










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEl hornero
versión impresa ISSN 0073-3407versión On-line ISSN 1850-4884
Hornero v.21 n.2 Buenos Aires ago./dic. 2006
Towards a mechanistic interpretation of bird migration in South America
Alex E. Jahn 1,4, Douglas J. Levey 1, Jennifer E. Johnson 2, Ana Maria Mamani 3 and Susan E. Davis 3
1 Department of Zoology, University of Florida. 223 Bartram Hall, Gainesville, FL 32611, USA.
2 Department of Biology, Swarthmore College. 500 College Ave., Swarthmore, PA 19081, USA.
3 Museo de Historia Natural Noel Kempff Mercado. Calle Irala N° 565, Santa Cruz, Bolivia.
4 ajahn@zoo.ufl.edu
Abstract
Research to date has demonstrated that bird migration is comprised of highly diverse and plastic behavioural patterns. Our objective is to highlight the importance of studying mechanisms underlying these patterns in austral migrants. We focus on the high incidence of overlap in breeding and non-breeding ranges as a particularly thought-provoking pattern. We then explore the opportunities afforded by partial migration theory to elucidate the mechanisms underlying seasonal range overlap. We propose that a mechanistic understanding of migration in South America will both provide a deeper appreciation of the ecology, physiology and evolution of migratory species in the New World, and improve the scientific foundation for their conservation.
Key words: mechanism, partial migration, population, range overlap, Tyrannus.
Resumen
Hacia una interpretación mecanística de la migración de aves en América del Sur
La investigación reciente sobre aves migratorias ha demostrado que constituyen un grupo que presenta comportamientos altamente diversos, plásticos y complejos. Nuestro objetivo general es resaltar la importancia de estudiar los mecanismos que generan los patrones que caracterizan la migración de aves en América del Sur. Para ello nos enfocamos en un patrón interesante (la alta incidencia de superposición en la distribución reproductiva y de invernada), analizando las oportunidades ofrecidas por la teoría de migración parcial para dilucidar los mecanismos que producen tal superposición. Proponemos que una comprensión mecanística de la migración de aves en América del Sur no solo proveería una apreciación más profunda sobre la ecología, la fisiología y la evolución de las especies migratorias del Nuevo Mundo, sino que también mejoraría los fundamentos científicos para su conservación.
Palabras clave: mecanismos, migración parcial, población, superposición de distribuciones, Tyrannus.
Received 8 March 2006, accepted 28 December 2006
Research on various aspects of bird migration - from biogeography to ecology and physiology - has demonstrated that migratory behaviour is an extraordinarily diverse and complex phenomenon. For example, it can evolve relatively rapidly (Able and Belthoff 1998, Berthold 1999, Piersma et al. 2005), is phylogenetically flexible (Böhning-Gaese and Oberrath 1999), and can appear and disappear through time within a lineage (Zink 2002, Joseph et al. 2003, Outlaw et al. 2003) or even within an individual's lifetime (e.g., Schwabl and Silverin 1990). In the New World, a continuum of migratory strategies exists, from long-distance migrations undertaken by all populations of a species to short-distance migrations undertaken by only some populations (Levey and Stiles 1992) or individuals (e.g., Ketterson and Nolan 1976). Migration can even play a central role in speciation (Winker 2000, Winker and Pruett 2006).
To solve persistent riddles about the evolution and regulation of migration requires teasing apart factors confounded in space and time. This is difficult to accomplish without a broad geographic scope. In the New World, almost all work on migration has been restricted to the north-temperate latitudes (Levey 1994, Jahn et al. 2004), with relatively little attention paid to migration within South America (austral migration, sensu Chesser 1994). Some species of Neotropical austral migrants move annually between the temperate zone and tropical latitudes and some species migrate within either tropical or temperate latitudes of the continent (Joseph 1997). There are more than 220 species of Neotropical austral migrants, comprising the largest migratory system in the Southern Hemisphere (Chesser 1994). Recent literature on migration in South America principally explores biogeographic patterns (e.g., Chesser 1994, da Silva 1999, Capllonch and Lobo 2005), evolution (Joseph et al. 2003), habitat associations (e.g., Chesser 1995, Stotz et al. 1996, Jahn et al. 2002) and the timing of migratory movements (e.g., Hayes et al. 1994).
The geographical patterns of migration in the Neotropics are complex (Morton 1977, Winker et al. 1997, Bildstein 2004). In some species all populations migrate, but in others different populations migrate in the same or in different directions. Furthermore, all of these strategies can occur within one genus (e.g., genus Tyrannus, Fig. 1; Ridgely and Tudor 1994, Chesser 1995). This stands in sharp contrast to the situation in North America, where all the populations of migratory species generally move in the same direction during each season.

