










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEl hornero
versión impresa ISSN 0073-3407versión On-line ISSN 1850-4884
Hornero v.21 n.2 Buenos Aires ago./dic. 2006
Annual survival of Red Knots (Calidris canutus rufa) using the San Antonio Oeste stopover site is reduced by domino effects involving late arrival and food depletion in Delaware Bay
Patricia M. González 1,4, Allan J. Baker 2,3 and María Eugenia Echave 1
1 Fundación Inalafquen. Roca 135, 8520 San Antonio Oeste, Río Negro, Argentina.
2 Royal Ontario Museum. 100 Queen's Park, Toronto, ON M5S 2C6, Canada.
3 Department of Zoology, University of Toronto. Toronto, Canada.
4 ccanutus@yahoo.com.ar
Abstract
Ecological conditions in breeding and non-breeding areas of migrant birds have been linked to their annual survival and production of young, but the role of stopover sites is under-appreciated. Through banding studies and censuses along the flyway from Tierra del Fuego to the Canadian Arctic, the drastic decline in 2000-2001 of Red Knots (Calidris canutus rufa) population summering in southern South America in the northern winter was shown to be related to the overharvesting of horseshoe crabs (Limulus polyphemus) in Delaware Bay, USA, their last stopover site before reaching their breeding grounds, and to the late arrival of the birds at this site. In San Antonio Oeste, Argentina, where 25-50% of the Tierra del Fuego Red Knots population congregates every northward migration season, annual survival of the cohort of experienced birds banded in March 1998 was impacted a year later than the general decline. Knots marked at San Antonio Oeste earlier in March arrived in Delaware Bay on average before those marked 15 days later. Additionally, early migrating knots with active body moult in San Antonio Oeste exhibited a higher return rate in the following years than late and non-moulting birds. Since the decline, birds arriving late in Delaware Bay have been at increased risk of not being able to refuel properly or on time because food is no longer superabundant at that stopover site. These domino effects indicate that there are fitness consequences to individual migration strategies adopted by birds at austral summering and stopover sites, which can be amplified by compressed timing in Delaware Bay when food is depleted at this final stopover site.
Key words: Calidris canutus, domino effects, migration, population decline, Red Knot, stopover ecology.
Resumen
Reducción de la supervivencia anual del Playero Rojizo (Calidris canutus rufa) en su escala migratoria de San Antonio Oeste, Argentina, por efectos dominó de llegada tardía y depresión del recurso trófico en Bahía Delaware
Si bien se ha relacionado la condición de las áreas de estadía no reproductiva y reproductiva de las aves migratorias con su supervivencia y producción de crías, el papel de los sitios de escala como limitante del tamaño poblacional es escasamente conocido. Mediante estudios de anillado y censos a lo largo de la ruta de vuelo desde Tierra del Fuego hasta el Ártico de Canadá, hemos relacionado la drástica declinación de la población de Playero Rojizo (Calidris canutus rufa) ocurrida durante 2000-2001 con la sobrepesca del cangrejo herradura (Limulus polyphemus) en su última escala en la migración hacia el norte (Bahía Delaware, EEUU) y la llegada tardía de las aves. En San Antonio Oeste, Argentina, donde se congrega el 25-50% de la población de Tierra del Fuego durante la migración al norte, el seguimiento de aves experimentadas de la cohorte anillada en marzo de 1998 permitió estimar que su supervivencia fue afectada un año más tarde que la declinación general. Las aves marcadas más temprano en marzo llegaron antes a Bahía Delaware, en promedio, que las marcadas 15 días después. Además, las aves tempranas con presencia de muda activa de plumaje corporal mostraron una tasa de retorno significativamente mayor en años posteriores que las aves sin muda activa. Desde la declinación, las aves tardías incrementaron el riesgo de no acumular reservas apropiadamente o a tiempo, debido a que el recurso trófico dejó de ser superabundante en Bahía Delaware. Estos efectos dominó indican que las estrategias de migración individuales originadas en las áreas de estadía austral y en los sitios de escala migratoria tienen consecuencias sobre la adecuación biológica y que éstas pueden ser amplificadas por la reducción en el tiempo de estadía en Bahía Delaware cuando el recurso trófico es escaso en este sitio.
Palabras clave: Calidris canutus, declinación poblacional, ecología de escala migratoria, efecto dominó, migración, Playero Rojizo.
