










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEl hornero
versión impresa ISSN 0073-3407versión On-line ISSN 1850-4884
Hornero v.21 n.1 Buenos Aires ene./ago. 2006
Escalas en ecología: su importancia para el estudio de la selección de hábitat en aves
Víctor R. Cueto
Grupo de Investigación en Ecología de Comunidades de Desierto (ECODES), Departamento de Ecología, Genética y Evolución, FCEyN, Universidad de Buenos Aires. Pab. 2, Piso 4, Ciudad Universitaria, C1428EHA Buenos Aires, Argentina. vcueto@ege.fcen.uba.ar
Resumen
En este trabajo discuto sobre qué entendemos por escala en ecología y cómo las dimensiones espaciales de nuestro estudio pueden influir en los patrones que observamos y en nuestras conclusiones sobre los procesos que los producen. Analizo la importancia de establecer la escala espacial de nuestros estudios cuando intentamos evaluar la selección de hábitat por las aves, discutiendo los métodos y conclusiones de un trabajo a múltiples escalas sobre la relación entre la vegetación y las aves. Concluyo que comprender la importancia de establecer la escala de nuestro trabajo no solo permite una mejor planificación del proyecto de investigación, sino que facilita la interpretación de los resultados obtenidos y mejora el potencial para encontrar explicaciones a nuestro problema de estudio.
Palabras clave: definición de escala, dominios de la escala, estudios a múltiples escalas, extensión, grano, selección de hábitat, tipos de escala.
Abstract
Scale in ecology: its importance for the study of avian habitat selection
I discuss about the meaning of scale in ecology, and how spatial dimensions of our study influence observed patterns and conclusions about processes producing them. I analyze the importance of establishing the spatial scale in our studies when we evaluate avian habitat selection, and I discuss methods and conclusions of a multiple-scale investigation on bird-vegetation relationships. I conclude that understanding the importance of scale in our investigations allows a better planning of the research project, facilitates the interpretation of the obtained results, and improves the potential to find explanations for the problem under study.
Key words: extent, grain, habitat selection, multiscale studies, scale definition, scale domains, scale types.
El interés de los ecólogos por el efecto de la escala en sus trabajos de investigación ha aumentado notablemente en los últimos tiempos. Antes de la década de 1980 en pocos trabajos se mencionaba explícitamente la escala del estudio, pero desde mediados de esa década se produjo un rápido incremento en las publicaciones que tienen en cuenta los problemas de la escala 1. Por ejemplo, Kareiva y Andersen 2 revisaron 100 experimentos de campo en trabajos de ecología de comunidades y encontraron que la mitad de ellos fueron realizados en parcelas de menos de 1 m de diámetro, a pesar de que había considerables diferencias en el tamaño y los tipos de organismos estudiados. Además, la literatura ecológica de décadas pasadas está llena de estudios en los cuales se colectaban datos a escala de una hectárea o menos y se extrapolaban las conclusiones a escalas mucho más amplias (incluso a todo el planeta). El resultado de tales extrapolaciones puede o no ser válido, pero es razonable reconocer que es posible que frente a esos cambios de escala las fuentes de error y variación involucradas se modifiquen o que cambien las relaciones con los factores ambientales 3. En gran medida la indiferencia de los ecólogos por el problema de la escala en sus estudios podría haberse debido a los siguientes motivos 4: (1) la teoría ecológica fue desarrollada sin tenerla en cuenta, y por lo tanto se puede aplicar en cualquier ámbito espacio-temporal; (2) los fenómenos con los que suelen trabajar los ecólogos son intuitivamente familiares y probablemente los perciben y estudian desde escalas antropocéntricas que concuerdan con su propia experiencia; y (3) existe una tradición de uso de cuadrantes y parcelas de un tamaño particular simplemente porque fueron los usados previamente por otros investigadores.
Los patrones y los procesos ecológicos que los producen son dependientes de la escala espacial y temporal sobre la cual son observados 4. Sin embargo, no se puede decir que un proceso esté restringido a una escala particular, sino que existen escalas de tiempo y espacio en las cuales un proceso resulta relevante o prevalece sobre otros 1. Por ejemplo, la hipótesis que propone que los procesos de especiación y extinción pueden haber contribuido a generar patrones observados a escala biogeográfica no puede extrapolarse a los patrones a escala local, y por lo tanto es poco apropiada como su explicación. De la misma forma, la sugerencia que la competencia interespecífica es importante en la determinación del patrón de uso de hábitat puede ser más relevante a escala local que a escala biogeográfica 5,6. Una solución al problema de identificar y compatibilizar la escala de los patrones y los procesos es diseñar estudios que incluyan un análisis de escalas múltiples 7.
Las características del hábitat tienen influencias importantes sobre las aves. Por ejemplo, la estructura y la composición florística de la vegetación pueden determinar, entre otros aspectos, la distribución y abundancia del alimento, la disponibilidad de perchas para cantar, de cobertura contra predadores y de sitios para nidificar 8. Considerando este papel del hábitat para las aves, muchos investigadores han estudiado cómo las especies lo utilizan y qué procesos podrían dar cuenta del patrón de selección de hábitat observado (e.g., interacciones competitivas, respuestas específicas de las especies 9). Sin embargo, para comenzar a entender esta relación proceso-patrón se necesita establecer primero el grado de variabilidad espacio-temporal de aquellos patrones. Por ejemplo, en bosques deciduos del noreste de Estados Unidos se encontró que Empidonax minimus influía negativamente sobre el uso de hábitat de Setophaga ruticilla 10. Sin embargo, estas especies estaban positivamente asociadas a escala regional 11. A partir de ello se interpretó que los factores que guían la selección de hábitat a gran escala (e.g., formaciones boscosas con diferentes características estructurales de la vegetación) se imponen en este caso a los efectos locales de la competencia interespecífica. En estos bosques también se encontró que el número de parejas reproductivas de Dendroica caerulescens estaba positivamente relacionado con la densidad de arbustos cuando se comparaban las características del hábitat entre distintas parcelas de bosque, pero un análisis a escala del territorio de los individuos indicó que esta variable no tiene incidencia en la selección de los territorios dentro de cada parcela 12. Por su parte, en las estepas arbustivas de América del Norte, las aves responden a las características estructurales del hábitat a escala biogeográfica, pero a escala regional estas asociaciones desaparecen y es más importante la composición florística del hábitat, mientras que a escala local no hay similitudes con los patrones observados a las escalas mayores 13–15.
