










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEl hornero
versión impresa ISSN 0073-3407versión On-line ISSN 1850-4884
Hornero v.23 n.1 Buenos Aires ene./ago. 2008
Extinciones locales de aves en fragmentos de bosque en la región de Santa Elena, Andes Centrales, Colombia
Gabriel J. Castaño-Villa 1 y Juan C. Patiño-Zabala 2
1 Grupo de Investigación en Ecosistemas Tropicales, Departamento de Recursos Naturales y Medio Ambiente, Universidad de Caldas. AA 275, Manizales, Colombia. gabriel.castano_v@ucaldas.edu.co
2 Ed. Centrum. Alberto Borges y Francisco de Orellana Piso 9 Of. 6, Guayaquil, Ecuador.
Resumen
Los registros históricos de la avifauna de la región de Santa Elena (Andes centrales colombianos), realizados entre 1879 y 1952, fueron comparados con los de las aves presentes actualmente en tres fragmentos remanentes de bosque de 41, 129 y 136 ha. De las 132 especies presentes originalmente solo se conservan 64 en los tres fragmentos. En los tres fragmentos han desaparecido el 78%, el 66% y el 63% de sus especies. Las especies con alta especificidad de hábitat, con peso corporal de 51–100 g, las que se alimentan en el estrato arbóreo y las que lo hacen en troncos tendieron a presentar una proporción de extinción mayor a lo esperado. Sin embargo, no hubo diferencias estadísticamente significativas entre la proporción de especies extintas observada y esperada para ninguna de las categorías (gremio trófico, estrato de alimentación, tamaño corporal y especificidad de hábitat), lo que indica que todos los grupos presentaron la misma vulnerabilidad a la extinción. El peso promedio de las aves extintas fue similar al de las aves aún presentes en los tres fragmentos. Las especies de las familias Grallaridae, Cotingidae, Thraupidae, Trogonidae y de la subfamilia Dendrocolaptinae fueron las más afectadas. Se discute por qué las características ecológicas evaluadas pueden no estar relacionadas con la extinción local de aves en estos fragmentos de bosque.
Palabras clave: Andes; Comunidad de aves; Extinción; Fragmentos boscosos; Vulnerabilidad.
Abstract
Local extinctions of birds in forest fragments in the Santa Elena region, central Andes, Colombia
We compared historic records from 1879 until 1952 of birds of Santa Elena region (Colombian central Andes) with those from a recent survey in three forest fragments of 41, 129 and 136 ha. Only 64 out of the 132 original species still remain in the three fragments; 78%, 66% and 63% of the original species have disappeared from the fragments. Species with high habitat specificity, those with body mass of 51–100 g, those which forage in the arboreal stratum, and those which forage on trunks showed a tendency to suffer more extinctions than expected. However, there were no statistical differences between observed and expected proportions of extinctions in any of the categories (trophic guild, foraging stratum, body mass and habitat specificity), indicating that all the bird groups presented the same vulnerability to extinction. Mean body mass of extinct birds was similar to that of the birds still remaining in the fragments. Species from the families Grallaridae, Cotingidae, Thraupidae, Trogonidae and from the subfamily Dendrocolaptinae were the most affected. We discuss why the evaluated ecological features could not be related with local extinction of bird species in these forest fragments.
Key words: Andes; Bird community; Extinction; Forest fragments; Vulnerability.
Recibido 28 septiembre 2006,
segunda versión corregida recibida 16 mayo 2008,
aceptado 17 agosto 2008
La fragmentación de los bosques tropicales ha sido reconocida como una de las mayores amenazas para la biodiversidad mundial (Laurance y Bierregaard 1997). En especial, se han documentado ampliamente sus efectos deletéreos (pérdida de la diversidad ocasionada por extinciones locales) sobre las comunidades de aves que éstos albergan (Turner 1996, Laurance et al. 2002). Algunas investigaciones han establecido que la vulnerabilidad de las especies de aves a la perturbación o alteración de su hábitat varía de acuerdo al gremio funcional al que pertenecen y a sus relaciones filogenéticas (Willis y Eisenmann 1979, Lovejoy et al. 1986, Bierregaard y Lovejoy 1989, Kattan et al. 1994, Stouffer y Bierregaard 1995, Renjifo 1999, Robinson 1999, Stratford y Stouffer 1999, Laurance et al. 2002, Sodhi et al. 2004). Se ha identificado, por ejemplo, que los frugívoros grandes del dosel, los insectívoros terrestres, las aves rapaces del interior del bosque y aquellas que se encuentran en su límite de distribución altitudinal son especialmente afectadas por la fragmentación (Kattan et al. 1994, Renjifo 1999, Kattan 2001). Lamentablemente, muy pocos estudios se han orientado a la determinación de los efectos de la fragmentación del hábitat en los Andes tropicales (Kattan y Álvarez-López 1996) y poco se conoce acerca de la respuesta de las comunidades de aves a la fragmentación en esta región.
