










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEl hornero
versión impresa ISSN 0073-3407versión On-line ISSN 1850-4884
Hornero v.20 n.2 Buenos Aires sept./dic. 2005
Éxito reproductivo del Tordo Renegrido (Molothrus bonariensis) en hospedadores de distinto tamaño corporal
Vanina D. Fiorini 1, Andrea A. Astié 1, Diego T. Tuero 1 y Juan C. Reboreda 1,2
1 Departamento de Ecología, Genética y Evolución, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Piso 4, Pabellón 2, Ciudad Universitaria, C1428EHA Buenos Aires, Argentina.
2 reboreda@ege.fcen.uba.ar
Resumen
Se analizaron los distintos componentes del éxito reproductivo del Tordo Renegrido (Molothrus bonariensis) en tres hospedadores que difieren marcadamente en su tamaño corporal: la Ratona Común (Troglodytes musculus), el Zorzal Chalchalero (Turdus amaurochalinus) y la Calandria Grande (Mimus saturninus). Para cada hospedador se estimó la supervivencia de sus nidos y la supervivencia de huevos, el éxito de eclosión y la supervivencia de pichones del parásito en nidos exitosos. En nidos de Troglodytes musculus y Mimus saturninus se analizó la asociación entre el número de huevos presentes en el nido durante la incubación y el éxito de eclosión de los huevos del parásito, y entre el número de pichones del hospedador y la supervivencia de los pichones parásitos. La supervivencia y el éxito de eclosión de los huevos del parásito no difirieron entre hospedadores, pero la supervivencia de sus pichones fue menor en los nidos de Turdus amaurochalinus. El éxito de eclosión estuvo asociado negativamente al número de huevos en los nidos de Troglodytes musculus pero no en los de Mimus saturninus, mientras que la supervivencia de los pichones estuvo asociada negativamente con el número de pichones del hospedador en los nidos de Mimus saturninus pero no en los de Troglodytes musculus. Los resultados de este y de otros estudios muestran que no existe una clara asociación entre el éxito reproductivo de Molothrus bonariensis y el tamaño corporal del hospedador, y señalan la importancia de tener en cuenta otras características de la historia de vida del hospedador además de su tamaño corporal.
Palabras clave: Éxito reproductivo; Molothrus bonariensis; Parasitismo de cría; Tordo Renegrido.
Abstract
Reproductive success of shiny cowbird (Molothrus bonariensis) in hosts of different body size
We analyzed different components of the reproductive success of Shiny Cowbird (Molothrus bonariensis) in three hosts that differ remarkably in body size: the House Wren (Troglodytes musculus), the Creamy-bellied Thrush (Turdus amaurochalinus), and the Chalk-browed Mockingbird (Mimus saturninus). For each host we estimated nest survival, and parasite's egg survival, hatching success and chick survival in successful nests. In Troglodytes musculus and Mimus saturninus nests we also analyzed the association between number of eggs and hatching success of the parasite, and between number of host chicks and chick survival of the parasite. The survival and hatching success of parasite's eggs did not differ between hosts, but chick survival was lower in Turdus amaurochalinus nests. Hatching success was negatively associated with number of eggs in Troglodytes musculus nests but not in those of Mimus saturninus, while chick survival was negatively associated with number of host chicks in Mimus saturninus nests but not in those of Troglodytes musculus. These results, as well as the ones of other studies, show that there is no clear association between the reproductive success of Molothrus bonariensis and the body size of the host and point out the importance of considering other host life history traits in addition to body size.
Key words: Brood parasitism; Molothrus bonariensis; reproductive success; Shiny Cowbird.
Recibido 14 enero 2005, aceptado 27 septiembre 2005
El Tordo Renegrido (Molothrus bonariensis) es un parásito de cría extremadamente generalista en cuanto a la selección de sus hospedadores. Los huevos de este parásito han sido encontrados en nidos de 243 especies y al menos 88 de ellas crían con éxito a sus pichones (Friedmann y Kiff 1985, Ortega 1998, Lowther y Post 1999, Fraga 2002, Lowther 2004). Si bien existen numerosos trabajos que presentan datos sobre la frecuencia e intensidad de parasitismo de Molothrus bonariensis, y en muchos de ellos se analiza el impacto de esta especie sobre el éxito reproductivo de sus hospedadores (se pueden ver las revisiones de Ortega 1998 y Reboreda et al. 2003), muy pocos estudios tienen información detallada del éxito reproductivo del parásito. Conocer el éxito reproductivo de Molothrus bonariensis cuando parasita a sus principales hospedadores resulta necesario para poder entender la dinámica poblacional del parásito y, de esta forma, poder predecir el impacto que éste puede producir sobre las poblaciones de especies hospedadoras (May y Robinson 1985, Wiley et al. 1991, Trine et al. 1998, Ney-Nifle et al. 2005).