Figure 1. Generalized seasonal distributions of Tyrannus savana savana (a), Tyrannus albogularis (b), and Tyrannus melancholicus melancholicus (c) in South America. Black polygons represent seasonal non-breeding ranges, white polygons represent seasonal breeding ranges and the gray polygon represents area of overlap in which permanent residents as well as non-breeding (i.e., overwintering) migratory individuals from the south occur together. Adapted from Chesser (1995).
Given the diversity of migration strategies evident within South America, it is clear that the phrase "bird migration in South America" encompasses a multitude of inter- and intra-specific patterns at smaller scales than the overall pattern at the continental level would suggest. Thus, to better understand how migratory birds respond to competing ecological and physiological demands, it will be essential to form links between patterns observed at different spatial and temporal scales and among taxonomic (e.g., families and species) and biological levels of organization (e.g., genotypes, individuals, ecosystems) (Levin 1992).
Our objective is to demonstrate that research on the mechanisms generating specific patterns of bird migration in South America is a prerequisite to gaining a better theoretical foothold, as well as for the ability to formulate sound, proactive conservation and management strategies. We use as an example the high incidence of seasonal range overlap in the distributions of South American migratory bird populations to highlight the power of existing theory on intrapopulation variation in migratory behaviour to explain such patterns.
A focus on mechanisms
Studying mechanisms - the proximate relationships between what an individual experiences and how it responds - is the key to being able to explain the causes for observed patterns, advancing both basic science (i.e., explanations, predictions, the formulation of original questions) as well as practical applications (i.e., conservation and environmental planning and management). Such rewards will only result from studies that are focused on specific questions and employ testable hypotheses (Vuilleumier 2004).
Tests of mechanistic hypotheses (i.e., how component parts of the phenomenon interrelate) - rather than phenomenological explanations (i.e., models that extrapolate future trends based on past trends) - are useful for interpreting the reasons for changes observed in a pattern (e.g., migratory timing or direction) (Koehl 1989), especially in light of the magnitude and speed of contemporary anthropogenic changes in global biogeochemical cycles (Lubchenco 1998). From an applied perspective, knowing where species are located during the entire annual cycle is necessary to formulate a basic conservation plan. However, an understanding of the factors influencing survival and reproduction at smaller spatial scales enables a more effective conservation strategy. For example, Marra et al. (1998) demonstrated that the quality of habitats occupied by Setophaga ruticilla individuals during the non-breeding season can affect their physical condition and thus their arrival date on North American breeding ranges, which has consequences for their reproductive success. In this case, knowledge of the life-history consequences of habitat use opens the door to the formulation of more detailed conservation priorities.
Studying the mechanisms that regulate the components of a system also allows patterns to be placed within an evolutionary context. As pointed out by Moore and Aborn (2000), research about habitat selection during migration has historically focused on describing habitat use rather than examining the mechanisms through which selection acts. Thus, to elucidate the processes responsible for observed patterns, it is important to consider the evolutionary history of a species as well as contemporary constraints. Taking just such an approach, Böhning-Gaese and Oberrath (2003) concluded that contemporary habitat preferences of migrants have been strongly influenced by the historical occupation of relatively open habitats in Africa (by ancestors of Holarctic migrants) and of Neotropical forests (by ancestors of Nearctic migrants), as well as by contemporary processes.
Nevertheless, an ability to understand the causes of patterns evident at the population level demands research on processes occurring at the individual level (Koehl 1989). This is because the evolutionary mechanisms that generate population-level migratory patterns originate from variation among individuals. Thus, a focus on the individual is essential for formulating and testing hypotheses about the evolution of migration (Bell 2000). Research focused at the level of the individual requires consideration of the ecological constraints encountered by individuals on different scales throughout their annual cycle. For example, a bird that forages on a scale of hundreds of meters may migrate hundreds of kilometers to search for similar resources at another site. Thus, migratory species are affected by processes occurring on vastly different temporal and spatial scales (e.g., Alerstam and Åkesson 2003). Forming links between these disparate scales is one of the central challenges not only of migrant bird ecology, but of science in general (Levin 1992).
Finally, since migratory behaviour is an attribute regulated by a suite of characters (e.g., physiological, social; Piersma et al. 2005), it is important to formulate hypotheses about these characters within an explicitly phylogenetic context (Zink 2002). In South America, the diversity of movement patterns, even within a single species (e.g., Myiarchus swainsoni; Joseph et al. 2003), may be a complex and long-term response to changing environmental conditions on the continent. However, diverse migratory behaviours can also appear on much shorter time scales. One notable example is the appearance of populations of Hirundo rustica that are beginning to reproduce within their historical non-breeding range, particularly in the coastal zone of the province of Buenos Aires, Argentina (Martínez 1983).
