










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkEl hornero
versão impressa ISSN 0073-3407versão On-line ISSN 1850-4884
Hornero v.22 n.2 Buenos Aires dez. 2007
Respuesta de las aves rapaces al uso de la tierra: un enfoque regional
Julieta Filloy 1,2 y M. Isabel Bellocq 1
1 ECOMA - Laboratorio de Ecología de Comunidades y Macroecología, Departamento de Ecología, Genética y Evolución, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Piso 4, Pab. 2, Ciudad Universitaria, C1428EHA Buenos Aires, Argentina.
2 jfilloy@ege.fcen.uba.ar
Resumen
La Región Pampeana ha sufrido cambios en toda su extensión debido principalmente a la agricultura, la ganadería y la urbanización. Actualmente, existe un gradiente regional agrícola-ganadero en el sentido N-S. En este trabajo se examina la respuesta de las aves rapaces a la intensidad de uso agrícola, relevando aves y uso de la tierra en 17 transectas que cubren el gradiente. Para explorar la relación entre la presencia y la abundancia de cada especie con la intensidad de uso agrícola se usaron regresiones logísticas y simples y autocorrelogramas. La Prueba de Mantel y el Análisis de Redundancia permitieron explorar la respuesta de la abundancia de las especies a la composición de los paisajes. En total se identificaron 11 especies de rapaces. La abundancia total respondió a la estructura espacial del uso de la tierra en la región, siendo el ensamble más abundante en paisajes ganaderos. Sin embargo, la mayor parte de las especies estuvo presente tanto en paisajes ganaderos como agrícolas. Desde una perspectiva regional, el ensamble fue afectado negativamente en mayor medida por la agricultura que por la ganadería, aunque la mayor parte de las especies fue tolerante al uso de la tierra. Sin embargo, dentro del paisaje agrícola o del paisaje ganadero las especies estudiadas manifestaron diferencias en cuanto al tipo de ambiente que explotan.
Palabras clave: Abundancia;, Agricultura; Argentina; Aves rapaces; Ganadería; Región Pampeana; Uso de la tierra.
Abstract
Raptorial bird responses to land use: a regional approach
The Pampean Region of Argentina has been gradually and completely transformed by agriculture (croplands and pastoral farming) and urbanization. Currently, there is a N-S gradient from a pure cropland through mix-farming to a pure pastoral landscape. We examine the response of raptorial birds to agricultural intensity by surveying birds and land use along 17 transects covering the gradient. To examine the relationship between species presence or abundance and agricultural intensity we used logistic and simple regressions and correlograms. We used Mantel Test and Redundancy Analysis to explore relationships between bird abundance and landscape elements. We recorded a total of 11 raptor species. Abundance of the raptor assemblage responded to the spatial structure of land use in the region, being more abundant in pastoral farming landscapes. Nonetheless, most species were found in both croplands and pastoral landscapes. From a regional perspective, the use of the land for crops seems to be more detrimental to raptors than pastoral farming, even though most raptor species showed to be tolerant to land use. However, within each particular landscape (cropland or pastoral land), the studied raptor species showed differences in the type of habitat they exploit.
Key words: Abundance; Agriculture; Argentina; Land use; Pampean Region; Pastoral farming; Raptorial birds.
Recibido 6 noviembre 2006, aceptado 30 noviembre 2007
Una de las ideas que impactaron en el pensamiento de la ecología tradicional durante la década de 1980 fue el entendimiento de que los patrones y procesos ecológicos pueden cambiar con la escala geográfica. Hasta entonces, la perspectiva estaba básicamente limitada a comunidades locales y los factores que gobiernan su estructura, como las perturbaciones, la heterogeneidad del hábitat o las relaciones interespecíficas. A partir entonces, se comprende que aunque algunos atributos como la riqueza específica o la abundancia son influenciados por muchos factores actuando a múltiples escalas, los atributos a escala local tienden a ser determinados por factores que actúan a escala local y los atributos a gran escala tienden a ser determinados por factores que operan a esa misma escala (Wiens 1989). Se intensificó entonces la búsqueda de las causas que determinan los patrones ecológicos espaciales a gran escala (ver revisiones de Rohde 1992 y Willig et al. 2003). Se destaca el estudio pionero que realizaron Rabinovich y Rapoport (1975) describiendo los cambios espaciales de riqueza específica de Passeriformes en Argentina y su asociación con variables ambientales, cuando los asuntos de escala todavía no eran un tema central en ecología. Más recientemente, con un marco teórico y metodológico mejor desarrollado, las aves siguieron proporcionando un buen modelo para el estudio de los factores que determinan patrones regionales tanto de riqueza específica (Cueto y Lopez de Casenave 1999, Bellocq y Gómez Insausti 2005) como de abundancia (Filloy y Bellocq 2006, 2007). Las aves rapaces, en particular, han sido utilizadas como modelos para responder preguntas centrales en ecología y conservación a gran escala en la última década: ¿qué factores determinan los patrones espaciales de riqueza específica (Diniz-Filho et al. 2004, Meynard et al. 2004, Bellocq y Gómez Insausti 2005, Sergio et al. 2005)?, ¿cómo responde la fauna nativa a los cambios en el uso de la tierra (Sánchez-Zapata y Calvo 1999, Herremans y Herremans-Tonnoeyr 2000, Sánchez-Zapata et al. 2003, Zurita y Bellocq 2007)?, ¿son las especies indicadoras útiles para el manejo de la biodiversidad (Rodríguez-Estrella et al. 1998)? Este trabajo intenta contribuir a responder alguna de estas preguntas.
