










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEl hornero
versión impresa ISSN 0073-3407versión On-line ISSN 1850-4884
Hornero v.19 n.1 Buenos Aires ene./ago. 2004
Comunidades de aves en un gradiente urbano de la ciudad de Mar del Plata, Argentina
Lucas M. Leveau 1,2 y Carlos M. Leveau 1
1 Alte. Brown 2420 1°A, 7600 Mar del Plata, Buenos Aires, Argentina.
2 lucasleveau@yahoo.com.ar
Resumen
El proceso de urbanización crece continuamente sobre las áreas nativas creando nuevos hábitats y situaciones para las aves. En este estudio se analiza la abundancia y distribución de aves presentes en un gradiente urbano de la ciudad de Mar del Plata durante la época reproductiva. Las aves se registraron en transectas en el centro comercial y en áreas suburbanas y periurbanas residenciales. La proporción de árboles y arbustos y de césped aumentó hacia las áreas menos urbanizadas. La similitud fue más alta entre las comunidades de aves de sitios suburbanos y periurbanos. La riqueza, la diversidad y la abundancia de aves aumentaron en las áreas suburbanas y estuvieron significativamente correlacionadas con la proporción de árboles y arbustos y de césped. La mayoría de los gremios siguieron la misma tendencia, excepto el de los omnívoros, los cuales fueron similarmente abundantes a lo largo del gradiente urbano. De las 15 especies que pudieron ser analizadas estadísticamente, aves exóticas como Columba livia y Passer domesticus fueron más abundantes en los sitios más urbanizados. Otras 8 especies fueron igualmente abundantes en áreas suburbanas y periurbanas. Milvago chimango, Turdus rufiventris y Agelaioides badius fueron más abundantes en los sectores periurbanos.
Palabras clave: Abundancia; Argentina; Aves; Gradiente urbano; Gremios; Mar del Plata; Riqueza.
Abstract
Bird communities along an urban gradient in Mar del Plata City, Argentina
The urbanization process grows continuously over pristine areas creating new habitats and situations for birds. In this study we analize the abundance and distribution of bird species along an urban gradient in Mar del Plata City during the breeding season. We recorded birds in transects in downtown, suburban residential areas and periurban residential areas. The proportion of trees and shrubs and of lawn increased in less urbanized areas. Richness, diversity and abundance of birds were higher at suburban and periurban sites, and were significantly correlated with the proportion of trees and shrubs, and of lawn. Most guilds followed the same trend, except omnivorous birds, which were similarly abundant along the urban gradient. Similarity was higher between bird communities in suburban and periurban sites. Of the 15 species that were statistically analized, exotic species like Columba livia and Passer domesticus were more abundant in the most urbanized areas. Other 8 species were equally abundant in suburban and periurban areas. Milvago chimango, Turdus rufiventris and Agelaioides badius were more abundant in the periurban areas.
Key words: Abundance; Argentina; Birds; Guilds; Mar del Plata; Richness; Urban gradient.
Recibido 22 enero 2004, aceptado 27 agosto 2004
Para el año 2025 se prevé que la población urbana de los países en vías de desarrollo, incluidos los países del Neotrópico, llegará a los cuatro billones de habitantes (tres veces más que la población urbana esperada para los países desarrollados), lo que provocará un gran impacto sobre los ecosistemas naturales y seminaturales (Marzluff et al. 2001). El conocimiento de los efectos de la urbanización sobre los ecosistemas podría ser una herramienta útil a escala local que permitiría a planificadores urbanos tener un papel decisivo en el mantenimiento o mejoramiento de la biodiversidad. A su vez, la percepción de los ciudadanos hacia la naturaleza urbana puede influenciar significativamente el manejo de la biodiversidad a una escala regional, nacional y aún global (ver referencias en Clergeau et al. 2001). Una de las maneras efectivas para estudiar los efectos de la urbanización sobre la biodiversidad es a través de un gradiente de urbanización (McDonnell y Pickett 1990). Otra aproximación ha sido la aplicación de la teoría ecológica de islas (ver MacArthur y Wilson 1967), principalmente en parques urbanos que difieren en tamaño o edad (Suhonen y Jokimäki 1988, Jokimäki 1999, Fernández-Juricic 2000).