El éxito reproductivo de un parásito de cría depende principalmente de: (1) la supervivencia de los nidos del hospedador y (2) la supervivencia de los huevos, el éxito de eclosión y la supervivencia de los pichones parásitos en nidos exitosos (i.e., en nidos en donde no hubo predación ni fueron abandonados). La mayoría de los estudios que han analizado el éxito reproductivo de Molothrus bonariensis presenta información sobre la proporción de huevos puestos por el parásito que llegan al estadio de volantones (Ortega 1998). Estos trabajos no discriminan entre las pérdidas que son consecuencia de la predación o del abandono del nido del hospedador y aquellas que resultan de la destrucción o falla de eclosión de los huevos o de la muerte de los pichones parásitos en nidos exitosos (i.e., los que produjeron volantones del hospedador). Otra aproximación para estimar el éxito reproductivo del parásito ha sido la utilizada por Mason (1986a, 1986b). Este autor evaluó la calidad de una comunidad de hospedadores de Molothrus bonariensis a partir de la supervivencia de sus nidos, si los hospedadores rechazaban o no huevos del parásito y si presentaban una dieta apropiada para sus pichones (i.e., que incluya proteínas animales). A partir de este análisis, Mason concluyó que el éxito reproductivo de Molothrus bonariensis sería mayor en hospedadores de tamaño corporal mayor que el parásito (i.e., mayores de 45-50 g).
Uno de los inconvenientes de suponer que el éxito reproductivo del parásito es función de la supervivencia de los nidos del hospedador es que ésta puede modificarse como resultado de la transformación del ambiente (Robinson et al. 1995) o puede diferir marcadamente entre ambientes (Mermoz y Reboreda 1998). Por el contrario, la supervivencia de los huevos, su éxito de eclosión y la supervivencia de los pichones del parásito dependen mayormente de las características del hospedador, por lo que sería de esperar que, para un determinado hospedador, estos componentes del éxito reproductivo del parásito no difieran mayormente entre ambientes. Por lo antes expuesto, al evaluar el éxito reproductivo del parásito cuando parasita un determinado hospedador resulta importante estimar en forma independiente la supervivencia de sus nidos y la supervivencia de huevos y pichones del parásito en nidos exitosos.
Existen distintos factores que pueden afectar la supervivencia de los huevos parásitos, su éxito de eclosión y la supervivencia de los pichones. En relación a la supervivencia de los huevos, las hembras de Molothrus bonariensis pican los huevos del hospedador y eventualmente los de otras hembras parásitas cuando visitan y parasitan los nidos (Hudson 1874, Höy y Ottow 1964). Este comportamiento, junto con el rechazo de huevos parásitos por parte de algunos hospedadores, sería la principal causa de pérdida de huevos de Molothrus bonariensis en nidos en donde no hubo predación ni abandono (Mermoz y Reboreda 2003). Por lo tanto, se espera que la supervivencia de los huevos del parásito esté asociada negativamente con la intensidad de parasitismo sobre el hospedador.
El éxito de eclosión de los huevos del parásito puede verse reducido por una mala sincronización del parasitismo con la puesta de huevos del hospedador (los huevos puestos cuando la incubación está avanzada no tendrían tiempo suficiente para completar su desarrollo; Fraga 1998) y por un corto período de incubación de los huevos del hospedador (los huevos parásitos tendrían una menor probabilidad de completar su desarrollo). El éxito de eclosión de los huevos del parásito también podría disminuir si los huevos del parásito y los del hospedador difieren en tamaño (Scott y Lemon 1996) o si el número de huevos en el nido es muy numeroso (Peer y Bollinger 1997).
La supervivencia de los pichones parásitos depende de que el alimento que entregan los hospedadores sea apropiado para el pichón parásito (Middleton 1991, Kozlovic et al. 1996). También puede influir que los pichones del hospedador tengan un mayor tamaño corporal o que éstos nazcan antes que los pichones del parásito (Scott y Lemon 1996). Otro factor que podría afectar negativamente la supervivencia de los pichones parásitos es el número de pichones del hospedador con los que debe competir por alimento. Sin embargo, Kilner (2003) y Kilner et al. (2004) han planteado que la presencia de pichones del hospedador podría aumentar la tasa de entrega de alimento al nido por parte de los padres y, de esta forma, beneficiar al pichón parásito.
El objetivo de este trabajo es analizar la supervivencia y el éxito de eclosión de los huevos y la supervivencia de los pichones de Molothrus bonariensis en tres hospedadores que difieren marcadamente en su tamaño corporal (Tabla 1): la Ratona Común (Troglodytes musculus) (= Troglodytes aedon musculus; Brumfield y Capparella 1996), el Zorzal Chalchalero (Turdus amaurochalinus) y la Calandria Grande (Mimus saturninus). Para Troglodytes musculus y Mimus saturninus se analiza la asociación entre el número de huevos presentes en el nido durante la incubación y el éxito de eclosión de los huevos del parásito, y entre el número de pichones del hospedador y la supervivencia de los pichones parásitos. Además, se comparan los resultados con los de estudios previos del éxito reproductivo de Molothrus bonariensis parasitando otros hospedadores.