La pérdida de hábitat debida a los cambios en el uso de la tierra afecta a todos los biomas. Las áreas de ambiente natural se están reduciendo a tasas muy altas y puede predecirse que solo una mínima proporción de tierra será mantenida dentro de reservas naturales. Se ha estimado que tan solo el 5% de la superficie terrestre está libre de modificaciones por acción humana (Meffe et al. 1997). Consecuentemente, el manejo de la matriz de uso productivo es fundamental para perpetuar poblaciones en el largo plazo, especialmente de especies que requieren grandes extensiones de hábitat como las aves rapaces. Se realizaron muchos estudios de poblaciones y comunidades de aves rapaces en paisajes con matrices de uso humano en América del Norte (Bird et al. 1996). Por ejemplo, la abundancia general de rapaces ha sido utilizada muy frecuentemente para evaluar el impacto de las transformaciones humanas de los ambientes (Rodríguez-Estrella et al. 1998, Sánchez-Zapata et al. 2003). Sin embargo, aún existen vacíos de información sobre las respuestas de las rapaces a cambios generados por el uso de la tierra en muchos biomas. En Argentina, solo en unos pocos estudios se ha relacionado de alguna manera a las aves rapaces con el uso de la tierra (e.g., Donázar et al. 1993, Travaini et al. 1995, Leveau y Leveau 2002, Zurita y Bellocq 2007).
La Región Pampeana en Argentina presenta un buen escenario para el estudio de los efectos del uso de la tierra sobre las comunidades. Originalmente dominada por estepas graminosas (Cabrera 1971), es una de las regiones de mayor productividad agrícola del planeta y se encuentra casi completamente transformada debido al uso intensivo agrícola y ganadero y a los asentamientos humanos (León et al. 1984). Sin embargo, dentro de la región la productividad de la tierra es heterogénea y el uso se encuentra zonificado (SAGPyA 2006). La productividad tiende a disminuir de norte a sur (i.e., desde la Pampa Ondulada hacia la Pampa Deprimida) debido a las características geomorfológicas de la región, determinadas principalmente por la depresión del río Salado (INTA 1990). Así, actualmente existe un gradiente norte-sur desde un paisaje predominantemente agrícola a uno predominantemente ganadero. Esta condición geográfica permite explorar asociaciones entre la abundancia de las aves rapaces y el uso de la tierra a lo largo de un gradiente de intensidad de disturbio dado por el uso agrícola o ganadero. El uso agrícola produce un disturbio más intenso que el ganadero, ya que el manejo agrícola incluye intervenciones con maquinarias y agroquímicos que implican un mayor impacto sobre el ambiente.
El objetivo de este trabajo es describir y analizar las respuestas de las aves rapaces a cambios espaciales en el uso de la tierra. Específicamente, se examina el patrón de presencia-ausencia (por especie) y de abundancia (total y por especie) de aves rapaces a lo largo del gradiente de uso de la tierra de la Región Pampeana, desde un paisaje agrícola hacia uno ganadero. Además, se explora la respuesta de la abundancia de cada una de las especies a la presencia y abundancia de elementos (i.e., usos específicos de la tierra) que caracterizan a esos paisajes.
Métodos
Área de estudio
El estudio se llevó a cabo en la parte central de la Región Pampeana (Cabrera 1971). Esta región presenta un clima templado con una temperatura promedio anual que oscila entre 13-17 °C. Las precipitaciones son relativamente constantes a lo largo del año, variando, en promedio, entre 1100 mm en el norte y 600 mm en el sur. La estepa graminosa que caracteriza a la región se encuentra dominada por pastos de los géneros Stipa, Piptochaetium, Aristida, Melica, Briza, Bromus, Eragrostis y Poa (Cabrera 1971).