En el Hemisferio Norte las aves han sido utilizadas para el estudio del gradiente urbano-rural, especialmente a partir de la década de 1990 (Blair 1996, 2001, Clergeau et al. 1998, Germaine et al. 1998; ver una revisión más completa en Clergeau et al. 2001 y en Marzluff et al. 2001). Por el contrario, en la Región Neotropical el conocimiento de los efectos de la urbanización sobre las comunidades de aves es pobre. Se han realizado algunos estudios en parches forestados y parques, principalmente en Brasil (Matarazzo-Neuberger 1992, 1995, Mendonça-Lima y Fontana 2000) y Argentina (Feninger 1983), pero solo en dos casos se estudiaron las aves a través de un gradiente urbano-rural (Ruszczyk et al. 1987, en Brasil; Reynaud y Thioulouse 2000, en la Guyana Francesa).
El objetivo de este estudio es analizar la respuesta de las especies de aves a la urbanización y su distribución a lo largo del gradiente urbano durante la época reproductiva, a través de parámetros como abundancia, riqueza, diversidad de especies y densidad de gremios. De acuerdo a los estudios realizados anteriormente, se esperan los siguientes patrones: (1) la riqueza y la diversidad de especies serán afectadas negativamente por el aumento de la urbanización (Huhtalo y Järvinen 1977, Blair 1996, 2001, Clergeau et al. 1998, Reynaud y Thioulouse 2000), (2) la abundancia de aves aumentará hacia las áreas menos urbanizadas (Huhtalo y Järvinen 1977, Blair 1996, Reynaud y Thioulouse 2000), y (3) el gremio de los omnívoros será más abundante en las áreas más urbanizadas, mientras que los otros gremios serán afectados negativamente por la urbanización (Clergeau et al. 1998).
Métodos
El estudio se llevó a cabo en la ciudad de Mar del Plata (38°00'N, 57°34'O), ubicada en el sudeste de la provincia de Buenos Aires, Argentina. La ciudad, con 562901 habitantes (censo 2001), recibió la llegada de 649058 turistas durante la temporada estival (diciembre-marzo) 2002-2003 (estadísticas de la Municipalidad de la ciudad de Mar del Plata). Posee una temperatura promedio anual de 14 °C y las precipitaciones alcanzan los 920 mm anuales. Los sitios de estudio fueron dos barrios forestados periurbanos (ubicados en la periferia de la ciudad): el Grosellar (54 ha; siete transectas) y el Bosque Patricio Peralta Ramos (216 ha; siete transectas); tres barrios forestados suburbanos (ubicados dentro de la matriz urbana): Los Troncos (169 ha; siete transectas), Pinos de Anchorena (18 ha; cuatro transectas) y Parque Luro (32 ha; cinco transectas); y el centro urbano o comercial (200 ha; 15 transectas). La mayoría de las especies de plantas de los sitios relevados son exóticas y ornamentales.
Desde octubre de 2002 hasta febrero de 2003 se recorrieron, en cada sitio, transectas fijas de 100 m de largo y de 50 m de ancho (0.5 ha), separadas por 200 m, durante las mañanas de domingo, entre las 06:00-09:00 h. Cada transecta fue recorrida dos veces y se calcularon los valores promedio de las dos recorridas de cada transecta, evitando de este modo la pseudoreplicación de los datos (n = 45). Se registraron todas las especies de aves vistas o escuchadas dentro del área de la transecta, excepto aquellas que volaban alto sin actividad de caza, como Milvago chimango o las golondrinas.
La estructura del hábitat, que comprende la cantidad de área cubierta por edificios, pavimento, césped, árboles y arbustos, fue estimada visualmente en las mismas transectas donde se relevaron las aves, mediante parcelas de 20 m de radio.
Los gremios fueron determinados sobre la base de datos de la literatura (Olrog 1959, de la Peña 1988, 1989) y de observaciones de campo. Cada especie fue clasificada usando dos componentes: su principal alimento (carnívora, frugívora, granívora, insectívora, nectarívora u omnívora) y el lugar donde busca el alimento (aérea, arborícola o terrícola). En el caso de Elaenia parvirostris, en estudios previos se ha registrado un alto grado de frugivoría (Marini y Cavalcanti 1998), pero en nuestra área de estudio la hemos visto alimentarse principalmente de insectos.