We now have the opportunity to design studies within South America to test theories formulated in other migration systems. With such an approach, we can both enrich our understanding of migration within South America and test the explanatory power of extant theories across migratory systems. We can begin to answer such questions as: do similar ecological, physiological, and genetic mechanisms underlie all migratory systems?; does migration within South America operate under different "rules" than in other systems? For example, the capacity for nocturnal compass orientation is highly conserved phylogenetically in migratory birds around the world (Piersma et al. 2005). In mid- and high-latitudes, migratory birds can use a magnetic inclination compass for orientation, but this system cannot function at equatorial latitudes (Wiltschko and Wiltschko 1995). Thus, what alternative cues can migrants use to orient and navigate in equatorial South America? Clearly, research in this area will allow us to test extant mechanistic theories as well as formulate new hypotheses about the migrations of birds.
In the next section, we use the seasonal overlap of ranges to launch a discussion of how to distinguish between populations of migratory species, as well as how to study the mechanisms operating within these populations.
Range overlap
What patterns characterize bird migration within South America? Stotz et al. (1996) and Chesser (1994) identified several key features: (1) taxonomic composition (in South America, the family Tyrannidae makes up nearly a third of all migratory species on the continent); (2) distance of migration (within South America, most migrants move over shorter distances than do Nearctic-Neotropical migrants); (3) proportion of migratory species along a latitudinal gradient (in South America, the gradient of increasing number of migratory species with latitude is less dramatic than in North-temperate latitudes); and (4) range overlap (approximately two-thirds of Neotropical austral migrants - 159 species - exhibit overlap in population ranges across seasons; i.e., migrants of one population migrate to areas already occupied by conspecifics that do not migrate). Although these four patterns are coarse-grained and described at a continental scale, they offer a point of departure from which to look for more specific patterns (Stotz et al. 1996). We focus on the pattern of range overlap among populations as an interesting pattern to explore the mechanisms that may underlie it.
Several biogeographical explanations have been offered to explain the causes of range overlap. Chesser (1994) offered two hypotheses. First, given that the South American continent is wider towards the equator, birds moving northwards towards the equator after the breeding season could experience a reduction in interspecific competition as a consequence of lower population densities due to the increasing land area, reducing their need to continue migrating northward. Second, there are no evident geophysical barriers to the east of the Andean Cordillera to segregate breeding and non-breeding ranges. In a similar vein, Hayes et al. (1994) proposed that the diminished land area at high latitudes corresponded to a reduced capacity to sustain breeding populations of migratory species, thus producing relatively short migrations between tropical and temperate latitudes.
To test mechanistic hypotheses on the pattern of range overlap, however, we must know something about the variation in migratory distance among individuals, since different strategies among individuals may be present across the species' range. In essence, seasonally overlapping ranges obscure any pattern of exactly where migratory individuals pass the non-breeding season (Stotz et al. 1996). Some populations may even be migrating within the area of overlapping ranges. For example, within any one species, there may be populations that are completely resident, as well as some that are partially migratory (ie., some individuals of a population migrate; Fig. 2). Thus, the area of overlap may "mask" substantial variation in migratory movements among populations. Furthermore, migrant and resident individuals may be partitioning the area of overlap in different ways. For example, Tellería and Pérez-Tris (2004) studied habitat associations in Erithacus rubecula, a European migrant species comprised of migratory populations whose non-breeding distribution overlaps with the distribution of resident populations in the south of the continent. They found that residents and migrants were physiologically different and occupied distinct habitats during the period of overlap. Consequently, in order to study the mechanisms underlying range overlap, it will be important to differentiate between populations and to define their "connectivity": the origin and destination of migratory populations (for a review, see Webster et al. 2002). Once the migratory patterns of populations have been characterized as migratory, resident, or partially migratory, research can turn to the question of which processes underlie their migratory pattern.
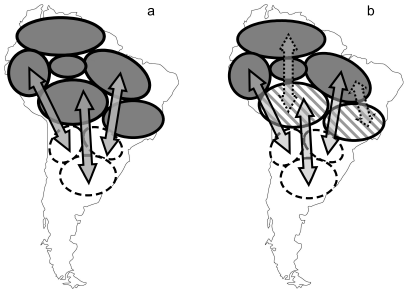
Figure 2. Two hypothetical patterns of population-level migration of Tyrannus melancholicus melancholicus: (a) migration exclusively of populations between a seasonal breeding area and an area of range overlap, and (b) migration of populations between a seasonal breeding area and an area of overlap, as well as partial migration of some populations within the area of overlap. Gray areas: populations of permanent residents, hatched areas: partially migratory populations, white areas: completely migratory populations, dashed arrows: partial migration, closed arrows: complete migration.