En la región andina colombiana se concentran los mayores centros urbanos y gran parte de las actividades agrícolas y pecuarias del país; como consecuencia de esto, sus ecosistemas naturales han perdido el 60.3% de su extensión original (Villarreal 2006). El deterioro ambiental de la región, sumado al alto nivel de endemismo de aves presente en los Andes centrales, condujo a su definición como área prioritaria para la conservación mundial (Stattersfield et al. 1998, Myers et al. 2000). Además, de acuerdo a Renjifo et al. (2002), el 23% de las especies de aves en peligro crítico en Colombia habitan en ella. De esta manera, es fundamental la comprensión de los efectos a largo plazo de la fragmentación sobre su diversidad. Aunque se han realizado algunas investigaciones en aves (Kattan et al. 1994, Renjifo 1999), es poco lo que se ha avanzado en el conocimiento de la respuesta a la fragmentación por parte de las especies de aves que persisten en los bosques remanentes fragmentados, en especial en lo relacionado con su vulnerabilidad. El propósito de este trabajo es determinar, desde una perspectiva histórica (Kattan et al. 1994, Kattan 2001, Sodhi et al. 2004), la asociación entre la vulnerabilidad y algunas características ecológicas (tamaño corporal, gremio trófico, estrato de alimentación y especificidad de hábitat) de las aves que persisten en tres remanentes de bosque luego de más de 71 años de deforestación y fragmentación en la región de Santa Elena.
Métodos
Área de estudio
La zona rural de la cuenca de la quebrada Santa Elena (Parque Regional Arví, corregimiento de Santa Elena) está localizada en el sector centro-oriental del municipio de Medellín (6°15'N,75°34'O), en el departamento de Antioquia, Andes centrales de Colombia (Fig. 1). Posee un área total de 34 km2, comprendida entre los 2000–2720 msnm. Su régimen climático está determinado por la influencia del clima ecuatorial de montaña, con una temperatura promedio de 17 °C y una precipitación anual de 2067.5 mm (Empresas Públicas de Medellín 1988). El territorio corresponde a la zona de vida Bosque Muy Húmedo Montano Bajo (bmh-MB) según el sistema de clasificación de Holdridge (Espinal 1985).

Figura 1. Región de Santa Elena, Andes centrales, Colombia. Se muestran en gris los tres fragmentos de bosque en donde se realizó el estudio. Las líneas gruesas discontinuas indican carreteras.
Históricamente, esta cuenca ha sido altamente intervenida debido a su proximidad a la ciudad de Medellín (el núcleo urbano se originó en su parte baja) y porque desde sus orígenes ha sido su despensa agropecuaria. Evidencia de esto es que para 1914 esta localidad era descripta como un área cubierta por unos pocos parches de bosque y muchos pastizales (Chapman 1917). Actualmente, se conservan algunos remanentes de bosque intervenidos en áreas de difícil acceso, inmersos en una matriz de paisaje dominada por grandes extensiones de tierra dedicadas a actividades agropecuarias de uso intensivo (floricultura, horticultura y ganadería), plantaciones forestales con fines productivos y protectores (varias especies del género Pinus y Cupressus lusitanica), asentamientos humanos con bajas densidades (fincas de recreo y casas campestres) y la zona urbana de Medellín.
Los fragmentos de bosque estudiados se ubican entre los 2000–2680 msnm. El fragmento I, de 41 ha, está localizado en la parte alta de la cuenca de la quebrada Las Antenas, sin conexión con otros hábitats boscosos e inmerso en una matriz de fincas de recreo y de producción agropecuaria. El fragmento II tiene una extensión de 129 ha y forma parte de la cuenca de la quebrada La Castro. Está rodeado por áreas abiertas (potreros abandonados) y separado por franjas de plantaciones forestales (Pinus patula) ubicadas en la cuenca de la quebrada Piedras Blancas. El fragmento III, con un área de 136 ha, circunda a las quebradas La Aguada y Santa Elena y forma parte de una reserva protegida por la Corporación Autónoma Regional del Centro de Antioquia. Desde 1928, cuando se construyó la ruta que une a Medellín con Rionegro, los tres fragmentos se encuentran aislados (González 1999). Las características de la estructura de la comunidad de plantas de esta zona están descriptas en detalle en Castaño y Arias (1999) y la forma de los fragmentos en Castaño-Villa y Patiño-Zabala (2007).