Para el análisis estadístico por especie se incluyeron aquellas que estuvieron presentes en más del 10% de las transectas. Se calculó la prueba no paramétrica de Spearman para relacionar las variables de estructura del hábitat con la densidad de gremios, abundancia (individuos/0.5 ha), riqueza (número de especies por conteo) y diversidad de especies. Luego se realizaron correcciones de los niveles de p mediante el Método de Bonferroni por inferencias simultáneas (Rice 1989). La diversidad de especies fue calculada para cada conteo con el Índice de Shannon-Wiener (Krebs 1995), H = -åpi log pi , donde pi es la proporción del total de la muestra correspondiente a la especie i. Las diferencias en las variables a lo largo del gradiente urbano fueron analizadas con la prueba no paramétrica de Kruskal-Wallis y análisis a posteriori de Tukey (Zar 1999). Para determinar el grado de similitud entre las comunidades de aves a lo largo del gradiente se usó el Índice de Sörensen (Clergeau et al. 2001), que se basa sólo en el número de especies registradas, S = 2 c / (a + b), donde c es el número de especies compartidas por las dos áreas, y a y b el número total de especies para cada área. Los valores de este índice varían entre 0 (comunidades completamente diferentes) y 1 (idénticas).
Resultados
Se registraron en total 31 especies: 7 en el sector urbano, 25 en el sector suburbano y 23 en el sector periurbano (Tabla 1). Tres especies fueron exóticas: Columba livia, Passer domesticus y Carduelis chloris. Los valores de similitud fueron bajos entre las comunidades del sector urbano y del sector suburbano y periurbano (0.35 y 0.34, respectivamente), mientras que se registró un alto grado de similitud (0.82) entre las dos últimas.
Tabla 1. Densidad promedio (± DE) de aves en tres tipos de barrio a lo largo del gradiente urbano de la ciudad de Mar del Plata. La densidad está expresada en número de aves/0.5 ha. Se muestran también el gremio al que pertenece cada especie y las diferencias estadísticas entre tipos de barrio para las especies observadas en más del 10% de los conteos. *: P < 0.05, **: P < 0.01, ***: P < 0.001. Diferentes letras indican diferencias significativas entre tipos de barrio (prueba no paramétrica de Tukey, P < 0.05).
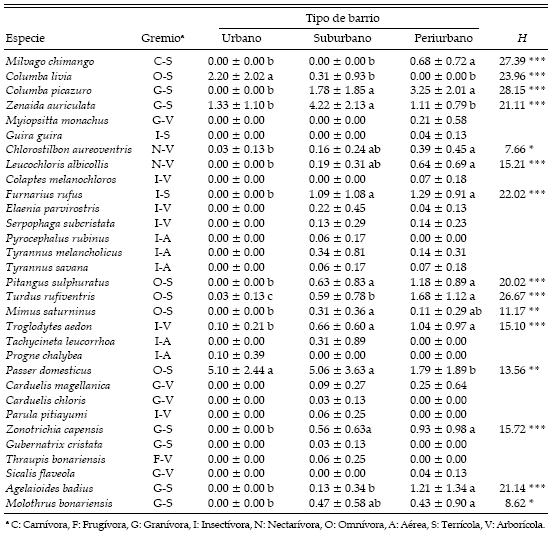
La cobertura de árboles, arbustos y césped siguió el mismo patrón en el gradiente urbano, aumentando desde el sector urbano hacia los sectores menos urbanizados (H = 33.63, P < 0.001 para árboles y arbustos; H = 35.28, P < 0.001 para césped; Fig. 1). Lo opuesto sucedió con la proporción de edificios a lo largo del gradiente, aumentando hacia los sectores más urbanizados (H = 35.82, P < 0.001; Fig. 1). El porcentaje cubierto por asfalto fue similar entre el sector urbano y suburbano, pero disminuyó marcadamente en el sector periurbano (H = 36.53, P < 0.001; Fig. 1).
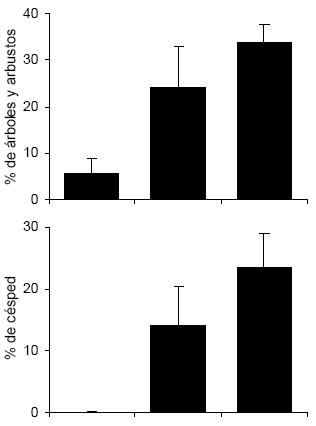
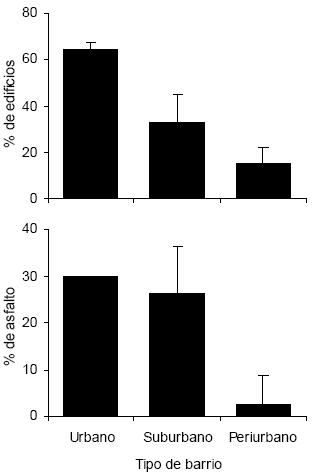
Figura 1. Estructura del hábitat en tres tipos de barrio a lo largo del gradiente urbano de la ciudad de Mar del Plata. Los valores son promedios + DE.