En este trabajo trataré de mostrar la importancia de la escala de estudio en las investigaciones ecológicas, particularmente en la evaluación de la selección de hábitat en aves. Mi análisis está lejos de ser una revisión exhaustiva; lo que intenta es plantear el tema y estimular su discusión, en especial durante las etapas de planificación de un proyecto de investigación. Mis comentarios estarán enfocados principalmente sobre los aspectos espaciales de la escala del estudio, incluyendo ejemplos concretos de mis investigaciones sobre la relación entre la vegetación y las aves. Sin embargo, los mismos pueden ser igualmente aplicados a los aspectos temporales de la escala de un trabajo de investigación.
¿Qué entendemos por escala?
Por "escala" nos referimos a las dimensiones espaciales y temporales de las entidades o fenómenos que observamos, lo que involucra (al menos implícitamente) mediciones y unidades de medición. Los objetos o eventos pueden ser caracterizados y diferenciados por sus escalas, como cuando hablamos del tamaño de un objeto o de la frecuencia de ocurrencia de un evento. Por regla general, deberíamos usar el término escala cuando somos capaces de asignar o identificar las dimensiones y unidades de medición de nuestro "objeto" de estudio (i.e., el ámbito en el cual realizamos las observaciones).
Lamentablemente, como sucede con otros términos en ecología, muy a menudo en la literatura el término escala es utilizado erróneamente, confundiéndolo con el término "nivel" y viceversa 16. Es común encontrar referencias donde se indica que se trabajó "a escala poblacional" o, inversamente, "a nivel de paisaje", lo que lleva a la pérdida de precisión de ambos términos y a que no se sepa con certeza de qué se está hablando cuando se los utiliza. Teniendo en cuenta que uno de los objetivos de la ciencia es la precisión, deberíamos ser cuidadosos en el uso de la terminología; vale la pena, entonces, aclarar las diferencias entre los dos conceptos. Por nivel nos referimos a los estados de organización de un sistema que se ordena en forma jerárquica. Los niveles de organización clásicos para los ecólogos son individuo, población, comunidad y ecosistema, aunque existen otras posibilidades 16. Los niveles de organización, aunque suelen estar asociados, no necesariamente dependen de la escala espacio-temporal de los objetos o eventos bajo estudio 17. Por ejemplo, a nivel poblacional podemos medir las tasas de natalidad y mortalidad desde escalas espacio-temporales de milímetros y minutos (e.g., bacterias en un medio de cultivo) hasta hectáreas y años (e.g., aves en bosques tropicales). A pesar de las grandes diferencias de escala, el nivel involucrado es el mismo.
¿Cómo expresamos la escala espacio-temporal en ecología?
La escala espacio-temporal de observación en un estudio ecológico tiene dos componentes: la extensión y el grano 18. Desde una perspectiva espacial, la extensión es el área total donde realizamos las observaciones durante un estudio (Fig. 1); este componente de la escala muchas veces es erróneamente considerado como la "escala del trabajo". El grano es el área (o tamaño) de nuestra unidad de observación (Fig. 1), el cual suele coincidir con la unidad de muestreo de los estadísticos. Podemos tomar como ejemplo a un investigador que está interesado en conocer la abundancia de las aves que habitan un algarrobal en el desierto del Monte y prepara el siguiente protocolo de estudio: contar aves en 20 parcelas circulares de 35 m de radio durante 10 min, ubicadas al azar y separadas como mínimo por 250 m, en un área rectangular de 40 ha, una vez por semana durante la época reproductiva (octubre–febrero). El grano de las observaciones es el tamaño de las parcelas circulares y el tiempo de conteo, mientras que la extensión está dada por el número total de parcelas circulares, la separación entre éstas, el tamaño del área rectangular y el número total de semanas que abarcarán las observaciones. La extensión y el grano definen los límites superior e inferior de resolución de un estudio, respectivamente. Toda inferencia acerca de un sistema escala-dependiente está restringida por la extensión y el grano de la investigación. No se puede extrapolar más allá de la extensión sin aceptar el supuesto de uniformidad del patrón bajo estudio (i.e., que la observación o la conclusión son independientes de la escala), un supuesto que muy a menudo no se cumple en los sistemas ecológicos, ni se puede detectar ningún elemento del patrón por debajo del grano.

Figura 1. Representación del grano (cuadrado pequeño, línea entera) y la extensión (cuadrado grande, línea entera) de un estudio en un paisaje con diferentes parches de hábitat. Cuando se aumenta la extensión del estudio (indicado por las flechas en el cuadrado grande) se incorporan nuevos parches de hábitat que no estaban presentes en el área de estudio original. Cuando se incrementa el tamaño del grano (indicado por las flechas en el cuadrado pequeño) algunos parches pequeños de hábitats que originalmente podían ser diferenciados, ahora son incluidos dentro de la misma muestra y las diferencias entre ellos son promediadas. Modificado a partir de Wiens 4.
