










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEl hornero
versión impresa ISSN 0073-3407versión On-line ISSN 1850-4884
Hornero v.20 n.2 Buenos Aires sept./dic. 2005
Dinámica estacional del ensamble de aves de un bosque norpatagónico de lenga (Nothofagus pumilio) y su relación con la disponibilidad de sustratos de alimentación
Rodrigo M. Becerra Serial1 y Dora Grigera1,2
1Centro Regional Universitario Bariloche, Universidad Nacional del Comahue. 8400 San Carlos de Bariloche, Río Negro, Argentina.
2dgrigera@crub.uncoma.edu.ar
Resumen
Se estudió la dinámica estacional de la composición específica y de la abundancia de las aves de un bosque de lenga en el valle del Ñireco, Parque Nacional Nahuel Nuapi, Argentina, y de los patrones de uso de los sustratos de alimentación. La diversidad específica y la densidad del ensamble fueron mucho más muy bajas en invierno que en primavera y verano. La riqueza de especies fue mínima en invierno, pero no presentó variaciones importantes entre las demás estaciones del año. Casi todas las especies presentes en invierno (7 de un total de 23) usaron principalmente los troncos y las ramas para alimentarse, dado que estos sustratos están disponibles y poseen alimento todo el año. Las aves ausentes en invierno utilizaron para alimentarse principalmente el suelo y, en menor medida que este sustrato, el follaje y el aire. Las usuarias del suelo no pueden permanecer en invierno en el área porque ese sustrato, a pesar de contener alimento, se encuentra cubierto de nieve. La falta de alimento y no la disponibilidad del sustrato de alimentación determinaría la ausencia invernal de las especies usuarias del follaje y del aire. El ensamble estudiado está compuesto mayormente por especies restringidas en el uso de los sustratos, principalmente en otoño e invierno. Cuatro especies de alto valor de conservación obtienen la mayor parte de su alimento de los troncos y de las ramas de las lengas; es probable que su sobrevivencia invernal dependa de la disponibilidad de este sustrato.
Palabras clave: Bosque de lenga; Nothofagus pumilio; Sustratos de alimentación; Patagonia.
Abstract
Seasonal dynamics of a bird assemblage in a north patagonian lenga (Nothofagus pumilio) forest and its relationship with the availability of foraging substrates
We describe the seasonal dynamics of species composition and abundance of a bird assemblage in a lenga forest at Ñireco Valley, Nahuel Huapi National Park, Argentina, and the use of feeding substrates. Species diversity and density were lower in winter than in spring and summer. Species richness was minimum in winter but it has not variations in the other seasons. Most of the species present in winter (7 over 23) mainly used tree trunks and branches as feeding substrates. These substrates and their food supply are available during all the year. Birds that were absent in winter mainly used the soil, and to a lesser extent foliage and air as feeding substrates. Soil users can not overwinter in the area because although the soil contains food, it is covered in snow. Lack of food may determine the absence of foliage- and air-users in winter. Most species used a low number of substrates for feeding, mainly in fall and winter. Four species of high conservation value mainly feed on tree trunks and branches; it is possible that their winter survival depends on the availability of these substrates.
Key words: Lenga forest; Nothofagus pumilio; Feeding substrates; Patagonia.
Recibido 23 agosto 2004, aceptado 26 mayo 2005
La abundancia y la diversidad son parámetros de primordial importancia en el estudio de las comunidades, ya que constituyen la base para la investigación de hipótesis sobre su funcionamiento y sobre los procesos de estructuración. Jaksic y Feinsinger (1991) señalaron la falta de estudios sobre la abundancia y la diversidad específica de aves andinopatagónicas y sobre sus patrones espacio-temporales de uso del hábitat, al mismo tiempo que destacaron su importancia no solo para acrecentar el conocimiento sino para la formulación de prioridades de conservación. Vuilleumier (1972) y Ralph (1985) describieron la variación de la abundancia y la diversidad específica de aves de la región andinopatagónica; si bien incluyeron bosques de lenga (Nothofagus pumilio) entre los ambientes estudiados, solo trabajaron durante la temporada reproductiva. Iglesias (datos no publicados) estudió las mismas variables durante dos ciclos anuales consecutivos, pero no trabajó en bosques puros de lenga. Así, este trabajo es el primero en describir estos parámetros a lo largo de todo el año en estos ecosistemas patagónicos.
