










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEl hornero
versión impresa ISSN 0073-3407
Hornero vol.28 no.1 Ciudad Autónoma de Buenos Aires ago. 2013
ARTÍCULOS
Dieta del Chorlito Doble Collar (Charadrius falklandicus) en Península Valdés, Patagonia, Argentina
Luciana Musmeci1,2,3,4, Luis O. Bala1,3 y María de los Ángeles Hernández1,3
1 Universidad Nacional de la Patagonia San Juan Bosco. Boulevard Brown 3100, 9120 Puerto Madryn, Chubut, Argentina.
2 Fundación Patagonia Natural. Marcos A. Zar 760, 9120 Puerto Madryn, Chubut, Argentina.
3 Centro Nacional Patagónico. Boulevard Brown 2915, 9120 Puerto Madryn, Chubut, Argentina.
4 lumusmeci@cenpat.edu.ar
Recibido 29 noviembre 2012, aceptado 30 agosto 2013
RESUMEN
El objetivo de este trabajo fue evaluar la dieta del Chorlito Doble Collar (Charadrius falklandicus) en Playa Colombo (Golfo Nuevo, Península Valdés) y compararla con la registrada previamente en Playa Fracasso (Golfo San José), sitios de características similares distanciados por 21 km. En febrero y marzo de 2003 se colectaron 49 heces y se conservaron individualmente. La reconstrucción de la dieta se realizó mediante el reconocimiento de estructuras clave presentes en las heces. Se identificaron siete ítems presa, siendo los más importantes en orden decreciente el poliqueto Travisia olens, la almeja Darina solenoides y los insectos. Se registraron 1.3 almejas por excremento, de una talla promedio de 10.6 mm y un equivalente en biomasa (expresada como peso seco libre de cenizas) de 6.4 mg. Se estimó la abundancia de invertebrados bentónicos para determinar la oferta trófica. Los chorlitos seleccionaron tallas de almejas de 4-12 mm. La dieta en los dos meses fue similar. En Playa Fracasso la presa más importante es Darina solenoides, seguida por Travisia olens. En comparación con Playa Fracasso, en Playa Colombo los chorlitos ingirieron una menor cantidad de almejas, pero de mayor talla y mayor biomasa.
PALABRAS CLAVE: Alimentación; Aves playeras; Charadrius falklandicus; Oferta trófica; Playa Colombo; Presas; Selección de tallas.
ABSTRACT
Diet of the Two-banded Plover (Charadrius falklandicus) in Península Valdés, Patagonia, Argentina
The objective of this study was to evaluate the diet of the Two-banded Plover (Charadrius falklandicus) in Playa Colombo (Golfo Nuevo, Península Valdés), and to compare it with the diet recorded in previous studies in Playa Fracasso (Golfo San José), a site with similar characteristics 21 km away. In February and March 2003, we collected 49 faeces that were conserved individually. The diet was reconstructed based on the identification of key structures present in the faeces. Seven prey items were identified, with the most important components being, in decreasing order, the polychaete Travisia olens, the clam Darina solenoides and insects. We recorded 1.3 clams per sample, with a mean size of 10.6 mm and an equivalent biomass (expressed as ash-free dry weight) of 6.4 mg. We estimated benthic invertebrate abundance in order to measure the trophic offer. Plovers selected clam sizes of 4-12 mm. The diet was similar between months. The most important prey in Playa Fracasso is Darina solenoides, followed by Travisia olens. Plovers eat a lower quantity of clams, but with higher size and biomass, in Playa Colombo than in Playa Fracasso.
KEY WORDS: Charadrius falklandicus; Foraging; Playa Colombo; Prey; Shorebirds; Size selection; Trophic offer.
