










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEl hornero
versión impresa ISSN 0073-3407versión On-line ISSN 1850-4884
Hornero v.21 n.2 Buenos Aires ago./dic. 2006
A re-evaluation of evidence raises questions about the fasting migration hypothesis for Swainson's Hawk (Buteo swainsoni)
Marc J. Bechard 1,2, Jose H. Sarasola 2 and Brian Woodbridge 3
1 Raptor Research Center, Department of Biology, Boise State University. Boise, ID 83725, USA. mbechard@boisestate.edu
2 Centro para el Estudio y Conservación de las Aves Rapaces en Argentina (CECARA), Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa. Avda. Uruguay 151, 6300 Santa Rosa, La Pampa, Argentina.
3 U. S. Fish and Wildlife Service. PO Box 1006, Yreka, CA 96097, USA.
Abstract
We examined the fasting migration hypothesis for Swainson's Hawks (Buteo swainsoni) by estimating the length, duration, and speed of the migration between North and South America and measuring changes in their body masses at various times throughout the year. We instrumented 34 adult Swainson's Hawks with satellite radios on their breeding grounds in western North America to determine the duration, length, and speed of the migration. Migrating south at 188 km/day, it took Swainson's Hawks 51 days to complete their 13504 km migration to their austral summer grounds. Averaging only 150 km/day on their return migration, it took them 60 days to complete the shorter 11952 km migration back to North America. Adult male and female Swainson's Hawks had average body masses of 872 g and 1131 g, respectively, when they departed from North America in September and their body masses upon arrival in Argentina averaged 759 g for adult males and 933 g for adult females, indicating they lost only an average of 18% of their body masses during migration. Adult male and female Swainson's Hawks had body masses of 792 g and 1013 g, respectively, in February prior to their departure from Argentina and they returned to the breeding grounds in North America weighing 802 g and 1087 g in April. Our results indicate that the fasting migration model does not predict the actual body masses of Swainson's Hawks during the migration season and it should be modified to account for either lower energy expenditures during migration or the possibility that the birds use a stopover strategy during migration, feeding and regaining, or maintaining, fat stores along the migration route.
Key words: body mass, Buteo swainsoni, fasting, migration, Swainson's Hawk.
Resumen
Nuevas evidencias cuestionan la hipótesis del ayuno durante la migración para el Aguilucho Langostero (Buteo swainsoni)
Se examinó la hipótesis del ayuno durante la migración para el Aguilucho Langostero (Buteo swainsoni) mediante la estimación de la longitud, la duración y la velocidad de migración entre América del Norte y del Sur, y la medición de los cambios en el peso corporal en varios momentos a lo largo del año. Se colocaron transmisores satelitales a 34 aguiluchos adultos en sus áreas de cría en el oeste de América del Norte para determinar la duración, la longitud y la velocidad de la migración. En su migración al sur, a 188 km/día, el Aguilucho Langostero tarda 51 días para completar los 13504 km hasta las áreas donde permanece durante el verano austral. A un promedio de solo 150 km/día en su migración de retorno, le toma 60 días completar los 11952 km de regreso a América del Norte. Los machos y hembras adultos de Aguilucho Langostero tuvieron un peso corporal promedio de 872 g y 1131 g, respectivamente, cuando partían de América del Norte en septiembre, y su peso corporal al arribar a Argentina promedió 759 g en los machos adultos y 933 g en las hembras adultas, indicando que pierden, en promedio, solo el 18% de su peso corporal durante la migración. En febrero, antes de su partida de Argentina, los machos y hembras adultos de Aguilucho Langostero tuvieron un peso corporal de 792 g y 1013 g, respectivamente, y llegaron a sus áreas de cría en América del Norte pesando 802 g y 1087 g, en abril. Los resultados indican que el modelo de ayuno durante la migración no predice los pesos corporales reales del Aguilucho Langostero durante la estación migratoria y que debería ser modificado para dar cuenta de menores gastos energéticos durante la migración o la posibilidad de que las aves usen una estrategia de paradas durante la migración, alimentándose e incorporando (o manteniendo) grasa a lo largo de la ruta migratoria.