El estudio abarcó aproximadamente 350 km en dirección norte-sur (desde los 33°54'S, 60°01'O hasta los 36°43'S, 58°59'O; Fig. 1). La extensión resultó suficiente como para cubrir el gradiente de uso de la tierra que presenta la región, desde un 87% de tierra agrícola al norte, cambiando gradual y complementariamente hasta alcanzar un 91% de tierra ganadera al sur (Filloy y Bellocq 2007). El área estudiada está ubicada completamente dentro del Distrito Oriental de la región (Cabrera 1971). Allí, el clima es relativamente constante, con temperaturas mínimas promedio de 7.5-9.5 °C, temperaturas máximas promedio de 21.5-23.5 °C y precipitaciones entre 800-1000 mm (De Fina 1992).
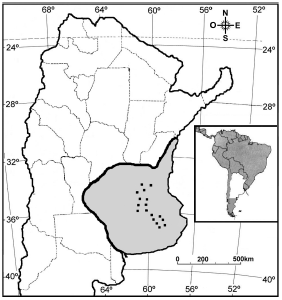
Figura 1. Ubicación aproximada de las 17 transectas en donde fueron relevadas las aves rapaces dentro de la Región Pampeana (área gris).
Muestreo de rapaces
Se realizaron relevamientos de aves rapaces diurnas por carretera a lo largo del gradiente ambiental durante noviembre de 1999, en la época de actividad reproductiva de la mayoría de las rapaces; los resultados hallados se restringen a este período. La realización de los relevamientos en primavera permite incorporar especies como Buteo swainsoni, que migran y se reproducen en el Hemisferio Norte durante el invierno del Hemisferio Sur. Se utilizó el método de puntos con radio ilimitado (Bibby et al. 1998). Se seleccionaron 17 transectas de 25 km (Fig. 1) dispuestas sobre caminos secundarios sin asfalto, con escaso tránsito (Ralph et al. 1995). Las mismas fueron recorridas con vehículo, realizando una parada de observación de 5 min cada 1 km (estableciendo un total de 26 puntos de observación por transecta). En cada punto se registró la cantidad de individuos de todas las especies de aves rapaces identificadas por vista o canto. Todos los relevamientos fueron realizados por los mismos observadores, entre las 6:00-11:00 h. En los análisis se consideró la presencia de cada especie y la abundancia total y por especie (expresada como la cantidad de individuos registrados por transecta).
Uso de la tierra: clasificación y cuantificación
Se examinaron las respuestas de las rapaces tanto al gradiente de uso de la tierra desde un paisaje con uso predominantemente agrícola a uno predominantemente ganadero como a la abundancia de los distintos elementos (i.e., usos específicos de la tierra) que componen los paisajes. A lo largo del recorrido de las 17 transectas se registró la distancia cubierta por cada uso específico de la tierra (Tabla 1) lindantes al camino (Dd y Di para derecha e izquierda del camino, respectivamente). Se calculó el porcentaje de la transecta ocupado por cada uso específico. Para ello, la suma de las distancias a ambos lados del camino ocupadas por cada uso se relativizó al doble de la longitud de la transecta (50 km) según la fórmula:
P = 100 (Dd + Di) / 50.
Luego, para cuantificar el gradiente agrícola-ganadero, se calculó el porcentaje de la tierra destinado a la agricultura por transecta. Para ello, dentro de cada transecta se sumaron los porcentajes ocupados por los usos específicos correspondientes a actividades agrícolas.
Tabla 1. Clasificación de los principales usos de la tierra encontrados en los paisajes agrícola y ganadero a lo largo de los relevamientos en la Región Pampeana.

Análisis de datos
Para examinar la respuesta general de las rapaces al gradiente espacial de usos de la tierra, se realizaron regresiones simples (lineales y no lineales) entre la abundancia total de rapaces y el porcentaje de tierra destinada a la agricultura. La ganadería y la agricultura son los principales usos de la región y, por lo tanto, son complementarios en cuanto al área que se destina a cada uno, por lo que una respuesta positiva a uno de ellos implica una respuesta negativa al otro y viceversa (más detalles en Filloy y Bellocq 2007).
Se controló la autocorrelación espacial de los datos generando un correlograma en base al Índice de Moran con los datos de abundancia por transecta y otro correlograma con los residuos de la abundancia luego de realizar la regresión y eliminar la variación explicada por la agricultura (Sánchez-Zapata et al. 2003, Diniz Filho et al. 2004). El correlograma permite analizar el grado de asociación de una variable con sí misma en función de la distancia entre los sitios donde los datos fueron tomados. A través de este análisis de los datos originales y de los residuos es posible distinguir entre posibles factores causales de esa estructura espacial y factores que correlacionan de manera espuria con la variable de interés (Diniz-Filho et al. 2004).
Para cada especie por separado se analizó el patrón de presencia-ausencia a lo largo del gradiente agrícola-ganadero. Se realizaron regresiones logísticas entre la presencia-ausencia (variable dependiente) y el porcentaje de tierra destinada a uso agrícola (variable independiente) por transecta.