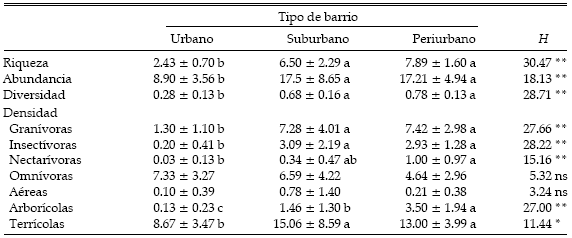
La riqueza, la abundancia y la diversidad de especies fueron menores en el sector urbano que en el sector suburbano y periurbano, sin diferencia entre estos dos últimos (Tabla 2). En cuanto a los gremios, la densidad de los omnívoros fue similar a lo largo del gradiente urbano, mientras que la densidad de las especies nectarívoras fue significativamente mayor en los sectores periurbanos que en el sector urbano (Tabla 2). Los granívoros y los insectívoros tuvieron niveles de abundancia similares en los sectores periurbanos y suburbanos, y menores en el sector más urbanizado (Tabla 2). El gremio de los carnívoros, representado solo por Milvago chimango, fue registrado únicamente en el sector periurbano. Las especies que se alimentan en el aire no fueron afectadas por el nivel de urbanización, mientras que las terrícolas fueron más abundantes en las áreas suburbanas y periurbanas (Tabla 2). Las especies que se alimentan en la vegetación fueron significativamente más abundantes en el sector periurbano, disminuyendo hacia los sectores más urbanizados (Tabla 2).
Tabla 2. Riqueza, abundancia (individuos/0.5 ha), diversidad y densidad de los gremios de aves en tres tipos de barrio a lo largo del gradiente urbano de la ciudad de Mar del Plata. Los valores son promedios ± DE. Se muestran también las diferencias estadísticas entre tipos de barrio. *: P < 0.01, **: P < 0.001, ns: no significativo. Diferentes letras indican diferencias significativas entre tipos de barrio (prueba no paramétrica de Tukey, P < 0.05).
De las 15 especies incluidas en los análisis estadísticos, 2 fueron más abundantes en el sector urbano: Columba livia (no observada en el sector periurbano) y Passer domesticus, aunque este último con niveles de abundancia similares en el sector suburbano (Tabla 1). Otras 8 especies (Columba picazuro, Chlorostilbon aureoventris, Leucochloris albicollis, Furnarius rufus, Pitangus sulphuratus, Troglodytes aedon, Zonotrichia capensis y Molothrus bonariensis) tuvieron abundancias similares en los sectores suburbanos y periurbanos, mientras que Milvago chimango, Turdus rufiventris y Agelaioides badius fueron más abundantes en los sectores periurbanos. Zenaida auriculata y Mimus saturninus fueron más abundantes en los sectores suburbanos.
La riqueza, la abundancia y la diversidad de especies se correlacionaron positivamente con la proporción de árboles y arbustos y de césped, y negativamente con la proporción de edificios y de asfalto (Tabla 3). La abundancia de granívoros, insectívoros, nectarívoros y carnívoros siguieron la misma tendencia. La abundancia de omnívoros no se correlacionó con ninguna variable. Las especies arborícolas se correlacionaron positivamente con las variables de vegetación y negativamente con la proporción de edificios y asfalto, mientras que la abundancia de las especies que buscan su alimento en el aire no se correlacionó con ninguna variable ambiental. Las especies terrícolas siguieron un patrón similar a las arborícolas, aunque su abundancia no se correlacionó con la proporción de asfalto.
Tabla 3. Valores del Coeficiente de Correlación de Spearman (rs) entre la riqueza, abundancia, diversidad y densidad de los gremios de aves y la cobertura de árboles y arbustos, de césped, de edificios y de asfalto a lo largo del gradiente urbano de la ciudad de Mar del Plata. *: P < 0.01, **: P < 0.001, ns: no significativo.