Cuando cambiamos la escala de medición de una variable, también modificamos la varianza de esa variable. La naturaleza del cambio depende de si modificamos el grano o la extensión de la escala 4. Manteniendo la extensión constante, un incremento del grano generalmente producirá una disminución de la varianza. Mac Nally y Quinn 19 presentan un ejemplo de cambios en la variabilidad de la densidad poblacional de Acanthiza lineata al modificar el grano espacial de sus observaciones. La variabilidad fue evaluada usando la desviación estándar del promedio entre muestreos sucesivos. Analizaron mediante transectas la densidad de esta especie con tres granos espaciales: 15 ha, 30 ha y 45 ha. Los conteos usando la transecta de 15 ha tuvieron una variabilidad entre muestreos de 0.47 individuos/ha, pero a medida que incrementaron el grano la variabilidad se fue reduciendo (30 ha: 0.32 individuos/ha; 45 ha: 0.25 individuos/ha), indicando una dependencia entre la escala de observación (grano) y la variabilidad de las mediciones. La densidad promedio de Acanthiza lineata, por el contrario, mostró pocos cambios con los tres granos utilizados (0.84, 0.73 y 0.72 individuos/ha, respectivamente). Es interesante notar que esta reducción en la variabilidad de las observaciones no se debió al incremento del número de transectas, sino al aumento del tamaño de la transecta, sugiriendo que el consejo usual de los estadísticos de "incrementar el número de muestras para reducir la variabilidad en la estimación de un parámetro" no es la única solución. Al mismo tiempo, dada esta dependencia entre la escala y la variabilidad de las mediciones, es de suma importancia indicar el grano y la extensión que se usaron para tomar las mediciones en un estudio.
Efecto de la escala sobre el grado de apertura de los sistemas ecológicos
Uno de los problemas más complejos que enfrentan los ecólogos es que los sistemas que estudian son en gran medida abiertos. Podemos considerar que un sistema ecológico es cerrado cuando la tasa de transferencia entre sistemas adyacentes se aproxima a cero o cuando las diferencias en los procesos entre ellos son tan grandes que sus dinámicas están efectivamente desacopladas. En cambio, un sistema es abierto cuando la tasa de transferencia es relativamente alta y las dinámicas de los elementos están acopladas entre sí en algún grado 4. Podemos establecer grados de apertura de los sistemas ecológicos a partir de considerar explícitamente la escala de observación. Consideremos un estudio hipotético sobre la dinámica poblacional de Crypturellus tataupa en islas del Paraná Medio que contienen un mosaico de hábitats (Fig. 2). La dinámica poblacional a escala de hábitat puede ser influenciada por la dispersión de individuos entre los parches de una isla y, por lo tanto, el sistema es abierto a esa escala. A una escala más amplia, por el contrario, la migración entre islas puede ser nula y el sistema, para la población de Crypturellus tataupa de una isla, resulta cerrado. Sin embargo, el sistema debería ser considerado nuevamente abierto si en el estudio se está evaluando el efecto de las fluctuaciones climáticas sobre la dinámica de las poblaciones que habitan todas las islas del Paraná Medio.

Figura 2. Un ejemplo de la relación entre la escala del estudio y el grado de apertura del sistema ecológico (ver detalles en el texto). Los pares de flechas indican la posibilidad de dispersión de los individuos entre parches de hábitat y entre islas.
Conocer el grado de apertura de los sistemas es de suma importancia para incrementar la probabilidad de que las mediciones efectuadas a una determinada escala revelen los mecanismos ecológicos que están operando. Por ejemplo, la diversidad de especies en una comunidad local puede estar determinada no solo por factores locales (e.g., heterogeneidad de hábitats, disponibilidad de recursos, abundancia de competidores), sino también por las tasas de especiación y extinción que operan a escala biogeográfica 20. Los cambios en el tamaño de una población a escala local pueden reflejar alteraciones del hábitat a escala regional o incluso eventos en otras partes de la distribución geográfica de la especie 21.
Importancia de la escala para detectar patrones e inferir procesos
Las características que podemos observar de un sistema ecológico pueden diferir si utilizamos una escala pequeña o grande, y el tipo de escala elegida determinará en gran medida la forma en que podemos estudiar el sistema (Tabla 1). Los estudios a escala pequeña son más útiles para evaluar en mayor detalle los mecanismos subyacentes, mientras que a grandes escalas es más probable encontrar patrones generales. La posibilidad de realizar experimentos replicados aumenta cuando reducimos la escala de la investigación. El potencial para cometer varios tipos de errores es menor a escalas grandes (aunque tal vez aumente el riesgo de enfatizar asociaciones espurias), pero a su vez se reduce la posibilidad de incrementar el esfuerzo de muestreo.
Tabla 1. Características generales de los atributos ecológicos y de las posibilidades de investigación que pueden depender de la escala de análisis (modificado a partir de Wiens 4).