Se examinó el grado de asociación entre los cambios ambientales (porcentaje de los usos de la tierra) y los cambios en la composición y abundancia de las especies por transecta a lo largo del gradiente. Se construyeron matrices de disimilitud entre transectas utilizando distancias euclidianas (van Tongeren 1995). La correlación entre ambas matrices se evaluó mediante la Prueba de Mantel basada en rangos (Legendre y Legendre 1998). Para examinar la respuesta de cada especie a los distintos usos que componen el paisaje agrícola y el ganadero se realizó un Análisis de Redundancia, suponiendo una respuesta lineal por parte de las especies a los cambios en las variables ambientales (ter Braak 1995). El Análisis de Redundancia ordena a las especies en función de las variables explicativas, maximizando las diferencias en sus respuestas. El modelo final consta de ejes ortogonales que, del primero al último, explican porcentajes decrecientes de la varianza total de la abundancia de las especies. Estos ejes son combinaciones lineales de las variables explicativas consideradas. Las especies y las variables ambientales son representadas por flechas; dos flechas que apuntan hacia el mismo lado indican una correlación positiva entre la especie y las variables, las que apuntan hacia lados opuestos indican una correlación negativa y si forman un ángulo recto indican una ausencia de correlación entre ellas (ter Braak 1995). Así, se incluyó en el análisis una matriz de abundancia por especie por transecta y otra matriz con los porcentajes de los distintos usos de la tierra por transecta. Se controló la dependencia espacial de los datos incorporados en ambos análisis utilizando la Prueba de Mantel, usando una matriz de distancia basada en la abundancia de las especies incluidas en la primera Prueba de Mantel (ver arriba) y en el Análisis de Redundancia y otra matriz de distancia basada en las coordenadas geográficas de las transectas (Legendre y Legendre 1998).
Resultados
A lo largo del gradiente agrícola-ganadero estudiado y luego de relevar las 17 transectas, se registró un total de 1666 individuos pertenecientes a 11 especies de aves rapaces (Tabla 2). Sin embargo, de estos registros, 1301 individuos correspondieron a Milvago chimango (aproximadamente el 78%).
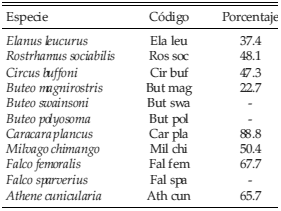
Tabla 2. Especies de aves rapaces identificadas durante los relevamientos en la Región Pampeana. Para cada una de las especies incluidas en el análisis de datos se indica el porcentaje de varianza de su abundancia explicado por el uso específico de la tierra de acuerdo al modelo del Análisis de Redundancia.
El ensamble de aves rapaces pareció responder al incremento en la intensidad agrícola a través de cambios en la abundancia (Fig. 2). La abundancia total de rapaces disminuyó de manera lineal con el incremento en el porcentaje de tierra destinado a la agricultura (F1,15 = 10.9880, P = 0.0047). Sin embargo, si no se considera a la especie dominante (Milvago chimango), la abundancia disminuyó exponencialmente (F1,15 = 25.7511, P < 0.0001). Los correlogramas evidenciaron, sobre la base de los datos de abundancia de las especies, la existencia de autocorrelación espacial positiva entre transectas cercanas y una autocorrelación espacial negativa entre transectas alejadas (Fig. 3). La intensidad de uso agrícola resultó un factor apropiado para explicar de manera parcial el patrón de abundancia observado, ya que luego de eliminar el efecto de la agricultura la estructura espacial de la abundancia de las rapaces no fue tan marcada. Los residuos no conservaron una estructura espacial ni a corta ni a larga distancia, con la excepción de las transectas que distan entre sí aproximadamente unos 200 km (Fig. 3).

Figura 2. Resultados del Análisis de Regresión Simple entre el porcentaje de tierra destinado a la agricultura y la abundancia total de aves rapaces (arriba) y la abundancia de rapaces excluyendo a Milvago chimango (abajo) en la Región Pampeana.
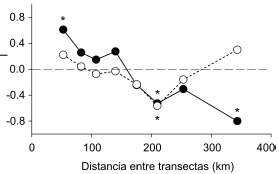
Figura 3. Correlograma en base al Índice I de Moran de la abundancia de aves rapaces en la Región Pampeana en función de la distancia entre transectas (círculos negros, línea llena) y de los residuos de la abundancia luego de eliminar la variación explicada por la agricultura (círculos blancos, línea rayada). *: P < 0.05.