Palabras clave: Aguilucho Langostero, ayuno, Buteo swainsoni, migración, peso corporal.
Received 2 January 2006, accepted 1 December 2006
Migration poses numerous behavioural and physiological challenges for birds. For most species, fat is the fuel used on migration. Prior to and during migration, birds deposit fat and then use it to meet their energy demands (Weis-Fogh 1952, Blem 1980, Ramenofsky 1990, McWilliams et al. 2004). Most warblers and shorebirds undergo rapid premigratory fattening depositing more than 50% of their body weight in fat to make long, non-stop flights over oceans, deserts, or other inhospitable areas where refuelling is difficult if not impossible (Blem 1980, Moore and Kerlinger 1987, Ramenofsky 1990, Jenni and Jenni-Eiermann 1998). However, for most species including raptors, migration is a slower, more extended process, involving a "stop and go" migration pattern with birds typically feeding en route as they encounter opportunities to eat. Using this strategy, migrants may maintain somewhat elevated levels of stored body fat, perhaps using this depot to carry them through periods of food scarcity. Nevertheless, they do not store enough premigratory fat to fuel their entire migration. This necessitates a refuelling strategy which spares their reserve energy supply and may augment their fat stores when food supplies are plentiful along the migration route (Berthold 1975).
There is very little quantitative information on how birds use stored fat during migration. Most studies that have dealt with this issue have focused on birds that use powered (flapping) flight during migration (Berger and Hart 1974, Blem 1980, Ramenofsky 1990). There is little information about fat deposition and use by falconiforms either before or during migration. However, several authors have maintained that migrating raptors take on fat deposits during migration. For example, American Kestrels (Falco sparverius) deposit 2-4% body fat during early autumn (Gessaman 1979) and it is well-known that Amur Falcons (F. amurensis) are extremely fat prior to their long migration over the Indian Ocean from India to East Africa (Ali and Ripley 1978). Western Honey-buzzards (Pernis apivoris) also undergo large variations in weight that are attributed to massive fat deposition prior to migration (Glutz von Blotzheim et al. 1971). Despite this, there is little information on the energy expenditure involved when birds use soaring flight in long-distance migrations.
The Swainson's Hawk (Buteo swainsoni) is one such long-distance, "stop and go" soaring migrant. Each year, it migrates back and forth between the plains, shrublands, steppes, and pampas of North and South America. Prior to its southward migration, it changes from a carnivore to an insectivore diet and hawks are frequently observed in flocks of hundreds of birds gorging themselves on insects such as grasshoppers (Johnson et al. 1987). As they migrate through the Mesoamerican land corridor at about 30°N latitude these small flocks coalesce into fewer, very large flocks consisting of tens of thousands of individuals. As they continue to migrate southward, the hawks are funnelled into extremely dense concentrations with an estimated 800000 hawks passing over the Isthmus of Panama during a nine-day period in late October (Bildstein, unpublished data). Some individuals drop out of the migration in Central America (Smith et al. 1986). Nevertheless, most continue their southward migration with some settling in Brazil, Uruguay, and Bolivia, but most not stopping until they reach their austral grounds in the pampas region of Argentina (England et al. 1997).
Based on the assertion that Swainson's Hawks observed roosting overnight in Panama seemed not to have cast pellets or defecated in roost trees (on the basis of a lack of "whitewash" on the forest floor below roosts), and because the species migrates in such large numbers over most of its migratory route, it has been suggested that Swainson's Hawks may complete most or all of their migration without feeding by fasting and metabolizing fat stores laid down before they depart on migration (Brown and Amadon 1968, Smith et al. 1986). Smith et al. (1986) presented a model based on estimates of the energetic cost and duration of this migration. Their model predicted that hawks would need to accumulate body fat deposits equalling about 55% of their lean body mass before migration to successfully complete the journey in either direction. Using the model's assumptions, a male Swainson's Hawk with a lean body mass of 600 g would have to store 330 g of body fat prior to migration. This model has received criticism (Kirkley 1991) and it is plagued with weaknesses because there have never been any records of premigratory weights for the Swainson's Hawk.