Los bosques de lenga se desarrollan en ambientes sometidos a condiciones físicas severas, entre ellas el incremento de la rigurosidad climática en el invierno y la disminución de la heterogeneidad espacial debida al carácter caducifolio de la lenga y a la cubierta de nieve permanente durante dicha estación. La disponibilidad de recursos es un factor determinante de la densidad para las poblaciones de aves (Raitt y Pimm 1976, Loiselle y Blake 1991, Marone 1992) y frecuentemente se recurre a la variabilidad en la disponibilidad de alimento para explicar los cambios estacionales en la estructura de los ensambles (e.g., Brooks 1997, Soave et al. 1999, Cueto y Lopez de Casenave 2000). Carrascal y Tellería (1985) mostraron la importancia del uso de los sustratos al encontrar en las aves de un bosque que dicha variable era la de mayor valor explicativo de la segregación entre las especies del ensamble. Por su parte, Lopez de Casenave (2001) estudió el uso de sustratos de alimentación como una de las cuatro variables (excluido el tipo de alimento) necesarias para establecer el patrón de alimentación de las aves.
Desde un punto de vista aplicado, teniendo en cuenta que la lenga es la especie forestal nativa de mayor valor económico de la Patagonia y la más codiciada para su explotación industrial (Laclau 1998), es importante conocer la estructura y el funcionamiento de los bosques de lenga en áreas libres de intervención humana a fin de tener datos testigo para poder evaluar el impacto de las acciones humanas sobre estos ambientes.
Los objetivos de este trabajo son (1) describir la composición específica y la abundancia del ensamble de aves de un bosque de lenga no perturbado a lo largo de un ciclo anual y (2) describir las variaciones en el uso de los sustratos de alimentación.
MétodosLos bosques de lenga ocupan la mayor superficie de la región de los bosques patagónicos y son los de mayor rango latitudinal y altitudinal entre los bosques de especies del género Nothofagus. Se extienden sobre la Cordillera de los Andes, desde los 37°S hasta los 55°S, en altitudes entre 1000-1700 msnm en el norte de su área de distribución y entre 0-600 msnm en Tierra del Fuego (Hueck 1978). Este estudio se llevó a cabo en un bosque puro de lenga, entre los 1280-1550 msnm en un área denominada “Valle del Ñireco” (41°14'S, 71°22'O), dentro de la Reserva del Parque Nacional Nahuel Huapi, Argentina (Fig. 1). En esta zona los inviernos son fríos y los veranos secos y templados con noches frías; más del 75% de las precipitaciones ocurre en la época invernal, entre abril y septiembre, cuando en las zonas de mayor altitud predominan las nevadas (Muñoz y Garay 1985). En el bosque estudiado el sotobosque es poco denso debido a la ausencia de caña coligüe (Chusquea culeou), y está constituido por arbustos bajos (e.g., Berberis serratodentata, Ribes magellanicum y Maytenus chubutensis) e hierbas (e.g., Vicia nigricans, Adenocaulon chilense y Alstroemeria aurea). Por la presencia de endemismos, entre otras características, el área de estudio forma parte de uno de los sitios de alto valor biológico de América Latina (Vila 2000).

Figura 1. Mapa de ubicación del área de estudio (valle del Ñireco) dentro de la Reserva del Parque Nacional Nahuel Nuapi, Argentina. Se muestran las unidades ambientales presentes en el parque según Mermoz et al. (2000).
