de las temporadas de cría estudiadas se observó que el volumen del tercer huevo fue significativamente menor comparado con los volúmenes de los primeros dos huevos (Tabla 4). En cambio, en las nidadas de dos huevos el volumen de ambos huevos fue similar (Tabla 4). Para el Cormorán Cuello Negro de Isla Isabel, el volumen promedio del primer (40.2 ± 4.3 cm3, n = 6) y del segundo huevo (40.1 ± 4.9 cm3, n = 6), en las nidadas de dos huevos, fueron similares (Prueba de Wilcoxon, Z = -0.41, P > 0.05).
Tabla 4. Volumen del primer, segundo y tercer huevo de Cormorán Imperial (con secuencia de puesta conocida) en nidadas de uno, dos y tres huevos en las colonias de Isla Isabel (1991) e Isla Galiano (1992), Golfo San Jorge. Los datos se expresan como promedios (± DE). Se muestran los valores de la comparación entre los volúmenes de los huevos (Prueba de t pareada). *: P < 0.01.

Éxito de eclosión y supervivencia de pichones
El período de incubación se estimó en 27.9 ± 2.4 días (n = 11 nidos) para el Cormorán Imperial. No fue posible estimar el período de incubación para el Cormorán Cuello Negro debido al bajo tamaño de muestra. Los huevos de los cormoranes Imperial y Cuello Negro eclosionaron mayoritariamente en forma asincrónica.
El éxito de eclosión para el Cormorán Imperial fue de 38.2% y de 52.5% en las temporadas 1991 y 1992, respectivamente (Tabla 5). Estas diferencias no resultaron estadísticamente significativas (X2 = 2.2, gl = 1, P > 0.05). Para el Cormorán Imperial, la proporción de pichones vivos al mes de nacidos respecto del número de pichones nacidos (n = 12) durante la temporada 1991 fue de 66.7%.
Tabla 5. Número de huevos puestos y huevos eclosionados de Cormorán Imperial en las colonias de Isla Isabel (1991) e Isla Galiano (1992), Golfo San Jorge.

Las curvas de crecimiento promedio elaboradas en base a medidas morfométricas y al peso de pichones de Cormorán Imperial mostraron un buen ajuste a la ecuación de Gompertz (tarso: r2 = 0.99, n = 30, F3,4 = 2.671, P < 0.01; culmen: r2 = 0.98, n = 60, F3,5 = 1.278, P < 0.01; peso: r2 = 0.99, n = 60, F3,5 = 4.691, P < 0.01). El incremento de cada variable estuvo definido por: tarso = 69.7e-1.87e^(-0.139T), culmen = 49.9e-1.95e^(-0.091T), y peso = 2.2e-4.41e^(-0.082T); donde T = edad (en días).
Tanto el incremento de peso como el crecimiento del culmen registraron valores máximos entre la segunda y la tercera semanas de vida, alcanzando valores próximos al asintótico a los 60 días. El crecimiento del tarso alcanzó valores máximos en la primera semana y valores asintóticos a los 30 días (Fig. 2). Considerando que el peso promedio de machos y hembras adultos en Bahía Bustamante a comienzos de la temporada de cría es de aproximadamente 2.2 kg (G Punta, datos no publicados), los pichones alcanzaron al mes de vida un peso promedio equivalente al 64% del peso de los adultos. A los aproximadamente 40 días de edad, los pichones de Cormorán Imperial fueron significativamente mayores que los de Cormorán Cuello Negro (Tabla 6).

Figura 2. Curvas de crecimiento de los pichones de Cormorán Imperial en las colonias de Isla Isabel e Isla Galiano, Golfo San Jorge. Para cada curva se indica la constante de crecimiento (K) y la asíntota (A) de la ecuación de Gompertz.
Tabla 6. Longitud del tarso, del culmen y peso de pichones (de aproximadamente 40 días de edad) de Cormorán Imperial y Cormorán Cuello Negro en las colonias de Isla Isabel (1991) e Isla Galiano (1991), Golfo San Jorge. Los datos se expresan como promedios (± DE).

El número de pichones sobrevivientes a la independencia por nido resultó similar entre años para el Cormorán Imperial (X2 = 4.5, gl = 2, P > 0.05; Tabla 7). Durante 1993, el número de pichones sobrevivientes a la independencia por nido fue similar para ambas especies (X2 = 1.4, gl = 1, P > 0.05; Tabla 7).
Tabla 7. Número de huevos puestos, número de pichones y número promedio de pichones por nido criados hasta la independencia del nido en las colonias de Cormorán Imperial y Cormorán Cuello Negro de Isla Isabel (1991) y de Isla Galiano (1993), Golfo San Jorge.