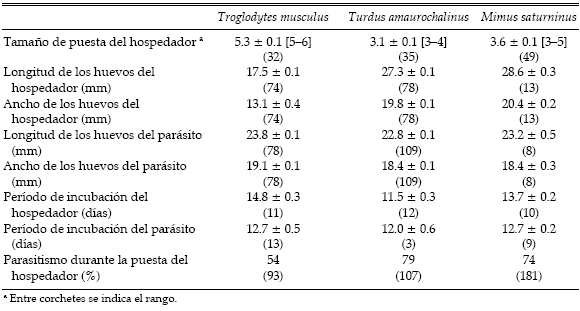
Tabla 1. Eficiencia (Ef) y éxito reproductivo (Ex) de Molothrus bonariensis cuando parasita diferentes hospedadores. Para cada hospedador se indican el peso corporal (PC, en gramos) y la frecuencia (Fr) e intensidad (In) de parasitismo. Se muestran los siguientes componentes del éxito reproductivo del parásito: supervivencia de nidos (SN), supervivencia de huevos (SH), éxito de eclosión (EE) y supervivencia de pichones (SP). Además, se indica el peso de los pichones parásitos antes de abandonar el nido (PP). Para la supervivencia de huevos y de pichones, el éxito de eclosión y el peso de los pichones se muestran el promedio ± EE, con el número de nidos utilizados para su cálculo entre paréntesis. Las variables están definidas en Métodos, Análisis de datos.

Métodos
Sitios de estudio
Los estudios de Troglodytes musculus y Mimus saturninus se realizaron en la Reserva Privada de Flora y Fauna Autóctona “El Destino”, ubicada en el partido de Magdalena, provincia de Buenos Aires (35°08'S, 57°23'O). La recolección de datos para Mimus saturninus abarcó las temporadas reproductivas (octubre-enero) de 2002-2003 y 2003-2004, mientras que para Troglodytes musculus se realizó durante la temporada reproductiva 2003-2004. El estudio de Turdus amaurochalinus se realizó en el departamento de Guaymallén, provincia de Mendoza (32°51'S, 68°42'O), durante las temporadas reproductivas (octubre-diciembre) de 1999-2002.
Recolección de datos
Los datos de Troglodytes musculus se obtuvieron en nidos construidos en cajas-nido de madera. Las cajas-nido tenían dimensiones internas de 23×14×10 cm (altura, ancho y profundidad) y fueron colocadas en talas (Celtis tala) a una altura de 1.5-2.0 m antes del inicio de la temporada reproductiva (ver más detalles en Tuero 2004). Los datos de Mimus saturninus fueron obtenidos en nidos construidos en talas (Celtis tala), coronillos (Scutia buxifolia) y molles (Schinus longifolius) a una altura de 1.5-2.5 m. Los datos de Turdus amaurochalinus fueron obtenidos en nidos construidos principalmente en vides y olivos a una altura de 1.5-4.5 m (ver más detalles en Astié 2004). Los nidos de Mimus saturninus y Turdus amaurochalinus fueron ubicados a través de una búsqueda sistemática en sitios en los que individuos adultos de estas especies emitían cantos de alarma.
Los nidos de cada especie fueron revisados regularmente cada 1-3 días desde el momento en que fueron encontrados hasta que produjeron volantones o fueron abandonados o predados. En cada visita se registró el número de huevos o pichones del hospedador y del parásito. A medida que los huevos eran puestos fueron medidos con un calibre (error: ±0.1 mm) y rotulados con tinta indeleble para permitir su seguimiento individual. Los pichones fueron marcados con tinta indeleble en el tarso y a los 7-9 días de edad se les colocó una combinación única de anillos de colores en sus tarsos. Los pichones fueron pesados regularmente con una balanza digital portátil (error: 0.1 g) en el caso de Troglodytes musculus y con balanzas de resorte de 10 y 50 g (error: 0.1 y 0.5 g, respectivamente) en el caso de Mimus saturninus y Turdus amaurochalinus.
Análisis de datos
La frecuencia de parasitismo (proporción de nidos que recibieron al menos un huevo parásito) y su intensidad (número promedio de huevos parásitos por nido parasitado) se estimaron a partir de una muestra de nidos encontrados en construcción o puesta que completaron la puesta. Se utilizó este criterio ya que: (1) la mayoría de los eventos de parasitismo de Molothrus bonariensis ocurren durante la puesta del hospedador (Massoni y Reboreda 1998, Mermoz y Reboreda 1999), y (2) la inclusión de nidos en donde hubo predación o que fueron abandonados durante el estadio de puesta podría producir una subestimación de la frecuencia de parasitismo, ya que estos nidos no sobrevivieron el tiempo suficiente como para ser parasitados.
Para estimar el tamaño de los huevos de cada hospedador se calculó la longitud y el ancho promedio de los huevos de cada nido y luego se promediaron estos valores (se consideraron a los nidos como datos independientes). En el caso de los huevos del parásito se consideró que cada huevo fue independiente, ya que no pudieron ser asignados a una hembra determinada.
El período de incubación del hospedador se estimó como el tiempo transcurrido desde la puesta del último huevo hasta la eclosión del último huevo (Nice 1954). Para estimar el período de incubación del parásito se siguió el siguiente criterio: (1) en los casos en los que el huevo fue puesto antes de la puesta del penúltimo huevo del hospedador, se estimó como el tiempo transcurrido desde la puesta del penúltimo huevo del hospedador (inicio de la incubación) hasta la eclosión del huevo del parásito (Briskie y Sealy 1990); y (2) en los casos en los que el huevo del parásito fue puesto luego de iniciada la incubación, se estimó como el tiempo transcurrido desde la puesta del huevo del parásito hasta su eclosión. El período de incubación de los hospedadores y del parásito se determinó a partir de una muestra de nidos que fueron visitados diariamente.
El porcentaje de parasitismo durante la puesta del hospedador se estimó como el cociente entre el número de huevos parásitos puestos durante este período y el número total de huevos puestos. Se consideró un período de puesta de cinco días para Troglodytes musculus, tres días para Turdus amaurochalinus y cuatro días para Mimus saturninus.