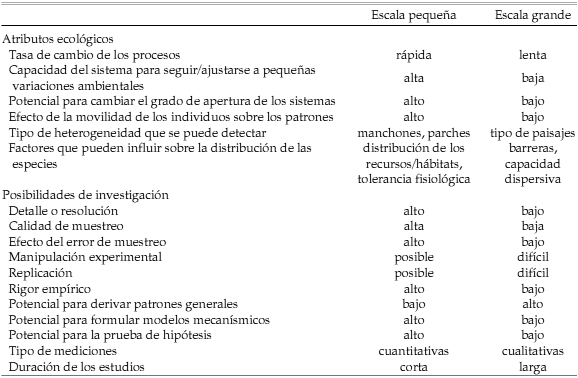
La escala de un estudio determina el rango de posibilidades para detectar patrones y procesos, por lo que si estudiamos un sistema en una escala inapropiada podemos cometer el error de identificar patrones que, en realidad, son artefactos del análisis. Por ejemplo, un investigador encuentra que dos especies de aves granívoras del mismo género difieren en la longitud del pico en simpatría pero no en alopatría, y concluye que dicho patrón es evidencia de desplazamiento de caracteres debido a la competencia interespecífica en el área de simpatría. Sin embargo, el patrón en la longitud del pico podría estar relacionado con una variación clinal en el peso de las dos especies (i.e., una respuesta alométrica) y, por lo tanto, el patrón observado podría ser un ejemplo de falso desplazamiento de caracteres 22. El investigador no detectó el cambio clinal debido a que la extensión de su trabajo no abarcó el rango completo de variación del rasgo morfológico considerado. Para el tipo de conclusión que quería alcanzar necesitaba un trabajo de una escala (extensión) mayor y, por lo tanto, realizó un muestreo incompleto. Quizás el mayor riesgo de seleccionar una escala inapropiada es que, como solemos ser muy ingeniosos, es probable que desarrollemos explicaciones para los datos obtenidos y creamos que comprendemos el funcionamiento del sistema, cuando en realidad solo hemos colectado un conjunto de datos que no permiten analizar confiablemente el problema que estamos tratando de estudiar 4.
Elección de la escala apropiada
Como se señaló anteriormente, una elección equivocada de la escala de estudio puede llevarnos a cometer errores de interpretación. La aproximación que se utiliza más frecuentemente para evaluar fenómenos a varias escalas es la selección arbitraria de varios puntos sobre el continuo de escalas posibles. Wiens et al. 23 reconocen cuatro escalas espaciales de utilidad general en las investigaciones ecológicas (Fig. 3): (1) escala biogeográfica: es un área suficientemente grande como para incluir diferentes climas, formaciones vegetales y ensambles de especies; (2) escala regional: una superficie que contiene muchos tipos de hábitats o poblaciones que pueden o no estar unidas por dispersión; (3) escala local: manchones de hábitat ocupados por muchos individuos de diferentes especies; y (4) escala del individuo: espacio ocupado por un individuo durante un período relevante de tiempo.

Figura 3. Ejemplos de estudios a las escalas espaciales generalmente utilizadas en las investigaciones ecológicas. (A) Biogeográfica. Se estudian patrones con muestras localizadas en sitios dispersos que abarcan varias provincias fitogeográficas (e.g., Monte, Espinal, Pampa). (B) Regional. El muestreo abarca 12 sitios de estudio en la porción austral del desierto del Monte. (C) Local. Las muestras son parcelas de 10 ha que permiten un análisis comparativo en un sitio de estudio. (D) Individuo. El muestreo está concentrado en el área de los territorios ocupados por individuos de una especie (zonas negras) y en las porciones no ocupadas, dentro de una parcela de estudio.
El problema de estas escalas es que están basadas en nuestra percepción particular de la naturaleza y pueden no ser las más relevantes para un caracol, una lagartija o un ave. Al estudiar distintos tipos de organismos se condiciona la escala de la investigación, aún cuando sean similares en algún aspecto particular. Por ejemplo, un escarabajo carroñero no se relaciona con su ambiente a la misma escala que un ave rapaz carroñera, aunque compartan el tipo de alimento. Estas diferencias entre organismos pueden ser vistas en términos de lo que Addicott et al. 24 denominaron vecindario ecológico ("ecological neighbourhood"), es decir áreas que son determinadas en función de un proceso ecológico, un período de tiempo o la capacidad de movilidad o actividad del organismo particular bajo estudio. Así, para un individuo de Parula pitiayumi que habita un bosque de Celtis tala el vecindario ecológico de sus desplazamientos durante toda su vida probablemente será de unos pocos kilómetros cuadrados, mientras que para un individuo de Geranoaetus melanoleucus involucrará un área de cientos de kilómetros cuadrados. Algunos autores han propuesto usar relaciones alométricas para generar funciones de escalamiento para organismos de diferentes tamaños 1,25. Sobre esta base, por ejemplo, el área de actividad (i.e., el vecindario ecológico) para un ave de 20 g sería del orden de 4 ha, mientras que para una de 200 g debería ser de aproximadamente 90 ha. La elección de la escala de estudio también depende en gran medida de las preguntas que buscamos responder con nuestra investigación. Los ecólogos de comportamiento, de poblaciones y de comunidades pueden compartir su interés en analizar la relación entre los recursos y sus consumidores, pero sus objetivos particulares los conducirán a trabajar en distintas escalas.
Como se desprende de los párrafos precedentes, no hay una receta sencilla para seleccionar la escala apropiada de un estudio. Sin embargo, hay dos factores significativos que guían esa selección. El primero es tener bien en claro cuál es la pregunta que deseamos responder con nuestro protocolo de investigación. El segundo es conocer la historia natural del sistema que vamos a estudiar. Si bien en ecología existe una fuerte tendencia a trabajar experimentalmente, todavía siguen siendo necesarios los trabajos basados en la observación de la historia natural de las especies, los cuales, con un buen diseño de muestreo, aportan las bases empíricas para proponer hipótesis que se pueden poner a prueba subsecuentemente con un protocolo experimental 26.