Received 10 March 2006, accepted 29 December 2006
The role of the condition of birds in non-breeding staging areas, breeding sites, or both in limiting long-term or year-to-year population sizes in migrants has been the subject of many studies in different bird taxa, especially in shorebirds (e.g., Evans and Pienkowski 1984, Evans et al. 1984, Pienkowski and Evans 1985, Newton 2004). In some species, habitats occupied in wintering areas (also known as "austral summering areas" in the Southern Hemisphere) and migration flyways, and their associated food supplies, can influence the body condition, migration dates and subsequent breeding success of migrants (Marra et al. 1998, Drent et al. 2003). Similarly, the numbers of young produced in one region could, through density-dependent processes, affect subsequent overall mortality in another region (e.g., Goss-Custard et al. 1995). Thus, events in breeding, migration and "wintering areas" are interdependent in their effects on bird numbers (reviewed in Newton 2004). Although less is known about long distance migrants, we hypothesize that population size might be limited by intra- or interspecific competition at key stopover sites if they face low food availability, higher predation risk, and increased disturbance or poor quality roosting places.
One of the best known shorebird species is the Red Knot Calidris canutus (Scolopacidae) which has a circumpolar breeding distribution in the Northern Hemisphere comprised of six discrete populations that are recognized as subspecies on the basis of differences in morphometrics and plumage (Piersma et al. 2005). Of these subspecies, Calidris canutus rufa travels the longest migration of about 16000 km twice a year between their breeding grounds in the Arctic and their summering (= northern wintering areas) sites in Tierra del Fuego and Patagonian Argentina (Morrison and Harrington 1992, Piersma and Davidson 1992, Harrington 2001, Tomkovich 2001). During their journeys, they congregate at the scarce wetlands extensive enough to support large flocks of birds. Such wetlands occur thousands of kilometres apart in the Atlantic flyway, like San Antonio Oeste at Río Negro Province in Argentina, Rio Grande do Sul and Maranhão in Brazil, and Delaware Bay in USA (Fig. 1).
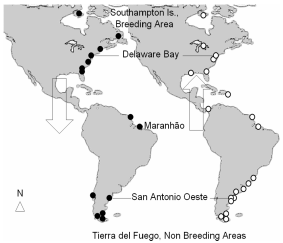
Figure 1. Red Knot flyway depicted on the basis of resightings of individuals colour-banded in Argentina from 1995 onwards. Black dots indicate resighting places during southern migration and white dots during northward migration. Key stopover study sites are indicated (González et al., unpublished data).
The majority of adults in the Tierra del Fuego population make a stopover in Delaware Bay every May, and mix with separate populations that spend the non-breeding season in Maranhão in northern Brazil and Florida, respectively (Atkinson et al. 2005, Baker et al. 2005). Until 2000 Red Knots and other migrant shorebirds fed almost exclusively on a superabundant supply of eggs of spawning horseshoe crabs (Limulus polyphemus) (Castro and Myers 1993, Tsipoura and Burger 1999), enabling them to store nutrients quickly and almost double their body mass for the final leg of migration to the Arctic breeding grounds. Extra stores are carried not only so that the birds can survive poor weather or lack of food after arrival, which can cause high levels of mortality (Morrison 1975, 2006, Boyd 1992), but also to enable the birds to undergo a series of physiological transformations from a state suitable for migration to one for breeding (Morrison et al. 2005). Beginning in 1990 and peaking in 1995-1996, there was a dramatic increase in commercial fishing to provide bait for eel and conch fisheries (Walls et al. 2002), which resulted in a six-fold decline in the numbers of horseshoe crabs caught in survey trawls in Delaware Bay (S. Michels, unpublished data; cited in Andres 2003). Since 2000, crab eggs are no longer superabundant in Delaware Bay.