Para estimar la eficiencia reproductiva del parásito (proporción de huevos que llegan al estadio de volantón en nidos exitosos) se analizó la supervivencia de los huevos del parásito, su éxito de eclosión y la supervivencia de sus pichones en nidos exitosos (i.e., los que produjeron al menos un volantón del hospedador o del parásito). Para estimar el éxito reproductivo total del parásito se calculó el producto de la eficiencia reproductiva y la supervivencia de los nidos del hospedador. Este es un estimador de la proporción de huevos parásitos puestos en nidos del hospedador que llegan al estadio de volantones, incluyendo tanto los efectos de predación y abandono de nidos como las pérdidas de huevos y pichones en nidos exitosos. La supervivencia de los nidos de cada hospedador se estimó como la proporción de nidos encontrados durante los estadios de construcción y puesta que produjeron al menos un volantón del hospedador o del parásito.
La supervivencia de huevos del parásito se estimó como el cociente entre el número de huevos del parásito que estaban presentes en el nido al final del período de incubación y el número de huevos parásitos puestos en el nido. Para el cálculo de esta variable se utilizaron los nidos encontrados en construcción y puesta que completaron la incubación. El éxito de eclosión se estimó como el cociente entre el número de huevos del parásito que eclosionaron y el número de huevos del parásito presentes en el nido al final del período de incubación. Para el cálculo de esta variable se utilizaron los nidos encontrados en construcción, puesta e incubación que completaron la incubación. La supervivencia de pichones del parásito se estimó como el cociente entre el número de pichones del parásito que abandonaron el nido exitosamente y el número de pichones parásitos que eclosionaron. Para el cálculo de esta variable se utilizaron los nidos encontrados en construcción, puesta e incubación que produjeron volantones. Se consideró que los pichones del parásito que se encontraban en el nido 1-2 días antes de la fecha en que los pichones del hospedador abandonaron el nido (12 días en Turdus amaurochalinus y Mimus saturninus y 15 días en Troglodytes musculus) fueron exitosos. En todos los casos observados estos pichones presentaban un desarrollo completo del plumaje y valores asintóticos de peso corporal.
Análisis del éxito reproductivo con diferentes hospedadores
Se compararon los distintos componentes del éxito reproductivo de Molothrus bonariensis cuando parasita diferentes hospedadores. Para ello se utilizaron los datos obtenidos en este estudio y datos de estudios previos realizados con Agelaius thilius (Massoni y Reboreda 1998, Massoni 2002), Mimus saturninus (Sackmann y Reboreda 2003), Turdus rufiventris (Lichtenstein 1998, Sackmann y Reboreda 2003) y Pseudoleistes virescens (Mermoz y Reboreda 2003). En todos los estudios la subespecie estudiada fue Molothrus bonariensis bonariensis (hembras: 40-45 g, machos: 50-55 g). Solo se tuvieron en cuenta estudios realizados en el área de distribución histórica de Molothrus bonariensis y en los que se pudo estimar en forma independiente la supervivencia de los nidos del hospedador y la supervivencia de huevos, éxito de eclosión y supervivencia de pichones parásitos en nidos exitosos. Para este último componente del éxito reproductivo sólo se tuvieron en cuenta estudios en los que fueron exitosos al menos seis nidos en los que nacieron pichones del parásito.
Análisis estadístico
Debido al reducido tamaño muestral de algunos grupos y a la falta de normalidad de los datos se utilizó estadística no paramétrica (Siegel y Castellan 1988). En el texto, los datos se presentan como promedios ± EE.
Resultados
Éxito reproductivo con distintos hospedadores
En la tabla 1 se presentan los datos de frecuencia e intensidad de parasitismo y de los distintos componentes del éxito reproductivo del parásito en nidos de Troglodytes musculus, Turdus amaurochalinus y Mimus saturninus. En la misma tabla se presentan datos similares para otras especies de hospedadores estudiadas previamente por otros autores.
La intensidad de parasitismo difirió significativamente entre los tres hospedadores estudiados, siendo mayor en Mimus saturninus que en Turdus amaurochalinus (Prueba de Kruskal- Wallis, H = 12.4, P = 0.002 y contrastes a posteriori, P < 0.05). Sin embargo, no se observaron diferencias entre hospedadores en la supervivencia de los huevos del parásito (H = 4.4, P = 0.11). Tampoco se observaron diferencias entre hospedadores en el éxito de eclosión de los huevos del parásito (H = 2.6, P = 0.27).
El período de incubación de los huevos del parásito fue similar en los tres hospedadores, pero el porcentaje de eventos de parasitismo sincronizados con la puesta del hospedador fue menor en nidos de Troglodytes musculus que en los de Turdus amaurochalinus y Mimus saturninus (Prueba de contingencia, χ2= 12.3, P = 0.003; Tabla 2). Sin embargo, si se tiene en cuenta que los huevos de Molothrus bonariensis tuvieron un período de incubación de dos días menos que los de Troglodytes musculus (Tabla 2), el 81% (75/93) de los huevos del parásito fue puesto de forma tal que su eclosión habría ocurrido con anterioridad o en forma simultánea a la eclosión del primer huevo del hospedador. Si se utiliza este criterio de sincronización del parasitismo, no se registraron diferencias significativas entre hospedadores (χ2= 1.6, P = 0.44).