Dominio de las escalas
Si la interpretación de los sistemas ecológicos dependiera de la escala en forma continua (i.e., si cada cambio en la escala promoviera una modificación de los patrones y procesos), entonces sería muy difícil encontrar alguna generalización, ya que el rango de extrapolación de los estudios quedaría muy limitado. En cambio, si el gradiente de escalas no es continuo, entonces podrían existir "dominios" de las escalas, que son rangos o "porciones" del gradiente de escalas en los que el patrón o proceso no cambia significativamente 4. Los dominios estarían separados por zonas de transición donde los patrones o procesos pasan de ser controlados por un grupo de factores a ser controlados por otros, de manera similar a los cambios de fase en los sistemas físicos 4. El dominio de una escala, siempre para un determinado patrón o proceso, definiría los límites dentro de los cuales es posible una generalización. Los descubrimientos a una escala particular pueden ser extrapolados a otras escalas dentro del dominio, pero la extensión de las conclusiones cruzando las zonas de transición sería peligroso 4. Las mediciones realizadas en dominios distintos no deberían ser comparadas directamente, pues las asociaciones entre variables dentro de un dominio podrían desaparecer o cambiar de signo cuando la escala es extendida más allá del mismo. Las explicaciones de un patrón derivadas de estudios mecanísmicos a escala pequeña diferirán dependiendo de si nosotros hemos ampliado la escala dentro del mismo dominio, entre dominios o cruzado varios de ellos.
Nuevamente surge la utilidad de la perspectiva de múltiples escalas, en este caso como una forma de identificar dominios a lo largo de un continuo de escalas. Los estudios ecológicos deberían ser realizados a varias escalas, con cambios en el grano y la extensión, independientes uno del otro. Esta aproximación provee una mejor resolución a los problemas de dominio y ayuda a incrementar el grado de generalidad de los patrones observados y de sus determinantes.
Enfoques a múltiples escalas: un ejemplo
Las aves muestran una estrecha relación con las características estructurales y florísticas de la vegetación cuando seleccionan el hábitat donde residen, pero la importancia de esos componentes puede depender de la escala del estudio (e.g., véanse los tres casos mencionados en la parte introductoria de este trabajo). Por este motivo, se analizó la relación entre la vegetación y las aves utilizando un enfoque de múltiples escalas 27. El trabajo fue realizado en tres escalas espaciales: biogeográfica, local y del individuo (ver Fig. 3).
A escala biogeográfica se analizó la influencia del clima y de la disponibilidad de estratos de vegetación sobre la riqueza de especies de aves passeriformes en la provincia de Buenos Aires 28. A escala local se estudió la respuesta del ensamble de aves que habitan los bosques de Celtis tala (talares) de la provincia de Buenos Aires a las alteraciones de la estructura y composición florística de la vegetación 29. Finalmente, a escala del individuo se evaluó la selección de microhábitats de alimentación por las aves en los talares 30. La descripción detallada de los métodos utilizados para analizar la relación entre la vegetación y las aves se puede encontrar en los trabajos originales. A continuación se describirán sucintamente, haciendo énfasis en la extensión y el grano de cada uno de los análisis.
Para realizar el trabajo a escala biogeográfica se subdividió la superficie de la provincia de Buenos Aires en 146 cuadrados de 50 km de lado. La extensión fue la superficie cubierta por los 146 cuadrados (aproximadamente 350000 km2), mientras que el grano fue la superficie de cada cuadrado (aproximadamente 2500 km2). En cada cuadrado se evaluó la presencia de las especies de aves passeriformes (obteniéndose la riqueza específica), el número de estratos de vegetación característico (derivado del tipo de vegetación predominante) y los valores de 13 variables relacionadas con la precipitación y la temperatura. La matriz de datos climáticos fue sometida a un Análisis de Componentes Principales, obteniéndose dos componentes (nuevas variables) que sintetizaron la mayor parte de la variación climática en la provincia. Las relaciones entre el clima, los estratos de vegetación y la riqueza de aves fueron evaluadas mediante Análisis de Correlación Simple y Parcial. Por su parte, a escala local se estudiaron las diferencias en los ensambles de aves que habitan talares con diferente estructura y composición florística de la Reserva El Destino (Partido de Magdalena). Los talares se encuentran ubicados sobre cordones de conchilla paralelos o subparalelos a la costa del Río de la Plata. Estos cordones se originaron por depósitos marinos como consecuencia de ingresiones del mar durante el Cuaternario, los cuales determinaron elevaciones de 1–2 m sobre el nivel de las áreas adyacentes. Esta característica topográfica conformó un paisaje de isletas de bosques de tamaño variable (entre 1–30 ha). En la reserva se encuentran talares que han sido protegidos de la explotación forestal y ganadera desde 1920 y otros que han sido explotados por tala selectiva hasta 1960 y que actualmente continúan siendo utilizados para la actividad ganadera (principalmente como refugio). Estos últimos se han regenerado principalmente a partir del rebrote de los tocones y, en la actualidad, presentan menor cobertura en el dosel y en los estratos bajos con respecto a los talares protegidos. Además, varias especies de árboles sufrieron una marcada disminución en su cobertura (especialmente Celtis tala, Jodina rhombifolia y Sambucus australis). Para evaluar los ensambles de aves se seleccionaron cuatro áreas de aproximadamente 15000 m2 (dos en cada tipo de talar). En cada área se establecieron al azar tres parcelas circulares de 1963 m2 cada una (el radio de cada parcela fue de 25 m), separadas entre sí por no menos de 150 m. La extensión del estudio fue de una superficie aproximada de 60000 m2 e incluía las 12 parcelas circulares, mientras que el grano era la superficie de una parcela circular (1963 m2). En cada parcela se contó el número de individuos de cada especie registrada, obteniéndose así la densidad de cada una. Finalmente, a escala del individuo se evaluó el comportamiento de alimentación y la selección de microhábitats de alimentación por las aves en el talar protegido de la Reserva El Destino. Para obtener los datos de comportamiento se recorrieron dos áreas de 15000 m2 en forma sistemática. Cada vez que un ave era observada alimentándose, se registraba la maniobra de alimentación empleada, el sustrato donde encontraba el alimento, la altura a la que capturaba su presa y la especie de planta en la que la encontraba. En este caso la extensión del trabajo fue de aproximadamente 30000 m2 (las dos áreas establecidas en el talar protegido) y el grano era el sitio donde el ave capturaba su alimento (un área menor a 1 m2).