Historia de los estudios ornitológicos de la región
La avifauna de la región de Santa Elena fue estudiada por TK Salmon en 1872 y sus registros publicados por Sclater y Salvin (1879). En 1914, L Miller y H Boyle estudiaron las aves de esta región como parte de la Expedición N°8 del American Museum of Natural History; sus resultados fueron publicados por Chapman (1917). Durante la segunda década del siglo XX, el hermano Nicéforo María colectó algunos ejemplares que se encuentran depositados actualmente en el Museo de Ciencias Naturales del Colegio San José de la Salle, en Medellín. Posteriormente, esta información fue compilada por Meyer de Schauensee (1948–1952) y Hilty y Brown (1986).
Colecta y análisis de datos
Entre el 5 de mayo de 1999 y el 30 de enero de 2000 se evaluó la riqueza de especies de aves en el interior de los tres fragmentos de bosque estudiados empleando puntos de conteo (Blondel et al. 1981). En los fragmentos de mayor tamaño (II y III) se ubicaron 12 puntos de conteo, mientras que en el I se ubicaron 9 puntos. En cada conteo, de una duración de 10 min, fueron registrados todos los individuos vistos y escuchados. Cada punto de conteo fue visitado en 10 ocasiones, a intervalos de tres semanas. También se realizaron búsquedas activas a través del interior de los fragmentos (entre las 06:00–11:30 h y entre las 14:00–18:00 h). Los fragmentos II y III fueron visitados durante 40 días cada uno, y el I durante 25 días, con un total de 315 h/persona de observación (120 h para los fragmentos II y III, y 75 h en el I).
Las extinciones locales, evaluadas para cada fragmento, fueron estimadas suponiendo que el número de especies presentes antes de la fragmentación era similar en todos los fragmentos. La estimación de la extinción observada (número de especies extintas localmente) presenta limitaciones asociadas a posibles subestimaciones en la riqueza de especies para esta región durante el siglo pasado, debido a que las técnicas empleadas para el registro de aves no se encontraban estandarizadas y a la imposibilidad para determinar posibles recolonizaciones durante el período en que ocurrieron las extinciones.
Las especies fueron categorizadas en grupos de acuerdo a su gremio trófico (frugívoras, frugívoro-insectívoras, frugívoro-insectívoro-nectarívoras, insectívoras, nectarívoro-insectívoras y rapaces), el estrato en el que se alimentan (arbóreo, sotobosque, troncos y varios), su tamaño corporal (menos de 25 g, 25–50 g, 51–100 g y más de 100 g; de acuerdo a Renjifo 1999) y su especificidad de hábitat (alta: especies que habitan bosques no perturbados, media: especies que raramente utilizan bosques perturbados o secundarios, baja: especies que utilizan regularmente hábitats modificados; de acuerdo a Loiselle y Blake 1992). La información se recopiló a partir de bibliografía (Hilty y Brown 1986, Stiles y Skutch 1989, Dunning 1993, Stotz et al. 1996) y observaciones de campo. La nomenclatura utilizada fue tomada de Remsen et al. (2007).
Para evaluar los patrones de extinción se compararon las distribuciones de las especies extintas en cada fragmento con una distribución esperada generada suponiendo que la proporción de especies extintas de cada grupo debería ser igual a la tasa total de extinción registrada en cada fragmento; las diferencias entre las distribuciones sugieren la tendencia de algunos grupos a ser más susceptibles a la extinción que otros (Kattan et al. 1994). Las distribuciones fueron comparadas por medio de la Prueba de G (Zar 1999), agrupando las categorías frugívoro-insectívora y frugívoro-insectívoro-nectarívora (gremios tróficos) para obtener valores esperados mayores a 5. Se empleó la Prueba U de Mann–Whitney para comparar el tamaño corporal de las especies extintas y las no extintas. No hubo control sobre las afinidades filogenéticas. Además, fueron excluidas de los análisis las especies cuyo patrón de distribución altitudinal se encuentra por fuera de los rangos del área de estudio, como es el caso de Pionopsitta pyrilia, que aunque fue registrada por Chapman (1917) tiene una distribución centrada en las tierras bajas tropicales (Hilty y Brown 1986, Stotz et al. 1996).
Resultados
La reconstrucción histórica de la avifauna de Santa Elena permitió establecer que para principios del siglo XX se encontraban asociadas a esta región 132 especies de aves residentes (Tabla 1). Actualmente, en los fragmentos persisten 64 de aquellas especies (Tabla 1), representadas en los fragmentos I, II y III por 28, 45 y 49 especies, respectivamente (i.e., han desaparecido el 78%, el 66% y el 63% de sus especies).
Tabla 1. Especies asociadas a bosques presentes en la región de Santa Elena, Andes centrales, Colombia. Para cada una se indica el gremio trófico al que pertenece, el estrato en el que se alimenta, la especificidad de hábitat, el tamaño corporal y su estatus local.