Several lines of evidence suggest linkages between events at stopover sites in San Antonio Oeste and Delaware Bay and the breeding grounds. First, studies that captured and colour-banded northbound Red Knots (9851 individuals) in Delaware Bay during each May from 1997 to 2002 showed that fewer Red Knots reached threshold departure masses of 180-200 g (Baker et al. 2004). As the abundance of crab eggs declined from 1997-1998 to 2001-2002, the predicted proportion of well-conditioned knots (200 g or greater) in Delaware Bay near departure time on 28 May decreased significantly by 70% (Baker et al. 2004). In the 2-3 days before the peak departure for the Arctic, mean body masses declined significantly from 183 g in 1997 to 162 g in 2002. Return rates of northbound adults caught in Delaware Bay also showed that Red Knots known to survive to a later year by being recaptured or resighted throughout the flyway were significantly heavier at initial capture than birds never seen again (Baker et al. 2004). Second, emerging evidence suggests that northbound birds from Tierra del Fuego may be arriving in Delaware Bay later in the spring than in earlier years, and that late birds are increasingly delaying departure for the breeding grounds (Baker et al. 2004). Late arrival on the breeding grounds often confers a strong reproductive disadvantage; late arrivals are predicted to have a lower probability of surviving and producing offspring if they attempt to breed. Resights of colour-banded birds showed that arrival time of the Tierra del Fuego birds in Delaware Bay relative to the more northerly summering birds in Florida and Maranhão have differed over time, although in most years the former on average arrive later in Delaware Bay than birds from northern Brazil or Florida. In 2000 and 2001 the highest proportion of Tierra del Fuego birds occurred at or after the peak aerial count, meaning that Florida and Maranhão birds may have departed earlier, or there were more late arrivals from Tierra del Fuego, or both (Baker et al. 2004).
Fitness consequences of reduced adult survival and recruitment are consistent with the alarming decline in population size of Red Knots in Tierra del Fuego from 51000 to 27000 in 2000-2002 (González et al. 2004, Morrison et al. 2004); such a rapid decline may seriously threaten the viability of this subspecies. Annual survival of adult birds at Delaware Bay arriving from Tierra del Fuego and northern South America decreased by 37% between May 2000 and May 2001. In addition, annual survival estimated from captured and marked birds (3644 individuals) from 1995 to 2003 in San Antonio Oeste and in Tierra del Fuego declined significantly from an average of 85% in the three migration years from 1994-1995 to 1997-1998 to 56% in the ensuing three-year period to 2000-2001. Recruitment, as measured by the proportion of second-year birds, comprised 19% of annual catches of 500-600 Knots in Tierra del Fuego in 1995, 16% in 2000 and 10% in 2001 (Baker et al. 2004).
In this paper we examine the role of two key stopover sites in the flyway of Calidris canutus rufa and assess the possible flow-on or domino effects (Piersma 1987) that can accrue after the birds leave the austral summering sites in Tierra del Fuego and migrate northwards en route to the breeding grounds. These sites are (1) San Antonio Oeste in Argentina, the main stopover site on the coast of Patagonia for Red Knots, where most adults undergo body moult into breeding plumage before undertaking long flights northwards, and (2) Delaware Bay in USA, the critical final spring refuelling site where they must accumulate large stores of nutrients before departing for the breeding grounds in the Arctic. San Antonio Oeste is located at 40°45'S, 64°55'W in Río Negro Province of Argentina, and hosts 25-50% of the total population from Tierra del Fuego during northward migration from late January to April (González et al. 2004). Here the primary food for Red Knots during the day (Sitters et al. 2001) is the mussel Brachidontes rodriguezi that they find on a rocky intertidal habitat locally known as "restinga" (González et al. 1996). Specifically, our objectives were to investigate whether habitat conditions and the timing of migration at a stopover site in the southern end of the flyway might be exacerbated by the declining food supplies in Delaware Bay, and the flow-on fitness consequences of decreased refuelling rates on late arriving Red Knots at this penultimate staging site.
Methods
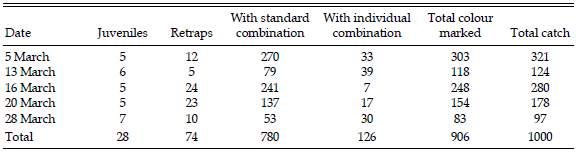
We analyzed arrival time of Red Knots in Delaware Bay in relation to their banding time in San Antonio Oeste. As part of an international research project on Red Knot populations throughout the flyway, a banding expedition to San Antonio Oeste was organized in 1998 (Baker et al. 1999, 2001, Piersma et al. 2005), during which knots were caught with cannon nets in five catches on 5, 13, 16, 20 and 28 March during the peak of migration (n = 1000, Table 1). Age class (juvenile or adult), body mass, body moult activity on the breast, and percentage of breeding plumage were recorded on captured birds. In addition to a metal numbered band most knots (n = 780) were banded with standard combinations of two colour bands and a coloured flag to identify "time cohorts" (distinctive combinations for 5 and 13 March, 16 and 20 March, and 28 March), while 126 birds were marked with individual schemes of orange flag and four colour bands.