El análisis biogeográfico sugirió que la riqueza de especies de aves passeriformes está asociada a la disponibilidad de estratos de vegetación (Fig. 4). A esta escala, la estructura de la vegetación es más importante que el clima en la determinación de los patrones de distribución geográfica de las aves.

Figura 4. Esquema de la relación entre las variables que influyen sobre la riqueza de especies de aves passeriformes en la provincia de Buenos Aires. La estructura de la vegetación está estimada a partir del número de estratos de los tipos de vegetación dominantes en la provincia. El primer factor climático está asociado a un gradiente NE-SO generado por un aumento de la precipitación y la humedad relativa y una disminución de la amplitud térmica y del número de heladas hacia el NE. El segundo factor climático está relacionado con un gradiente SE-NO determinado por un aumento de las temperaturas de enero y de la amplitud térmica hacia el NO. Se muestran los valores de las correlaciones parciales estadísticamente significativas entre variables (líneas llenas). Para más detalles ver Cueto y Lopez de Casenave 28. NS: no significativo.
A escala local, tanto la estructura de la vegetación como la composición florística influyen en el uso de hábitat. Las aves insectívoras responden a las diferencias en estructura de la vegetación (Fig. 5A), siendo menos abundantes en los bosques con una menor disponibilidad de follaje en altura (Fig. 6). Las frugívoro-insectívoras, por su parte, responden más a la composición florística del bosque (Fig. 5B), porque son menos abundantes en los sitios que presentan una menor abundancia y riqueza de especies de plantas productoras de frutos (e.g., Celtis tala, Jodina rhombifolia, Sambucus australis; Fig. 7) solo durante la primavera y el verano, precisamente la época de mayor oferta de frutos en los talares.

Figura 5. Densidad estacional promedio (+ EE), en ind/ha, de aves insectívoras (A) y frugívoro-insectívoras (B) que habitan los talares protegidos (barras negras) y explotados (barras blancas) de la Reserva El Destino, Magdalena, Buenos Aires. Las densidades de las aves insectívoras y frugívoro-insectívoras fueron estadísticamente mayores en los talares protegidos que en los explotados (ANOVA de 2 Factores con Medidas Repetidas en un Factor, Efecto Principal Tipo de Talar; F = 32; gl = 1,10; P < 0.01 y F = 5.5; gl = 1,10; P < 0.05). Para más detalles ver Cueto y Lopez de Casenave 29.

Figura 6. Perfil de cobertura de follaje en altura en los talares protegidos (puntos negros) y explotados (puntos blancos) de la Reserva El Destino, Magdalena, Buenos Aires. Los asteriscos indican diferencias significativas entre los dos tipos de talar (Prueba de t a dos colas; *: P < 0.05, **: P < 0.01, n = 8).

Figura 7. Cobertura promedio (+ DE) de las especies de árboles en los talares protegidos (barras negras) y explotados (barras blancas) de la Reserva El Destino, Magdalena, Buenos Aires. Los asteriscos indican diferencias significativas entre los dos tipos de talar (Prueba de t a dos colas o Prueba de Mann-Whitney; *: P < 0.05, **: P < 0.001, n = 8). SC: Scutia buxifolia, CT: Celtis tala, LL: Ligustrum lucidum, SL: Schinus longifolius, JR: Jodina rhombifolia, SA: Sambucus australis.
Dos de las especies insectívoras estudiadas a escala del individuo son recolectoras de presas en el follaje (Parula pitiayumi y Polioptila dumicola), mientras que la otra se alimenta mediante el revoloteo (Serpophaga subcristata). Las dos primeras seleccionan el estrato superior del bosque para alimentarse, mientras Serpophaga subcristata se alimenta en todos los estratos (Fig. 8). Elaenia parvirostris, una especie frugívoro-insectívora, se alimenta principalmente mediante la caza por revoloteo y también captura su alimento en todos los estratos del bosque (Fig. 8). Estos resultados sugieren que a escala del individuo la estructura de la vegetación no debería afectar de la misma manera a las cuatro especies. Serpophaga subcristata y Elaenia parvirostris no seleccionan estratos de vegetación para alimentarse y, por lo tanto, no deberían sufrir una disminución en la oferta de sitios de alimentación cuando se simplifica la estructura del bosque. Este resultado se refleja parcialmente a escala local cuando se analizó la respuesta poblacional de estas especies. Parula pitiayumi y Polioptila dumicola fueron menos abundantes en los bosques con menor oferta de microhábitats de alimentación (los talares explotados), mientras que Serpophaga subcristata no mostró marcadas variaciones en su densidad entre ambos tipos de bosques (Tabla 2). En cambio, para Elaenia parvirostris la respuesta poblacional no fue la esperada en función de su selección de microhábitats: aunque no selecciona microhábitats a escala del individuo, sus densidades difieren entre tipos de bosque a escala local (Tabla 2). Esta diferencia se debería, como ya se indicó, a la disminución en la abundancia o a la falta de especies productoras de frutos en los talares explotados (ver Fig. 7).