Tabla 2. Parámetros reproductivos de los tres hospedadores estudiados y de Molothrus bonariensis en nidos de estos hospedadores. Para cada variable se muestran el promedio ± EE y, entre paréntesis, el número de nidos (o huevos en el caso del tamaño de los huevos del parásito y del porcentaje de parasitismo) utilizados para su cálculo.
La supervivencia de los pichones parásitos fue significativamente menor en los nidos de Turdus amaurochalinus que en los de los otros dos hospedadores (H = 21.9, P < 0.001; contrastes a posteriori, P < 0.05).
Número de huevos y éxito de eclosión
Se analizó la asociación entre el número de huevos presentes en el nido durante el período de incubación y el éxito de eclosión de los huevos del parásito. Este análisis se realizó en Troglodytes musculus y Mimus saturninus, los hospedadores con tamaños corporales extremos. El número de huevos del hospedador presentes en el nido durante la incubación fue 4.0 ± 0.2 (rango: 2-6, n = 30) en Troglodytes musculus, y 2.3 ± 0.2 (rango: 0-4, n = 39) en Mimus saturninus. En ambas especies el éxito de eclosión de los huevos del parásito no estuvo asociado al número de huevos del hospedador (Correlación de Spearman; rS= 0.07, Z = 0.37, P = 0.71, n = 30 para Troglodytes musculus; rS= 0.22, Z = 1.35, P = 0.18, n = 39 para Mimus saturninus). Tampoco se observó una asociación significativa entre el número total de huevos presentes (i.e., hospedador más parásito) y el éxito de eclosión de los huevos del parásito en nidos de Mimus saturninus (número de huevos: 4.4 ± 0.2, rango: 1-8, rS= -0.16, Z = -0.97, P = 0.32, n = 39), pero esta asociación fue significativa en nidos de Troglodytes musculus (número de huevos: 5.8 ± 0.2, rango: 3-8, rS= -0.36, Z = -1.94, P = 0.05, n = 30).
Número de pichones y supervivencia
La supervivencia de los pichones del parásito no estuvo asociada al número de pichones de Troglodytes musculus (rS= 0.26, Z = 1.10, P = 0.29, n = 18; Fig. 1A) ni al número total de pichones en el nido (rS= 0.05, Z = 0.19, P = 0.85, n = 18). Por el contrario, en nidos de Mimus saturninus se observó una asociación negativa entre el número de pichones del hospedador y la supervivencia de los pichones del parásito (rS= -0.63, Z = -2.81, P = 0.005, n = 21; Fig. 1B).
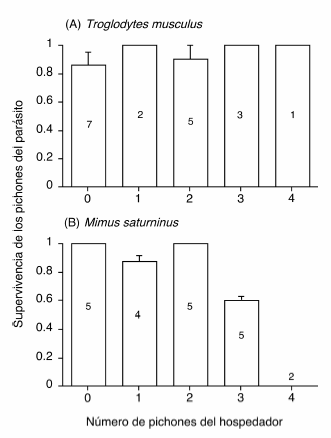
Figura 1. Supervivencia promedio (+ EE) de los pichones de Molothrus bonariensis en función del número de pichones que eclosionaron del hospedador cuando éste era Troglodytes musculus (A) y Mimus saturninus (B). Los números en las barras indican el número de nidos.
Discusión
Los resultados obtenidos muestran que la supervivencia de los huevos de Molothrus bonariensis fue similar en los tres hospedadores analizados. Esta ausencia de diferencias resulta llamativa, ya que los hospedadores estudiados tuvieron distintas intensidades de parasitismo y, como se mencionó más arriba, se espera que la supervivencia de los huevos del parásito esté asociada negativamente con la intensidad de parasitismo del hospedador. Si bien el número de especies analizadas es pequeño, nuestros resultados y los de estudios realizados en otros hospedadores (Tabla 1) no sustentan esta hipótesis. Una posible explicación para la ausencia de esta asociación es que la picadura afecte preferentemente a los huevos del hospedador. Esto fue observado previamente por Mermoz y Reboreda (1999) y podría deberse a que el mayor espesor de la cáscara de los huevos parásitos (Mermoz y Ornelas 2004) los haga más resistentes a las picaduras, o a que las hembras parásitas dirijan su comportamiento de picaduras preferentemente hacia los huevos del hospedador. Esto último se esperaría en el caso en que la hembra parásita ponga más de un huevo en el mismo nido, como ocurre en Molothrus ater (McLaren et al. 2003), ya que de esta forma sería menos probable que destruya sus propios huevos. En concordancia con esta idea, Soler et al. (1999) hallaron que Clamator glandarius no pica ni remueve los huevos del parásito presentes en el nido, y sugirieron que esto podría deberse a que las hembras de esta especie frecuentemente ponen dos huevos en el mismo nido.
