










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEl hornero
Print version ISSN 0073-3407On-line version ISSN 1850-4884
Hornero vol.19 no.1 Buenos Aires Jan./Aug. 2004
Caracterización de las asociaciones de alimentación multiespecíficas de aves marinas en la Ría Deseado, Santa Cruz, Argentina
Patricia B. Nasca 1,2, Patricia A. Gandini 1,3 y Esteban Frere 1,3
1 Centro de Investigaciones Puerto Deseado, Universidad Nacional de la Patagonia Austral. Av. Prefectura s/n, 9050 Puerto Deseado, Santa Cruz, Argentina.
2 patonasca@yahoo.com.ar
3 CONICET y Wildlife Conservation Society.
Resumen
El objetivo de este trabajo fue caracterizar las asociaciones de alimentación multiespecíficas de aves marinas en la Ría Deseado, Santa Cruz. Durante un año, se registraron la composición específica, la abundancia de las especies presentes y la duración de los ensambles. Se registró un total de 91 asociaciones durante el otoño, la primavera y el verano, estando ausentes en el invierno, cuando los peces formadores de los cardúmenes están alejados de la costa y no entran a la ría. El tamaño más frecuente fue de 100 individuos y estuvieron conformadas, en promedio, por tres especies. Su duración promedio (± DE) fue de 28.8 ± 15.8 min. Se identificaron 12 especies de aves y la Tonina Overa (Cephalorhynchus commersonii) en las asociaciones. Las especies más abundantes y más participativas fueron los gaviotines (Sterna hirundinacea y Thalasseus sandvicensis), la Gaviota Cocinera (Larus dominicanus) y el Pingüino Patagónico (Spheniscus magellanicus). De las especies que nidifican en la ría y que formaron parte de las asociaciones, el Cormorán Cuello Negro (Phalacrocorax magellanicus) fue la que menos participó. El Pingüino Patagónico y los gaviotines se asociaron positivamente. En la estación reproductiva, la Gaviota Cocinera y los gaviotines se encontraron juntos. En la Ría Deseado, las asociaciones estudiadas fueron de Tipo I, en las cuales los pingüinos cumplirían un papel importante en su formación y permanencia. Esto se debe a que, al buscar sus presas buceando, podrían conducirlas hacia la superficie quedando más disponibles para los gaviotines y las gaviotas, las cuales al alimentarse superficialmente y de manera conspicua estarían actuando como especies catalíticas.
Palabras clave: Argentina; Asociaciones de alimentación multiespecíficas; Aves marinas; Gaviotines; Pingüinos; Puerto Deseado.
Abstract.
Characterization of multispecies feeding flocks of seabirds in the Firth of Deseado, Santa Cruz, Argentina
The objective of this work was to study the multispecies feeding flocks of marine birds in the Firth of Deseado, Santa Cruz. During one year, we observed specific composition, species abundance and duration of the assemblages. A total of 91 associations were recorded during the fall, spring and summer, being absent in winter, when fishes that form schools are far away from the coast and not come into the firth. The most frequent size was 100 individuals and they were formed, on average, by three species. Their average duration (± SD) was 28.8 ± 15.8 min. We identified 12 species of birds and the Commerson´s dolphin (Cephalorhynchus commersonii) in the flocks. The most abundant and frequent species were terns (Sterna hirundinacea and Thalasseus sandvicensis), Kelp Gull (Larus dominicanus) and Magellanic Penguin (Spheniscus magellanicus). The Rock Shag (Phalacrocorax magellanicus) was the least frequent one among the species that breed in the firth and which formed part of the associations. Magellanic Penguins and terns were positively associated. In the breeding season, Kelp Gulls and terns were found together. In the Firth of Deseado, studied associations were Type I, in which the penguins would be carrying out an important role in their formation. Since they are divers, they could take their prey to the surface that remains then more available for terns and gulls. These birds feed on the surface and in a conspicuous way, so they would be acting like catalysts species.
Key words: Argentina; Multispecies feeding flocks; Penguins; Puerto Deseado; Seabirds; Terns.