Las especies estudiadas no mostraron un patrón de presencia-ausencia a lo largo del gradiente agrícola-ganadero. Milvago chimango fue registrada en todas las transectas (Fig. 4a) y Caracara plancus en casi todas (Fig. 4b). El resto de las especies se observaron tanto en transectas con predominio de actividad agrícola como en otras de uso ganadero (e.g., Rostrhamus sociabilis; Fig. 4c), evidenciando una tendencia a aparecer en ambos tipos de paisaje (P > 0.05 en todos los casos). Contrariamente a lo esperado por ser una especie muy común en toda la región, Falco sparverius sólo estuvo representado en algunas transectas con baja intensidad de uso agrícola (P < 0.05) (Fig. 4d). Buteo swainsoni (n = 4) y Buteo polyosoma (n = 1) fueron observadas en una única transecta (con un porcentaje de tierra de uso ganadero mayor al 90%), por lo que no fue posible evaluar el patrón de presencia-ausencia. Estas últimas tres especies fueron excluidas de los análisis posteriores.

Figura 4. Patrones de presencia-ausencia de especies de aves rapaces en función del porcentaje de tierra destinado a la agricultura en la Región Pampeana: (a) Milvago chimango, (b) Caracara plancus, (c) Rostrhamus sociabilis y (d) Falco sparverius.
Al considerar las respuestas teniendo en cuenta la abundancia por transecta y por especie, y las coordenadas geográficas de las transectas, la Prueba de Mantel reveló una ausencia de autocorrelación espacial entre ellas (rSpearman = -0.299, P = 0.998). Es decir, las transectas cercanas no resultaron más parecidas entre sí (en cuanto a la abundancia de las rapaces presentes) que lo esperado por azar, y las transectas alejadas no resultaron más diferentes que lo esperado por azar. La Prueba de Mantel reveló, en cambio, una correlación positiva entre la matriz de distancia basada en las abundancias y la matriz ambiental basada en el porcentaje de usos de la tierra presentes en cada transecta (rSpearman = 0.812, P = 0.001). Esto indica que cuanto más parecidas son dos transectas en cuanto a sus componentes (usos de la tierra y porcentaje ocupado por cada uno), también son más parecidas en cuanto a la composición y abundancia de las especies de aves rapaces.
Las especies respondieron a diferentes componentes del paisaje agrícola y del ganadero. Algunas especies se encontraron asociadas positivamente a variables que forman parte del paisaje agrícola: Elanus leucurus y Athene cunicularia estuvieron positivamente asociados a campos cultivados con avena, y Buteo magnirostris a campos con trigo y rastrojo (Fig. 5). Otras estuvieron asociadas positivamente a variables del paisaje ganadero: Caracara plancus y Rostrhamus sociabilis a los campos con presencia de ganado, Milvago chimango y Circus buffoni a los campos con pasturas seminaturales altas y sin ganado (Fig. 5). El modelo del Análisis de Redundancia mostró un alto grado de asociación entre los usos de la tierra y la abundancia de las especies (r = 0.82 y una varianza explicada del 45.3% para el primer eje) y explicó entre un moderado y un alto porcentaje la variabilidad en la abundancia de las especies a lo largo del gradiente (Tabla 2).
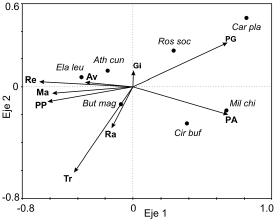
Figura 5. Resultado del Análisis de Redundancia basado en las abundancias de las especies de aves rapaces y los usos de la tierra de los paisajes agrícola y ganadero en la Región Pampeana. Los códigos de los usos de la tierra y de las especies son los mismos que en las tablas 1 y 2.
Discusión
La abundancia total de aves rapaces parece responder a la estructura espacial del uso de la tierra en la Región Pampeana, debido a que fueron más abundantes en paisajes ganaderos que en paisajes agrícolas. Sin embargo, la respuesta general del ensamble estuvo influenciada por la de la especie más abundante, Milvago chimango. La respuesta de esta especie suavizó la tendencia del resto del ensamble; la abundancia de rapaces disminuyó más marcadamente con el incremento en la intensidad de uso agrícola cuando se excluyó del análisis a Milvago chimango. Esto indica que el ensamble en general es sensible al uso de la tierra y que el uso agrícola tiene un impacto negativo mayor que el uso ganadero. De acuerdo a los correlogramas, los cambios espaciales en el uso de la tierra explicaron satisfactoriamente los cambios espaciales en la abundancia de rapaces. Sin embargo, se encontró que el uso de la tierra no pudo explicar las diferencias en la abundancia a distancias de aproximadamente 200 km. Esto indica la influencia de un factor no incorporado en los análisis, que actúa a una escala intermedia (Diniz-Filho et al. 2003). Dentro del área de estudio las zonas más altas y más bajas topográficamente se encuentran a una distancia aproximada de 200 km (INTA 1990). De esta manera, el drenaje diferencial y sus efectos sobre los ambientes (e.g., humedad del suelo y efecto sobre la microfauna) podrían explicar las diferencias en la abundancia de rapaces a esas distancias. La abundancia de rapaces se encuentra determinada por la oferta de recursos del ambiente (Preston y Beane 1996, Herremans y Herremans-Tonnoeyr 2000). Así, es posible que la respuesta encontrada se deba a que la ganadería favorece una mejor oferta de recursos (e.g., ganado en descomposición) o a que este uso de la tierra preserve recursos más similares a los ofrecidos por el pastizal original que el uso agrícola. Las especies estudiadas requieren y explotan diferentes recursos para alimentarse. El análisis de las respuestas individuales (ver más adelante) permitiría un entendimiento más profundo acerca de cómo opera el uso de la tierra sobre la abundancia de las rapaces.