Table 1. Details of Red Knots catches (number of individuals) made at San Antonio Oeste, Río Negro, Argentina, in March, 1998. Retraps indicates knots banded previously in this or other expeditions. Analyses were based only on the 906 colour-banded birds.
In May 1998 we compared the return rates in Delaware Bay of birds previously colour-banded at San Antonio Oeste during the first half ("early cohort": knots colour-banded during 5 and 13 March) and the second half ("late cohort": knots colour-banded during 16 and 20 March) of March 1998. To keep resighting probabilities approximately equal for the two time cohorts the sample from 28 March was not included in the analysis. Three observers regularly carried out scans of the flocks of Red Knots at different sites on the shores of Delaware Bay in New Jersey and Delaware and recorded the numbers of banded individuals relative to the numbers of unbanded birds. To avoid any biases introduced by the visual attraction of colour-banded birds, we made sure that all observed birds received scores as we examined as many different parts of feeding flocks as possible. We analyzed the data for each time cohort using Binary Logistic Regression with dependent variable (1: banded; 0: non-banded) on independent categorical variables (shore, observer) and day as a continuous variable.
In subsequent years from 1999 to 2003, resighting efforts of individually colour-banded adults at San Antonio Oeste were used to estimate annual apparent survival of the "1998 year cohort". The data conformed to Cormack-Jolly-Seber assumptions according to Choquet et al. (2001). Survival analyses were run in Mark 3.2 (White and Burnham 1999) using Cormack-Jolly-Seber models by cohort. Model selection was based on the Akaike Information Criterion. Binary Logistic Regression with logit link and sigma-restricted parameterization was used to analyze the return rate in the six following years (1: seen again, 0: not seen again) from recapture and resighting of banded birds (n = 228) related to the presence of active body moult, percentage of breeding plumage (categorical independent variables), body mass and day of initial capture (continuous independent variables).
Results
Arrival times in Delaware Bay
Return rates in Delaware Bay estimated from resightings or recaptures of both early and late cohorts of Red Knots banded in San Antonio Oeste in 1998 (n = 19884) increased through the season (Day b = 0.064, SE = 0.02, P < 0.001; Fig. 2), either because birds from the unbanded Maranhão or Florida summering populations were leaving or more banded knots from San Antonio Oeste were present at the end of the season, or both. Despite the similar numbers of birds banded at San Antonio Oeste in both time cohorts, the late cohort had a lower return rate than the earlier banded cohort (Fig. 2), indicating that the late cohort was on average arriving later in Delaware Bay, that at least some birds had not reached the bay by the end of May, or that a higher proportion of the late cohort used sites other than Delaware Bay, or a combination of these factors. Red Knots from San Antonio Oeste were more represented on the shores of Delaware than New Jersey (Shore b = 0.36, SE = 0.10, P < 0.0001), but the same trend in true detection rates was apparent in both states.
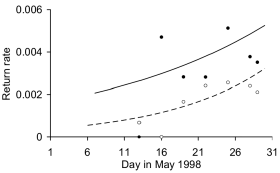
Figure 2. Return rates in Delaware Bay of "early" (filled circles) and "late" (open circles) cohorts of Red Knots banded at San Antonio Oeste, Río Negro, Argentina. The time dependent model of the return rate of colour-banded knots during the first half of March 1998 (early cohort, solid line; n = 421, b = 0.049, SE = 0.024, P < 0.043) is compared with the model for those banded in the second half of March 1998 (late cohort, dashed line; n = 402, b = 0.092, SE = 0.034, P < 0.007). Number of checked birds was 19884.
Apparent survival
Because we have shown previously (Baker et al. 2004) that a two time period model (1998-1999 to 2000-2001) was the best-fitting model for the Tierra del Fuego population, we compared this model for the 1998 San Antonio Oeste cohort with another that allowed a one year lag in survival effects between the periods 1998-2001 and 2001-2002. We also computed standard time dependent and constant survival and resighting probability models to check for goodness of fit. The model with the one year lag in the decline of annual survival and with a constant resighting probability was the best model (Table 2A). Annual survival of the San Antonio Oeste 1998 cohort was estimated to drop from 80.3% between 1998 and 2001 to 65.9% between 2001 and 2002, one year after the general population passing through Delaware Bay had suffered a similar drop in annual survival (Table 2B). Although the 95% confidence intervals of the estimates of annual survival in the San Antonio Oeste 1998 cohort model before and after 2001 partly overlap because of the relatively small size of the colour-banded 1998 cohort (n = 126), there is no indication of a decline in annual survival before 2001.