Information on connectivity of populations and, at a local scale, on habitat use by different individuals (i.e., migrants vs. residents) has obvious relevance for conservation policy. For example, if one population is in decline in the breeding area, knowledge of where it spends the non-breeding season within the area of overlap can greatly help to pinpoint the part of the life cycle in which the greatest threats to the survival of that population occur.
We now focus on some ideas about how to study mechanisms that operate on an intra-population level and produce population-level patterns. We use partial migration as an example (Fig. 2). For a recent review of other population-level migration patterns, see Boulet and Norris (2006).
Partial migration
Partial migration is perhaps the most common type of bird migration in the world (Berthold 2001). From an evolutionary standpoint, partial migration is thought to be an intermediate step in the evolution of complete migratory behaviour in birds (Berthold 1999). Consequently, studying the processes that produce partial migration could be a key to understanding more about the proximate and ultimate processes that govern migratory movements in species with overlapping ranges.
Before discussing the processes producing partial migration, it is important to distinguish between population-level partial migration and intra-population partial migration. In population-level partial migration, some populations of a species migrate and other populations do not. For example, in Tyrannus savana, the nominate subspecies is a Neotropical austral migrant, while another subspecies (Tyrannus savana sanctaemartae) remains as permanent resident in the northern part of the continent (Chesser 1995, Stiles 2004). This population-level variation in migration characterizes at least 70% of the migratory species in South America (Parker et al. 1996). In the case of intra-population partial migration, some individuals of the same population migrate after the breeding season and others do not (sensu Lack 1943; for a review, see Berthold 2001, Jahn et al. 2004). Differences in migratory behaviour between individuals in the same population have been widely documented in other migratory systems but have not yet been documented in South America. Hereafter, when we refer to "partial migration", we consider specifically intra-population partial migration.
The first studies on partial migration tended to be descriptive or to focus on evolutionary, population-level processes (e.g., Lack 1943, 1954, Kalela 1954, Cohen 1967, von Haartman 1968, Biebach 1983). More recently, emphasis has moved towards identifying and weighing the differences between individuals. Within this context, "migrant" and "resident" are considered to be alternative strategies with different benefits for different individuals in a population (e.g., Swingland 1983). Individual asymmetries (e.g., age, size) are postulated to determine migratory status because of differences in competitive advantage between individuals. Lundberg (1987) contributed to this framework by adding the parameter of frequency-dependent choice, in which individuals of different social rank (e.g., juveniles low in the social hierarchy) decide whether to migrate, depending upon the relative frequencies of the dominant and subdominant individuals in the population with which they have to compete. Indeed, factors such as age, sex and social status in a population have been shown to determine migratory behaviour (e.g., Gauthreaux 1982, Schwabl 1983, Adriaensen and Dhondt 1990). For example, Able and Beltoff (1998) demonstrated that among Carpodacus mexicanus migrating within North America, younger individuals were characterized by a higher tendency to migrate. These demographic and social conditions are often tightly linked to such parameters as competitive ability, physiological tolerance and habitat associations, which in turn are postulated to affect migratory status (Ketterson and Nolan 1983, Cristol et al. 1999, Tellería and Pérez-Tris 2004). Thus, a test of the relationship between these parameters (e.g., physiological tolerance, social dominance) and migratory strategy (e.g., migratory timing, migratory distance) produces mechanistic models on the causes of such patterns as range overlap. It follows that identifying demographic (e.g., age, sex), social (e.g., dominance), and morphological (e.g., body size) differences between individuals is a prerequisite to studying the processes responsible for variation in the migratory behaviour among individuals of a partially migratory population (i.e., whether or not an individual migrates).