Desde una perspectiva regional, varias especies de aves rapaces mostraron ser tolerantes a la transformación ambiental que sufrió la Región Pampeana desde la introducción del ganado en el siglo XVII, registrándose su presencia tanto en paisajes agrícolas como ganaderos. Aparentemente, ambos paisajes generan recursos atractivos para las rapaces. A una escala menor, Preston y Beane (1996) encontraron que la mayor parte de las 12 especies de rapaces que estudiaron resultaron tolerantes a la actividad humana. Los resultados para Falco sparverius, Buteo polyosoma y Buteo swainsoni sugieren que estas especies no son tolerantes a la actividad agrícola, ya que solo fueron registradas en zonas preponderantemente ganaderas. Sin embargo, es ampliamente reconocido que Falco sparverius es muy común y habita en toda la región (Narosky y Di Giacomo 1993) y que, aunque este estudio no lo reflejó, es muy frecuente en campos cultivados (e.g., Hernández Vázquez et al. 2000). Además, Preston y Beane (1996) encontraron que la presencia de Buteo swainsoni era independiente de la intensidad de la actividad humana. Una investigación particular para estas especies podría clarificar estas aparentes contradicciones, sobre todo porque la interpretación de los resultados se basa en un número bajo de registros.
A una escala menor, las rapaces respondieron de manera diferente a los usos de la tierra (i.e., al cultivo o al estado de la pastura que se presenta en cada campo). Dos paisajes con alto porcentaje de tierra agrícola podrían diferir en cuanto a la abundancia e incluso presencia de las especies de rapaces porque, por ejemplo, se encuentran ocupados por distintos cultivos. Esto explicaría por qué, al analizar los patrones de presencia-ausencia a lo largo del gradiente, la mayor parte de las especies estuvo presente en algunas transectas agrícolas o ganaderas y no en otras. Existen evidencias de explotación diferencial de los recursos disponibles a escala local en algunas condiciones particulares del paisaje agrícola (Smallwood et al. 1996). Caracara plancus respondió positivamente a un incremento en el porcentaje de campos bajo pastoreo al momento de los relevamientos, posiblemente por la mayor disponibilidad de alimento (fundamentalmente carroña). Esta especie responde a la actividad humana (Rodríguez-Estrella 1996, 2007) y específicamente a cambios en la disponibilidad de carroña (Rodríguez-Estrella 1996). En coincidencia con los resultados de este trabajo, Leveau y Leveau (2004) encontraron que Milvago chimango resultó más abundante en campos con pasturas que en campos cultivados. También hay evidencia de que las actividades agrícolas impactan negativamente en algunas especies de rapaces. Por ejemplo, se ha registrado la destrucción de nidos de Athene cunicularia por la acción de maquinarias en campos agrícolas (Bellocq 1993, 1997) y la mortalidad masiva de individuos de Buteo swainsoni debido a la aplicación de insecticidas químicos (Hartley et al. 1996, Goldstein et al. 1999).
En general, las respuestas de las rapaces a la transformación de los hábitats originales por parte del hombre son variadas. Los resultados muestran que varias especies de la Región Pampeana, al estar presentes a lo largo de casi todo el gradiente, son tolerantes tanto al uso agrícola como ganadero. Sin embargo, a pesar de esa tolerancia, las rapaces resultaron sensibles a la estructura espacial del uso de la tierra, mostrando cambios en la abundancia, ya sea total o por especie. Las rapaces de regiones de clima templado suelen no responder a alteraciones moderadas del ambiente porque se adecuan rápidamente a estos nuevos sistemas (Rodríguez-Estrella et al. 1998). Por el contrario, las especies de climas tropicales son particularmente vulnerables a los cambios en el ambiente (Thiollay 1996, Zurita y Bellocq 2007), mientras que las de clima árido pueden verse favorecidas por el uso de la tierra (Rodríguez-Estrella 2007). Este trabajo sugiere que en la Región Pampeana las actividades agrícolas y ganaderas no limitan la presencia de varias especies de aves rapaces. Sin embargo, la distribución espacial de su abundancia puede encontrarse influenciada por la forma en que el uso de la tierra se estructura dentro de la región.