Due to the lack of information on migrant Swainson's Hawks, we undertook a study to document the timing and geographic length of the Swainson's Hawk migration and the changes in its body mass prior to its departure on migration and after its arrival on their breeding and austral summer grounds. We used these body mass estimates to test the fasting migration model proposed by Smith et al. (1986) in an effort to determine if Swainson's Hawks store sufficient amounts of fat to fuel their migration without stopping to eat along the way.
Methods
We carried out our main sampling work in two study areas. One was in the Snake River Plain of southern Idaho in North America and the other was located in the pampas region of South America in Buenos Aires Province and La Pampa Province, central Argentina. Both study areas have undergone extensive agricultural alteration and currently most of the landscape is dominated by croplands planted in row crops and plantations of exotic tree species including black locust (Robinia pseudoacacia) trees in Idaho and eucalyptus (Eucalyptus spp.) trees in Argentina.
To track the migration of hawks between the two study areas, we captured breeding female Swainson's Hawks at their nest sites throughout their North American breeding grounds and instrumented them with Microwave Electronics, Inc. satellite transmitters (PTTs). Details on trapping methods, attachment techniques, and duty cycles of PTTs can be found in Fuller et al. (1998). We used the locations obtained from PTTs to determine the length of the migration route, the duration of migration period, and the speed of migration between North and South America.
To estimate fat stores before and after migration, we captured and weighed male and female Swainson's Hawks between 1998-2001 to obtain body mass estimates for Swainson's Hawks throughout the time they were on their breeding grounds in North America and on their austral summer grounds in Argentina. We captured hawks using either bal-chatri traps with mouse lures or dho-gaza nets with Great Horned Owl (Bubo virginianus) lures. Captured hawks were weighed using 1000 and 2000 g Pesola scales. We also took measurements of wing chord, tail length, culmen length, tarsus length and width, and hallux length using steel rulers or calipers. Breeding adults were sexed on the basis of their behaviour at their nest sites. Juveniles and adults captured on their austral summer grounds were sexed using blood samples and PCR amplification of chromohelicase-DNA genes located on the two sex chromosomes (Griffiths et al. 1998). Blood samples were obtained from the brachial vein of captured birds using 1 ml syringes, and blood was stored in either EDTA buffer solution or 80% alcohol (Burgoyne et al. 1994).
Results
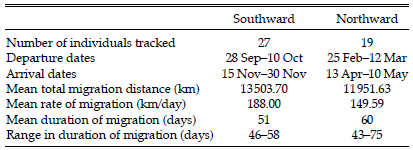
We captured and attached satellite radios to 34 breeding Swainson's Hawks throughout the species' North American breeding range in nine states and provinces. From these instrumented hawks, we obtained 27 complete southward and 19 complete northward migration tracks (Table 1). All of the tracks extended between western North America and central Argentina and both the southward and northward tracks followed a similar route, almost entirely inland except through southern Mexico and Central America where some of the segments of the tracks were along the coast. There were no crossings of large bodies of water. Departure dates on the southward migration varied between late September and early October, and on the northward migration departure dates varied between late February and mid-March. The mean total distance hawks migrated southward was slightly over 13500 km, compared to a mean of slightly less than 12000 km on the northward migration. Hawks migrating southward migrated at an average rate of 188 km/day, and those migrating northward averaged 150 km/day. The duration of the southward and northward migrations averaged 51 and 60 days, respectively. Arrival dates in Argentina varied between mid- to late November. Arrival dates on the North American breeding grounds varied between mid-April and early May.
Table 1. Southward and northward migration tracks of satellite-tagged Swainson's Hawks.
We captured and obtained body mass measurements for a total of 41 adult males and 25 adult females on their North American breeding grounds in 1998 between the time of their arrival in April and May until their departure on migration in September and October (Table 2). Of these 66 individuals captured in their breeding area, 21 adults were captured shortly after their arrival at nesting areas in April and May. An additional 18 breeding adults were captured in July prior to the fledging of their young. In September, 27 adults were captured in the vicinity of their nesting areas prior to their departure on migration. In Argentina, we captured a total of 74 adult males and 37 adult females and 75 juvenile males and 39 juvenile females between the time of their arrival on and departure from their austral summer grounds. Of these 225 captured hawks in Argentina, a total of 59 adults and 39 juveniles were captured in December, whereas 52 adults and 75 juveniles were captured in January and February prior to their departure on the northward migration (Table 2). From the literature, we also obtained estimates of the body mass data for 9 adult male and 12 adult female Swainson's Hawks and 14 juvenile male and 6 juvenile female Swainson's Hawks that were captured in October from 1972-1983 in Panama, approximately midway through their southward migration (Smith et al. 1986).