Table 2. Apparent survival of the San Antonio Oeste 1998 cohort from Cormack-Jolly-Seber capture-recapture analysis: (A) model selection, (B) real function parameters of the best-fitting model for individually colour-banded Red Knots. AIC: Akaike Information Criterion, φ: apparent survival, p: probability of recapture or resighting, (.): constant model, (t): time dependent model, t1: 1998-2001, t2: 2001-2002, t3: 1998-2000, t4: 2000-2002. Model selection based on correction for over-dispersion with c-hat = 1.509.

Variables explaining return rates
The best logistic regression model for return rates of San Antonio Oeste 1998 knots according to the Akaike Information Criterion included the effect of both day of initial capture and presence of body moult they were undergoing (P < 0.0001). The probability for a 1998 cohort bird to be seen in the following years was negatively correlated (b = -0.090, SE = 0.025) with day of initial capture (P < 0.0001), indicating that birds captured late in March were less likely to be seen again than those captured in the early half of the month. Red Knots in body moult had a higher probability of being seen in future years than birds that were not moulting (b = 4.43, SE = 0.277, P < 0.0001), thereby relating the condition of the bird to an indirect measure of survival. Although percentage of breeding plumage and body mass are indices of body condition and thus we would expect that redder-plumaged and heavier birds exhibit a higher probability of survival, these variables are collinear (positively correlated) with day of initial capture through the season. Thus, a model with percentage of breeding plumage and presence of body moult showed that redder knots had a lower likelihood of being seen in following years (b = -0.412, SE = 0.16, P < 0.01; number of birds seen again = 57, number of birds not seen again = 171). A model including only body mass and presence of body moult as independent variables was not significant for body mass because late cohort knots were heavier than early cohort birds (128.0 ± 0.9 g, n = 122, and 123.7 ± 1.0 g, n = 106, respectively, mean ± SE; ANOVA: F1, 226 = 13.66, P < 0.0003). However, moulting birds were heavier on average than non-moulting birds (127.2 ± 0.8 g, n = 191, and 122.5 ± 1.7 g, n = 37, respectively; ANOVA: F1, 226 = 6.26, P < 0.0130). These results suggest that later captured Red Knots stayed later at the San Antonio Oeste stopover site to gain mass and complete the acquisition of redder breeding plumage than did earlier captured Red Knots.
Discussion
Domino effects at stopover sites
Baker et al. (2004) argued that food supplies at Delaware Bay, the last stopover site before the flight to the High Arctic breeding grounds, limited the Patagonian wintering population of Red Knots. The new analyses presented here further suggest that different segments of the population may be differently affected by staging site problems; in particular, the late migration strategy adopted by some individuals is now associated with greater risks of mortality than in the past. Long-distance migrants from the Red Knot population in southern South America are more restricted in their timing of migration into Delaware Bay than are northern populations. On average they arrive later in Delaware Bay than do northerly wintering knots as shown here for the migration season of 1998, when conditions in Delaware Bay were good (see also Baker et al. 2004). In the years of severe food limitation that occurred in Delaware Bay after 2000, we predicted that Red Knots would suffer potentially drastic consequences on survival and recruitment. Prior to 2000, knots refuelling in Delaware Bay departed en masse for the Arctic in the period May 28-30, but as the supply of horseshoe crab eggs was depleted in subsequent years more birds have been delaying their departure by 7-14 days (Baker et al. 2004, pers. obs.). A domino effect between the two stopover sites in San Antonio Oeste and Delaware Bay has thus appeared and been exacerbated mainly by poor refuelling conditions in the final stopover site. Drent et al. (2003:274) emphasized the "critical role of the final take-off site" of Pink-footed Geese (Anser brachyrhynchus) as suggested by a positive relationship between en route body condition and subsequent breeding success.