Se realizaron censos de aves durante cuatro días consecutivos en marzo (otoño), agosto (invierno) y octubre (primavera) de 1997 y en enero (verano) de 1998, mediante el método de línea de transecta de Emlen modificado por Bibby et al. (1992). Este método es el más apropiado para cubrir áreas grandes y uniformes que albergan pocas especies e individuos, como es el caso del bosque estudiado. Los censos comenzaron siempre al amanecer, en días soleados y sin viento, a fin de que los datos de las distintas estaciones fuesen comparables y de minimizar la incidencia de factores que disminuyen la actividad de las aves o perturban su detectabilidad (Bibby et al. 1992). La transecta, de 3500 m de longitud, fue recorrida en un tiempo promedio (± DE) de 2.35 ± 0.18 h en cada ocasión y el tiempo total de relevamiento fue de 37.33 h. Se registraron todos los individuos vistos u oídos en 11 bandas contiguas situadas a cada lado de la transecta y paralelas entre sí. El ancho de cada una de las primeras 10 bandas fue de 5 m y el de la undécima de 50 m. En cada registro se identificó la especie, el número de banda en la que se encontraba, el sustrato que ocupaba y si estaba buscando o ingiriendo alimento. Los sustratos, sobre la base de Holmes y Recher (1986), fueron clasificados en: suelo; hierbas y arbustos, que incluye los renovales de lenga de porte inferior a 1.5 m; troncos caídos; troncos en pie; ramas, que comprende las ramas de diámetro superior a 10 cm; follaje, constituido por las ramas de diámetro menor a 10 cm; y aire.
La estructura del ensamble en cada estación se describió mediante los siguientes parámetros: (1) riqueza de especies; (2) densidad de individuos por especie y densidad total (la suma de las densidades de cada especie), obtenidas a partir del promedio de los cuatro días de censo por estación (según Bibby et al. 1992); este promedio subestima la densidad de las especies poco abundantes o conspicuas, pero evita la sobreestimación que causa el uso de los números máximos, que pueden ser obtenidos por recuentos dobles; (3) diversidad específica, estimada mediante el índice de Shannon-Wiener, H' (Zar 1999); y (4) equitatividad, estimada como J' = H' / H'máx= H' / log10 k, donde H'máx es el máximo valor que podría tener H' si todas las poblaciones tuviesen la misma densidad y k es el número de especies (Zar 1999). Los valores del índice de diversidad específica fueron comparados con la Prueba t de Hutchenson (Zar 1999). Para determinar la significación estadística de este análisis se fijó un nivel de P < 0.05.
Para comparar la composición específica y el tamaño de las poblaciones de los ensambles, se calcularon las distancias euclidianas entre los valores de densidad por especie obtenidos en cada estación (Krebs 1989).
Se estudió la relación existente entre la permanencia de las especies a lo largo del año y el uso de los sustratos de donde obtienen su alimento. Los datos de frecuencia de uso de sustratos de alimentación fueron transformados en proporciones y ordenados en cuatro matrices (una por cada estación, para diferenciar estacionalmente los ensambles) de 7 × n, siendo 7 los sustratos y n el número de especies presentes en cada caso. Cada matriz fue sometida a un Análisis de Agrupamiento, utilizando distancias euclidianas para obtener la matriz de similitud y ligamiento promedio aritmético no ponderado (“UPGMA”) a fin de construir el dendrograma (Crisci y López Armengol 1983). Para fijar el nivel de similitud que define los grupos en cada dendrograma se consideraron simultáneamente dos criterios: (1) la distancia máxima entre agrupamientos sucesivos del dendrograma y (2) el promedio de similitud entre todos los pares de especies del ensamble (Lopez de Casenave 2001). Si ambos criterios no eran coincidentes se optó por aquel que incluye la menor cantidad de asociaciones entre especies (i.e., mayor cantidad de grupos, por lo tanto más pequeños), ya que en el Análisis de Agrupamiento la distorsión se incrementa hacia los grupos más grandes (Crisci y López Armengol 1983). Los grupos obtenidos fueron denominados en función del sustrato de alimentación más usado por las especies que los componen.