Recibido 21 abril 2004, aceptado 30 agosto 2004
Aves, mamíferos y peces comúnmente forman ensambles de alimentación multiespecíficos en el mar (Au y Pitman 1986, Harrison et al. 1991, Mills 1998, Camphuysen y Webb 1999). Algunas de estas asociaciones se forman cuando las presas se encuentran agrupadas, atrayendo a varias especies de predadores que pueden interactuar entre sí. Se argumenta que muchas especies se benefician cuando se alimentan en grupos, debido a que otras especies localizan o concentran a las presas. Las asociaciones pueden estar compuestas por especies que juegan roles particulares en la iniciación, mantenimiento y disgregación del grupo (Sealy 1973, Chilton y Sealy 1987, Harrison et al. 1991, Mahon 1992, Ostrand 1999).
Al usar diferentes métodos de alimentación, como predación aérea y buceo, las especies difieren en su capacidad para encontrar y capturar presas. Así, aquellas especies que encuentran una concentración de presas y que, además, poseen un comportamiento de alimentación conspicuo, funcionan como especies catalíticas, pues indican a otras aves la ubicación de las presas. Las aves buceadoras son capaces de encontrar presas bajo la superficie y pueden influenciar la distribución de éstas, concentrándolas en la superficie. Este comportamiento se ha observado en mamíferos marinos y en pingüinos, los cuales conducen a los peces hacia la superficie mientras se alimentan (Jehl 1974, Boswall y MacIver 1975, Harrison et al. 1991, Mills 1998). Además, las aves marinas que se alimentan en la superficie pueden ser atraídas por la presencia de otros predadores más que por las presas en sí mismas (Harrison et al. 1991).
La Ría Deseado (provincia de Santa Cruz, Argentina) concentra una alta diversidad de aves marinas y costeras (Gandini y Frere 1998). En su interior, muchas de estas especies nidifican y se alimentan, y cuando ingresan a la ría cardúmenes de peces pelágicos como la sardina fueguina (Sprattus fuegensis), se forman agrupaciones alimentarias multiespecíficas. El objetivo de este trabajo fue caracterizar estas asociaciones, describiendo: (1) la composición específica, el tamaño del grupo, la abundancia de cada especie y la frecuencia de participación de cada especie en las distintas asociaciones, (2) la duración de las asociaciones, y (3) la presencia de asociaciones entre las especies que conforman los grupos de alimentación.
Métodos
Área de estudio
El trabajo se llevó a cabo en la Ría Deseado (47°45'S, 65°54'O; Fig. 1). La ría tiene una longitud aproximada de 40-42 km, con un ancho en su boca de aproximadamente 1.5 km. El área se caracteriza por la presencia de fuertes vientos y una baja precipitación anual que no excede los 200 mm. Presenta playas rocosas, acantilados, cabos e islas, algunas de las cuales se conectan con el continente en las mareas bajas. La ría es utilizada por aves (marinas y costeras) y por mamíferos marinos como sitio de alimentación. Además, 12 especies de aves marinas nidifican en su interior, entre ellas el Pingüino Patagónico (Spheniscus magellanicus), la Gaviota Cocinera (Larus dominicanus), tres cormoranes (el Cormorán Gris Phalacrocorax gaimardi, el Cormorán Cuello Negro Phalacrocorax magellanicus y el Biguá Phalacrocorax olivaceus) y dos gaviotines (Sterna hirundinacea y Thalasseus sandvicensis).
Figura 1. Ubicación del sitio de estudio dentro de la Ría Deseado, Santa Cruz, Argentina.