La Península Valdés, ubicada en el noreste de la Patagonia argentina, es utilizada por diversas especies de aves playeras entre las cuales se incluye el Chorlito Doble Collar (Charadrius falklandicus) (Bala et al. 2001, 2002). Esta especie posee migración neotropical, nidificando en la Patagonia durante el verano y migrando parcial o totalmente hacia el norte en el período no reproductivo (Blanco et al. 2006). García-Peña et al. (2008) indican que en la zona esta especie se reproduce entre octubre y diciembre. Dentro de Península Valdés, el Chorlito Doble Collar utiliza distintos intermareales para alimentarse y reproducirse, entre los que se destacan Playa Fracasso (Bala et al. 2001, 2002, García-Peña et al. 2008), ubicada en el Golfo San José, y Playa Colombo (Musmeci 2005), ubicada en el Golfo Nuevo (Fig. 1). Ambos intermareales, distanciados por 21 km, son de características similares: presentan principalmente arena fina, son de forma semicircular y en bajamar presentan gran amplitud. Están sometidos a un régimen de marea semidiurno que determina la disponibilidad de alimento para las aves playeras. Las mareas en los golfos Nuevo y San José se encuentran desfasadas en medio ciclo; cuando en uno de ellos es pleamar en el otro es bajamar y viceversa.

Figura 1. Ubicación de Playa Colombo (sitio de estudio) y Playa Fracasso en Península Valdés, en el noreste de la Patagonia argentina.
Se cuenta con estudios previos de dieta del Chorlito Doble Collar realizados en Playa Fracasso (D'Amico 2001, D'Amico y Bala 2004, D'Amico et al. 2004). Allí, los chorlitos se alimentan mayormente de la almeja Darina solenoides (77%), el poliqueto Travisia olens (47%), insectos (43%) y el cangrejo Cyrtograpsus affinis (41%), entre otras presas de menor importancia como Glycera americana (13%). Los autores reportaron un promedio de 4.6 almejas por excremento y que las aves seleccionan positivamente tallas de almejas comprendidas en el rango 1-4 mm (con una talla promedio de 2.1 mm). En Playa Fracasso, el Chorlito Doble Collar tiene una presencia regular entre marzo y agosto, con una abundancia máxima en marzo y abril (Pagnoni 1997, D'Amico 2001), periodo que coincide con los registros de abundancias en Playa Colombo (Musmeci 2005).
El objetivo de este estudio fue evaluar la dieta del Chorlito Doble Collar en Playa Colombo y compararla con la registrada en Playa Fracasso, a fin de determinar si los chorlitos aprovechan de manera diferente los recursos tróficos en ambos sitios. Estas aves representan el último eslabón de la cadena trófica que incluye a las comunidades de invertebrados bentónicos en estas playas, que a su vez representan una alta oferta de alimento. Conocer cómo las aves playeras utilizan los ambientes en los sitios de estadía y reproducción, así como a lo largo de su migración, es importante para la conservación de estas especies, debido a que dependen del buen estado de dichos sitios.
MÉTODOS
Sitio de estudio
Playa Colombo está ubicada en el Golfo Nuevo (42°38'S, 64°14'O; Fig. 1), en la provincia de Chubut. Presenta una longitud de 4.5 km y un intermareal que alcanza una extensión de 1.8 km en su parte central durante la bajamar. Las mareas presentan una amplitud promedio de 3.93 m y máxima de 5.76 m.
Oferta trófica
Se realizaron muestreos del bentos a lo largo de dos transectas perpendiculares a la línea de marea en febrero de 2003 con el fin de ponderar la oferta trófica para las aves. Se consideraron diferentes estaciones de muestreo, en función de la altura del nivel de marea (cada 0.5 m de altura), las cuales quedaron establecidas de acuerdo con el plano de reducción de las tablas de marea del Servicio de Hidrografía Naval. Cada una de las transectas se dividió desde el nivel 1 (1 m de altura, correspondiente a la bajamar) hasta el nivel 4 (4 m de altura, correspondiente a los horizontes superiores de la playa). En cada estación se obtuvieron seis muestras con un cilindro de 20 cm de diámetro interno y 35 cm de profundidad, que se utilizó para extraer panes de sustrato con los organismos infaunales incluidos. Se lavó in situ cada muestra con un cedazo metálico de 0.50 mm de abertura de malla y los organismos encontrados se conservaron a -20 °C hasta su posterior análisis. En el laboratorio, las muestras fueron lavadas sobre dos tamices superpuestos de 1.00 mm y 0.50 mm de abertura de malla y se identificaron, midieron y cuantificaron los organismos presentes.