Resultados
Se registró un total de 23 especies, 17 passeriformes y 6 no passeriformes. Cuatro especies observadas fuera de los censos (Polyborus plancus, Sephanoides sephaniodes, Xolmis pyrope y Phrygilus unicolor) no se incluyeron en los análisis al no disponer de registros de distancias.
De las 19 especies registradas en los muestreos, solo 7 permanecieron en el área de estudio durante el invierno (Tabla 1). Entre las aves presentes en invierno se distinguió a las no migratorias (aquellas cuya densidad no mostró mayores variaciones entre el invierno y el verano) y a las migratorias parciales (aquellas cuya densidad durante el invierno disminuyó en un 50% o más con relación al verano). En el grupo de especies ausentes en invierno se pudo diferenciar entre las registradas en el bosque en al menos dos de las estaciones restantes (las emigrantes invernales) y aquellas que son visitantes ocasionales del bosque, presentes en solo una estación fuera de la invernal y en bajas densidades.
Tabla 1. Densidad estacional (ind/100 ha) y categoría de residencia de las especies de aves del bosque de lenga (Nothofagus pumilio) del valle del Ñireco, Parque Nacional Nahuel Nuapi, Argentina. Para cada estación se muestra además la densidad total (ind/100 ha), la riqueza (número de especies), la diversidad específica (H') y la equitatividad (J').

La diversidad específica fue máxima en verano y mínima en invierno (Tabla 1). Excepto entre otoño e invierno (P = 0.337), los valores del índice fueron significativamente distintos entre estaciones (P < 0.05). La densidad mostró el mismo patrón general de variación (máxima en verano y mínima en invierno). Los valores de riqueza del otoño, la primavera y el verano difirieron poco entre sí y fueron relativamente altos en comparación con el número de especies presentes durante el invierno. Por el contrario, la equitatividad fue máxima en esa estación (Tabla 1).
La mayor similitud en la composición y abundancia relativa de los ensambles de aves se observó entre el otoño y el invierno, mientras que la mayor disimilitud ocurrió entre el invierno y el verano (Tabla 2). Durante el invierno no se encontró ninguna especie exclusiva, mientras que se registraron dos especies exclusivas en cada una de las estaciones restantes (Tabla 1).
Tabla 2. Distancia euclidiana entre estaciones, calculada a partir de los valores de densidad por especie, para las aves del valle del Ñireco, Parque Nacional Nahuel Nuapi, Argentina.

Los sustratos de alimentación más usados por las especies presentes en invierno fueron los troncos y las ramas (en todas las estaciones), en particular por los pícidos (Campephilus magellanicus, Picoides lignarius y Colaptes pitius), Pygarrhichas albogularis y Enicognathus ferrugineus (Tabla 3). También Aphrastura spinicauda usó mayormente los troncos y las ramas para alimentarse, aunque además usó otros sustratos. Phrygilus patagonicus no fue vista alimentándose en invierno. A diferencia de las otras especies presentes en invierno, esta especie fue vista mayormente alimentándose en el follaje en otoño y primavera, y usó ramas, follaje, hierbas y arbustos en verano. Las especies ausentes en invierno obtuvieron su alimento principalmente del suelo y, en menor medida, del follaje y el aire (Tabla 3). Cinclodes fuscus, Pteroptochos tarnii, Scytalopus magellanicus y Upucerthia dumetaria usaron exclusivamente el suelo, mientras que Troglodytes musculus y Turdus falcklandii lo utilizaron en mayor proporción que a arbustos y ramas. Elaenia albiceps usó mayormente el follaje en primavera y las ramas en verano. En ambas estaciones Carduelis barbata utilizó principalmente el follaje para alimentarse y Tachycineta meyeni sólo se alimentó en el aire.