En este estudio se define una asociación de alimentación como un grupo de 10 o más individuos de dos o más especies alimentándose activamente, mientras se encuentran a una distancia no mayor a los 3 m unos de otros. Según Hoffman et al. (1981), a las especies que participan en las asociaciones de alimentación se las puede agrupar en cuatro grupos funcionales: catalíticas, buceadoras, cleptoparásitas y supresoras. Las catalíticas son aquellas especies cuya activa búsqueda y forma de alimentación son muy conspicuas y, por lo tanto, son utilizadas por otras aves como un indicador de áreas con alta densidad de presas. Por consiguiente, determinan el rápido desarrollo de un ensamble. En la Ría Deseado, tanto la Gaviota Cocinera como los gaviotines presentan este comportamiento. Entre los buceadores se incluyen aves que buscan su alimento realizando buceos con persecución y aquellas que se zambullen desde el aire y nadan bajo el agua para capturar a sus presas. En este estudio, tales especies están representadas por los cormoranes, el Pingüino Patagónico y la tonina overa (Cephalorhynchus commersonii). Las cleptoparásitas obtienen su alimento robándolo a otras aves; en este estudio están representadas por la Gaviota Cocinera. Las supresoras son aquellas aves que, por su forma de alimentación, hacen disminuir la disponibilidad de presas para otros miembros de la asociación.
La mayor parte de las observaciones se realizaron desde un punto elevado en la costa (250 m), con binoculares (10×42) y telescopio (12-36×50), entre marzo de 1999 y marzo de 2000. Los muestreos se llevaron a cabo en tres bloques horarios: 08:30-09:30, 13:00-14:00 y 19:00-20:00 h. Fuera de estos bloques también se tomaron datos cuando se observaba una asociación. Para cada asociación se registraron la composición específica, la abundancia de las especies presentes y, cuando fue posible, la duración (en min). También se registraron las condiciones meteorológicas (intensidad del viento con la escala Beaufort) y oceanográficas (estado de marea: subiendo o bajando). Los datos de abundancia de individuos que se reproducen en la ría se obtuvieron de Gandini y Frere (1998).
Para el análisis de los datos se utilizaron métodos no paramétricos y se aceptó como significativo un valor crítico de P < 0.05. Para probar diferencias entre grupos se utilizó la Prueba de Kruskal-Wallis y, para las comparaciones múltiples, el Método de Dunn (Siegel y Castellan 1988). Para probar asociaciones significativas entre variables se usó el Análisis de Correlación de Spearman. Se utilizó la Prueba de Chi-Cuadrado para evaluar la significación de las distribuciones de frecuencia.
Resultados
Durante el otoño, la primavera y el verano se registraron 91 asociaciones, estando ausentes durante el invierno. Participaron 12 especies de aves y la tonina overa (Tabla 1). Las especies que participaron con más individuos en las asociaciones fueron los gaviotines, la Gaviota Cocinera y el Pingüino Patagónico.
Tabla 1. Especies involucradas en asociaciones de alimentación multiespecíficas en la Ría Deseado. Se indica el método de alimentación, la abundancia de las aves que nidifican en la ría (número de individuos durante la época reproductiva), la frecuencia de participación en las asociaciones (en porcentaje) y la abundancia promedio (con el rango entre paréntesis) de aves y cetáceos presentes en las asociaciones observadas.

El 66% de los ensambles se formaron cuando la marea estaba subiendo (c2 = 9.24, P < 0.002) y el 77% lo hicieron en días sin viento y cuando la ría se encontraba calma (c2 = 26.38, P < 0.0001). El tamaño más frecuente fue de 100 individuos y estuvieron conformadas, en promedio, por tres especies. Tanto para el tamaño de las asociaciones como para la riqueza específica se encontraron diferencias significativas entre estaciones (H = 22.47, n = 91, P = 0.0001 para el tamaño; H = 13.96, n = 91, P = 0.003 para la riqueza; Fig. 2). El tamaño de las asociaciones fue significativamente mayor durante el verano de 1999 con respecto al verano siguiente (Dunn = 21.19, P < 0.0167), mientras que en el otoño el número de individuos por asociación fue menor que en la primavera (Dunn = 24.13, P < 0.0167)(Fig. 2). Entre la primavera y el verano siguiente la riqueza específica y el tamaño de las asociaciones también disminuyeron significativamente (Dunn = 27.7, P < 0.0167 para la riqueza; Dunn = 35.94, P < 0.0167 para el tamaño; Fig. 2).