Análisis de la dieta
Una de las técnicas para el estudio de la dieta en aves playeras es el análisis de heces o egagrópilas. Los resultados que se obtienen tienen alto nivel de confianza, siendo una técnica de uso aceptada en la comunidad científica (Dekinga y Piersma 1993, González et al. 1996, Petracci 2002, Hernández et al. 2004, 2008). Puede presentar dificultades cuando las aves se alimentan de presas que no contienen estructuras duras, pudiendo únicamente realizarse inferencias cualitativas, subestimando la ingesta total (Barret et al. 2007). El tiempo de retención de la comida en el tracto digestivo de las aves playeras parece ser corto (20-50 min en el Playero Rojizo, Calidris canutus; Piersma 1994) y debido a que ingieren sus presas enteras el análisis de las heces revela qué han comido localmente (Van Gils et al. 2005).
Se colectaron 49 heces (34 el 24 de febrero y 15 el 21 de marzo de 2003) de bandadas monoespecíficas de Chorlito Doble Collar luego de observar a los individuos alimentándose en el lugar como mínimo unos 50 min, para asegurar que las heces se correspondieran con la ingesta en el sitio. Durante los muestreos la marea estaba bajando y las heces fueron colectadas entre las alturas del nivel de marea de 3 y 3.5 m. Las heces se conservaron individualmente a -20 °C.
Para reconstruir la dieta se utilizó la metodología descripta por Dekinga y Piersma (1993) con modificaciones (Hernández et al. 2004), identificando estructuras clave presentes en las heces que permiten reconocer las especies componentes de la dieta (e.g., restos de valvas de moluscos, quetas y mandíbulas de poliquetos, apéndices de crustáceos e insectos). Se disgregó cada muestra en agua y se la observó bajo lupa binocular, verificando la presencia o ausencia de quetas de poliquetos o pequeñas estructuras tales como apéndices de crustáceos que pueden sobrenadar en el medio. Luego, cada muestra se tamizó sobre un cedazo de 0.25 mm de abertura de malla y el material retenido se observó nuevamente bajo lupa a fin de extraer los elementos clave para identificar las presas consumidas. Para el caso particular de Darina solenoides es posible determinar si las charnelas encontradas corresponden a valvas derechas o izquierdas. Así, los condróforos de igual tamaño correspondientes a valvas opuestas se consideraron como pertenecientes a una misma almeja, permitiendo cuantificar el número de individuos ingeridos por excremento. Además, con mediciones de estas estructuras es posible estimar la talla (en mm) y la biomasa (en mg de peso seco libre de cenizas) de las almejas al momento de ser ingeridas, usando análisis de regresión. La regresión entre la altura de la charnela (h) y la talla fue: Talla = 13.476 h - 0.1191 (r = 0.99, n = 165). La regresión entre la talla y la biomasa fue: Biomasa = 0.0000094 Talla 2.60196 (r = 0.98, n = 60).
Debido a que no todas las presas ingeridas son cuantificables, se calculó la frecuencia de ocurrencia de presas (en porcentaje) con respecto al total de heces examinadas. Entre estas presas no cuantificables se encuentran poliquetos no mandibulados (identificables por la presencia de sus quetas), crustáceos (por la presencia de apéndices) e insectos (por la presencia de artejos).
Se examinó la relación entre las tallas de almejas ingeridas y las que se encontraban disponibles en el intermareal. Para determinar la clase de tallas de individuos seleccionados positivamente por las aves se utilizó el índice de Savage (Manly et al. 1993): Wi= Ui /pi, donde Ui corresponde a la proporción de cada clase de talla de presa consumida y pi es la proporción de dicha clase de talla presente en la población, calculada como la proporción de almejas de dicha clase de talla respecto de la totalidad de almejas ofertadas de todas las tallas. El estadístico presenta un valor crítico asociado a una distribución chi-cuadrado con un grado de libertad (Manly et al. 1993).