Table 2. Numbers of adult and juvenile Swainson's Hawks captured in southern Idaho and Argentina.

Adult male and female Swainson's Hawks arrived on their breeding grounds in April and May averaging (± SD) 802 ± 54 g and 1087 ± 87 g, respectively (Fig. 1). During the breeding season, adults lost body mass and averaged only 707 ± 29 g for males and 1013 ± 38 g for females in July. By September, they had regained body masses averaging 872 ± 61 g for males and 1131 ± 72 g for females. Body mass measurements reported for Swainson's Hawks in Panama (Smith et al. 1986), approximately midway through their southward migration, showed that adult male and female Swainson's Hawks had lower body masses and averaged only 730 ± 42 g, and 980 ± 134 g, respectively. Likewise, juvenile male and female Swainson's Hawks in Panama had body masses of only 715 ± 38 g and 941 ± 51 g, respectively (Smith et al. 1986). Adult hawks arriving in Argentina had slightly higher body masses for adult males which averaged 759 ± 86 g, but about 47 g lower for adult females which averaged 933 ± 61 g. Juvenile hawks arrived in Argentina with lower body masses averaging 678 ± 29 g for juvenile males and 802 ± 68 g for juvenile females. After they settled onto their austral summer grounds in Argentina both sexes gained body mass. By January and February, adult males averaged 792 ± 50 g, adult females averaged 1013 ± 70 g, juvenile males averaged 714 ± 54 g, and juvenile females averaged 932 ± 99 g (Fig. 1).

Figure 1. Annual variations in the body masses (in g) of adult (open circles) and juvenile (filled circles) female and adult (open squares) and juvenile (filled squares) male Swainson's Hawks in North, Central, and South America.
Discussion
The fasting model proposed by Smith et al. (1986) is based on an assumption of a fat-free body mass of 600 g for a male Swainson's Hawk, and it assumes that males must gain 330 g of fat to fuel their 9000 km southward migration. Then, they must regain that same amount of body fat before their northward migration in March (Fig. 2). This model does not address females; therefore, we assumed a lean body mass of 800 g for a female Swainson's Hawk and assumed that they also would need to deposit at least 330 g of body fat prior to their southward and northward migrations (Fig. 3). Comparison of our body mass measurements with those of the model showed that, while both sexes and age classes underwent large losses of body mass during their migration, they did not fluctuate as much as the fasting model predicted. Both adult male and female Swainson's Hawks arrived on their breeding grounds with masses greater than those predicted. Nevertheless, prior to their departure on their southward migration, their body masses were nearly identical to those predicted by the model. Body mass measurements for adult and juvenile hawks of both sexes were also similar to those predicted by the model when the hawks were midway through their southward migration in Panama. Our measurements for adult and juvenile male Swainson's Hawks showed opposite trends for the remainder of the migration. Juvenile males lost a small amount of body mass (average of 37 g), whereas adult males gained body mass (average of 29 g) between Panama and Argentina. Both adult and juvenile females continued to lose body mass between Panama and Argentina. Nevertheless, upon their arrival in Argentina, both sexes and age classes had body masses in excess of those predicted by the model.

Figure 2. Comparison of annual variations in the body masses (in g) of adult (open squares) and juvenile (filled squares) male Swainson's Hawks to those predicted by the fasting migration hypothesis model (filled triangles, broken line).

Figure 3. Comparison of annual variations in the body masses (in g) of adult (open circles) and juvenile (filled circles) female Swainson's Hawks to those predicted by the fasting migration hypothesis model (open triangles, broken line).