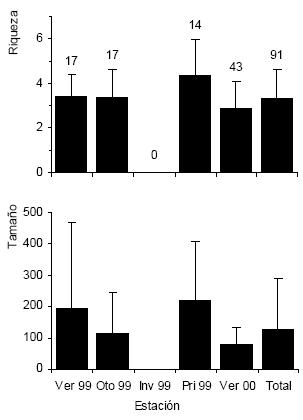
Figura 2. Riqueza específica (número de especies/asociación; arriba) y tamaño (número de individuos/asociación; abajo) de las asociaciones de alimentación multiespecíficas en la Ría Deseado en cada estación. Los datos se expresan como promedios + DE y el tamaño de muestra se indica sobre las barras.
En general, los gaviotines fueron los más numerosos y los que más participaron de las asociaciones. Se encontraron presentes en todas las estaciones (excepto en el invierno) y participaron en el 95% de las asociaciones (Tablas 1 y 2).
Tabla 2. Frecuencia de participación en las asociaciones de alimentación multiespecíficas (en porcentaje) y abundancia promedio (con el rango entre paréntesis) de aves y cetáceos presentes en las asociaciones observadas en la Ría Deseado en cada estación.
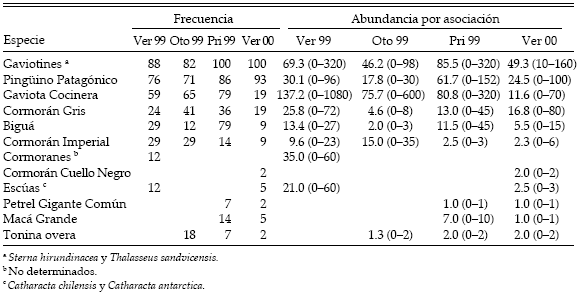
La población de Pingüino Patagónico que nidifica en la ría fue cinco veces mayor que cualquier otra especie del estudio. Sin embargo, participaron del 85% de las asociaciones (Tabla 1). El número de pingüinos presentes en las asociaciones varió estacionalmente (H = 10.86, n = 91, P = 0.0125; Tabla 2). La abundancia aumentó significativamente entre el otoño y la primavera, con el comienzo de la estación reproductiva y la llegada al área de estudio (Dunn = 29.21, P < 0.0167) y fue menor en el verano de 1999 con respecto a la primavera (Dunn = 24.62, P < 0.0167).
La abundancia y la participación de la Gaviota Cocinera variaron con las estaciones (H = 30.81, n = 91, P < 0.0001 para la abundancia; H = 22.28, n = 91, P = 0.0001 para la participación; Tabla 2). Se encontraron en mayor número en verano, otoño y primavera de 1999 con respecto al verano de 2000 (Dunn = 20.09 para verano de 1999 vs verano de 2000, Dunn = 23.38 para otoño de 1999 vs verano de 2000, Dunn = 33.06 para primavera de 1999 vs verano de 2000; P < 0.0167; Tabla 2). De la misma manera varió la frecuencia de participación (Dunn = 18.29 para verano de 1999 vs verano de 2000, Dunn = 20.97 para otoño de 1999 vs verano de 2000, Dunn = 27.28 para primavera de 1999 vs verano de 2000; P < 0.0167; Tabla 2).
De las cuatro especies de cormoranes que participaron de los ensambles, tres nidifican en las aguas interiores de la ría, siendo la más abundante el Cormorán Gris. Dicha especie participó en el 26% de las asociaciones (Tabla 1). El Biguá participó en el 24% de las asociaciones (Tabla 1). Su abundancia aumentó significativamente entre el otoño y la primavera, y disminuyó entre la primavera y el verano de 2000 (H = 30.81, n = 91, P < 0.0001; Dunn = 31.86 para otoño de 1999 vs primavera de 1999, Dunn = 32.52 para primavera de 1999 vs verano de 2000, P < 0.0167; Tabla 2). El Cormorán Imperial (Phalacrocorax atriceps) participó en un 18% de las asociaciones pese a que esta especie no nidifica en la ría (Tabla 1). De todas las especies de aves marinas que nidifican en el interior de la ría y que, a su vez, formaron parte de las asociaciones, el Cormorán Cuello Negro fue la que menos participó (Tabla 1).
Las toninas solo estuvieron presentes en el 5% de los ensambles (Tabla 1). Este porcentaje podría estar subestimado, debido a que son difíciles de observar mientras se están alimentando.