Para evaluar si existen diferencias en la dieta entre meses (febrero y marzo) se utilizó un Análisis de Similitud (ANOSIM) usando el programa PRIMER v5 (Plymouth Routines In Multivariate Ecological Research; Clarke y Warwick 2001). Se obtuvo el valor del estadístico R, que varía entre -1 y 1 y para el cual los valores cercanos a 0 indican la similitud máxima. Utilizando una matriz de Bray-Curtis se estimó la similitud entre muestras de cada mes mediante un Análisis de Similitud Porcentual (SIMPER), que permite identificar la contribución de cada ítem presa a dichos valores de similitud.
RESULTADOS
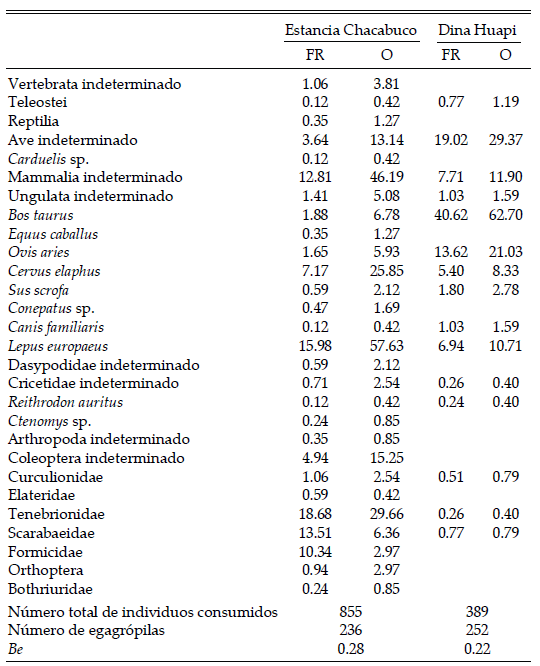
La almeja Darina solenoides presentó mayores abundancias en los niveles de marea superiores (Tabla 1). El poliqueto Travisia olens, encontrado en todos los niveles, mostró abundancias heterogéneas a lo largo de los niveles de marea. El gasterópodo Buccinanops globulosus y el ostrácodo Cycloleberis pouseni solo se encontraron en la parte inferior del intermareal. Los anfípodos tuvieron mayores abundancias en los sectores intermedio e inferior, mientras que, por el contario, el isópodo Excirolana armata fue más abundante en los niveles superiores.
Tabla 1. Abundancia promedio (± DE) de los invertebrados presentes a distintas alturas del nivel de marea en Playa Colombo, Península Valdés, noreste de la Patagonia argentina. La altura de 1.5 m corresponde a bajamar y la de 4.0 m a pleamar. Los valores están expresados en individuos/m2.
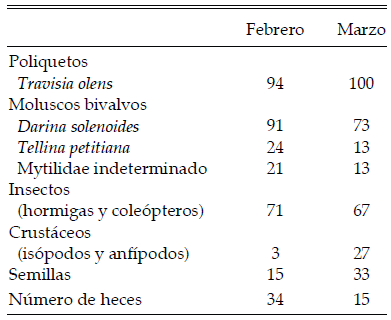
Se distinguieron siete ítems presa en la dieta del Chorlito Doble Collar (Tabla 2). Travisia olens estuvo presente en casi todas las muestras, seguido en orden de importancia por Darina solenoides y los insectos (hormigas y coleópteros). Otras especies con menor presencia relativa fueron la almeja Tellina petitiana, semillas, crustáceos (anfípodos e isópodos) y mitílidos.
Tabla 2. Frecuencia de ocurrencia (en porcentaje) de los distintos ítems presa encontrados en la dieta del Chorlito Doble Collar (Charadrius falklandicus) en Playa Colombo, Península Valdés, noreste de la Patagonia argentina, en febrero y marzo de 2003.