Tabla 3. Porcentaje de uso de sustratos de alimentación por parte de las aves del valle del Ñireco, Parque Nacional Nahuel Nuapi, Argentina, durante las cuatro estaciones del año. Para cada especie se indica el número total de individuos observados. Su: suelo, TrC: troncos caídos, HyA: hierbas y arbustos, Ra: ramas, Tr: tronco, Fo: follaje, Ai: aire.
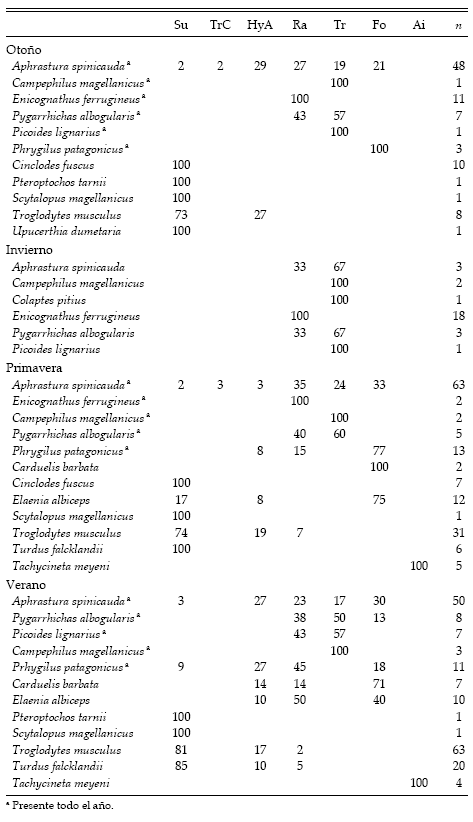
En la figura 2 se presentan los dendrogramas resultantes del Análisis de Agrupamiento. Aunque en este análisis se incluyeron algunas especies que contaron con un bajo número de registros de uso de sustrato de alimentación, la mayoría de estos casos son consistentes con la información reportada en la bibliografía (Canevari et al. 1991, Grigera et al. 1994). En otoño, invierno y verano, los dos criterios utilizados para determinar los grupos produjeron resultados coincidentes. No habiendo sido así en la primavera, se optó por el promedio de las distancias euclidianas entre todos los pares de especies. A lo largo del ciclo anual se identificaron cinco grupos de especies, que fueron denominados (1) usuarias de suelo, (2) de ramas, (3) de tronco, (4) de follaje y (5) de aire. Los grupos de usuarias de tronco y de ramas estuvieron presentes en todas las estaciones, aunque en el verano, junto con las que usaron mayormente el follaje, formaron un único conjunto que se denominó usuarias de árbol. Los grupos de aves que se alimentan principalmente en el suelo, el follaje y el aire (este último integrado solo por Tachycineta meyeni), estuvieron ausentes en invierno. En otoño se identificaron cuatro grupos. Los grupos de usuarias de tronco y de suelo fueron los de mayor número de especies, aunque el segundo estuvo compuesto por menos individuos que en el verano.

Figura 2. Dendrogramas resultantes de los Análisis de Agrupamiento estacionales basados en el uso de sustratos de alimentación por parte de las aves del valle del Ñireco, Parque Nacional Nahuel Nuapi, Argentina. La línea discontinua indica la distancia máxima entre agrupamientos sucesivos del dendrograma y la línea continua el promedio de similitud entre todos los pares de especies del ensamble. Los códigos de los sustratos (a la izquierda del dendrograma) son los mismos que en la tabla 3 y los de las especies son los mismos que en la Tabla 1.