En los tres hospedadores estudiados la mayoría de los huevos del parásito fueron puestos en forma sincrónica con los huevos del hospedador. Además, la duración del período de incubación y el éxito de eclosión de los huevos de Molothrus bonariensis no difirió entre hospedadores. Payne (1977) planteó que en aquellos casos en que el huevo del parásito es de menor tamaño que el del hospedador no habría un buen contacto entre éste y el parche de incubación y, por lo tanto, disminuiría la eficiencia de incubación. Ese mismo argumento fue utilizado por Wiley (1985) para explicar el menor éxito de eclosión de los huevos de Molothrus bonariensis en nidos de Quiscalus niger. Sin embargo, Soler (1990) observó un alto éxito de eclosión de los huevos de Clamator glandarius en nidos de Corvus corone aun cuando los huevos de este hospedador duplican en volumen a los del parásito. Nuestros resultados son coincidentes con los de Soler (1990) e indican que la diferencia de tamaño entre los huevos del hospedador y de Molothrus bonariensis no afectaría en forma significativa la eficiencia de incubación de estos últimos. El éxito de eclosión de los huevos de Molothrus bonariensis sólo se vio afectado por el número total de huevos presentes en el nido (hospedador más parásito) en el caso del hospedador más pequeño estudiado (Troglodytes musculus). Sin embargo, debido a que en esta especie la frecuencia de nidos con muchos huevos es baja, el éxito de eclosión promedio de los huevos del parásito no difirió con respecto al observado cuando parasitaba los otros hospedadores.
La supervivencia de los pichones de Molothrus bonariensis fue similar en los nidos de Troglodytes musculus y Mimus saturninus (hospedadores que tienen tamaños corporales considerablemente menor y mayor al del parásito, respectivamente), pero fue bastante menor en los nidos de Turdus amaurochalinus (de tamaño corporal levemente mayor al del parásito). La menor supervivencia de los pichones de Molothrus bonariensis en nidos de Turdus amaurochalinus se debió probablemente a que este hospedador presenta un conjunto de características que resultan particularmente desfavorables para el parásito (Astié 2004). Estas características son: (1) inicio de la incubación con la puesta del segundo huevo y período de incubación más corto que el del parásito (aun cuando el parasitismo esté sincronizado con la puesta del hospedador, los pichones del hospedador nacen antes o simultáneamente con los del parásito); (2) al nacer, los pichones del hospedador tienen un mayor tamaño corporal que los del parásito; y (3) con frecuencia ocurre reducción de la nidada, y ésta afecta al pichón de menor tamaño (que, en nidos parasitados, es el pichón parásito).
Los valores de supervivencia de los pichones de Molothrus bonariensis en nidos de Turdus amaurochalinus fueron los más bajos observados para esta especie en los hospedadores estudiados hasta ahora (Tabla 1). Los únicos valores similares fueron los observados por Lichtenstein (1998) en nidos de Turdus rufiventris. Esta autora postuló que el bajo éxito de Molothrus bonariensis en nidos de Turdus rufiventris se debería a que en esta especie hospedadora los padres discriminan activamente en la entrega de alimento entre sus pichones y los de Molothrus bonariensis (Lichtenstein 2001). En nuestro estudio no se analizó si los padres de Turdus amaurochalinus discriminaban entre sus pichones y los del parásito. Sin embargo, teniendo en cuenta que en este hospedador el pichón parásito fue siempre el de menor tamaño en el nido y que sobrevivió solamente cuando estaba solo o con un pichón del hospedador (Astié 2004), resulta más parsimonioso atribuir la baja supervivencia a la asimetría en la competencia por alimento con los pichones del hospedador. Resultados similares fueron obtenidos por Soler (1990), quien observó que la supervivencia de los pichones de Clamator glandarius fue muy baja en un hospedador de mayor tamaño corporal (Corvus corone). Este autor atribuyó esta menor supervivencia a una competencia más intensa por alimento entre los pichones del parásito y los del hospedador, quienes tempranamente superaban en tamaño corporal a los del parásito. En forma semejante, la asociación negativa entre la supervivencia de los pichones del parásito y el número de pichones del hospedador en nidos de Mimus saturninus pero no en los de Troglodytes musculus indica que las diferencias de tamaño corporal entre los pichones del hospedador y del parásito son importantes bajo condiciones de alta competencia por alimento en el nido. Este resultado también muestra que el comportamiento de picadura de huevos de Molothrus bonariensis aumentaría la supervivencia de los pichones del parásito (y, por lo tanto, su éxito reproductivo) en hospedadores de mayor tamaño corporal.
En un estudio comparativo de distintos hospedadores de Molothrus ater, Kilner (2003) observó que los pichones parásitos tenían mayor probabilidad de sobrevivir cuando estaban en nidos de hospedadores de tamaño intermedio y compartían el nido con uno o dos pichones del hospedador. Esta autora postuló que estos resultados podrían deberse a que los pichones del hospedador ayudarían a aumentar la tasa de pedido de alimento. Sin embargo, es interesante resaltar que salvo en cuatro casos, todos los hospedadores con los que trabajaron eran de un tamaño corporal menor al del parásito.