Our satellite telemetry data showed that Swainson's Hawks undergo a migration pattern that Berthold (1993) has described as a concentration migration. It begins on a broad front at its onset in North America and then funnels down to a narrow flight path as the southward migration moves through Mexico to the southern Gulf of Mexico. The flight path remains narrow throughout Central and South America. All of the hawks we tracked spent the austral summer in central Argentina and none went to Florida or central California where small numbers of wintering Swainson's Hawks are regularly reported. The northward migration again fans out with birds becoming more widely dispersed as they traverse Mexico and enter the southern United States. Hawks making the southward migration departed from their North American breeding grounds by early October and completed their 13500 km journey in 51 days. For the migration north, hawks departed from their austral grounds by mid-March, migrating more slowly and completing their 11950 km migration in 60 days. The northward migration is shorter because, unlike southward migrants which cross to the Pacific Ocean side of Central America following the coastline, northward migrants remain on the Caribbean side of Central America shortening their migration. Nevertheless, the hawks took longer to complete the migration primarily due to their slower rate of travel. In their model for fasting migration, Smith et al. (1986) assumed that between Las Cruces (New Mexico) and Argentina the length of the migration route was 9000 km and the migration took 37.5 days for completion. Because we tracked migrants the entire length of the migration route from breeding areas to austral grounds in Argentina, our estimated length for the total migration route was over 4500 km longer than the length used by Smith et al. (1986) in their model. Consequently, our estimated time for the duration of the migration was 13 days longer in autumn and 22 days longer in spring, or 35 and 59% longer, respectively, than assumed in the model.
Our body mass measurements showed that, like other migrants, Swainson's Hawks undergo large-scale annual changes in their body masses. While little is known about the size of fat deposits of diurnal raptors, it is generally assumed that sizeable changes in body masses at different times of the year represent changes mainly in the amount of stored fat (Gessaman 1979, Newton 1979, Blem 1980). Our results showed that adult male and female Swainson's Hawks gained in excess of 150 g of fat following the breeding season in preparation for their southward migration. Males departed weighing an average of 872 g and females departed weighing an average of 1131 g. Upon their arrival in Argentina in late November and early December, both sexes had lost nearly all of this fat reserve losing an average of 127 and 143 g, respectively. Adults gained body mass on the austral grounds gaining an average of only approximately 50 g of fat by January and February. Goldstein et al. (1999) found that body masses of Swainson's Hawks increased from approximately 800 g to 900 g over this same time period. Surprisingly, male and female Swainson's Hawks arrived on their breeding grounds in North America weighing 802 and 1087 g, or more than they did when they were in Argentina. Fat levels shown by juveniles of both sexes were consistently lower than those we recorded in adults. Juveniles of both sexes had sustained greater losses in body mass than adults when they arrived in Argentina. Juveniles also departed from Argentina with less body fat than adults had.
Our test of the fasting migration model indicated that the body mass fluctuations that Swainson's Hawks experienced prior to and after migration were not great enough for them to have made their migration using only stored fat. Neither adult nor juvenile male Swainson's Hawks had body masses as low as the lean body mass of 600 g that the model is based upon. Similarly, neither adult nor juvenile females had body masses as low as the 800 g lean body mass that we used as an estimate in our comparison. Both sexes arrived in North America and South America weighing more that they should have, if they conformed to the fasting model, and they all departed from Argentina weighing far less than they should have to successfully complete the northward migration using only stored fat. Clearly, the fasting model does not predict the actual body masses of this species throughout the year. The hawks do not arrive lean enough nor do they depart fat enough to accomplish their migration solely using pre-departure fat stores of sufficient magnitude as predicted by the fasting migration hypothesis.