Fitness consequences of individual migration strategies
The migration schedules of the austral summering population in Tierra del Fuego potentially could have fitness consequences, as northbound knots arriving later at their first southern stopover site in San Antonio Oeste on average also arrive later in Delaware Bay and thus have compressed refuelling time and later departure for the breeding grounds. However, arrival timing in San Antonio Oeste could be related to habitat condition in Tierra del Fuego or differences in individual strategies of Red Knots or both, rather than to a limitation from poor quality stopover. This is because day of initial capture at San Antonio Oeste does not necessarily reflect departure date, as some earlier arrivals left in a few days while others remained until the end of the season in April, and thus late arrivals might depart together with some early birds (González et al., unpublished data). The rate of storage of nutrients at the San Antonio Oeste stopover site is around 0.5 g/day (Piersma et al. 2005), which is very low compared with the average 4.6 g/day that knots achieve at Delaware Bay, the highest recorded among the world's subspecies and staging sites (Piersma et al. 2005). Thus, birds departing from San Antonio Oeste in early March are likely to have lower body mass than those that remain until the middle of April. Birds that are minimizing the cost of energy transport are expected to stop at all useful sites along the route to carry the smallest possible fuel loads (Gudmundsson et al. 1991, Alerstam and Hedenström 1998); while our results suggest that early cohort knots in San Antonio Oeste are following this general strategy, late cohort knots in contrast seems to adjust to time-selected migration, where birds should accrue substantial fuel loads to minimize migration time to Delaware Bay.
These differences in individual migratory strategies indicate why body mass could not explain the likelihood of return rates at San Antonio Oeste, whereas at Delaware Bay it is a significant explanatory variable. While Red Knots at Delaware Bay appear more synchronized in migration timing for refuelling and departures because the Arctic breeding grounds are only available during a short period in the year, at San Antonio Oeste Red Knots have a broader window of time where individual strategies can be employed.
Apparent survival estimates for the San Antonio Oeste 1998 cohort indicate that birds in this cohort did not suffer the decline that the average adult passage population in Delaware Bay did between 2000-2001, but instead had a lag until one year later when they had a significant drop in survival. We interpret this to mean that early migrating and experienced birds in the San Antonio Oeste 1998 cohort (whose survivors would have been older than the general population in 2000-2001) were able to avoid mortality better than the general population in Tierra del Fuego which includes younger adult birds, and only were affected late in the population decline.
Role of stopover sites in population limitation
The importance of the last stopover as a refuelling site before departing for the breeding grounds and of late arrival has been shown to have fitness consequences in birds (Alerstam and Hedenström 1998, Madsen 2001, Drent et al. 2003, Morrison 2006). Morrison (2006) found that Calidris canutus islandica departing from their last stopover site in Iceland in better than average condition had a higher probability of being seen again following a series of years with difficult weather conditions in the Arctic. This shows that being in superior condition was linked with higher survival; in this case conditions were normal at the final stopover area but the birds encountered unusually difficult conditions on the breeding grounds. The situation in Delaware Bay again demonstrates the link between condition and survival; birds were unable to reach suitable departure condition because of a reduction in the available food supplies resulting in reduced survival. In the Wadden Sea, reduction in food stocks for shorebirds due to overharvesting in the cockle fishery has resulted in a concomitant reduction in bird numbers, providing another example of how human activities can severely impact population sizes in migratory shorebirds (Stroud et al. 2006). Under conditions of food depletion in Delaware Bay there can be a domino effect from the timing of refuelling in southern stopover sites in South America like San Antonio Oeste, as birds arriving late in the USA will then be delayed further in their departure or be underfuelled, resulting in overall lower breeding success and increased mortality in the population. This emphasizes the migratory connectivity of populations of knots at these and other sites as well as the role of individual migration strategies. An integrated flyway-wide approach to management and recovery is required for effective conservation of this rapidly declining population. In the absence of effective management at sites throughout the flyway we can expect the worldwide decline in shorebird numbers to continue, and face the grim prospect of extinction of populations or species at an accelerating rate.
Acknowledgements. We acknowledge financial support from the Royal Ontario Museum Foundation and the Natural Sciences and Engineering Research Council of Canada, NOAA, US Fish and Wildlife Service and the Canadian Wildlife Service. We thank N. Garfield and B. Archer for supporting our work in South America, and Birders Exchange of American Birding Association for donating binoculars and telescopes. We thank CODEMA in Río Negro, Prefectura Naval Argentina, and the Baraschi and Echave families for providing logistical support in San Antonio Oeste, and DNREC and NJFWS in Delaware Bay. We thank Theunis Piersma and Guy Morrison for many helpful suggestions which improved the manuscript, and two anonymous referees for their constructive comments. The authors owe a debt of gratitude to numerous researchers, volunteers, teachers and students, especially to T. Piersma, P. de Goeij, D. Price, P. Ireland, H. Sitters, M. Carbajal, G. Escudero, G. Murga, R. I. G. Morrison, R. Pissaco, L. Benegas and G. Guerrero who made it possible to catch, process and resight knots in Argentina.