What, then, are some specific, testable mechanistic hypotheses that can begin to tease apart potential causes for range overlap? Partial migration is one class of the broader category of differential migration, in which individuals within a population undertake migrations of varying distances (e.g., Ketterson and Nolan 1976). In partial migrants, the distance travelled by some members of the population falls at an extreme of a gradient: zero migration (i.e., residence). Thus, since partial migration is a class of differential migration (Alerstam and Hedenström 1998), much of the theory about differential migration is applicable to partial migration and vice versa. A great variety of theories attempt to explain partial and differential migration, but the majority are variations on three general hypotheses (Bell 2005). The first one is the Dominance Hypothesis. Subdominants are poor competitors for available food; when there are not sufficient resources for all the individuals in a population, subdominants are therefore more likely to migrate in order to avoid competition with dominant individuals. Supporting data are principally indirect, based upon the observation that in different species younger individuals or females are subdominant and migrate longer distances (e.g., Junco hyemalis; Ketterson and Nolan 1976). The second hypothesis is the Arrival Time Hypothesis. Individuals that establish territories at the beginning of the breeding season are less likely to migrate as far because a short migration distance ensures a rapid return to the breeding range and access to the best territories. For example, in a study of Anser caerulescens, Bêty et al. (2004) found a significant relationship between the arrival date on the breeding grounds and the probability of reproducing (although excessively early arrivals suffered from negative climatic effects). Finally, the third hypothesis is the Body Size Hypothesis. In accordance with the relationship between body surface area and volume, larger individuals can better withstand lower temperatures and endure food limitations, giving them a lower probability of migrating away for the winter. For example, in Carpodacus mexicanus, females are socially dominant, but also smaller in size and tend to migrate further than males (Belthoff and Gauthreaux 1991). Studies evaluating these hypotheses have generally failed to produce data that predict migratory distance (e.g., Junco hyemalis; Ketterson and Nolan 1985). Future research in South America could test the explanatory power of these hypotheses in a context independent of the North-temperate system within which the hypotheses were originally formulated.
Future directions
Given the challenge of understanding the mechanisms underlying such a complex system, it is timely to note several themes that may put this type of research in perspective.
Comparisons among common taxa in the New World
Current theory postulates a Neotropical origin for migratory species in the New World, both Nearctic-Neotropical and Neotropical austral migrants (Levey and Stiles 1992, Rappole 1995, Joseph 1997, Chesser and Levey 1998, Joseph et al. 1999, Böhning-Gaese and Oberrath 2003). A review of the data from Parker et al. (1996) reveals that at least 34 families, 56 genera and 23 species have populations of both Nearctic-Neotropical and Neotropical austral migrants. For example, Pyrocephalus rubinus have populations that are Nearctic-Neotropical migrants and other populations that are Neotropical austral migrants (Parker et al. 1996). Intraspecific comparisons of populations employing different strategies (Neotropical austral vs. Nearctic-Neotropical migration) are therefore possible and have the advantage of avoiding the confounding effects of phylogeny that commonly haunt interspecific comparative studies. Specifically, intraspecific research among populations of one species rather than comparisons between species decreases the effects of a shared phylogenetic history, which is problematic because it leads to a lack of independence in the parameters being compared (e.g., Gittleman and Luh 1992, Garland and Adolph 1994).
Conducting comparative studies among migratory systems offers the additional benefit of avoiding the circular logic of attempting to test a hypothesis explaining the causes of a pattern in the same system within which the hypothesis was originally developed. For example, within South America, at least two distinct migratory sub-systems have evolved (Joseph 1997). Extant hypotheses can be evaluated in each sub-system independently.
Available techniques and technologies
The complexity of migration within South America demands an interdisciplinary research approach, incorporating a variety of techniques and technologies (Alerstam and Åkesson 2003, Barlein 2003). What tools can be applied to migration research in South America? New technologies exist that can supply data on diverse temporal scales (between years, months, or days), spatial scales (meters or kilometers), and between levels of biological organization (cellular, organismal, population-level). One particularly exciting new tool is stable isotope analysis, which can be used to determine the origins of migratory individuals (see Hobson 2005 for a review, and Torres Dowdall et al. 2006, in this volume). Base isotope maps do not currently exist for most isotopes throughout the whole of South America, but may be available in the near future.
International collaboration
Because migratory species confront varying constraints throughout the annual cycle (Sillett and Holmes 2002), and since events in one season can exert substantial influence over processes in subsequent seasons (e.g., Marra et al. 1998, Norris et al. 2004), research should be conducted throughout all phases of the annual cycle. This may most easily be accomplished by establishing collaborative international networks of researchers and conservation practitioners (e.g., Barlein 2003). Such associations could standardize methods and share data about the same species and populations. These activities will be essential for advancing the study of patterns and mechanisms (see Stiles 2004 for suggestions and interesting questions), as well as advancing the conservation of populations of migratory species that cross political boundaries.
Research at the individual level
We emphasize the value of collecting individual-specific data (e.g., through colour-banding and genetics). Mechanisms apparent at the population level originate at the individual level; therefore, descriptive studies conducted at the individual level, such as McNeil's (1982) work documenting winter site fidelity in Elaenia parvirostris, are particularly useful to guide the development and testing of mechanistic hypotheses.