Discusión
Si bien este estudio fue realizado en un solo sitio y durante un único año, es probable que los patrones observados no cambien sustancialmente en otros bosques del mismo tipo. Los valores bajos de riqueza, abundancia y diversidad de la avifauna del bosque estudiado son comunes al conjunto de los bosques subantárticos, y son consecuencia de procesos geológicos y paleoclimáticos (como la orogenia andina y las glaciaciones del cuaternario) que afectaron a la región patagónica dejando a estos bosques con una fauna empobrecida y en condiciones de insularidad (Vuilleumier 1985, Villagrán 1991). Durante el verano las condiciones ambientales (temperatura promedio, disponibilidad de alimento y heterogeneidad ambiental) son las más favorables del ciclo anual y, aunque la riqueza tuvo escasa variación con respecto a la primavera y al otoño, la diversidad y la densidad de individuos fueron más altas que en el resto de las estaciones, sobre todo en comparación con el invierno. Si bien en el otoño y en la primavera se dan situaciones supuestamente similares en cuanto a temperatura, precipitaciones y heterogeneidad espacial, la mayor densidad observada durante la primavera permite suponer que en el otoño hay menos alimento disponible, debido probablemente a la existencia de una menor cantidad de agua en el suelo durante esa estación, a causa del déficit hídrico del verano precedente y también al menor vigor de la vegetación por la inminente caída de las hojas de la lenga. La emigración de la mayoría de las especies en el invierno y la disminución de la abundancia de algunas de las que permanecen, determinan un gran cambio en la estructura del ensamble en dicha estación. Esta variación puede explicarse en términos de una respuesta de las especies a la rigurosidad climática del invierno y a la disminución de la disponibilidad de alimento debida a la baja productividad de ciertos sustratos (e.g., el follaje), o bien a la disminución de su accesibilidad.
La dinámica de los cinco grupos identificados a lo largo del año guarda relación con la variación en la disponibilidad y productividad de los sustratos, factores que influyen sobre la oferta de alimento en cada estación. Los grupos presentes en todas las estaciones están compuestos por aves que se alimentan en tronco y en ramas, sustratos disponibles y con alimento durante todo el año. Aún en invierno hay larvas de insectos sobre y debajo de la corteza de los troncos y las ramas (M Khun, com. pers.), y sobre las lengas crecen plantas de Misodendrum spp. y líquenes (Usnea spp. y Protousnea spp.). Las larvas de insectos son el alimento de las tres especies de pícidos que forman parte de estos grupos, así como de Pygarrhichas albogularis, que puede extraer larvas ubicadas debajo de la corteza sondeando con su pico (Grigera et al. 1996). Aphrastura spinicauda es menos hábil para esta tarea por las características de su pico (Grigera 1982) y se alimenta también de otros sustratos no productivos en el invierno (follaje, hierbas y arbustos) o no accesibles (troncos caídos); por esta razón, quizás no pueda sustentarse la población entera durante esta estación y parte de la misma deba migrar. Enicognathus ferrugineus, usuaria de las ramas, en primavera y en verano consume casi exclusivamente flores, semillas y follaje de lenga, mientras que durante el invierno se alimenta de los brotes de Misodendrum spp. (Díaz 2004) y de líquenes (R Becerra Serial, obs. pers.), estrategia que le permite permanecer en el bosque todo el año. En el invierno están ausentes los frutos y semillas que forman parte de la dieta de Phrygilus patagonicus; sin embargo, esta especie es la segunda en valores de densidad con relación a las otras aves presentes en esa estación. Está documentada la inclusión de insectos en su dieta, pero en mínima cantidad y durante el verano (Grigera 1976), de modo que es necesaria mayor información para explicar la presencia de esta especie en la estación más desfavorable del año.
El grupo de usuarias del suelo no puede permanecer en invierno porque ese sustrato, a pesar de contener distintos estadios de los invertebrados que constituyen su alimento (M Khun, com. pers.) es inaccesible al encontrarse cubierto por la nieve. En el otoño, las secuelas de la sequía del verano podrían afectar a las especies usuarias del suelo. El carácter caducifolio de las lengas y la ausencia de flores, semillas y frutos determinan que las aves usuarias del follaje tampoco puedan permanecer durante el invierno.