El número promedio (±DE) de individuos de Darina solenoides presentes por excremento para las muestras de febrero fue de 1.3±1.5 (n = 34), con una talla de 10.6±4.9 mm (n = 45). Estos valores representan un aporte en biomasa (expresada como peso seco libre de cenizas) de 6.4±9.2 mg por excremento (n = 34).
En las heces colectadas en marzo, Darina solenoides tuvo alta frecuencia de ocurrencia pero en pocas oportunidades se logró medir su charnela.
Las tallas de almejas ingeridas por el Chorlito Doble Collar variaron entre 3.9-24.1 mm (Fig. 2), mientras que en el intermareal se encontraba disponible un amplio rango de tallas (2-32 mm), con dos cohortes de individuos con tallas de 10 y 20 mm, respectivamente. Los valores del índice de Savage indican que los chorlitos seleccionaron positivamente (P < 0.001) las tallas comprendidas en el rango 4-12 mm.
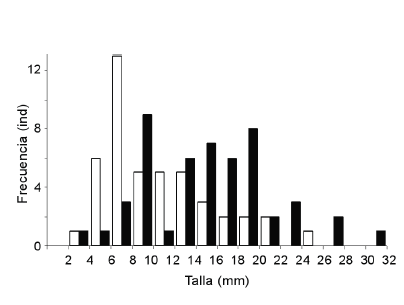
Figura 2. Distribución de frecuencias de las tallas disponibles de la almeja Darina solenoides (barras negras) y de las tallas ingeridas por el Chorlito Doble Collar (Charadrius falklandicus) (barras blancas) en Playa Colombo, Península Valdés, noreste de la Patagonia argentina.
La dieta fue similar en los dos meses (R = 0.107, P = 0.058), diferenciándose en un 32.0%. Las muestras de febrero fueron similares en un 64.9% y las de marzo en un 71.9%. En ambos casos, más del 75% de la similitud se debió al aporte de dos ítems presa: Travisia olens (febrero 49.4%, marzo 39.2%) y Darina solenoides (febrero 25.7%, marzo 36.1%).
DISCUSIÓN
En Playa Colombo (Golfo Nuevo), el Chorlito Doble Collar muestra una amplia preferencia por el poliqueto Travisia olens, seguida en importancia por un par de ítems presa: la almeja Darina solenoides e insectos (hormigas y coleópteros). Esta preferencia es inversa a la reportada en Playa Fracasso (Golfo San José), donde la presa más importante es Darina solenoides, seguida por Travisia olens (D'Amico 2001, D'Amico y Bala 2004). Sin embargo, en ambas playas las presas principales son estas mismas dos especies. La ausencia de Glycera americana en la dieta en Playa Colombo se debe a que allí esta especie está ausente.
Los chorlitos ingirieron menor cantidad de almejas en Playa Colombo que en Playa Fracasso (1.3 y 4.6 individuos por excremento, respectivamente). Sin embargo, éstas presentaron una talla mucho mayor (10.7 y 2.1 mm, respectivamente). Por ello, en Playa Colombo las almejas aportaron, en equivalente de biomasa por excremento, un valor 16 veces superior al reportado en Playa Fracasso (6.4 y 0.4 mg, respectivamente). La selección de tallas de Darina solenoides también fue diferente en Playa Colombo (4-12 mm) con respecto a Playa Fracasso (1-4 mm; D'Amico 2001). Esto puede deberse a la baja oferta de almejas de tallas menores a 10 mm en Playa Colombo. Teniendo en cuenta la escasa oferta de almejas de esta playa, así como su importancia en la dieta de las aves playeras en Península Valdés (D'Amico 2001, Musmeci 2005, Hernández 2007, Hernández et al. 2008), la ingesta de otros ítems presa se explicaría por la necesidad de estas aves de ampliar su espectro trófico a fin de compensar sus requerimientos energéticos.