La duración promedio (± DE) de los ensambles de alimentación fue de 28.8 ± 15.8 min (n = 40). Tan solo el 10% de las asociaciones observadas tuvieron una duración mayor a 50 min, mientras que el 45% duró no más de 20 min. La máxima duración fue de 65 min. En todas las asociaciones se encontraron presentes especies buceadoras (cormoranes, pingüinos o toninas). No se encontraron diferencias significativas entre la riqueza específica y la duración promedio de las asociaciones (Fig. 3). La tabla 3 ilustra las diferencias en el tamaño y la persistencia de las asociaciones con y sin individuos de Gaviota Cocinera, Pingüino Patagónico, Cormorán Gris y Biguá. Los gaviotines estuvieron presentes en todos los ensambles y, en consecuencia, no es posible realizar este tipo de comparación. Cuando el Pingüino Patagónico participó de las asociaciones, el tamaño fue mayor (H = 3.95, n = 40, P = 0.047) y su duración tendió a ser mayor con respecto a aquellas en las cuales se encontró ausente. Los ensambles que contaron con la presencia del Cormorán Gris también tuvieron mayor tamaño (H = 4.06, n = 40, P = 0.044), pero su persistencia no fue significativamente diferente (H = 0.31, n = 40, P = 0.575).
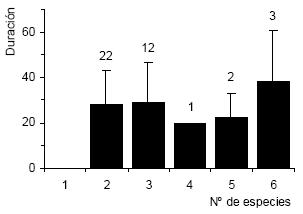
Figura 3. Duración (en min) de las asociaciones de alimentación multiespecíficas en la Ría Deseado en función de la riqueza específica. Los datos se expresan como promedios + DE y el tamaño de muestra se indica sobre las barras.
Tabla 3. Tamaño (número de individuos) de las asociaciones de alimentación multiespecíficas en la Ría Deseado y su duración (en min) en presencia y ausencia de Gaviota Cocinera, Pingüino Patagónico, Cormorán Gris y Biguá. Para cada una de estas especies se indica, entre paréntesis, el porcentaje de presencia en las asociaciones. Los datos se expresan como promedios ± DE.
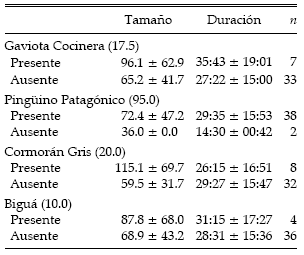
Algunas de las especies de aves marinas en la Ría de Puerto Deseado se encontraron juntas. Independientemente de la estación, se encontró una asociación positiva entre el Pingüino Patagónico y los gaviotines (r = 0.33, n = 91, P = 0.001). A su vez, se encontraron juntos a la Gaviota Cocinera y los gaviotines durante la estación reproductiva (r = 0.40, n = 57, P = 0.002).
Discusión
Las asociaciones de alimentación se forman cuando se concentra una cantidad abundante de alimento para los predadores y una vez que una o varias aves comienzan a alimentarse activamente, mientras que otras que se encuentran próximas al parche también se acercan para capturar presas. De acuerdo a Hoffman et al. (1981), las asociaciones en la Ría Deseado fueron del Tipo I, es decir, relativamente pequeñas, de corta duración y que se formaron una vez que un cardumen era descubierto. Su ausencia en los meses de invierno coincide con la temporada en la cual los peces formadores de los cardúmenes se encuentran alejados de la costa y, por lo tanto, no entran en la ría (Sánchez et al. 1995, Cousseau y Perrotta 1998).