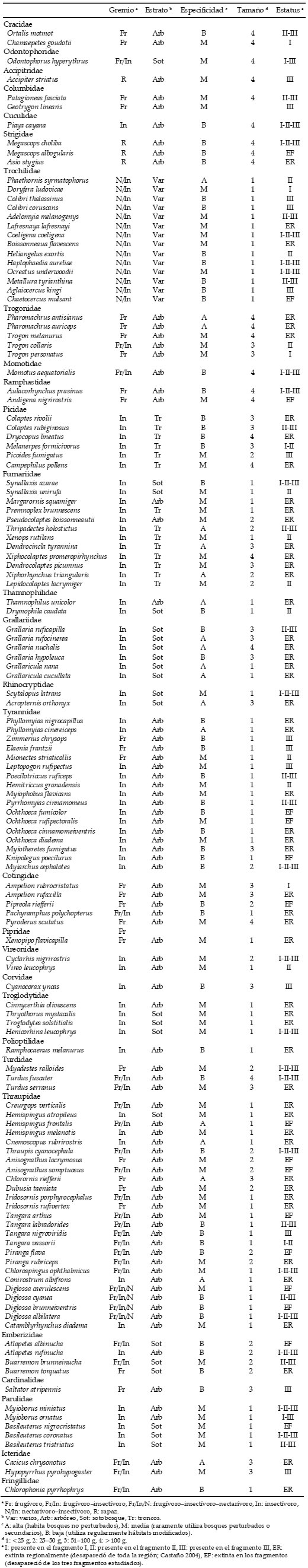
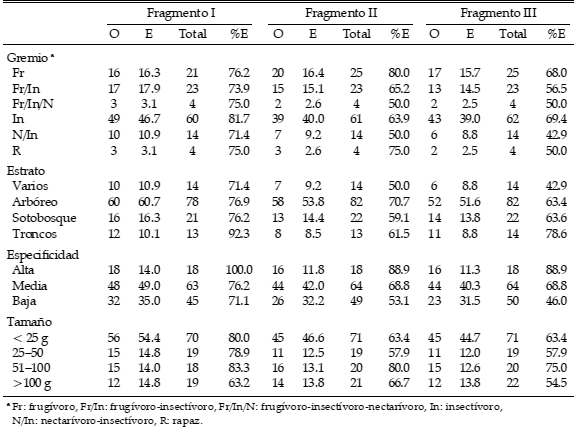
Aunque para las aves con alta especificidad de hábitat o con peso corporal entre 51–100 g se registró un número mayor de extinciones observadas que esperadas en todos los fragmentos (Tabla 2), no hubo diferencias significativas entre ambas distribuciones ni entre los grupos según la especificidad de hábitat (para el fragmento I: G = 0.67, gl = 2, P = 0.717; fragmento II: G = 1.36, gl = 2, P = 0.507; fragmento III: G = 2.20, gl = 2, P = 0.334) ni entre los grupos de distinto tamaño corporal (fragmento I: G = 0.34, gl = 3, P = 0.952; fragmento II: G = 0.42, gl = 3, P = 0.936; fragmento III: G = 0.28, gl = 3, P = 0.963). Entre las aves frugívoro-insectívoras y nectarívoro-insectívoras se registró un número menor de especies extintas que lo esperado, pero las diferencias tampoco fueron significativas entre los gremios tróficos (para el fragmento I: G = 0.22, gl = 3, P = 0.974; fragmento II: G = 0.38, gl = 3, P = 0.944; fragmento III: G = 1.38, gl = 3, P = 0.711; Tabla 2). En el fragmento II se registró un número de especies extintas mayor que el esperado entre las aves que se alimentan en el estrato arbóreo, y en los fragmentos I y III entre las que se alimentan en troncos, pero tampoco hubo diferencias entre ambas distribuciones entre los grupos según el estrato en el que se alimentan las especies (para el fragmento I: G = 0.21, gl = 3, P = 0.975; fragmento II: G = 0.63, gl = 3, P = 0.889; fragmento III: G = 0.76, gl = 3, P = 0.859).
Tabla 2. Número observado (O) y esperado (E) de especies extintas, número total de especies (total) y porcentaje de extinción (%E) para cada grupo de las diferentes categorías de aves (gremio trófico, estrato en el que se alimenta, especificidad de hábitat y tamaño corporal) en los tres fragmentos de bosque estudiados en la región de Santa Elena, Andes centrales, Colombia.
El peso promedio de las aves extintas fue similar al de las aves aún presentes en los tres fragmentos (P > 0.094, Prueba U de Mann–Whitney), tanto cuando se evaluaron las aves frugívoras (P > 0.5801) como las insectívoras (P > 0.2948), lo cual indica que el tamaño corporal por sí solo no está asociado a una mayor vulnerabilidad.