Según D'Amico y Bala (2004), el aporte de Darina solenoides en Playa Fracasso es prácticamente despreciable si se lo compara con el de los poliquetos. Considerando el elevado porcentaje de ocurrencia de Travisia olens en Playa Colombo, estos poliquetos podrían estar jugando un rol importante en el aporte energético brindado a las aves, ya que prácticamente no produce desperdicios. Hernández (2007) encontró que en Península Valdés las almejas Darina solenoides son las presas que mayor aporte energético brindan (en Playa Colombo: 20.2 KJ/g, expresado en peso seco libre de cenizas), siendo menor el de Travisia olens (Playa Colombo: 10.9 KJ/g). Sin embargo, es importante destacar que en Playa Colombo hay sectores del intermareal en donde las densidades de Travisia olens son elevadas.
Algunos ítems presa como Tellina petitiana, los mitílidos, las semillas y los insectos podrían considerarse como remanentes de otros sitios donde las aves se estuvieron alimentando, aún cuando las heces se recolectaron después de observar a las bandadas más de 50 min. Los restos de mitílidos sugieren que las aves se habían estado alimentando en sustratos rocosos, mientras que la presencia de semillas e insectos indicaría que lo habían hecho en la sección superior del intermareal (en Playa Colombo o en otra localidad). El análisis de la oferta trófica indica que la almeja Tellina petitiana se encuentra ausente en Playa Colombo (Musmeci 2012, Musmeci et al. 2013), aunque se la encontró en otras playas de Península Valdés (Hernández 2007, Musmeci 2012, Musmeci et al. 2013). La ingesta de esta presa sugiere que los chorlitos estuvieron alimentándose en otro intermareal. Se ha reportado que los mismos individuos de Playero Rojizo utilizan distintas playas dentro de Península Valdés para alimentarse y que, en ocasiones, utilizan playas situadas en los golfos Nuevo y San José durante el mismo ciclo de marea (Musmeci et al. 2012). En función de la ingesta observada, es probable que el Chorlito Doble Collar tenga un comportamiento similar de utilización de más de un intermareal. Estudios futuros deberían incluir el seguimiento de individuos marcados con el fin de conocer los movimientos internos dentro de las diferentes playas de Península Valdés.
En estudios realizados en simultáneo con este trabajo (Musmeci 2005) se encontró que, además del Chorlito Doble Collar, otras tres especies de aves playeras hacían uso del intermareal: el Playero Rojizo, el Playerito Rabadilla Blanca (Calidris fuscicollis) y el Playerito Blanco (Calidris alba). En los días en que fue realizado el muestreo de este trabajo, las abundancias de estas aves fueron muy pequeñas, no superando los 400 individuos de cada una (Musmeci 2005). Los estudios de dieta de las otras especies presentes en Playa Colombo indican que existe una superposición trófica, aunque poseen diferentes longitudes de pico y estrategias de alimentación, por lo que estarían seleccionando distintos rangos de tallas de almejas (Musmeci 2005, Hernández 2007).
AGRADECIMIENTOS
Se agradece al Centro Nacional Patagónico y a la Universidad Nacional de la Patagonia San Juan Bosco. El trabajo fue realizado con financiamiento otorgado por la Fundación Patagonia Natural.