Finalización del ciclo reproductivo y dispersión
Los pichones de Cormorán Imperial comenzaron a abandonar progresivamente sus colonias luego de haber completado la muda juvenil (aproximadamente a los 3 meses de edad), aunque la mayor parte de los individuos lo hicieron entre la última semana de marzo y la primera de abril. A partir de estas fechas, la mayor parte de la población de Cormorán Imperial parece dispersarse, alejándose de las zonas de reproducción. Durante las visitas efectuadas a las zonas de cría fuera de la temporada reproductiva, se observó la presencia de un número relativamente bajo de Cormoranes Imperiales. Además, un total de 21 individuos anillados en las colonias de las islas Isabel y Galiano fueron recuperados distantes en hasta 600 km durante otoño e invierno en localidades al norte de la provincia.
Los pichones de Cormorán Cuello Negro comenzaron a frecuentar el mar a fines de marzo. A diferencia de lo observado para el Cormorán Imperial, el número de individuos adultos de Cormorán Cuello Negro se mantuvo relativamente constante a lo largo de todo el año. Durante las visitas a las zonas de cría en temporada invernal se observó un gran número de Cormoranes Cuello Negro en las islas y sus inmediaciones. En cuatro visitas mensuales efectuadas entre mayo y agosto de 1992 a Isla Galiano, se observó en la isla el 40–50% del total de aves presentes en la colonia durante la temporada anterior. Se desconoce el número de individuos que se encontraban alimentándose en aguas adyacentes.
Discusión
La cronología de los eventos del ciclo reproductivo del Cormorán Imperial en Bahía Bustamante varió en más de un mes durante los años estudiados, abarcando el rango de fechas observado para otras colonias de Chubut como Punta Tombo (Malacalza 1984) y Punta León (Yorio et al. 1994, Malacalza y Navas 1996), para Isla Deseada, Santa Cruz (Arrighi y Navarro 1998) y para las Islas Malvinas (Woods y Woods 1997). Las fechas de inicio de puesta observadas, sin embargo, fueron anteriores a las registradas en la colonia ubicada sobre el muelle abandonado de Comodoro Rivadavia, en donde la reproducción comienza a fines de noviembre (Libenson 1997). El Cormorán Cuello Negro en Bahía Bustamante, por su parte, mostró una mayor variación entre años en la época de puesta que el Cormorán Imperial. Al igual que lo observado para el Cormorán Imperial, las fechas de inicio de puesta en la zona de estudio abarcaron el rango de fechas observado en otras colonias de la provincia de Chubut (Malacalza 1995, Libenson 1997) y en las Islas Malvinas (Woods y Woods 1997).
En dos de las temporadas los Cormoranes Imperiales se reprodujeron más de un mes antes que los Cormoranes Cuello Negro. También han sido registradas diferencias en la cronología del ciclo reproductivo en otras localidades del norte de la Patagonia (Malacalza 1995, Libenson 1997), en Punta Tombo (G Punta, datos no publicados) y en otras colonias del norte del Golfo San Jorge (G Punta, datos no publicados). Las diferencias observadas en las fechas de inicio de las puestas entre los cormoranes Imperial y Cuello Negro durante los ciclos reproductivos estudiados podrían atribuirse a diferencias en su ecología alimentaria, como ha sido sugerido para otras aves marinas (Lack 1971, Croxall y Prince 1980, Schreiber y Burger 2002). En la zona de Bahía Bustamante ambas especies se alimentan en gran proporción de peces bentónicos, aunque el Cormorán Imperial muestra un rango de alimentación más amplio y una mayor utilización de cardúmenes de presas pelágicas (Punta et al. 1993, Punta et al., en prensa). Estas diferencias en la ecología alimentaria podrían permitirle al Cormorán Imperial alcanzar con anterioridad una condición física adecuada para comenzar la reproducción.
Para el Cormorán Imperial, el número de huevos por nido al mes de iniciada la puesta fue menor durante 1991. Esta diferencia pudo deberse a los disturbios ocasionados durante ese año por ataques de Petreles Gigantes Comunes (Macronectes giganteus) sobre Cormoranes Imperiales adultos (Punta y Herrera 1995). El sobrevuelo de los petreles ocasionó el abandono de nidos en repetidas oportunidades, resultando en la depredación de huevos por Gaviotas Cocineras (Larus dominicanus), Gaviotas Grises (Leucophaeus scoresbii) y Palomas antárticas (Chionis alba) (Punta y Herrera 1995). El número de huevos por nido al mes de iniciada la puesta fue también menor en 1991 que en 1993 para el Cormorán Cuello Negro. Aunque durante esa temporada no se observaron intentos de depredación de Petreles Gigantes Comunes sobre esta especie, la proximidad de las colonias de Cormorán Imperial y Cormorán Cuello Negro sugiere que pueden haber sido también afectados por esos disturbios. Cabe considerar que las diferencias observadas en el número de huevos entre temporadas pueden también ser el resultado de cambios en la disponibilidad de alimento, aunque se carece de la información necesaria para evaluar esta hipótesis.