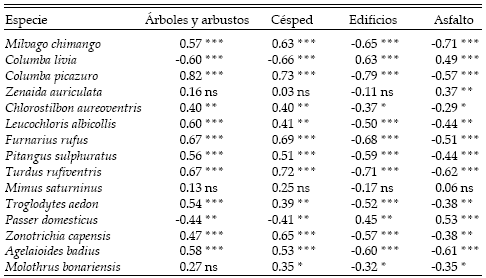
Las abundancias de Passer domesticus y Columba livia se correlacionaron positivamente con la proporción de edificios y de asfalto, y negativamente con la de césped y de árboles y arbustos (Tabla 4). Otras 10 especies se correlacionaron positivamente con la proporción de cubierta arbórea y de césped, y negativamente con la de edificios y de asfalto (Tabla 4). La abundancia de Zenaida auriculata solo se correlacionó positivamente con la proporción de asfalto, la de Mimus saturninus no se correlacionó con ninguna de las variables ambientales, mientras que la de Molothrus bonariensis se correlacionó negativamente con la proporción de edificios y de asfalto, y positivamente con la de césped, pero no se correlacionó con la proporción de árboles y arbustos (Tabla 4).
Tabla 4. Valores del Coeficiente de Correlación de Spearman (rs) entre la densidad de las especies de aves observadas en más del 10% de los conteos y la cobertura de árboles y arbustos, de césped, de edificios y de asfalto a lo largo del gradiente urbano de la ciudad de Mar del Plata. *: P < 0.05, **: P < 0.01, ***: P < 0.001, ns: no significativo.
Discusión
Las comunidades de aves fueron muy similares en los sectores suburbanos y periurbanos, pero éstas fueron poco similares a las del centro urbano, resultado similar al de Clergeau et al. (1998, 2001). Esto se debería principalmente a las diferencias estructurales del hábitat (áreas suburbanas y periurbanas con importante cobertura de vegetación y áreas del centro urbano con alta cobertura de edificios).
Tal como se ha reportado en estudios similares, la diversidad y la riqueza de aves disminuyeron marcadamente con el nivel de urbanización (Huhtalo y Järvinen 1977, Blair 1996, Clergeau et al. 1998, Reynaud y Thioulouse 2000). En este estudio, la proporción de árboles y arbustos y la de césped fueron favorables para la mayoría de las especies de aves observadas. Sin embargo, la mayoría de las especies registradas en los sectores suburbanos y periurbanos están ampliamente distribuidas en la región (Narosky e Yzurieta 1987, Narosky y Di Giacomo 1993), colonizando las áreas urbanas. Por lo tanto, es crucial diferenciar entre calidad y cantidad de especies al momento de mantener la diversidad biológica (Blair 1996). Solo se registraron 4 especies (Milvago chimango, Troglodytes aedon, Zonotrichia capensis y Molothrus bonariensis) de las observadas por Isacch y Martínez (2001) en un pastizal de paja colorada (Paspalum quadrifarium), resultando esto en la desaparición de más de 20 especies típicas. De las especies de aves que se pueden registrar en los bosques de tala (Celtis tala) de la zona (Leveau, obs. pers.), Serpophaga subcristata, Elaenia parvirostris y Parula pitiayumi fueron registradas en menos del 10% de nuestros relevamientos, mientras que Polioptila dumicola estuvo ausente. De esta forma, este estudio demuestra que la urbanización puede provocar extinción local e invasión de especies (Blair 1996, 2001).
El sitio de nidificación que utilizan las aves tiene una importancia crucial en su éxito como colonizadores urbanos. Las especies que anidan en árboles están mejor adaptadas a la urbanización, probablemente debido a que los sitios de nidificación están más disponibles por la plantación de árboles ornamentales en las áreas residenciales (Lim y Sodhi 2004). De forma similar, las especies que anidan en cavidades artificiales, como Passer domesticus y ciertas especies de golondrinas, se ven beneficiadas porque usan estructuras humanas para nidificar. Por el contrario, las especies que nidifican en el suelo o en la vegetación herbácea están dramáticamente afectadas por la urbanización (Haire et al. 2000). De las 31 especies registradas en este estudio y de las 80 especies registradas en un estudio en Singapur (Lim y Sodhi 2004), solo 1 especie en cada caso nidificaba en el suelo (en nuestro caso, Zonotrichia capensis). Estos resultados indican que el impacto de la urbanización sobre ambientes de pradera podría provocar la extinción local de la mayoría de las especies que anidan en esos ambientes y la colonización, y consecuente expansión geográfica, de especies nidificantes en árboles.