Agradecimientos. Los comentarios y sugerencias de G. Zurita y tres revisores anónimos contribuyeron a mejorar una versión preliminar del manuscrito. El trabajo fue financiado por el Consejo Nacional de Investigaciones Científicas y Técnicas de Argentina, la Universidad de Buenos Aires y el Lincoln Park Zoo.
Bibliografía Citada
Bellocq MI (1993) Reproducción, crecimiento y mortalidad de Athene cunicularia (lechucita de las vizcacheras) en agrosistemas de Argentina. Hornero 13:272-276 [ Links ]
Bellocq MI (1997) Ecology of the Burrowing owl in agrosystems of central Argentina. Journal of Raptor Research Reports 9:35-41 [ Links ]
Bellocq MI y Gómez-Insausti JR (2005) Raptorial birds and environmental gradients in the southern Neotropics: a test of species-richness hypotheses. Austral Ecology 30:900-906 [ Links ]
Bibby C, Jones M y Mardsen S (1998) Expedition field techniques. Bird Surveys. Royal Geographical Society, Londres [ Links ]
Bird DM, Varland D y Negro JJ (1996) Raptors in human landscapes. Academic Press, Londres [ Links ]
ter Braak CJF (1995) Ordination. Pp. 91-173 en: Jongman RHG, ter Braak CJF y van Tongeren OFR (eds) Data analysis in community and landscape ecology. Cambridge University Press, Cambridge [ Links ]
Cabrera A (1971) Fitogeografía de la Argentina. Boletín de la Sociedad Argentina de Botánica 14:1-43 [ Links ]
Cueto VR y Lopez de Casenave J (1999) Determinants of bird species richness: role of climate and vegetation structure at a regional scale. Journal of Biogeography 26:487-492 [ Links ]
De Fina AL (1992) Aptitud agroclimática de la República Argentina. Academia Nacional de Agronomía y Veterinaria, Buenos Aires [ Links ]
Diniz-Filho JAF, Bini LM y Hawkins BA (2003) Spatial autocorrelation and red herrings in geographical ecology. Global Ecology and Biogeography 12:53-64 [ Links ]
Diniz-Filho JAF, Rangel TFLVB y Hawkins BA (2004) A test of multiple hypothesis of the diversity gradient of South American owls. Oecologia 140:633-638 [ Links ]
Donázar JA, Ceballos O, Travaini A e Hiraldo F (1993) Roadside raptor surveys in the Argentinean Patagonia. Journal of Raptor Research 27:106-110 [ Links ]
Filloy J y Bellocq MI (2006) Spatial variation in the abundance of Sporophila seedeaters in the southern Neotropics: contrasting the effects of agricultural development and geographical position. Biodiversity and Conservation 15:3329-3340 [ Links ]
Filloy J y Bellocq MI (2007) Patterns of bird abundance along the agricultural gradient of the Pampean region. Agriculture, Ecosystems and Environment 120:291-298 [ Links ]
Goldstein MI, Lacher TE, Woodbridge B, Bechard MJ, Canavelli SB, Zaccagnini ME, Cobb GP, Scollon EJ, Tribolet R y Hooper MJ (1999) Monocrotophos-induced mass mortality of Swainson’s hawks in Argentina, 1995-95. Ecotoxicology 8:201-214 [ Links ]
Hartley RR, Hustler K y Mundy PJ (1996) The impact of man on raptors in Zimbabwe. Pp. 337-354 en: Bird DM, Varland D y Negro JJ (eds) Raptors in human landscapes. Academic Press, Londres [ Links ]
Hernández Vázquez S, Durand Martínez BC, Esparza Salas R y Valadez González C (2000) Distribución temporal de aves rapaces diurnas en la Reserva “Playón de Mismaloya”. Revista de Biología Tropical 48:1015-1018 [ Links ]
Herremans M y Herremans-Tonnoeyr D (2000) Land use and the conservation status of raptors in Botswana. Biological Conservation 94:31-41 [ Links ]
INTA (1990) Atlas de suelos de la República Argentina. Instituto Nacional de Tecnología Agropecuaria, Buenos Aires [ Links ]
Legendre P y Legendre L (1998) Numerical ecology. Elsevier, Amsterdam [ Links ]
León RJC, Rusch GM y Oesterheld M (1984) Pastizales pampeanos — impacto agropecuario. Phytocoenologia 12:201-218 [ Links ]
Leveau LM y Leveau CM (2002) Uso de hábitat por aves rapaces en un agroecosistema pampeano. Hornero 17:9-15 [ Links ]