Factores ambientales pueden afectar a los peces y, a su vez, la distribución de éstos afecta a las aves (Safina y Burger 1988). Cuando la claridad del agua es alta, los peces son más visibles desde la superficie y tienden a bajar hacia aguas más profundas escapando de las aves predadoras. Safina y Burger (1988) estudiaron grupos monoespecíficos de Gaviotín Golondrina (Sterna hirundo) y encontraron que estas aves no incrementan las capturas de presas al aumentar la claridad del agua, y que cuando el viento disminuye la visibilidad de la superficie del mar, esto dificulta a los gaviotines la localización de presas. También los ciclos de marea frecuentemente afectan la actividad de alimentación de estos gaviotines, siendo mayor con la bajante debido a que bajos niveles de agua pueden concentrar peces y remover del fondo a los invertebrados, haciendo más disponible la comida para estas aves. En nuestro estudio, los ensambles se formaron en días sin viento y con la ría calma, donde las asociaciones entre especies que se alimentan en la superficie, como los gaviotines, y aquellas que lo hacen buceando, como el Pingüino Patagónico, parece ser crítica en la formación de estos ensambles. Por un lado, los pingüinos pueden conducir hacia la superficie a grupos de presas que quedan disponibles para otras especies y evitar que bajen hacia aguas más profundas (Jehl 1974, Boswall y MacIver 1975, Harrison et al. 1991, Mills 1998). Por otro lado, los gaviotines y las gaviotas, que buscan su alimento desde el aire, podrían visualizar mejor los cardúmenes más superficiales. Además, al alimentarse de manera conspicua estarían actuando como especies indicadoras de áreas con alta densidad de presas (catalíticas). A diferencia de lo observado por Safina y Burger (1988), la mayoría de las asociaciones se formaron cuando la marea estaba subiendo, coincidiendo con la entrada de los cardúmenes hacia el interior de la ría. Pese a que las toninas incrementan su actividad de alimentación en ese momento del ciclo de mareas (C Righi, com. pers.), su participación en los ensambles fue muy baja.
Según Mills (1998), otro factor importante en la formación de las asociaciones es el reclutamiento visual. Las asociaciones de mayor duración pueden atraer un mayor número de especies y un mayor número de individuos. En este estudio, aunque se observó cierta relación positiva entre el número de especies y la duración, ésta no se pudo corroborar. El Pingüino Patagónico mostró un efecto positivo para el inicio y el mantenimiento de las asociaciones, pues cuando estuvo presente los ensambles tuvieron un mayor tamaño y mostraron una tendencia a aumentar su persistencia. La falta de significación de estos resultados podría deberse al bajo número de asociaciones en las cuales fue posible calcular la duración.
La abundancia de las especies de aves que conforman las asociaciones de alimentación no fue un simple reflejo de su abundancia en la ría en la estación reproductiva. Los gaviotines fueron el grupo más abundante y participativo en las asociaciones, pero los terceros en importancia en cuanto a número de parejas nidificantes, mientras que el Pingüino Patagónico es la especie de mayor tamaño poblacional que nidifica en la ría aunque sólo una mínima proporción de estos individuos integró las asociaciones. Los pingüinos tienen rangos de alimentación que oscilan entre 30-50 km de distancia durante la estación reproductiva (Frere y Gandini, datos no publicados), por lo que la mayoría se estaría alimentando fuera de la ría y, por lo tanto, no participarían de las asociaciones que se forman dentro de la ría. Además, no todas las especies que nidifican en el interior de la ría participaron de las asociaciones, como fue el caso del Cormorán Cuello Negro. Tanto el Cormorán Gris como el Cormorán Cuello Negro nidifican en las paredes de acantilados de la ría y poseen una dieta muy similar, basada en peces e invertebrados bentónicos (Gandini y Frere 1995, 1998, Malacalza et al. 1997, Frere y Gandini 2001, Frere et al. 2002, Millones et al., datos no publicados). Mientras que el Cormorán Gris participa de las bandadas de alimentación de manera oportunista (Frere et al. 2002), la ausencia del Cormorán Cuello Negro podría deberse a que se alimenta en la desembocadura de la ría y no en su interior como lo hace el Cormorán Gris (Quintana et al., datos no publicados).
En conclusión, este estudio representa una primera aproximación al estudio de bandadas mixtas de alimentación entre aves marinas en un área geográfica de indudable importancia para la reproducción y alimentación de aves y mamíferos marinos. Varios interrogantes quedan pendientes para futuras investigaciones, tales como los complejos procesos de formación de estas asociaciones (i.e., qué especies las inician), el orden o secuencia de llegada de las diferentes especies a la asociación y cuáles actúan como supresoras desmembrándola.