En las colonias de Bahía Bustamante, gran parte de los individuos de Cormorán Cuello Negro permanecieron en la zona durante el invierno, mientras que la mayor parte de los Cormoranes Imperiales se dispersaron fuera del área de cría. La información obtenida sobre dispersión de Cormoranes Imperiales adultos y juveniles en este estudio sugiere un movimiento post-reproductivo hacia el norte de sus localidades de cría. Aunque se desconoce su procedencia, individuos de esta especie han sido anteriormente reportados a lo largo de la costa atlántica en la provincia de Buenos Aires y en Uruguay durante la temporada de otoño-invierno (Murphy 1936, Meyer de Schauensee 1966, Escalante 1970, del Hoyo 1992, Rasmussen 1994).
Tanto la Bahía Bustamante como los sectores costeros adyacentes constituyen importantes zonas de extracción de macroalgas, particularmente Gracilaria verrucosa. La explotación guanera también se ha llevado a cabo intermitentemente en algunas de las colonias insulares de Cormorán Imperial (Punta 1996). Recientemente ha comenzado la explotación turística de algunos sitios. El conocimiento de la biología reproductiva de los cormoranes Imperial y Cuello Negro es particularmente relevante al momento de establecer pautas de manejo costero para ordenar el desarrollo de actividades humanas y minimizar el disturbio resultante. Por ejemplo, una de las principales precauciones a tener en cuenta durante la extracción de guano debe ser evitar el disturbio producido por el ingreso de personas a la colonia para desarrollar actividades extractivas durante momentos críticos del ciclo reproductivo (Punta 1996). Las operaciones de extracción de guano deberían efectuarse después de la novena semana de nacidos los pichones y antes del asentamiento de los reproductores en la siguiente temporada. Las variaciones entre años en el patrón temporal de reproducción imposibilitan el establecimiento de fechas de extracción prefijadas, debiéndose determinar año tras año los períodos de actividad extractiva para cada colonia.
Agradecimientos. Agradecemos a la Dirección General de Intereses Marítimos de Chubut y a la Universidad Nacional de la Patagonia San Juan Bosco por el apoyo al presente estudio. A Soriano S.A. y a los Señores Miguel José y Esteban De Felice por su apoyo logístico. Finalmente, a Sandro Sollazzo por la colaboración en los trabajos de campo y a Flavio Quintana por sus comentarios durante la elaboración del manuscrito.
Bibliografía Citada
Arrighi A y Navarro JL (1998) Ecología reproductiva del Cormorán Imperial (Phalacrocorax atriceps), en Isla Deseada (Santa Cruz, Argentina). Hornero 15:64–67 [ Links ]
Boswall J y Prytherch RJ (1972) Some notes on the birds of Point Tombo, Argentina. Bulletin of the British Ornithologists Club 92:118–129 [ Links ]
Croxall JP y Prince PA (1980) Food, feeding ecology and ecological segregation of seabirds at South Georgia. Biological Journal of the Linnean Society 14:103–131 [ Links ]
Escalante R (1970) Aves marinas del Río de la Plata y aguas vecinas del Océano Atlántico. Barreiro y Ramos, Montevideo [ Links ]
Frere E y Gandini P (1996) Conceptos generales para la evaluación y monitoreo de poblaciones de aves marinas. Informes Técnicos del Plan de Manejo Integrado de la Zona Costera Patagónica–Fundación Patagonia Natural 8:1–29 [ Links ]
del Hoyo J, Elliott A y Sargatal J (1992) Handbook of the birds of the world. Volume 1. Ostrich to ducks. Lynx Edicions, Barcelona [ Links ]
Hoyt DF (1979) Practical methods of estimating volume and fresh weight of birds eggs. Auk 96:73–77 [ Links ]
Humphrey PS, Bridge D, Reynolds PW y Peterson RT (1970) Birds of Isla Grande (Tierra del Fuego). Smithsonian Institution, Washington DC [ Links ]
Lack D (1971) Ecological isolation in birds. Blackwell, Oxford [ Links ]