Figura 8. Distribución vertical de las actividades de alimentación de cuatro especies de aves que habitan los talares protegidos de la Reserva El Destino, Magdalena, Buenos Aires. Los datos son porcentajes de observaciones a distintas alturas (puntos negros) y se muestra también la disponibilidad de follaje en altura (puntos blancos). Existen diferencias estadísticamente significativas entre la disponibilidad de follaje y la distribución vertical de la actividad de alimentación para Polioptila dumicola y Parula pitiayumi (χ2 = 38.4, gl = 4, n = 45, P < 0.005 y χ2 = 38.4, gl = 6, n = 66, P < 0.0001, respectivamente). Para más detalles ver Cueto y Lopez de Casenave 30.
Tabla 2. Densidad promedio anual (± DE) de cuatro especies de aves que habitan los talares protegidos y explotados de la Reserva El Destino, Magdalena, Buenos Aires.

Los resultados de los análisis a escalas mayores reflejaron solo parcialmente los obtenidos a escala del individuo, quizás porque los últimos enfatizan los aspectos más relacionados con la percepción ambiental de los factores últimos de la selección de hábitat de las aves, ya que están asociados con la adquisición de sus recursos alimenticios y con sus condicionamientos comportamentales y morfológicos. A escala del individuo, la estructura de la vegetación y la composición florística son dos características que estarían asociadas con la obtención del alimento (el que actuaría como factor último) y que servirían como guías para la selección del hábitat (i.e., como factores próximos). Sin embargo, cuando se analizan los resultados a mayores escalas no todas las especies responden de la manera predicha por los estudios a escala del individuo, enfatizándose la importancia de otros factores que afectan la selección de hábitat por las aves. Por ejemplo, analicemos qué sucede con las cuatro especies en las tres escalas. Una revisión de su distribución geográfica en la provincia de Buenos Aires muestra que las cuatro son muy frecuentes en las zonas costeras 31, coincidiendo muy bien con la distribución de los bosques costeros (i.e., selva en galería y talares). Así, a escala biogeográfica las características estructurales del paisaje restringen la presencia de estas especies (i.e., donde no hay hábitats boscosos no están presentes), aunque a medida que reducimos la escala la respuesta es diferente. Para algunas especies, a escala local y del individuo sólo es importante que se encuentren presentes estructuras arbóreas (Serpophaga subcristata), mientras que para otras hay otros factores, como la disponibilidad de distintos estratos de vegetación (Parula pitiayumi y Polioptila dumicola) o la presencia de árboles con frutos (Elaenia parvirostris) que determinan su uso del hábitat. Sin un enfoque a múltiples escalas estas diferencias no podrían ser detectadas y, por lo tanto, la extrapolación de los patrones determinados a una escala podría llevar a conclusiones erróneas cuando se los aplica a otras escalas.
Conclusión
En este trabajo intenté reflexionar sobre la importancia de los aspectos metodológicos de la escala en los estudios ecológicos. Si bien se expusieron los problemas espaciales de la escala, también es importante tener en cuenta los temporales 23. Por ejemplo, nuestra capacidad para determinar qué factores influyen en la selección de hábitat por las aves depende de si el estudio involucra la época reproductiva o todo el año, ya que es probable que los requerimientos de uso de hábitat sean distintos durante el período de nidificación y durante el invierno. Comprender la importancia de establecer la escala espacio-temporal de nuestro trabajo no solo permitirá una mejor planificación de nuestro proyecto de investigación (tanto conceptual como logística), sino que facilitará la interpretación de los datos obtenidos y mejorará la posibilidad de desarrollar explicaciones acerca de nuestro problema de estudio.
Agradecimientos. A Javier Lopez de Casenave por su amistad y por compartir más de 10 años en la búsqueda de entender cómo se organizan los ensambles de aves. También le agradezco por su energía en las largas jornadas en el campo, las charlas sobre las aves de los talares y por haber insistido para que escribiera este trabajo. A John A. Wiens, quien no creo que haya leído alguna vez mis "trabajos", porque mi lectura de los suyos ha contribuido enormemente a mi formación como científico. A Luis Marone mi agradecimiento es por los mismos motivos que al Dr. Wiens, pero su caso es aún más meritorio porque él sí ha sufrido la lectura de mis manuscritos (incluido éste...). A Fernando Milesi y a un revisor anónimo por sus sugerencias y comentarios para mejorar el manuscrito. También agradezco a los alumnos de los cursos 2004 y 2005 de Ecología de Comunidades y Ecosistemas (FCEyN, UBA) por las estimulantes discusiones cuando analizábamos el problema de la escala en ecología. Casi todo mi trabajo de investigación sobre selección de hábitat fue en gran medida "auto-financiado"; sin embargo, en los últimos tiempos debo agradecer a CONICET y ANPCyT (actualmente a través del PICT 01-12199) de Argentina por el financiamiento recibido. Esta es la contribución N° 48 del Grupo de Investigación en Ecología de Comunidades de Desierto (Ecodes), de IADIZA (CONICET) y FCEyN (UBA).