La existencia en verano de un grupo de aves que utilizan los árboles, formado por especies que en otras estaciones componen tres grupos, probablemente se debe al aumento de la cantidad de alimento, lo que les permite utilizar una mayor diversidad de sustratos de alimentación y así ampliar la expresión de su nicho trófico.
Al analizar globalmente el uso de los sustratos de alimentación por parte del ensamble estudiado, las características más notables que surgen son que la mayoría de las especies están restringidas en el uso de los sustratos, principalmente en otoño e invierno, y que los grupos más ricos en especies contienen a las especies más abundantes. La restricción en el uso de microhábitats fue señalada por Grigera et al. (1994) como una característica de todos los vertebrados de los bosques del Parque Nahuel Huapi. En cuanto a las aves que se encontraron presentes en invierno, parece que su permanencia está determinada primariamente por la disponibilidad de los sustratos portadores de alimento. Debería estudiarse si la coexistencia en esta estación es posible por la segregación en el uso de microhábitats más que por el tipo de alimento o por la temporalidad, dado que, con excepción de Enicognathus ferrugineus, todas las especies del gremio son insectívoras y de hábitos diurnos (Grigera 1982, Grigera et al. 1994).
Entre las especies presentes en invierno, Enicognathus ferrugineus, Pygarrhichas albogularis y Aphrastura spinicauda constituyen endemismos regionales de nivel genérico (Vuilleumier 1985), en tanto que Campephilus magellanicus posee un alto valor de conservación (Úbeda y Grigera 1995, Grigera et al. 1996) y fue seleccionada como especie focal de la Ecorregión Valdiviana (Vila 2000). Estas cuatro especies mostraron un uso importante de los troncos para alimentarse, por lo cual la disponibilidad de este sustrato es crucial para su sobrevivencia invernal en los bosques de lenga. Esta información debería ser tenida en cuenta, tanto dentro como fuera de las áreas protegidas, para el establecimiento de políticas de conservación y prácticas de manejo.
Agradecimientos. Agradecemos a Claudio Knaup por su ayuda en el trabajo de campo, a Carmen Úbeda por sus constructivos comentarios y a los revisores que con sus observaciones enriquecieron este artículo.
Bibliografía citada
BIBBY CJ, BURGESS ND Y HILL DA (1992) Bird census techniques. Academic Press, Londres [ Links ]
BROOKS DM (1997) Avian seasonality at a locality in the central Paraguayan Chaco. Hornero 14:193-203 [ Links ]
CANEVARI M, CANEVARI P, CARRIZO GR, HARRIS G, RODRÍGUEZ MATA J Y STRANEK R (1991) Nueva guía de las aves argentinas. Fundación Acindar, Buenos Aires [ Links ]
CARRASCAL LM Y TELLERÍA JL (1985) Estudio multidimensional del uso del espacio en un grupo de aves insectívoras forestales durante el invierno. Ardeola 32:95-113 [ Links ]
CRISCI JV Y LÓPEZ ARMENGOL MF (1983) Introducción a la teoría y práctica de la taxonomía numérica. OEA, Washington DC [ Links ]
CUETO VR Y LOPEZ DE CASENAVE J (2000) Bird assemblages of protected and exploited coastal woodlands in east-central Argentina. Wilson Bulletin 112:395-402 [ Links ]
DÍAZ JIS (2004) Predación predispersiva de Enicognathus ferrugineus en bosques de Nothofagus pumilio: patrones y efectos. Tesis de licenciatura, Universidad Nacional del Comahue, San Carlos de Bariloche [ Links ]
GRIGERA D (1976) Ecología alimentaria de cuatro especies de Fringillidae frecuentes en la zona del Nahuel Huapi. Physis, C 35:279-292 [ Links ]