Al comparar los resultados de los estudios en los que se analizó en forma independiente la supervivencia de los nidos del hospedador y la supervivencia de huevos, éxito de eclosión de huevos y supervivencia de pichones del parásito en nidos exitosos (Tabla 1), surge que la supervivencia de los nidos del hospedador explica la mayor parte de la variación en el éxito reproductivo de Molothrus bonariensis. Como se mencionó más arriba, la supervivencia de los nidos varía considerablemente entre ambientes, por lo que inferir el éxito reproductivo del parásito (y, por lo tanto, la calidad del hospedador) a partir de un único estudio puede llevar a conclusiones erróneas. De la tabla 1 surge también que la supervivencia y el éxito de eclosión de los huevos del parásito en los nidos de los hospedadores estudiados son similares (0.76-0.97 para la supervivencia de huevos y 0.62-0.82 para el éxito de eclosión) y que, a excepción de Turdus amaurochalinus y de uno de los estudio realizados en Turdus rufiventris (Lichtenstein 1998), la supervivencia de los pichones tampoco difiere mucho entre hospedadores (0.62-0.89). El peso asintótico de los pichones de Molothrus bonariensis que abandonaron exitosamente el nido de sus hospedadores tampoco muestra una clara asociación con el tamaño corporal del hospedador (Tabla 1).
Los resultados presentados en este trabajo, así como los de estudios previos (Lichtenstein 1998, Massoni y Reboreda 1998, Mermoz y Reboreda 2003, Sackman y Reboreda 2003), muestran que no existe una clara asociación entre el éxito reproductivo del parásito y el tamaño corporal del hospedador. Por el contrario, estos estudios muestran que el éxito reproductivo de Molothrus bonariensis es relativamente similar en hospedadores muy distintos y plantean la importancia de tener en cuenta otras características de la historia de vida del hospedador además de su tamaño corporal.
Agradecimientos. Agradecemos a dos revisores por las sugerencias realizadas a una versión previa del manuscrito. Durante la realización de este trabajo Andrea A. Astié y Vanina D. Fiorini fueron becarias de posgrado del Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) y Diego T. Tuero fue becario doctoral de la Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT). Juan C. Reboreda es miembro de la Carrera del Investigador Científico del CONICET. Este trabajo fue financiado mediante un subsidio de Neotropical Grassland Conservancy otorgado a Vanina D. Fiorini y subsidios de la Universidad de Buenos Aires (X158) y de la ANPCyT (01-09237) otorgados a Juan C. Reboreda.
Bibliografía citada
ASTIÉ AA (2004) Interacciones entre el parásito de cría Molothrus bonariensis y uno de sus hospedadores, Turdus amaurochalinus. Tesis doctoral, Universidad de Buenos Aires, Buenos Aires [ Links ]
BRISKIE JV Y SEALY SG (1990) Evolution of short incubation periods in the parasitic cowbirds, Molothrus sp. Auk 107:789-794 [ Links ]
BRUMFIELD RT Y CAPPARELLA AP (1996) Genetic differentiation and taxonomy in the House Wren species group. Condor 98:547-556 [ Links ]
FRAGA RM (1998) Interactions of the parasitic screaming and shiny cowbirds (Molothrus rufoaxillaris and M. bonariensis) with a shared host, the bay-winged cowbird (Molothrus badius). Pp. 173-193 en: ROTHSTEIN SI Y ROBINSON SK (eds) Parasitic birds and their hosts: studies in coevolution. Oxford University Press, Nueva York [ Links ]
FRAGA RM (2002) Notes on new or rarely reported Shiny Cowbird hosts from Argentina. Journal of Field Ornithology 73:213-219 [ Links ]
FRIEDMANN H Y KIFF LF (1985) The parasitic cowbirds and their hosts. Proceedings of the Western Foundation of Vertebrate Zoology 2:226-304 [ Links ]
HÖY G Y OTTOW J (1964) Biological and oological studies of the molothrine cowbirds (Icteridae) of Argentina. Auk 81:186-203 [ Links ]
HUDSON WH (1874) Notes on the procreant instincts of the three species of Molothrus found in Buenos Aires. Proceedings of the Zoological Society 11:153-174 [ Links ]
KILNER RM (2003) How selfish is a cowbird nestling? Animal Behaviour 66:569-576 [ Links ]
KILNER RM, MADDEN JR Y HAUBER ME (2004) Brood parasitic cowbird nestlings use host young to procure resources. Science 305:877-879 [ Links ]
KOZLOVIC DR, KNAPTON RW Y BARLOW JC (1996) Unsuitability of the House Finch as a host of the Brownheaded Cowbird. Condor 98:253-258 [ Links ]
LICHTENSTEIN G (1998) Parasitism by Shiny Cowbirds of Rufous-bellied Thrushes. Condor 100:680-687 [ Links ]
LICHTENSTEIN G (2001) Low success of shiny cowbird chicks parasitizing rufous-bellied thrushes: chick- chick competition or parental discrimination? Animal Behaviour 61:401-413 [ Links ]
LOWTHER PE (2004) Molothrine cowbird host list. Field Museum, Chicago (URL: http://fm1.fieldmuseum.org/ aa/Files/lowther/CBList.pdf) [ Links ]