Conclusion
Numerous questions are wide-open lines of research - both descriptive and hypothesis-based - in South America. Much progress has been made on other continents about problems that remain unsolved in South America, such that existing bodies of theory, technologies and methods could be readily applied to research on South American migration. Interesting questions concerning the evolution of migration for which we know little in South America include: (1) biogeography (what is the relationship between speciation rates and migratory behaviour across clades?, how does this relationship compare in South America to other migratory systems?); (2) ecology (what is the winter ecology of Neotropical austral migrants?, what is the relationship between migratory timing and biotic vs. abiotic factors?); (3) physiology (do Neotropical austral migrants employ a similar navigation system as migrants in other systems?, what are the energetic constraints to migration in South America relative to other systems?); (4) life-history (which part of the life cycle is most limiting in terms of reproductive success and survival?, are there carry-over effects for reproductive success between seasons?). The answers to such questions will provide a basis upon which to ask more questions and further develop more detailed, mechanistic hypotheses. Given the complexity that characterizes austral migration, our understanding of New World bird migration will be greatly enriched when we undertake a multidisciplinary approach that incorporates both descriptive and theoretical research to elucidate the origins and maintenance of migratory patterns in South America.
Acknowledgements. We thank Víctor Cueto for his invitation to participate in the symposium "Aves migratorias americanas: algunos apuntes para conocerlas" and the organizers of the XI Reunión Argentina de Ornitología for their support. Comments from two anonymous reviewers served to greatly improve the quality of this paper. These ideas are based in great part on the present research of the authors which is financed in part by the National Science Foundation (OISE-0313429, 0612025), American Ornithologists' Union, Wilson Ornithological Society, Western Bird Banding Association, School of Natural Resources and Environment-University of Florida, and Estancia Caparú (Bolivia).
Literature Cited
Able KP and Belthoff JR (1998) Rapid "evolution" of migratory behaviour in the introduced house finch of eastern North America. Proceedings of the Royal Society of London B 265:2063-2071 [ Links ]
Adriaensen F and Dhondt AA (1990) Population dynamics and partial migration of the European Robin (Erithacus rubecula) in different habitats. Journal of Animal Ecology 59:1077-1090 [ Links ]
Alerstam T and Åkesson S (2003) Long-distance migration: evolution and determinants. Oikos 103:247-260 [ Links ]
Alerstam T and Hedenström A (1998) The development of bird migration theory. Journal of Avian Biology 29:343-369 [ Links ]
Barlein F (2003) The study of bird migrations - some future perspectives. Bird Study 50:243-253 [ Links ]
Bell CP (2000) Process in the evolution of bird migration and pattern in avian ecogeography. Journal of Avian Biology 31:258-265 [ Links ]
Bell CP (2005) Inter- and intrapopulation migration patterns: ideas, evidence, and research priorities. Pp. 41-52 in: Greenberg R and Marra PP (eds) Birds of two worlds. The ecology and evolution of migration. Johns Hopkins University Press, Baltimore [ Links ]
Belthoff JR and Gauthreaux SA (1991) Partial migration and differential winter distribution of House Finches in the eastern United States. Condor 93:374-382 [ Links ]
Berthold P (1999) A comprehensive theory for the evolution, control and adaptability of avian migration. Ostrich 70:1-11 [ Links ]
Berthold P (2001) Bird migration: a general survey. Second edition. Oxford University Press, New York [ Links ]
Bêty J, Giroux JF and Gauthier G (2004) Individual variation in timing of migration: causes and reproductive consequences in greater snow geese (Anser caerulescens atlanticus). Behavioral Ecology and Sociobiology 57:1-8 [ Links ]
Biebach H (1983) Genetic determination of partial migration in the European Robin (Erithacus rubecola). Auk 100:601-606 [ Links ]
Bildstein K (2004) Raptor migration in the Neotropics: patterns, processes and consequences. Ornitología Neotropical 15(Suppl.):83-99 [ Links ]
Böhning-Gaese K and Oberrath R (1999) Phylogenetic effects on morphological, life-history, behavioral, and ecological traits of birds. Evolutionary Ecology Research 1:347-364 [ Links ]