Más del 76% de las especies de las familias Grallaridae, Cotingidae, Thraupidae y de la subfamilia Dendrocolaptinae desaparecieron de los tres fragmentos; algo similar ocurrió con la familia Trogonidae, de la que solo dos especies persistieron en fragmentos diferentes (Tabla 1). Por el contrario, para los colibríes (Trochilidae) la proporción de especies extintas en los tres fragmentos fue inferior al 43%, mucho menor a la esperada para todos los fragmentos, que fue superior al 63%.
Discusión
En ninguna de las categorías de aves se observó una propensión a la extinción (i.e., una proporción de extinciones mayor a lo esperado). Este resultado se contrapone al de varios trabajos en los que se ha registrado una mayor vulnerabilidad en algunos gremios tróficos (e.g., insectívoros y frugívoros), para especies que se alimentan en algunos estratos particulares o para especies con cierta especificidad de hábitat (Newmark 1991, Kattan 1992, Kattan et al. 1994, Ribon et al. 2003, Castaño 2004). Otros autores, sin embargo, tampoco han encontrado una relación entre características ecológicas como estrato de forrajeo, tamaño corporal o gremio trófico y una mayor propensión a la extinción en aves o mamíferos en fragmentos de bosque (Laurance 1991, Renjifo 1999, Crooks 2002, dos Anjos 2006). Al respecto, Laurance (1991) determinó que la abundancia de los mamíferos en la matriz circundante del paisaje fragmentado en Australia fue un mejor predictor de su vulnerabilidad que características ecológicas como tamaño corporal, nivel trófico o especialización en la dieta, debido a que los hábitats modificados en torno a los fragmentos permiten la dispersión o supervivencia de las especies que usan la matriz. También ha sido reportado que otras características muy variables entre las especies de aves, como la capacidad para tolerar el deterioro del hábitat y la de dispersión, afectan su persistencia dentro de un paisaje alterado en fragmentos en Kenia (Lens et al. 2002).
Algunos de los elementos anteriormente descriptos podrían tener relación con la riqueza de especies actual de los fragmentos y con la no detección de grupos con mayor vulnerabilidad en relación a las variables evaluadas. Es posible que otras características ecológicas no incluidas en esta investigación tengan una relación más estrecha con la extinción local. En la actualidad, en los fragmentos persisten únicamente dos especies con alta especificidad de hábitat (Phaethornis syrmatophorus y Thripadectes holostictus) y un 80% de las aves que aún se encuentran en ellos están asociadas a los bordes (Castaño-Villa y Patiño-Zabala 2007). Estas aves se desplazan a través de la matriz del paisaje (e.g., cañadas arboladas, cercos), por lo que poseen cierta capacidad para dispersarse a través de la cuenca. Esto puede favorecer su persistencia en pequeños fragmentos, como ha sido mencionado para la Amazonia (Antogiovanni y Metzger 2005). Por el contrario, las especies con alta especificidad de hábitat y que además tienen limitaciones para la dispersión a través de la matriz del paisaje podrían ser más vulnerables. El análisis en conjunto de un número mayor de características ecológicas de las especies puede conducir a la identificación de patrones de vulnerabilidad, pero esto podría seguir siendo muy ambicioso, ya que, como han registrado Ferraz et al. (2003) en fragmentos en la Amazonia brasileña, cada especie difiere ampliamente en su probabilidad de extinción.
Otros factores como la cacería o la abundancia de mesopredadores en los fragmentos podrían estar más relacionados con la desaparición de algunas especies que sus características ecológicas. Este puede ser el caso de la Pava Maraquera (Chamaepetes goudotii), que aunque es frugívora, arborícola y de gran tamaño, aún se conserva en el fragmento más pequeño, el único en donde no se registró cacería. Otras especies que son traficadas como mascotas (Anisognathus lacrymosus y Anisognathus somptuosus, ambas frugívoras de tamaño intermedio) han desaparecido de todos los fragmentos quizás debido a su cacería. La desaparición de algunas especies en los fragmentos puede estar asociada a la predación de nidos por serpientes (Loiselle y Hoppes 1983), ya que en todos ellos son comunes varias especies (e.g., Liophis pseudocobella y Lampropeltis triangulum) que pueden amenazar la persistencia de las aves. Además, el tamaño y la ubicación de los fragmentos (en áreas suburbanas y rurales) pueden incrementar las tasas de predación de nidos (Arango-Vélez y Kattan 1997, Danielson et al. 1997).
Es importante resaltar que el 80% de las aves insectívoras de más de 100 g (e.g., Campephilus pollens y Xiphocolaptes promeropirhynchus) desaparecieron de todos los fragmentos; es decir que bosques con áreas menores a 136 ha parecen ser poco capaces de soportar poblaciones de especies con estas características a largo plazo. Esto concuerda con lo reportado por Willis (1979) y Christiansen y Pitter (1997), quienes aseguran que este grupo de aves puede desaparecer de los fragmentos pequeños debido a sus requerimientos de grandes territorios. La tendencia observada de una mayor extinción en aves de 51–100 g en todos los fragmentos se debe posiblemente a una combinación de altos requerimientos metabólicos diarios y baja movilidad en organismos de tamaño intermedio, que los haría más vulnerables (véanse citas en Cardillo y Bromhan 2001).