Bibliografía Citada
1 Schneider DC (1994) Quantitative ecology: spatial and temporal scaling. Academic Press, San Diego [ Links ]
2 Kareiva P y Andersen M (1988) Spatial aspects of species interactions: the weeding of models and experiments. Pp. 38–54 en: Hastings A (ed) Community ecology. Springer-Verlag, Nueva York [ Links ]
3 Peterson DL y Parker VT (1998) Ecological scale: theory and applications. Columbia University Press, Nueva York [ Links ]
4 Wiens JA (1989) Spatial scaling in ecology. Functional Ecology 3:385–397 [ Links ]
5 Wiens JA (1983) Avian community ecology: an iconoclastic view. Pp. 355–403 en: Brush AH y Clark GA (eds) Perspectives in ornithology. Cambridge University Press, Cambridge [ Links ]
6 Wiens JA (1986) Spatial and temporal variation in studies of shrubsteppe birds. Pp. 154–172 en: Diamond J y Case TJ (eds) Community ecology. Harper & Row, Nueva York [ Links ]
7 Kotliar NB y Wiens JA (1990) Multiple scales of patchiness and patch-structure: a hierarchical framework for the study of heterogeneity. Oikos 59:253–260 [ Links ]
8 Cody ML (1985) Habitat selection in birds. Academic Press, Nueva York [ Links ]
9 Wiens JA (1989) The ecology of bird communities. Cambridge University Press, Cambridge [ Links ]
10 Sherry TW y Holmes RT (1988) Habitat selection by breeding American Redstarts in response to a dominant competitor, the Least Flycatcher. Auk 105:350–364 [ Links ]
11 Holmes RT y Sherry TW (1988) Assessing population trends of New Hampshire forest birds: local vs. regional patterns. Auk 105:756–768 [ Links ]
12 Steele BB (1992) Habitat selection by breeding black-throated blue warblers at two spatial scales. Ornis Scandinavica 23:33–42 [ Links ]
13 Rotenberry JT y Wiens JA (1980) Habitat structure, patchiness, and avian communities in North American steppe vegetation: a multivariate analysis. Ecology 61:1228–1250 [ Links ]
14 Wiens JA y Rotenberry JT (1981) Habitat associations and community structure of birds in shrubsteppe environments. Ecological Monographs 51:21–41 [ Links ]
15 Wiens JA, Rotenberry JT y Van Horne B (1987) Habitat occupancy patterns of North American shrubsteppe birds: the effects of spatial scale. Oikos 48:132–147 [ Links ]
16 O'Neill RVO y King AW (1998) Homage to St. Michel; or, why are there so many books on scale? Pp. 3–15 en: Peterson DL y Parker VT (eds) Ecological scale: theory and applications. Columbia University Press, Nueva York [ Links ]
17 Allen TFH (1998) The landscape "level" is dead: persuading the family to take it off the respirator. Pp. 35–54 en: Peterson DL y Parker VT (eds) Ecological scale: theory and applications. Columbia University Press, Nueva York [ Links ]
18 Allen TFH y Starr TB (1982) Hierarchy: perspectives for ecological complexity. University of Chicago Press, Chicago [ Links ]
19 Mac Nally R y Quinn GP (1998) Symposium introduction: the importance of scale in ecology. Australian Journal of Ecology 23:1–7 [ Links ]
20 Ricklefs RE (1987) Community diversity: relative roles of local and regional processes. Science 235:167–171 [ Links ]
21 Marra PP, Hobson KA y Holmes RT (1998) Linking winter and summer events in a migratory bird using stable-carbon isotopes. Science 282:1884–1886 [ Links ]
22 Grant PR (1975) The classical case of character displacement. Evolutionary Biology 8:237–337 [ Links ]
23 Wiens JA, Addicott JF, Case T y Diamond J (1986) The importance of spatial and temporal scale in ecological investigations. Pp. 145–153 en: Diamond J y Case TJ (eds) Community ecology. Harper & Row, Nueva York [ Links ]
24 Addicott JF, Aho JM, Antolin MF, Padilla DK, Richardson JS y Soluk DA (1987) Ecological neighbourhoods: scaling environmental patterns. Oikos 49:340–346 [ Links ]
25 Schneider DC (1998) Applied scaling theory. Pp. 253–269 en: Peterson DL y Parker VT (eds) Ecological scale: theory and applications. Columbia University Press, Nueva York [ Links ]
26 Marone L, Lopez de Casenave J y Cueto VR (2001) Historia natural local y ciencia ecológica universal: los sistemas semillas-granívoros en desiertos de Sudamérica y Norteamérica. Pp. 678–679 en: Primack R, Rozzi R, Feinsinger P, Dirzo R y Massardo F (eds) Fundamentos de conservación biológica: perspectivas latinoamericanas. Fondo de Cultura Económica, México DF [ Links ]
27 Cueto VR (1996) Relación entre los ensambles de aves y la estructura de la vegetación. Un análisis a tres escalas espaciales. Tesis doctoral, Universidad de Buenos Aires, Buenos Aires [ Links ]
28 Cueto VR y Lopez de Casenave J (1999) Determinants of bird species richness: role of climate and vegetation structure at a regional scale. Journal of Biogeography 26:487–492 [ Links ]
29 Cueto VR y Lopez de Casenave J (2000) Bird assemblages of protected and exploited coastal woodlands in east-central Argentina. Wilson Bulletin 112:395–402 [ Links ]
30 Cueto VR y Lopez de Casenave J (2002) Foraging behavior and microhabitat use of birds inhabiting coastal woodlands in eastcentral Argentina. Wilson Bulletin 114:342–348 [ Links ]
31 Narosky T y Di Giacomo AG (1993) Las aves de la Provincia de Buenos Aires: distribución y status. Asociación Ornitológica del Plata, Vázquez Mazzini y L.O.L.A., Buenos Aires [ Links ]