Al igual que lo registrado en varios trabajos anteriores (Huhtalo y Järvinen 1977, Blair 1996, Clergeau et al. 1998, Reynaud y Thioulouse 2000), la abundancia de aves disminuyó hacia los sectores más urbanizados. Sin embargo, Clergeau et al. (1998) registraron un aumento de la abundancia en las zonas más urbanizadas de la ciudad de Rennes (Francia) e hipotetizaron que éste podría ser el patrón más común en áreas urbanas. Blair (1996) sugirió que la abundancia sería mayor en los niveles intermedios de un gradiente urbano-rural.
A diferencia de los resultados encontrados por Clergeau et al. (1998), las aves omnívoras mantuvieron abundancias similares a lo largo del gradiente urbano, coincidiendo esto con los resultados obtenidos por Lim y Sodhi (2004). Esto se podría deber a que, en nuestro estudio, el gremio de los omnívoros estuvo compuesto tanto por especies exóticas que se alimentan de desperdicios humanos como por especies nativas que no lo hacen. Las aves insectívoras y granívoras se vieron favorecidas por niveles intermedios y bajos de urbanización, coincidiendo con lo encontrado por Clergeau et al. (1998). Reynaud y Thioulouse (2000), en su estudio de la ciudad tropical de Cayenne (Guyana Francesa), distinguieron un total de 11 gremios tróficos, de los cuales los granívoros, 3 clases de insectívoros, los nectarívoros y los omnívoros fueron constantes a lo largo del gradiente urbano, aunque los carnívoros registraron un patrón similar al de este estudio. Esto indica que la determinación de gremios como indicadores biológicos del nivel de urbanización puede variar ampliamente entre ciudades debido al contexto regional. Por ejemplo, Cayenne está rodeada por selvas tropicales, las cuales brindan una mayor cantidad de especies y tipos de recursos, resultando esto en una mayor variedad de gremios en comparación con Mar del Plata, que está rodeada por agroecosistemas de climas templados. En cuanto a las tácticas de caza, las especies que buscan su alimento en el aire no fueron afectadas por el grado de urbanización, mientras que las especies terrícolas fueron igualmente abundantes en los niveles intermedios y bajos de urbanización. Las especies que buscan alimento en la vegetación fueron afectadas por los niveles altos e intermedios de urbanización, aparentemente dependiendo de altos niveles de cobertura vegetal.
Las distintas especies de aves registradas en este estudio respondieron diferencialmente al nivel de urbanización (ver también Blair 1996). Las especies exóticas como Columbia livia y Passer domesticus son capaces de aprovechar los recursos brindados por los humanos en forma directa, siendo comunes en otros centros urbanos del mundo (Huhtalo y Järvinen 1977, Blair 1996, Clergeau et al. 1998, Lim y Sodhi 2004). Si bien Progne chalybea no pudo ser analizada estadísticamente, solo se registró en el centro urbano, donde los edificios altos le proporcionan sitios de nidificación. Esta especie también sería beneficiada por los altos niveles de urbanización, colonizando nuevas áreas. Ocho especies fueron abundantes en áreas suburbanas y periurbanas; éstas se vieron favorecidas principalmente por el aumento en la riqueza estructural del hábitat, debido a la plantación de vegetación ornamental que también aporta sitios de nidificación y perchas. Zenaida auriculata fue más abundante en el sector suburbano y solo se correlacionó con la proporción de asfalto. Como esta especie busca su alimento en el suelo, es muy posible que las calles asfaltadas le proporcionen un sustrato con alta visibilidad del alimento. Mimus saturninus no se correlacionó con ninguna variable ambiental, lo que podría indicar que está adaptada a niveles intermedios de urbanización. Chlorostilbon aureoventris y Leucochloris albicollis parecen ser beneficiadas por las áreas residenciales, debido a la presencia de plantas y árboles ornamentales como el limpiatubos (Callistemon speciosus), los cuales les proporcionan un recurso alimenticio. Leucochloris albicollis también se ve beneficiado por la plantación de árboles exóticos del género Eucalyptus, y en especial Eucalyptus globulus, el cual florece durante otoño-invierno, constituyendo una fuente de alimentación durante ese período y favoreciendo su expansión en la provincia de Buenos Aires (Montaldo 1984). Milvago chimango, Turdus rufiventris y Agelaioides badius fueron más abundantes en lugares con bajo nivel de urbanización, y parecen estar muy asociadas con altos niveles de cobertura vegetal.