Los mayores porcentajes de extinción observados en Cotingidae, Grallariidae y Dendrocolaptinae, así como la baja sensibilidad registrada en Trochilidae, concuerdan con lo encontrado en otras investigaciones (Leck 1979, Johnels y Cuadros 1986, Kattan et al. 1994, Stouffer y Bierregard 1995, Bierregard y Stouffer 1997, Renjifo 1999, Ribon et al. 2003). Las familias Trogonidae y Thraupidae emergen como grupos altamente vulnerables ante la fragmentación a largo plazo. Entre los Trogonidae, Pharomachrus antisianus y Pharomachrus auriceps desaparecieron de todos los fragmentos, mientras que Trogon collaris y Trogon personatus fueron registradas solamente en dos ocasiones en los fragmentos I y II, respectivamente. Pharomachrus auriceps también desapareció de otra región andina en Colombia (Kattan et al. 1994). Esto se ha explicado en función de su condición de migradora altitudinal (Powell y Bjork 1995); aunque en la zona estudiada hay bosques a otras alturas, en su mayoría se encuentran muy distantes, lo cual puede conducir a una baja supervivencia de esta especie en los fragmentos. Explicar la mayor vulnerabilidad de las especies de Thraupidae es más complicado, debido a la amplia gama de hábitos que se presentan en esta familia.
En relación con las prioridades para la conservación de aves en Colombia, de las cuatro especies endémicas dos han desaparecido de estos fragmentos (Grallaria rufocinerea y Xenopipo flavicapilla), mientras que Odontophorus hyperythrus y Hypopyrrhus pyrohypogaster aún persisten en algunos de ellos, aunque en grupos muy pequeños. Es de especial interés los casos de Grallaria rufocinerea, Grallaria cucullata, Ampelion rufaxilla, Xenopipo flavicapilla e Iridosornis porphyrocephalus, que se encuentran en alguna categoría de amenaza de acuerdo al Libro Rojo de Aves de Colombia (Renjifo et al. 2002), ya que todas ellas han desaparecido de los tres fragmentos estudiados (que quizás son los remanentes de bosque de mayor área en la región).
Por último, aunque no fue posible encontrar una relación entre las características ecológicas evaluadas y la propensión a la extinción entre las aves en estos tres fragmentos, se evidenciaron los posibles efectos deletéreos de la fragmentación a largo plazo sobre la diversidad de aves en una región de los Andes centrales colombianos. A pesar de que los resultados de esta investigación pueden hacer pensar que estos pequeños fragmentos poseen un bajo valor para la conservación, ellos cumplen un importante papel para la investigación y la educación, y además forman parte del patrimonio ambiental y territorial de esta región.
Agradecimientos. Los autores expresan su agradecimiento a Álvaro Lema Tapias, Jaime Vicente Estévez, Luisa Fernanda Lema y Juan Alejandro Morales, y a otros correctores anónimos por sus comentarios y sugerencias. A la Sociedad Antioqueña de Ornitología que aportó recursos para el desarrollo de la primera fase de esta investigación de la cual participaron en la colección de datos de campo Alejandro Arias, Ana Castaño, Pablo Flórez, Carlos Delgado y Julio Saenz; así como a Tomás Cuadros Carvajal por su apoyo y asesoría durante esta investigación. Por último, al proyecto Conservación de la biodiversidad de los Andes en Colombia de la Universidad de Caldas.