There are several possible explanations for the failure of the fasting model to account for our body mass data. It is possible that the model is based on incorrect assumptions. The assumption that lean male Swainson's Hawks weigh 600 g seems to be too low. The leanest males we measured were captured in July in the middle of the breeding season when energy demands would have been extremely high for males. None of the males we captured had a body mass of 600 g and only one male, the lightest, weighed 655 g. Most males had body masses greater than 700 g, indicating that the estimated lean body mass of male Swainson's Hawks should be increased to about 700 g. A second questionable assumption of the fasting model is the metabolic cost of migration. The model assumes that the cost of soaring flight is twice that of basal metabolic rate (BMR). Smith et al. (1986) suggest that the value for the costs of migratory flight used in their model may vary between 1-3 times BMR. In order for the model to fit the body mass fluctuations we observed, a flight metabolism rate of 1 BMR would need to be assumed as the estimated cost of soaring flight. When we did so, the model conformed to our body mass data much more closely. This implies that migrating Swainson's Hawks would need to use far less energy for flight during their migration than has been previously considered. There are few estimates of the cost of soaring flight in large migratory raptors. The question our data raises is whether it is possible for soaring raptors to expend no more energy than they do at rest. Preliminary data on the heart rates of migrating Turkey Vultures (Cathartes aura) indicate that the heart rate of vultures actually falls below their resting rate (Mandel and Bildstein, unpublished data). Like Turkey Vultures, Swainson's Hawks may also experience very little added energy costs while soaring. A confounding issue is that there are no data available concerning the amount of time that Swainson's Hawks spend using flapping flight during migration. Swainson's Hawks have been observed in Chiapas (Mexico) using flapping flight in the early morning and near roosts in the evening, and during windy days, particularly when there is a head wind (Kirkley, unpublished data). While there are no good estimates of the metabolic costs of flapping flight, if it is used to any extent during the migration period, this would increase the metabolic costs of migration necessitating an adjustment in the model's estimated cost of migration. Clearly, the fasting model needs to be revised extensively in order for it to accurately reflect the trends in body mass we observed.
One puzzling aspect of our findings for the annual body mass fluctuations of Swainson's Hawks is the fact that these hawks depart from the austral summer grounds with far less fat than they should have stored and they arrive on their breeding grounds with far more fat than they should have left. One obvious possibility is that Swainson's Hawks feed during their migratory journeys. Our satellite data indicate that Swainson's Hawks remain in specific locations along the migration route for as long as one week (unpublished data). For example, some hawks remain in Texas for 10 days and some linger in Colombia for nearly one week. Flocks of Swainson's Hawks have been observed feeding on crickets and grasshoppers in Texas (Littlefield 1973) and Costa Rica (Slud 1964). Our telemetry data suggest that, during migration, flocks of hawks may settle into strategic stopover places along the route where they feed and regain fat stores. Their stopover in Texas appears to be important because it allows them to gain fat fuel to power their migration through the Mesoamerican land corridor. It may be speculated that they settle again in Central America or northern South America where they regain fat stores to fuel the remainder of the migration to Argentina. Or, perhaps, they simply feed continually while on migration. Swainson's Hawks become insectivorous during the migration season. They can potentially feed in flight on airborne insects such as dragonflies, grasshoppers, and crickets that collect in columns of rising air called thermals. We have observed flying Swainson's Hawks catching and eating what appear to be insects while migrating through Colombia (unpublished data). The implication is that Swainson's Hawks feed while soaring by eating insects that are in the same thermals they use while migrating.
While our findings argue convincingly that the fasting migration hypothesis for Swainson's Hawks is not correct, there is a need for more data to fully explain how this species successfully completes its long-distance migration each year between North and South America. Further studies are needed which document the body masses of migrating Swainson's Hawks midway through their migration. Capture of migrants in Costa Rica, Panama, and Colombia would better document our estimates of the body masses and fat stores of these birds when they are halfway through their migration. Observations of flocks at roost sites in Central America are also needed. Further documentation of opportunistic feeding and the occurrence of pellets and faeces at roost sites will enable us to ascertain if these birds do, in fact, utilize a stopover strategy to feed and maintain fat stores along the migratory route.
Acknowledgements. Major financial support and other assistance has been provided by the US Department of Interior, Department of Defense, Department of Agriculture, Boise State University, the University of Minnesota, the Canadian Wildlife Service, the Argentine Instituto Nacional de Tecnología Agropecuaria (INTA), and the states of Oregon, Arizona, and Idaho. The American Bird Conservancy, National Fish and Wildlife Foundation, and Novartis, Inc have also provided support. Swainson's Hawks were captured by J. McKinley, M. Martel, G. Holroyd, U. Banasch, D. Matiatos, and others.