Böhning-Gaese K and Oberrath R (2003) Macroecology of habitat choice in long-distance migratory birds. Oecologia 137:296-303 [ Links ]
Boulet M and Norris DR (2006) The past and present of migratory connectivity. Ornithological Monographs 61:1-13 [ Links ]
Capllonch P and Lobo R (2005) Contribution to the knowledge of migration of three Elaenia species (Tyrannidae) from Argentina. Ornitología Neotropical 16:145-161 [ Links ]
Chesser RT (1994) Migration in South America: an overview of the Austral system. Bird Conservation International 4:91-107 [ Links ]
Chesser RT (1995) Biogeographic, ecological, and evolutionary aspects of South American austral migration, with special reference to the family Tyrannidae. PhD dissertation, Louisiana State University, Louisiana [ Links ]
Chesser RT and Levey DJ (1998) Austral migrants and the evolution of migration in New World birds: diet, habitat and migration revisited. American Naturalist 152:311-319 [ Links ]
Cohen D (1967) Optimization of seasonal migratory behavior. American Naturalist 101:5-17 [ Links ]
Cristol DA, Baker MB and Carbone C (1999) Differential migration revisited: latitudinal segregation by age and sex class. Current Ornithology 15:33-88 [ Links ]
Garland T Jr and Adolph SC (1994) Why not to do two-species comparative studies: limitations on inferring adaptation. Physiological Zoology 67:797-828 [ Links ]
Gauthreaux SA Jr (1982) The ecology and evolution of avian migration systems. Pp. 93-168 in: Farner DS and King JR (eds) Avian biology. Volume 6. Academic Press, New York [ Links ]
Gittleman JL and Luh HK (1992) On comparing comparative methods. Annual Review of Ecology and Systematics 23:383-404 [ Links ]
von Haartman L (1968) The evolution of resident versus migratory habit in birds: some considerations. Ornis Fennica 45:1-7 [ Links ]
Hayes FE, Scharf PA and Ridgely RS (1994) Austral bird migrants in Paraguay. Condor 96:83-97 [ Links ]
Hobson KA (2005) Stable isotopes and the determination of avian migratory connectivity and seasonal interactions. Auk 122:1037-1048 [ Links ]
Jahn AE, Davis SE and Saavedra AM (2002) Patrones en la migración austral de aves entre temporadas y hábitats en el Chaco boliviano, con notas de observaciones raras y una lista de especies. Ecología en Bolivia 37:31-50 [ Links ]
Jahn AE, Levey DJ and Smith KG (2004) Reflections across hemispheres: a system-wide approach to New World bird migration. Auk 121:1005-1013 [ Links ]
Joseph L (1997) Towards a broader view of Neotropical migrants: consequences of a re-examination of austral migration. Ornitología Neotropical 8:31-36 [ Links ]
Joseph L, Lessa EP and Christidis L (1999) Phylogeny and biogeography in the evolution of migration: shorebirds of the Charadrius complex. Journal of Biogeography 26:329-342 [ Links ]
Joseph L, Wilke T and Alpers D (2003) Independent evolution of migration on the South American landscape in a long-distance temperate-tropical migratory bird, Swainson's flycatcher (Myiarchus swainsoni). Journal of Biogeography 30:925-937 [ Links ]
Kalela O (1954) Populationsökologische gesichtspunkte zur entstehung des vogelsugs. Annales Botanici Societatis Zoologicae Botanicae Fennicae Vanamo 16:1-31 [ Links ]
Ketterson ED and Nolan V Jr (1976) Geographic variation and its climatic correlates in the sex ratio of eastern-wintering Dark-eyed Juncos (Junco hyemalis). Ecology 57:679-693 [ Links ]
Ketterson ED and Nolan V Jr (1983) The evolution of differential bird migration. Current Ornithology 1:357-402 [ Links ]
Ketterson ED and Nolan V Jr (1985) Intraspecific variation in avian migration: evolutionary and regulatory aspects. Contributions in Marine Science 27(Suppl.):553-579 [ Links ]
Koehl MR (1989) Discussion: from individuals to populations. Pp. 39-53 in: Roughgarden J, May RM and Levin SA (eds) Perspectives in ecological theory. Princeton University Press, Princeton [ Links ]
Lack D (1943) The problem of partial migration. British Birds 37:122-131 [ Links ]
Lack D (1954) The natural regulation of animal numbers. Oxford University Press, London [ Links ]
Levey DJ (1994) Why we should adopt a broader view of Neotropical migrants. Auk 111:233-236 [ Links ]
Levey DJ and Stiles FG (1992) Evolutionary precursors of long-distance migration: resource availability and movement patterns in Neotropical landbirds. American Naturalist 140:447-476 [ Links ]
Levin SA (1992) The problem of pattern and scale in ecology. Ecology 73:1943-1967 [ Links ]
Lubchenco J (1998) Entering the century of the environment: a new social contract for science. Science 279:491-497 [ Links ]