LOWTHER PE Y POST W (1999) Shiny Cowbird (Molothrus bonariensis). Pp. 1-24 en: POOLE A Y GILL F (eds) The birds of North America. Academy of Natural Sciences y American Ornithologists' Union, Philadelphia y Washington DC [ Links ]
MASON P (1986a) Brood parasitism in a host generalist, the Shiny Cowbird (Molothrus bonariensis): I. The quality of different species as hosts. Auk 103:52-60 [ Links ]
MASON P (1986b) Brood parasitism in a host generalist, the Shiny Cowbird (Molothrus bonariensis): II. Host selection. Auk 103:61-69 [ Links ]
MASSONI V (2002) Parasitismo de cría del Tordo Renegrido sobre el Varillero Ala Amarilla. Tesis doctoral, Universidad de Buenos Aires, Buenos Aires [ Links ]
MASSONI V Y REBOREDA JC (1998) Costs of brood parasitism and the lack of defenses on the Yellowwinged Blackbird-Shiny Cowbird system. Behavioral Ecology and Sociobiology 42:273-280 [ Links ]
MAY RM Y ROBINSON SK (1985) Population dynamics of avian brood parasitism. American Naturalist 130:161-167 [ Links ]
MAYFIELD H (1975) Suggestions for calculating nest success. Wilson Bulletin 87:456-466 [ Links ]
MCLAREN CM, WOOLFENDEN BE, GIBBS HL Y SEALY SG (2003) Genetic and temporal patterns of multiple parasitism by Brown-headed Cowbirds (Molothrus ater) on Song Sparrows (Melospiza melodia). Canadian Journal of Zoology 81:281-286 [ Links ]
MERMOZ ME Y ORNELAS JF (2004) Phylogenetic analysis of life-history adaptations in parasitic cowbirds. Behavioral Ecology 15:109-119 [ Links ]
MERMOZ ME Y REBOREDA JC (1998) Nesting success in Brown-and-yellow Marshbirds: effects of time of the breeding season, nest site and brood parasitism. Auk 115:871-878 [ Links ]
MERMOZ ME Y REBOREDA JC (1999) Egg laying behavior by Shiny Cowbirds parasitizing Brown-andyellow Marshbirds. Animal Behaviour 58:873-882 [ Links ]
MERMOZ ME Y REBOREDA JC (2003) Reproductive success of Shiny Cowbird (Molothrus bonariensis) parasitizing the larger Brown-and-yellow Marshbird (Pseudoleistes virescens). Auk 120:1128-1139 [ Links ]
MIDDLETON ALA (1991) Failure of Brown-headed Cowbird parasitism in nests of the American Goldfinch. Journal of Field Ornithology 62:200-203 [ Links ]
NEY-NIFLE M, BERNSTEIN C, REBOREDA JC Y KACELNIK A (2005) Population dynamics and avian brood parasitism: persistence and invasions in a three species system. Journal of Animal Ecology 74:274-284 [ Links ]
NICE MM (1954) Problems of incubation periods in North American birds. Condor 56:173-197 [ Links ]
ORTEGA CP (1998) Cowbirds and other brood parasites. University of Arizona Press, Tucson [ Links ]
PAYNE RB (1977) The ecology of brood parasitism in birds. Annual Review of Ecology and Systematics 8:1-28 [ Links ]
PEER BD Y BOLLINGER EK (1997) Explanations for the infrequent cowbird parasitism on Common Grackles. Condor 99:151-161 [ Links ]
REBOREDA JC, MERMOZ ME, MASSONI V, ASTIÉ AA Y RABUFFETTI FL (2003) Impacto del parasitismo de cría del Tordo Renegrido (Molothrus bonariensis) sobre el éxito reproductivo de sus hospedadores. Hornero 18:77-88 [ Links ]
ROBINSON SK, THOMSON FR III, DONOVAN TM, WHITEHEAD DR Y FAABORG J (1995) Regional forest fragmentation and the nesting success of migratory birds. Science 267:1987-1990 [ Links ]
SACKMANN P Y REBOREDA JC (2003) A comparative study of Shiny Cowbird parasitism in two large hosts: Chalk-browed Mockingbird and Rufous-bellied Thrush. Condor 105:728-736 [ Links ]
SCOTT DM Y LEMON RE (1996) Differential reproductive success of Brown-headed Cowbirds with Northern Cardinals and three other hosts. Condor 98:259-271 [ Links ]
SIEGEL S Y CASTELLAN NJ (1988) Nonparametric statistics for the behavioral sciences. Segunda edición. McGraw-Hill, Singapur [ Links ]
SOLER M (1990) Relationships between the Great spotted Cuckoo Clamator glandarius and its corvid hosts in a recently colonized area. Ornis Scandinavica 21:212-223 [ Links ]
SOLER M, SOLER JJ Y PÉREZ-CONTRERAS T (1999) The cost of host egg damage caused by a brood parasite: experiments on Great spotted Cuckoo (Clamator glandarius) and magpies (Pica pica). Behavioral Ecology and Sociobiology 46:381-386 [ Links ]
TRINE CL, ROBINSON WD Y ROBINSON SK (1998) Consequences of Brown-headed Cowbird parasitism for host population dynamics. Pp. 273-295 en: ROTHSTEIN SI Y ROBINSON SK (eds) Parasitic birds and their hosts: studies in coevolution. Oxford University Press, Nueva York [ Links ]
TUERO DT (2004) Impacto del parasitismo de cría del Tordo Renegrido (Molothrus bonariensis) sobre el éxito reproductivo de un hospedador de pequeño tamaño, la Ratona Común (Troglodytes aedon). Tesis de Licenciatura, Universidad de Buenos Aires, Buenos Aires [ Links ]
WILEY JW (1985) Shiny Cowbird Molothrus bonariensis parasitism in two avian communities in Puerto Rico. Condor 87:165-176 [ Links ]
WILEY JW, POST W Y CRUZ A (1991) Conservation of the Yellow-shouldered Blackbird Agelaius xanthomus, an endagered West Indian Species. Biological Conservation 55:119-138 [ Links ]