Bibliografía Citada
dos Anjos L (2006) Bird species sensitivity in a fragmented landscape of the Atlantic forest in Southern Brazil. Biotropica 38:229–234 [ Links ]
Antogiovanni M y Metzger JP (2005) Influence of matrix habitats on the occurrence of insectivorous birds species in Amazonian forest fragments. Biological Conservation 122:441–451 [ Links ]
Arango-Vélez N y Kattan GH (1997) Effects of forest fragmentation on experimental nest predation in Andean cloud forest. Biological Conservation 81:137–143 [ Links ]
Bierregaard RO y Lovejoy TE (1989) Effects of forest fragmentation on Amazonian understory bird communities. Acta Amazonica 19:215–241 [ Links ]
Bierregaard RO y Stouffer PC (1997) Understory birds and dynamic habitat mosaics in Amazonian rainforests. Pp. 138–155 en: Laurance WE y Bierregaard RO (eds) Tropical forest remmants. Ecology, management, and conservation of fragmented communities. University of Chicago Press, Chicago [ Links ]
Blondel J, Ferry C y Frochot B (1981) Point counts with unlimited distance. Studies in Avian Biology 6:414–420 [ Links ]
Cardillo M y Bromhan L (2001) Body size and risk of extinction in Australian mammals. Conservation Biology 15:1435–1440 [ Links ]
Castaño GJ (2004) Evaluación del riesgo de extinción de aves en hábitats fragmentados a largo plazo mediante el análisis de sus características ecológicas. Tesis de Maestría, Universidad Nacional de Colombia, Medellín [ Links ]
Castaño GJ y Arias A (1999) Evaluación de la avifauna de la microcuenca de la quebrada Santa Helena, zona centro oriental de Medellín. Instituto para el Manejo Integral de la Cuenca del Río Medellín y Sociedad Antioqueña de Ornitología, Medellín [ Links ]
Castaño-Villa GJ y Patiño-Zabala JC (2007) Composición de la comunidad de aves en bosques fragmentados en la región de Santa Elena. Boletín Científico del Centro de Museos 11:47–60 [ Links ]
Chapman FM (1917) The distribution of bird-life in Colombia: a contribution to a biological survey of South America. Bulletin of the American Museum of Natural History 36:1–729 [ Links ]
Christiansen MB y Pitter E (1997) Species loss in a forest bird community near Lago Santa in southeastern of Brazil. Biological Conservation 80:23–32 [ Links ]
Crooks KR (2002) Relative sensitivities of mammalian carnivores to habitat fragmentation. Conservation Biology 16:488–502 [ Links ]
Danielson WR, DeGraaf RM y Fuller TK (1997) Rural and suburban forest edges: effect on egg predators and nest predation rates. Landscape and Urban Planning 38:25–36 [ Links ]
Dunning JB Jr (1993) Handbook of avian body masses. CRC Press, Boca Raton [ Links ]
Empresas Públicas de Medellín (1988) Cuenca de la quebrada Piedras Blancas: estudio geomorfológico. Empresas Públicas de Medellín, Medellín [ Links ]
Espinal LS (1985) Geografía ecológica del departamento de Antioquia (Zonas de vida–Formaciones vegetales). Revista de la Facultad Nacional de Agronomía 38:24–39 [ Links ]
Ferraz G, Russell GF, Stouffer PC, Bierregaard RO, Pimm SL y Lovejoy TE (2003) Rates of species loss from Amazonian forest fragments. Proceedings of the National Academy of Sciences 100:14069–14073 [ Links ]
González LF (1999) Caminos republicanos en Antioquia. Los caminos de Medellín a Rionegro, las rutas por Santa Elena 1800–1928. Corporación Autónoma Regional del Centro de Antioquia, Medellín [ Links ]
Hilty SL y Brown WL (1986) A guide to the birds of Colombia. Princeton University Press, Princeton [ Links ]
Johnels SA y Cuadros TC (1986) Species composition and abundance of bird fauna in a disturbed forest in the central Andes of Colombia. Hornero 12:235–241 [ Links ]
Kattan GH (1992) Rarity and vulnerability: the birds of the Cordillera Central of Colombia. Conservation Biology 6:64–70 [ Links ]
Kattan GH (2001) Extinción de especies y fragmentación del hábitat en el Neotrópico. Pp. 205–206 en: Primack R, Rozzi R, Feinsinger P, Dirzo R y Massardo F (eds) Fundamentos de conservación biológica: perspectivas latinoamericanas. Fondo de Cultura Económica, México DF [ Links ]
Kattan GH y Álvarez-López H (1996) Preservation and management of biodiversity in fragmented landscapes in the Colombia Andes. Pp. 3–18 en: Schelhas J y Greenberg R (eds) Forest patches in tropical landscapes. Island Press, Washington DC [ Links ]
Kattan GH, Álvarez-López H y Giraldo M (1994) Forest fragmentation and bird extinctions: San Antonio eighty years later. Conservation Biology 8:138–146 [ Links ]
Laurance WF (1991) Ecological correlates of extinction proneness in Australian tropical rain forest mammals. Conservation Biology 5:79–89 [ Links ]