De las 15 especies de aves más observadas en la ciudad de Mar del Plata, el 67% está catalogada, a nivel regional, como abundante, y el 27% y el 7% restante como común y escasa, respectivamente (Narosky y Di Giacomo 1993). Esto significa que las áreas urbanas favorecen la colonización por especies ampliamente distribuidas y capaces de explotar nuevos recursos. Además, esto podría indicar que la colonización de áreas urbanas comienza con poblaciones de aves no urbanas relativamente densas (Tomialojc 1998), aunque es muy posible que especies como Columba livia o Passer domesticus colonicen ciudades desde otras áreas urbanas (Erz 1966, Tomialojc 1998).
Independientemente del uso de la tierra, el diseño y conservación de paisajes en un área urbana pueden influenciar fuertemente la presencia de una especie determinada (Hostetler y Knowles-Yanez 2003). Varios estudios enfatizan la importancia de implementar medidas a nivel sitio-específico, mediante la plantación de determinadas especies de árboles y arbustos (Clergeau et al. 1998, 2001, Germaine et al. 1998, Hostetler y Knowles-Yanez 2003). La plantación de árboles nativos no solo puede incrementar la riqueza de aves nativas, sino también beneficiar a las personas elevando el valor de sus propiedades (Mills et al. 1989, Germaine et al. 1998; ver referencias en Fernández-Juricic y Jokimäki 2001). En Mar del Plata, la preservación de parches con pastizales nativos en la periferia (escala regional) beneficiaría la presencia de especies típicas de esos ambientes, y la plantación de árboles autóctonos como el tala o el coronillo (Scutia buxifolia) en los jardines (escala local) podría favorecer la presencia de sus especies de aves acompañantes en estos ambientes urbanos.
Agradecimientos. Agradecemos a Fernando Mapelli por sus sugerencias en una versión anterior y a Marcelo Kittlein por el envío de bibliografía. Los aportes del editor y de dos revisores anónimos fueron muy útiles.
Bibliografía Citada
Blair RB (1996) Land use and avian species diversity along an urban gradient. Ecological Applications 6:506-519 [ Links ]
Blair RB (2001) Creating a homogeneous avifauna. Pp. 459-486 en: Marzluff JM, Bowman R y Donnelly R (eds) Avian ecology and conservation in an urbanizing world. Kluwer Academic, Norwell [ Links ]
Clergeau P, Jokimäki J y Savard JPL (2001) Are urban bird communities influenced by the bird diversity of adjacent landscapes? Journal of Applied Ecology 38:1122-1134 [ Links ]
Clergeau P, Savard JPL, Mennechez G y Falardeau G (1998) Bird abundance and diversity along an urban-rural gradient: a comparative study between two cities on different continents. Condor 100:413-425 [ Links ]
Erz W (1966) Ecological principles in the urbanization of birds. Ostrich (Supplement) 6:357-364 [ Links ]
Feninger O (1983) Estudios cuantitativos sobre aves en áreas urbanas de Buenos Aires con densa población urbana. Hornero Número Extraordinario:174-191 [ Links ]
Fernández-Juricic E (2000) Bird community composition patterns in urban parks of Madrid: the role of age, size, and isolation. Ecological Research 15:373-383 [ Links ]
Fernández-Juricic E y Jokimäki J (2001) A habitat island approach to conserving birds in urban landscapes: case studies from southern and northern Europe. Biodiversity and Conservation 10:2023-2043 [ Links ]
Germaine SS, Rosenstock SS, Schweinsburg RE y Richardson WS (1998) Relationships among breeding birds, habitat, and residential development in Greater Tucson, Arizona. Ecological Applications 8:680-691 [ Links ]
Haire SL, Bock CB, Cade BS y Bennett BC (2000) The role of landscape and habitat characteristics in limiting abundance of grassland nesting songbirds in an urban open space. Landscape and Urban Planning 48:65-82 [ Links ]
Hostetler M y Knowles-Yanez K (2003) Land use, scale, and bird distributions in the Phoenix metropolitan area. Landscape and Urban Planning 62:55-68 [ Links ]