Libenson L (1997) Aspectos de la biología reproductiva de Phalacrocorax magellanicus y P. albiventer en el puerto de Comodoro Rivadavia (Chubut: Argentina). Neotropica 43:73–77 [ Links ]
Malacalza VE (1984) Biología reproductiva de Phalacrocorax albiventer. I. Nidificación en Punta Tombo. Contribución Centro Nacional Patagónico 98:1–13 [ Links ]
Malacalza VE (1995) Aportes al conocimiento de la biología reproductiva de Phalacrocorax magellanicus (Aves: Phalacrocoracidae). Neotropica 41:27–30 [ Links ]
Malacalza VE y Navas JR (1996) Biología y ecología reproductiva de Phalacrocorax albiventer (Aves: Phalacrocoracidae) en Punta León, Chubut, Argentina. Ornitología Neotropical 7:53–61 [ Links ]
Meyer de Schauensee R (1966) The species of birds of South America with their distribution. Livingston, Narberth [ Links ]
Murphy RC (1936) Oceanic birds of South America. Volume 2. Macmillan Company y American Museum of Natural History, Nueva York [ Links ]
de la Peña M (1980) Notas nidológicas sobre biguaes y cormoranes (Aves: Anhingidae y Phalacrocoracidae). Historia Natural 1:109–112 [ Links ]
Punta G (1989) Guaneras de la Provincia del Chubut. Potencialidad productiva y fundamentos para su manejo racional. Dirección de Impresiones Oficiales, Rawson [ Links ]
Punta G (1996) Estado de situación del recurso guanero en la República Argentina. Informes Técnicos del Plan de Manejo Integrado de la Zona Costera Patagónica - Fundación Patagonia Natural 6:1–19 [ Links ]
Punta G y Herrera G (1995) Predation by Southern Giant Petrels Macronectes giganteus on adult Imperial Cormorants Phalacrocorax atriceps. Marine Ornithology 23:166–167 [ Links ]
Punta G y Saravia J (1993) Distribución, abundancia y aspectos de la biología reproductiva del Cormorán Cuello Negro Phalacrocorax magellanicus en la Provincia del Chubut, Argentina. Hornero 13:295–298 [ Links ]
Punta GE, Saravia JRC y Yorio PM (1993) The diet and foraging behaviour of two Patagonian cormorants. Marine Ornithology 21:27–36 [ Links ]
Punta G, Yorio P y Herrera G (en prensa) Temporal patterns in the diet and food partitioning in Imperial Cormorants (Phalacrocorax atriceps) and Rock Shags (P. magellanicus) breeding at Bahía Bustamante, Argentina. Wilson Bulletin [ Links ]
Rasmussen PC (1994) Geographic variation in morphology and allozymes of South American Imperial Shags. Auk 111:143–161 [ Links ]
Ricklefs RE (1968) Patterns of growth in birds. Ibis 110:419–451 [ Links ]
Ricklefs RE (1971) Patterns of growth in birds. II. Growth rate and mode of development. Ibis 115:177–201 [ Links ]
Schreiber EA y Burger J (2002) Biology of marine birds. CRS Press, Boca Ratón [ Links ]
Siegel-Causey D (1986) Behaviour and affinities of the Magellanic Cormorant. Notornis 33:249–257 [ Links ]
Sokal RR y Rohlf FS (1981) Biometry. WH Freeman & Company, San Francisco [ Links ]
Woods W y Woods A (1997) Atlas of breeding birds of the Falkland Islands. Anthony Nelson, Shropshiere [ Links ]
Yorio P, Frere E, Gandini P y Conway W (1999) Status and conservation of seabirds breeding in Argentina. Bird Conservation International 9:299–314 [ Links ]
Yorio P, García Borboroglu P, Bertellotti M, Lizurume ME, Giaccardi M, Punta G, Saravia J, Herrera G, Sollazzo S y Boersma D (1998) Distribución reproductiva y abundancia de las aves marinas de Chubut. Parte II: Norte del Golfo San Jorge, de Cabo Dos Bahías a Comodoro Rivadavia. Pp. 76–117 en: Yorio P, Frere E, Gandini P y Harris G (eds) Atlas de la distribución reproductiva de aves marinas en el litoral patagónico argentino. Fundación Patagonia Natural y Wildlife Conservation Society, Buenos Aires [ Links ]
Yorio P, Quintana F, Campagna C y Harris G (1994) Diversidad, abundancia y dinámica espacio-temporal de la colonia mixta de aves marinas de Punta León, Patagonia. Ornitología Neotropical 5:69–77 [ Links ]