Laurance WF y Bierregaard RO (1997) Tropical forest remnants: ecology, management, and conservation of fragmented communities. University of Chicago Press, Chicago [ Links ]
Laurance WF, Lovejoy TE, Vasconcelos HL, Bruna EM, Didham RK, Stouffer PC, Gascon C, Bierregaard RO, Laurance SG y Sampaio E (2002) Ecosystem decay of Amazon forest fragments: a 22-year investigation. Conservation Biology 16:605–618 [ Links ]
Leck CF (1979) Avian extinctions in an isolated tropical wet forest preserve, Ecuador. Auk 96:343–352 [ Links ]
Lens L, Van Dongen S, Norris K, Githiru M y Matthysen E (2002) Avian persistence in fragmented rainforest. Science 298:1236–1238 [ Links ]
Loiselle BA y Blake JG (1992) Population variation in a tropical bird community: implications for conservation. BioScience 42:838–845 [ Links ]
Loiselle BA y Hoppes WG (1983) Nest predation in insular and mainland lowland forest in Panama. Condor 85:93–95 [ Links ]
Lovejoy TE, Bierregaard RO Jr, Rylands AB, Malcolm JR, Quintela CE, Harper LH, Brown KS Jr, Powell AH, Powell GVN, Schubart HOR y Hays MB (1986) Edge and other effects of isolation on Amazon forest fragments. Pp. 257–285 en: Soulé ME (ed) Conservation Biology. The science of scarcity and diversity. Sinauer Associates, Sunderland [ Links ]
Meyer de Schauensee R (1948–1952) The birds of the Republic of Colombia. Caldasia 5:251–1224 [ Links ]
Myers N, Mittermeier RA, Mittermeier CG, Da Fonseca GAB y Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858 [ Links ]
Newmark WD (1991) Tropical forest fragmentation and the local extinction of understory birds in the Eastern Usambara mountains, Tanzania. Conservation Biology 5:67–78 [ Links ]
Powell GV y Bjork R (1995) Implications of intratropical migration on reserve design: a case study using Pharomachrus mocinno. Conservation Biology 9:354–362 [ Links ]
Remsen JV, Cadena CD, Jaramillo A, Nores M, Pacheco JF, Robbins MB, Schulenberg TS, Stiles FG, Stotz DF y Zimmer KJ (2007) A classification of the bird species of South America. American Ornithologists' Union, Baton Rouge (URL: http://www.museum.lsu.edu/~Remsen/SACCBaseline.html) [ Links ]
Renjifo LM (1999) Composition changes in a subandean avifauna after long-term forest fragmentation. Conservation Biology 13:1124–1139 [ Links ]
Renjifo LM, Franco-Maya AM, Amaya-Espinel JD, Kattan GH y López-Lanús B (2002) Libro rojo de aves de Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt y Ministerio del Medio Ambiente, Bogotá [ Links ]
Ribon R, Simon JE y De Mattos GT (2003) Bird extinctions in Atlantic forest fragments of the Viçosa region, southeastern Brazil. Conservation Biology 17:1827–1839 [ Links ]
Robinson WD (1999) Long-term changes in the avifauna of Barro Colorado Island, Panama, a tropical forest isolate. Conservation Biology 13:85–97 [ Links ]
Sclater PL y Salvin O (1879) On the birds collected by the late Mr. T. K. Salmon in the State of Antioquia, United States of Colombia. Proceedings of the Zoological Society of London 1879:486–550 [ Links ]
Sodhi NS, Liow LH y Bazzaz FA (2004) Avian extinctions from tropical and subtropical forests. Annual Review of Ecology, Evolution and Systematics 35:323–345 [ Links ]
Stattersfield AJ, Crosby MJ, Long AJ y Wege DC (1998) Endemic bird areas of the world. Priorities for biodiversity conservation. Burlington Press, Cambridge [ Links ]
Stiles FG y Skutch AF (1989) A guide to the birds of Costa Rica. Cornell University Press, Ithaca [ Links ]
Stotz DF, Fitzpatrick JW, Parker TA III y Moskovist DK (1996) Neotropical birds: ecology and conservation. University of Chicago Press, Chicago [ Links ]
Stouffer PC y Bierregaard RO Jr (1995) Effects of forest fragmentation on understory hummingbirds in Amazonian Brazil. Conservation Biology 9:1085–1094 [ Links ]
Stratford JA y Stouffer PC (1999) Local extinctions of terrestrial insectivorous birds in a fragmented landscape near Manaus, Brazil. Conservation Biology 13:1416–1423 [ Links ]
Turner IM (1996) Species loss in fragments of tropical rainforest: a review of the evidence. Journal of Applied Ecology 33:200–209 [ Links ]
Villareal H (2006) Ecosistemas terrestres naturales. Pp. 43–66 en: Chaves ME y Santamaría M (eds) Informe sobre el avance en el conocimiento y la información de la biodiversidad 1998–2004. Tomo II. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá [ Links ]
Willis EO (1979) The composition of avian communities in remanescent woodlots in southern Brazil. Papéis Avulsos de Zoologia 33:1–25 [ Links ]
Willis EO y Eisenmann E (1979) A revised list of birds of Barro Colorado Island, Panama. Smithsonian Contributions to Zoology 291:1–31 [ Links ]
Zar JH (1999) Biostatistical analysis. Cuarta edición. Prentice-Hall, Upper Saddle River [ Links ]