Huhtalo H y Järvinen D (1977) Quantitative composition of the urban bird community in Tornio, Northern Finland. Bird Study 24:179-185 [ Links ]
Isacch JP y Martínez MM (2001) Estacionalidad y relaciones con la estructura del hábitat de la comunidad de aves de pastizales de paja colorada (Paspalum quadrifarium) manejados con fuego en la provincia de Buenos Aires, Argentina. Ornitología Neotropical 12:345-354 [ Links ]
Jokimäki J (1999) Occurrence of breeding bird species in urban parks: effects of park structure and broad-scale variables. Urban Ecosystems 3:21-34 [ Links ]
Krebs CJ (1995) Ecología: estudio de la distribución y la abundancia. Editorial Harla, Mexico DF [ Links ]
Lim HC y Sodhi NS (2004) Responses of avian guilds to urbanization in a tropical city. Landscape and Urban Planning 66:199-215 [ Links ]
MacArthur RH y Wilson EO (1967) The theory of island biogeography. Princeton University Press, Princeton [ Links ]
Marini MÂ y Cavalcanti RB (1998) Frugivory by Elaenia flycatchers. Hornero 15:47-50 [ Links ]
Marzluff JM, Bowman R y Donnelly R (2001) A historical perspective on urban bird research: trends, terms, and approaches. Pp. 1-17 en: Marzluff JM, Bowman R y Donnelly R (eds) Avian ecology and conservation in an urbanizing world. Kluwer Academic, Norwell [ Links ]
Matarazzo-Neuberger WM (1992) Avifauna urbana de dois municipio da Grande Sao Paulo, SP (Brasil). Acta Biológica Paranaense 21:89-106 [ Links ]
Matarazzo-Neuberger WM (1995) Comunidades de aves de cinco parques e praças da Grande Sao Paulo, Estado de Sao Paulo. Ararajuba 3:13-19 [ Links ]
McDonnell MJ y Pickett STA (1990) Ecosystem structure and function along urban-rural gradients: an unexploited opportunity for ecology. Ecology 71:1232-1237 [ Links ]
Mendonça-Lima A y Fontana CS (2000) Composição, freqüencia e aspectos biológicos da avifauna no Porto Alegre Coutry Clube, Rio Grande do Sul. Ararajuba 8:1-8 [ Links ]
Mills GS, Dunning JB y Bates JM (1989) Effects of urbanization on breeding bird community structure in southwestern desert habitats. Condor 91:416-428 [ Links ]
Montaldo NH (1984) Asociación de dos especies de picaflores con árboles del género Eucalyptus (Myrtaceae) en la Provincia de Buenos Aires. Hornero 12:159-162 [ Links ]
Narosky T y Di Giacomo AG (1993) Las aves de la provincia de Buenos Aires: distribución y estatus. Asociación Ornitológica del Plata, Vázquez Mazzini Editores y L.O.L.A, Buenos Aires [ Links ]
Narosky T e Yzurieta D (1987) Guía para la identificación de las aves de Argentina y Uruguay. Asociación Ornitológica del Plata, Buenos Aires [ Links ]
Olrog CC (1995) Las aves argentinas. Una guía de campo. Instituto Miguel Lillo, Tucumán [ Links ]
de la Peña, MR (1988) Guía de aves argentinas. Tomo 5. Dendrocolaptidae a Tyrannidae. L.O.L.A., Buenos Aires [ Links ]
de la Peña, MR (1989) Guía de aves argentinas. Tomo 6. Rhinocryptidae a Corvidae. L.O.L.A., Buenos Aires [ Links ]
Reynaud RA y Thioulouse J (2000) Identification of birds as ecological markers along a neotropical urban-rural gradient (Cayenne, French Guiana), using co-inertia analysis. Journal of Environmental Management 59:121-140 [ Links ]
Rice WR (1989) Analizing tables of statistical tests. Evolution 43:223-225 [ Links ]
Ruszczyk A, Rodrigues JJS, Roberts TMT, Bendati MMA, del Pino RS, Marques JCV y Melo MTQ (1987) Distribution patterns of eight bird species in the urbanization gradient of Porto Alegre, Brazil. Ciência e Cultura 39:14-19 [ Links ]
Suhonen J y Jokimäki J (1988) A biogeographical comparison of the breeding bird species assemblages in twenty Finnish urban parks. Ornis Fennica 65:76-83 [ Links ]
Tomialojc L (1998) Breeding bird densities in some urban and non-urban habitats: the Dijon case. Acta Ornithologica 33:159-171 [ Links ]
Zar JH (1999) Biostatistical analysis. Cuarta edición. Prentice Hall, Upper Saddle River [ Links ]