Leveau LM y Leveau CM (2004) Efectos de la urbanización en la distribución y abundancia del chimango Milvago chimango en Mar del Plata, Argentina, durante un ciclo anual. Pp. 12-13 en: Actas del Primer Simposio Argentino sobre Investigación y Conservación de Rapaces, Museo de La Plata, La Plata, Buenos Aires, Argentina, 1 y 2 de octubre de 2004. Museo de La Plata, La Plata [ Links ]
Meffe GK y Carroll CR (1997) Principles of conservation biology. Segunda edición. Sinauer Associates, Sunderland [ Links ]
Meynard C, Samaniego H y Marquet P (2004) Biogeografía de aves rapaces de Chile. Pp. 129-143 en: Muñoz-Pedreros A, Rau J y Yañez J (eds) Aves rapaces de Chile. CEA Ediciones, Valdivia [ Links ]
Narosky T y Di Giacomo AG (1993) Las aves de la Provincia de Buenos Aires: distribución y estatus. Asociación Ornitológica del Plata, Vázquez Mazzini Editores y LOLA, Buenos Aires [ Links ]
Preston CR y Beane RD (1996) Occurrence and distribution of diurnal raptors in relation to human activity and other factors at Rocky Mountain Arsenal, Colorado. Pp. 365-374 en: Bird DM, Varland D y Negro JJ (eds) Raptors in human landscapes. Academic Press, Londres [ Links ]
Rabinovich JE y Rapoport EH (1975) Geographical variation of diversity in Argentine passerine birds. Journal of Biogeography 2:141-157 [ Links ]
Ralph CJ, Geupel GR, Pyle P, Martin TE, DeSante DF y Milá B (1995) Manual de métodos de campo para el monitoreo de aves terrestres. USDA Forest Service General Technical Report PRW-GTR, Albany [ Links ]
Rodríguez-Estrella R (1996) Response of common Black hawks and Crested caracaras to human activities in Mexico. Pp. 355-364 en: Bird DM, Varland D y Negro JJ (eds) Raptors in human landscapes. Academic Press, Londres [ Links ]
Rodríguez-Estrella R (2007) Land use changes affect distributional patterns of desert birds in the Baja California peninsula, Mexico. Diversity and Distributions 13:877-889 [ Links ]
Rodríguez-Estrella R, Donázar JA e Hiraldo F (1998) Raptors as indicators of environmental change in the scrub habitat of Baja California Sur, Mexico. Conservation Biology 12:921-925 [ Links ]
Rohde K (1992) Latitudinal gradients in species diversity: the search for the primary cause. Oikos 65:514-527 [ Links ]
SAGPyA (2006) La integración de la ganadería argentina. Situación de la ganadería pampeana. Secretaría de Agricultura, Ganadería, Pesca y Alimentos, Ministerio de Economía y Producción de la República Argentina, Buenos Aires [ Links ]
Sánchez-Zapata JA y Calvo JF (1999) Raptor distribution in relation to landscape composition in semi-arid Mediterranean habitats. Journal of Applied Ecology 36:254-262 [ Links ]
Sánchez-Zapata JA, Carrete M, Gravilov A, Sklyarenko S, Cevallos O, Donázar JA e Hiraldo F (2003) Land use changes and raptor conservation in steppe habitats of eastern Kazakhstan. Biological Conservation 111:71-77 [ Links ]
Sergio F, Newton I y Marchesi L (2005) Top predators and biodiversity. Nature 436:192 [ Links ]
Smallwood SK, Nakamoto BJ y Geng S (1996) Association analysis of raptors on a farming landscape. Pp. 177-190 en: Bird DM, Varland D y Negro JJ (eds) Raptors in human landscapes. Academic Press, Londres [ Links ]
Thiollay JM (1996) Rain forest raptor communities in Sumatra: the conservation value of traditional agroforest. Pp. 245-261 en: Bird DM, Varland D y Negro JJ (eds) Raptors in human landscapes. Academic Press, Londres [ Links ]
van Tongeren OFR (1995) Cluster analysis. Pp. 174-212 en: Jongman RHG, ter Braak CJF y van Tongeren OFR (eds) Data analysis in community and landscape ecology. Cambridge University Press, Cambridge [ Links ]
Travaini A, Rodríguez A, Ceballos O, Donázar JA e Hiraldo F (1995) Roadside raptor surveys in Central Argentina. Hornero 14:64-66 [ Links ]
Wiens JA (1989) Spatial scaling in ecology. Functional Ecology 3:385-387 [ Links ]
Willig MR, Kaufman DM y Stevens RD (2003) Latitudinal gradients of biodiversity: pattern, process, scale and synthesis. Annual Review of Ecology and Systematics 34:273-309 [ Links ]
Zurita GA y Bellocq MI (2007) Pérdida y fragmentación de la Selva Paranaense: efectos sobre las aves rapaces diurnas. Hornero 22:141-147 [ Links ]