Literature Cited
Ali S and Ripley D (1978) Handbook of birds of India and Pakistan. Oxford University Press, Oxford [ Links ]
Berger M and Hart JS (1974) Physiology and energetics of flight. Pp. 416-477 in: Farner DS, King JR and Parkes KC (eds) Avian biology. Volume 4. Academic Press, New York [ Links ]
Berthold P (1975) Migration: control and metabolic physiology. Pp. 77-128 in: Farner DS and King JR (eds) Avian biology. Volume 5. Academic Press, New York [ Links ]
Berthold P (1993) Bird migration: a general survey. Oxford University Press, Oxford [ Links ]
Blem C (1980) The energetics of migration. Pp. 175-224 in: Gauthreaux SA (ed) Animal migration, orientation, and navigation. Academic Press, New York [ Links ]
Brown LH and Amadon D (1968) Eagles, hawks, and falcons of the world. McGraw-Hill, New York [ Links ]
Burgoyne L, Kijas J, Hallsworth P and Turner J (1994) Safe collection, storage, and analysis of DNA from blood. Pp. 36-54 in: Proceedings of the Fifth International Symposium on Human Identification. Promega Corporation, Scottsdale [ Links ]
England AS, Bechard MJ and Houston CS (1997) Swainson's Hawk (Buteo swainsoni). Pp. 1-28 in: Poole A and Gill F (eds) The birds of North America. Academy of Natural Sciences and American Ornithologists' Union, Philadelphia and Washington DC [ Links ]
Fuller MR, Seegar WS and Schueck LS (1998) Routes and travel rates of migrating Peregrine Falcons Falco peregrinus and Swainson's Hawks Buteo swainsoni in the western hemisphere. Journal of Avian Biology 29:433-440 [ Links ]
Gessaman JA (1979) Premigratory fat in the American Kestrel. Wilson Bulletin 91:625-626 [ Links ]
Glutz von Blotzheim UN, Bauer KM and Bezzel E (1971) Handbuch der vögelmitteleuropas. Volume 4. Falconiformes. Akademische Verlagsgesellschaft, Frankfurt [ Links ]
Goldstein MJ, Bloom PH, Sarasola JH and Lacher TE (1999) Post-migration weight gain of Swainson's Hawks in Argentina. Wilson Bulletin 111:428-432 [ Links ]
Griffiths R, Double MC, Orr K and Dawson RJG (1998) A DNA test to sex most birds. Molecular Ecology 7:1071-1075 [ Links ]
Jenni L and Jenni-Eiermann S (1998) Fuel supply and metabolic constraints in migrating birds. Journal of Avian Biology 29:521-528 [ Links ]
Johnson CG, Nickerson LA and Bechard MJ (1987) Grasshopper consumption and summer flocks of non-breeding Swainson's Hawks. Condor 89:676-678 [ Links ]
Kirkley JS (1991) Do migrant Swainson's Hawks fast en route to Argentina? Journal of Raptor Research 25:82-86 [ Links ]
Littlefield CD (1973) Swainson's Hawks preying on fall army worms. Southwestern Naturalist 17:433 [ Links ]
McWilliams SR, Guglielmo C, Pierce B and Klaassen M (2004) Flying, fasting, and feeding in birds during migration: a nutritional and physiological ecology perspective. Journal of Avian Biology 35:377-393 [ Links ]
Moore FR and Kerlinger P (1987) Stopover and fat deposition by North American Wood Warblers (Parulinae) following spring over the Gulf of Mexico. Oecologia 74:47-54 [ Links ]
Newton I (1979) Population ecology of raptors. Buteo Books, Vermillion [ Links ]
Ramenofsky M (1990) Fat storage and fat metabolism in relation to migration. Pp. 215-231 in: Gwinner E (ed) Bird migration. Springer-Verlag, Berlin [ Links ]
Slud P (1964) The birds of Costa Rica. Bulletin of the American Museum of Natural History 128:1-430 [ Links ]
Smith NG, Goldstein DL and Bartholomew GA (1986) Is long-distance migration possible for soaring hawks using only stored fat? Auk 103:607-611 [ Links ]
Weis-Fogh T (1952) Fat combustion and metabolic rate of flying locusts (Schistocerca gregaria Forshal). Philosophical Transactions of the Royal Society of London, B 237:1-36 [ Links ]