Agradecimientos. Agradecemos a la Wildlife Conservation Society y a la Universidad Nacional de la Patagonia Austral por proveer los fondos necesarios para llevar a cabo este trabajo. A Chantal Torlaschi por su colaboración en las tareas de campo.
Bibliografía Citada
Au D y Pitman R (1986) Seabird interactions with dolphins and tuna in the eastern tropical Pacific. Condor 88:304-317 [ Links ]
Boswall J y MacIver D (1975) The Magellanic Penguin Spheniscus magellanicus. Pp. 271-306 en: Stonehouse B (ed) The biology of penguins. MacMillen Press, Londres [ Links ]
Camphuysen CJ y Webb A (1999) Multi-species feeding associations in North Sea seabirds: jointly exploiting a patchy environment. Ardea 87:177-198 [ Links ]
Chilton G y Sealy SG (1987) Species roles in mixed-species feeding flocks of seabirds. Journal of Field Ornithology 58:456-463 [ Links ]
Cousseau MB y Perrotta RG (1998) Peces marinos de Argentina: biología, distribución, pesca. INIDEP, Mar del Plata [ Links ]
Frere E y Gandini PA (2001) Aspects of the breeding biology of the red-legged cormorant Phalacrocorax gaimardi on the Atlantic coast of South America. Marine Ornithology 29:67-70 [ Links ]
Frere E, Quintana F y Gandini PA (2002) Diving behaviour of the red-legged cormorant in southeastern patagonia, Argentina. Condor 104:440-444 [ Links ]
Gandini PA y Frere E (1995) Distribución, abundancia y ciclo reproductivo del cormorán gris Phalacrocorax gaimardi en la costa patagónica, Argentina. Hornero 14:57-60 [ Links ]
Gandini PA y Frere E (1998) Seabirds and shorebirds diversity and associated conservation problems in Puerto Deseado, Patagonia, Argentina. Ornitologia Neotropical 9:13-22 [ Links ]
Harrison NM, Whitehouse MJ, Heinemann D, Prince PA, Hunt GL Jr y Veit RR (1991) Observations of multispecies seabirds flocks around South Georgia. Auk 108:801-810 [ Links ]
Hoffman W, Heinemann D y Wiens JA (1981) The ecology of seabirds feeding flocks in Alaska. Auk 98:437-456 [ Links ]
Jehl JR Jr (1974) The distribution and ecology of marine birds over the continental shelf of Argentina in winter. Transactions of the San Diego Society of Natural History 17:217-234 [ Links ]
Mahon TE (1992) The role of Marbled Murrelets in mixed-species feeding flocks in British Columbia. Wilson Bulletin 104:738-743 [ Links ]
Malacalza VE, Bertellotti NM y Poretti TI (1997) Variación estacional de la dieta de Phalacrocorax magellanicus (Aves: Phalacrocoracidae) en Punta Loma (Chubut, Argentina). Neotropica 43:35-37 [ Links ]
Mills KL (1998) Multispecies seabird feeding flocks in the Galápagos Islands. Condor 100:277-285 [ Links ]
Ostrand WD (1999) Marbled murrelets as initiators of feeding flocks in Prince William Sound, Alaska. Waterbirds 22:314-318 [ Links ]
Safina C y Burger J (1988) Ecological dynamics among prey fish, bluefish and foraging common terns in an Atlantic coastal system. Pp. 95-173 en: Burger J (ed) Seabirds and other marine vertebrates. Columbia University Press, New York [ Links ]
Sanchez RP, Remeslo A, Madirolas A y Ciechomski JDZ (1995) Distribution and abundance of post-larvae and juveniles of the Patagonian sprat, Sprattus fuegensis and related hydrographical conditions. Fisheries Research 23:47-81 [ Links ]
Sealy SG (1973) Interespecific feeding assemblages of marine birds off British Columbia. Auk 90:796-802 [ Links ]
Siegel S y Castellan NJ Jr (1988) Nonparametric statistics for the behavioral sciences. Segunda edición. McGraw Hill International, Singapore. [ Links ]