Literature Cited
Alerstam T and Hedenström A (1998) The development of bird migration theory. Journal of Avian Biology 29:343-369 [ Links ]
Andres BA (2003) Delaware Bay shorebird-horseshoe crab assessment report - Biological Assessment Shorebird Technical Committee. Division of Migratory Bird Management, US Fish and Wildlife Service, Arlington [ Links ]
Atkinson P, Baker AJ, Bevan RM, Clark NA, Cole KB, González PM, Newton J, Niles LJ and Robinson RA (2005) Unravelling the migratory strategies of a long-distance migrant using stable isotopes: Red Knot Calidris canutus movements in the Americas. Ibis 147:738-749 [ Links ]
Baker AJ, González PM, Minton CDT, Carter DB, Niles LJ, do Nascimento I and Piersma T (2001) Hemispheric problems in the conservation of Red Knots (Calidris canutus rufa). Pp. 21-28 in: Proceedings of the VI Neotropical Ornithological Congress, International Shorebird Symposium, Monterrey, México. Western Hemisphere Shorebird Reserve Network, Manomet [ Links ]
Baker AJ, González PM, Piersma T, Niles LJ, do Nascimento ILS, Atkinson PW, Clark NA, Minton CDT, Peck MK and Aarts G (2004) Rapid population decline in Red Knots: fitness consequences of decreased refuelling rates and late arrival in Delaware Bay. Proceedings of the Royal Society of London B 271:875-882 [ Links ]
Baker AJ, González PM, Serrano I, Júnior WRT, Efe MA, Rice S, D'Amico VL, Rocha MC and Echave ME (2005) Assessment of the wintering area of Red Knots in Maranhão, northern Brazil, in February 2005. Wader Study Group Bulletin 107:10-18 [ Links ]
Baker AJ, Piersma T and Greenslade A (1999) Molecular versus phenotypic sexing in Red Knots. Condor 101:887-893 [ Links ]
Boyd H (1992) Arctic summer conditions and British Knot numbers: an exploratory analysis. Wader Study Group Bulletin 64(Supp.):144-152 [ Links ]
Castro G and Myers JP (1993) Shorebird predation on eggs of horseshoe crabs during spring stopover on Delaware Bay. Auk 110:927-930 [ Links ]
Choquet R, Reboulet AM, Pradel R and Lebreton JM (2001) U-care (Utilities-CApture-REcapture) user's guide. CEFE, CNRS, Montpellier [ Links ]
Drent R, Both C, Green M, Madsen J and Piersma T (2003) Pay-offs and penalties of competing migratory schedules. Oikos 103:274-292 [ Links ]
Evans PR, Goss-custard JD and Hale WG (1984) Coastal waders and wildfowl in winter. Cambridge University Press, Cambridge [ Links ]
Evans PR and Pienkowski MW (1984) Population dynamics of shorebirds. Pp. 83-123 in: Burger J and Olla BL (eds) Behavior of Marine Animals 5. Plenum Press, New York [ Links ]
González PM, Carbajal M, Morrison RIG and Baker AJ (2004) Tendencias poblacionales del Playero Rojizo (Calidris canutus rufa) en el sur de Sudamérica. Ornitología Neotropical 15(Suppl.):357-365 [ Links ]
González PM, Piersma T and Verkuil Y (1996) Food, feeding and refuelling of Red Knots during northward migration at San Antonio Oeste, Río Negro, Argentina. Journal of Field Ornithology 67:575-591 [ Links ]
Goss-custard JD, Clarke RT, Briggs KB, Ens BJ, Exo K-M, Smit C, Beintema AJ, Caldow RWG, Catt DC, Clark NA, Le Vdit Durell SEA, Harris MP, Hulscher JB, Meininger PL, Picozzi N, Prys-Jones R, Safriel UN and West AD (1995) Population consequences of winter habitat loss in a migratory shorebird. 1. Estimating model parameters. Journal of Applied Ecology 32:320-336 [ Links ]
Gudmundsson GA, Lindstrom A and Alerstam T (1991) Optimal fat loads and long-distance flights by migrating Knots Calidris canutus, Sanderlings C. alba and Turnstones Arenaria interpres. Ibis 133:140-152 [ Links ]