GRIGERA D (1982) Ecología alimentaria de algunas paseriformes insectívoras frecuentes en la zona del Nahuel Huapi. Ecología 7:67-84 [ Links ]
GRIGERA D, ÚBEDA C Y CALÍ S (1994) Caracterización ecológica de la asamblea de tetrápodos del Parque y Reserva Nacional Nahuel Huapi. Revista Chilena de Historia Natural 67:273-298 [ Links ]
GRIGERA D, ÚBEDA C Y RECA A (1996) Estado de conservación de las aves del Parque y Reserva Nacional Nahuel Huapi. Hornero 14:1-13 [ Links ]
HOLMES RT Y RECHER HF (1986) Determinants of guild structure in forest bird communities: an intercontinental comparison. Condor 88:427-439 [ Links ]
HUECK K (1978) Los bosques de Sudamérica. Sociedad Alemana de Cooperación Técnica, Eschborn [ Links ]
JAKSIC FM Y FEINSINGER P (1991) Bird assemblages in temperate forests of North and South America: a comparison of diversity, dynamics, guild structure, and resource use. Revista Chilena de Historia Natural 64:491-510 [ Links ]
KREBS CJ (1989) Ecological methodology. Harper Collins, Nueva York [ Links ]
LACLAU P (1998) Los ecosistemas forestales y el hombre en el sur de Chile y Argentina. Boletín Técnico 34, Fundación Vida Silvestre Argentina, Buenos Aires [ Links ]
LOISELLE BA Y BLAKE JG (1991) Temporal variation in birds and fruits along an elevational gradient in Costa Rica. Ecology 72:180-193 [ Links ]
LOPEZ DE CASENAVE J (2001) Estructura gremial y organización de un ensamble de aves del desierto del Monte. Tesis doctoral, Universidad de Buenos Aires, Buenos Aires [ Links ]
MARONE L (1992) Seasonal and year-to-year fluctuations of bird populations and guilds in the monte desert, Argentina. Journal of Field Ornithology 63:294-308 [ Links ]
MERMOZ M, ÚBEDA C, GRIGERA D, BRION C, MARTÍN C, BIANCHI E Y PLANAS H (2000) El Parque Nacional Nahuel Huapi: sus características ecológicas y su estado de conservación. Administración de Parques Nacionales y Universidad Nacional del Comahue, San Carlos de Bariloche [ Links ]
MUÑOZ E Y GARAY AF (1985) Caracterización climática de la provincia de Río Negro. INTA, San Carlos de Bariloche [ Links ]
RAITT RJ Y PIMM SL (1976) Dynamics of bird communities in the Chihuahuan Desert, New Mexico. Condor 78:427-442 [ Links ]
RALPH CJ (1985) Habitat association patterns of forest and steppe birds of northern Patagonia, Argentina. Condor 87:141-157 [ Links ]
SOAVE GE, MARATEO G, REY P, GLAZ D Y DARRIEU C (1999) Evolución estacional de los ensambles de aves de un talar del nordeste de la provincia de Buenos Aires, Argentina. Comisión de Investigaciones Científicas, La Plata [ Links ]
ÚBEDA C Y GRIGERA D (1995) Recalificación del estado de conservación de la fauna silvestre argentina, Región Patagónica. Consejo Asesor Regional Patagónico de la Fauna Silvestre, Subsecretaría de Recursos Naturales, Buenos Aires [ Links ]
VILA A (2000) Análisis de la biodiversidad y conservación de la Ecorregión Valdiviana. Memoria del taller, Segunda etapa. Boletín Técnico 52, Fundación Vida Silvestre Argentina, Buenos Aires [ Links ]
VILLAGRÁN C (1991) Historia de los bosques templados del sur de Chile durante el Tardiglacial y Postglacial. Revista Chilena de Historia Natural 64:447-460 [ Links ]
VUILLEUMIER F (1972) Bird species diversity in Patagonian forests (Temperate South America). American Naturalist 106:266-271 [ Links ]
VUILLEUMIER F (1985) Forest birds of Patagonia: ecological geography, speciation, endemism, and faunal history. Ornithological Monographs 36:255-304 [ Links ]
ZAR JH (1999) Biostatistical analysis. Cuarta edición. Prentice-Hall, Upper Saddle River [ Links ]