Lundberg P (1987) Partial bird migration and evolutionarily stable strategies. Journal of Theoretical Biology 125:351-360 [ Links ]
Marra PP, Hobson KA and Holmes RT (1998) Linking winter and summer events in a migratory bird by using stable-carbon isotopes. Science 282:1884-1886 [ Links ]
Martínez MM (1983) Nidificación de Hirundo rustica erythrogaster (Boddaert) en la Argentina (Aves, Hirundinidae). Neotropica 29:83-86 [ Links ]
McNeil R (1982) Winter resident repeats and returns of austral and boreal migrant birds banded in Venezuela. Journal of Field Ornithology 53:125-132 [ Links ]
Moore FR and Aborn DA (2000) Mechanisms of en route habitat selection: how do migrants make habitat decisions during stopover? Studies in Avian Biology 20:34-42 [ Links ]
Morton ES (1977) Intratropical migration in the Yellow-green Vireo and Piratic Flycatcher. Auk 94:97-106 [ Links ]
Norris DR, Marra PP, Kyser TK, Sherry TW and Ratcliffe LM (2004) Tropical winter habitat limits reproductive success on the temperate breeding grounds in a migratory bird. Proceedings of the Royal Society of London B 271:59-64 [ Links ]
Outlaw DC, Voelker G, Mila B and Girman DJ (2003) Evolution of long-distance migration in and historical biogeography of Catharus thrushes: a molecular phylogenetic approach. Auk 120:299-310 [ Links ]
Parker TA III, Stotz DF and Fitzpatrick JW (1996) Ecological and distributional databases. Pp. 113-436 in: Stotz DF, Fitzpatrick JW, Parker TA III and Moskovits KD (eds) Neotropical birds: ecology and conservation. University of Chicago Press, Chicago [ Links ]
Piersma T, Pérez-Tris J, Mouritsen H, Bauchinger U and Bairlein F (2005) Is there a "migratory syndrome" common to all migrant birds? Annals of the New York Academy of Sciences 1046:282-293 [ Links ]
Rappole JH (1995) The ecology of migrant birds: a Neotropical perspective. Smithsonian Institution Press, Washington DC [ Links ]
Ridgely RS and Tudor G (1994) The birds of South America. Volume II: the suboscine passerines. University of Texas Press, Austin [ Links ]
Schwabl H (1983) Ausprägung und Bedeutung des Teilzugverhaltens einer südwestdeutschen Population der Amsel Turdus merula. Journal für Ornithologie 124:101-116 [ Links ]
Schwabl H and Silverin B (1990) Control of partial migration and autumnal behaviour. Pp. 144-155 in: Gwinner E (ed) Bird migration: physiology and ecophysiology. Springer-Verlag, Berlin [ Links ]
Sillett T and Holmes R (2002) Variation in survivorship of a migratory songbird throughout its annual cycle. Journal of Animal Ecology 71:296-308 [ Links ]
da Silva JMC (1999) Seasonal movements and conservation of seedeaters of the genus Sporophila in South America. Studies in Avian Biology 19:272-280 [ Links ]
Stiles FG (2004) Austral migration in Colombia: the state of knowledge, and suggestions for action. Ornitología Neotropical 15(Suppl.):349-355 [ Links ]
Stotz DF, Fitzpatrick FW, Parker TA III and Moskovits DK (1996) Neotropical birds: ecology and conservation. University of Chicago Press, Chicago [ Links ]
Swingland IR (1983) Intraspecific differences in movements. Pp. 102-115 in: Swingland IR and Greenwood PJ (eds) The ecology of animal movement. Clarendon Press, Oxford [ Links ]
Tellería JL and Pérez-Tris J (2004) Consequences of the settlement of migrant European Robins, Erithacus rubecula, in wintering habitats occupied by conspecific residents. Ibis 146:258-268 [ Links ]
Torres Dowdall J, Farmer A and Bucher EH (2006) Uso de isótopos estables para determinar conectividad migratoria en aves: alcances y limitaciones. Hornero 21:73-84 [ Links ]
Vuilleumier F (2004) A critique of Neotropical ornithology: is research on Neotropical birds scientific? Ornitología Neotropical 15:41-60 [ Links ]
Webster MS, Marra PP, Haig SM, Bensch S and Holmes RT (2002) Links between worlds: unraveling migratory connectivity. Trends in Ecology and Evolution 17:76-83 [ Links ]
Wiltschko R and Wiltschko W (1995) Magnetic orientation in animals. Springer, Berlin [ Links ]
Winker K (2000) Migration and speciation. Nature 404:36 [ Links ]
Winker K, Escalante P, Rappole JH, Ramos MA, Oehlenschlager RJ and Warner DW (1997) Periodic migration and lowland forest refugia in a "sedentary" Neotropical bird, Wetmore's Bush-Tanager. Conservation Biology 11:692-697 [ Links ]
Winker K and Pruett CL (2006) Seasonal migration, speciation, and morphological convergence in the genus Catharus (Turdidae). Auk 123:1052-1068 [ Links ]
Zink R (2002) Towards a framework for understanding the evolution of avian migration. Journal of Avian Biology 33:433-436 [ Links ]