Harrington BA (2001) Red Knots (Calidris canutus). Pp. 1-32 in: Poole A and Gill F (eds) The birds of North America. Academy of Natural Sciences and American Ornithologists' Union, Philadelphia and Washington DC [ Links ]
Madsen J (2001) Spring migration strategies in pink-footed goose Anser brachyrhynchus and consequences for spring fattening and fecundity. Ardea 89:43-55 [ Links ]
Marra PP, Hobson KA and Holmes RT (1998) Linking winter and summer events in a migratory bird by using stable-carbon isotopes. Science 282:1884-1886 [ Links ]
Morrison RIG (1975) Migration and morphometrics of European Knot and Turnstone on Ellesmere Island, Canada. Bird-Banding 46:290-301 [ Links ]
Morrison RIG (2006) Body transformations, condition, and survival in Red Knots Calidris canutus travelling to breed at Alert, Ellesmere Island, Canada. Ardea 94:607-618 [ Links ]
Morrison RIG, Davidson NC and Piersma T (2005) Transformations at high latitudes: why do Red Knots bring body stores to the breeding grounds? Condor 107:449-457 [ Links ]
Morrison RIG and Harrington BA (1992) The migration system of the Red Knots Calidris canutus rufa in the New World. Wader Study Group Bulletin 64(Suppl.):71-84 [ Links ]
Morrison RIG, Ross RK and Niles LJ (2004) Declines in wintering populations of Red Knots in southern South America. Condor 106:60-70 [ Links ]
Newton I (2004) Population limitation in migrants. Ibis 146:197-226 [ Links ]
Pienkowski MW and Evans PR (1985) The role of migration in the population dynamics of birds. Pp. 331-352 in: Sibly RM and Smith RH (eds) Behavioural ecology. Ecological consequences of adaptive behaviour. Blackwell Scientific Publications, Oxford [ Links ]
Piersma T (1987) Production by intertidal benthic animals and the limits to their predation by shorebirds - a heuristic model. Marine Ecology Progress Series 38:187-196 [ Links ]
Piersma T and Davidson NC (1992) The migrations and annual cycles of five subspecies of Knots in perspective. Wader Study Group Bulletin 64(Suppl.):187-197 [ Links ]
Piersma T, Rogers DI, González PM, Zwarts L, Niles LJ, Nascimento I, Minton CDT and Baker AJ (2005) Fuel storage rates before northward flights in Red Knots worldwide. Pp. 262-273 in: Greenberg R and Marra PP (eds) Birds of two worlds: ecology and evolution of migration. John Hopkins University Press, Baltimore [ Links ]
Sitters H, González PM, Piersma T, Baker AJ and Price D (2001) Day and night feeding habitat of Red Knots in Patagonia: profitability versus safety? Journal of Field Ornithology 72:86-95 [ Links ]
Stroud DA, Baker A, Blanco DE, Davidson NC, Delany S, Ganter B, Gill R, González P, Haanstra L, Morrison RIG, Piersma T, Scott DA, Thorup O, West R, Wilson J and Zöckler C (2006) The conservation and population status of the world's waders at the turn of the millennium. Pp. 643-648 in: Boere GC, Galbraith CA and Stroud DA (eds) Waterbirds around the world. A global overview of the conservation, management and research of the world's waterbird flyways. The Stationery Office, Edinburgh [ Links ]
Tomkovich PS (2001) A new subspecies of Red Knots Calidris canutus from the New Siberian islands. Bulletin of the British Ornithologists' Club 121:257-263 [ Links ]
Tsipoura N and Burger J (1999) Shorebird diet during spring migration stopover on Delaware Bay. Condor 101:635-644 [ Links ]
Walls EA, Berkson J and Smith SA (2002) The horseshoe crab, Limulus polyphemus: 200 million years of existence, 100 years of study. Reviews in Fisheries Science 10:39-73 [ Links ]
White GC and Burnham KP (1999) Program MARK: survival estimation from populations of marked animals. Bird Study 46(Suppl.):120-139 [ Links ]