BIBLIOGRAFÍA CITADA
1. BALA LO, D'AMICO VL Y STOYANOFF P (2002) Migrating shorebirds at Peninsula Valdes, Argentina: report for the year 2000. Wader Study Group Bulletin 98:6-9. [ Links ]
2. BALA LO, HERNÁNDEZ MA Y D'AMICO VL (2001) Shorebirds present on Fracasso Beach (San José Gulf, Valdés Peninsula, Argentina): report of the 1999's migrating season. Wader Study Group Bulletin 94:27-30. [ Links ]
3. BARRETT RT, CAMPHUYSEN CJ, ANKER NILSSEN T, CHARDINE JW, FURNESS RW, GARTHE S, HUPPOP O, LEOPOLD MF, MONTEVECCHI WA Y VEIT RR (2007) Diet studies of seabirds: a review and recommendations. Journal of Marine Science 64:1675-1691. [ Links ]
4. BLANCO D, YORIO P, PETRACCI P Y PUGNALI G (2006) Distribution and abundance of non-breeding shorebirds along the coasts of the Buenos Aires Province, Argentina. Waterbirds 29:381-390. [ Links ]
5. CLARKE KR Y WARWICK RM (2001) Change in marine communities: an approach to statistical analysis and interpretation. Segunda edición. PRIMER-E, Plymouth. [ Links ]
6. D'AMICO VL (2001) Ecología trófica de Charadrius falklandicus en playas de Península Valdés. Tesis de licenciatura, Universidad Nacional de la Patagonia San Juan Bosco, Puerto Madryn. [ Links ]
7. D'AMICO VL Y BALA LO (2004) Prey selection and feeding behaviour of Two-banded Plover (Charadrius falklandicus) in Península Valdés, Patagonia, Argentina. Waterbirds 27:264-269. [ Links ]
8. D'AMICO VL, HERNÁNDEZ MA Y BALA LO (2004) Selección de presas en relación con las estrategias de forrajeo de aves migratorias en Península Valdés, Argentina. Ornitología Neotropical 15:357-364. [ Links ]
9. DEKINGA A Y PIERSMA T (1993) Reconstructing diet composition on the basis of faeces in a mollusceating wader, the Knot Calidris canutus. Bird Study 40:144-156. [ Links ]
10. GARCÍA-PEÑA GE, HERNÁNDEZ MA Y BALA LO (2008) Ecología reproductiva del Chorlo de Doble Collar (Charadrius falklandicus) en Península Valdés, Argentina. Ornitología Neotropical 19 (Suppl.):445-452. [ Links ]
11. GONZÁLEZ PM, PIERSMA T Y VERKUIL Y (1996) Food, feeding, and refueling of Red Knots during northward migration at San Antonio Oeste, Río Negro, Argentina. Journal of Field Ornithology 67:575-591. [ Links ]
12. HERNÁNDEZ MA (2007) Ecología trófica del Playero Rojizo (Calidris canutus rufa), Playero de Rabadilla Blanca (Calidris fuscicollis) y Playero Blanco (Calidris alba) en Península Valdés. Tesis doctoral, Universidad Nacional del Comahue, San Carlos de Bariloche. [ Links ]
13. HERNÁNDEZ MA, BALA LO Y MUSMECI LR (2008) Dieta de tres especies de aves playeras migratorias en Península Valdés, Patagonia argentina. Ornitología Neotropical 19 (Suppl.):605-611. [ Links ]
14. HERNÁNDEZ MA, D'AMICO VL Y BALA LO (2004) Presas consumidas por el Playero Rojizo (Calidris canutus) en Bahía San Julián, Santa Cruz, Argentina. Hornero 19:7-11. [ Links ]
15. MANLY B, MCDONALD L Y THOMAS D (1993) Resource selection by animals. Statistical design and analysis for field studies. Chapman and Hall,Londres. [ Links ]
16. MUSMECI LR (2005) Evaluación de playa Colombo (Península Valdés, Chubut) por la utilización de las aves playeras migratorias. Tesis de licenciatura, Universidad Nacional de la Patagonia San Juan Bosco, Puerto Madryn. [ Links ]
17. MUSMECI LR (2012) Evaluación de humedales costeros de Península Valdés (Chubut) utilizados como sitios de parada por los Playeros Rojizos (Calidris canutus rufa).Tesis doctoral, Universidad Nacional del Comahue, San Carlos de Bariloche. [ Links ]
18. MUSMECI LR, HERNÁNDEZ MA, BALA LO Y SCOLARO JA (2012) Use of Peninsula Valdes (Patagonia Argentina) by migrating Red Knots (Calidris canutus rufa). Emu 112:357-362. [ Links ]
19. MUSMECI L, HERNÁNDEZ M, SCOLARO J Y BALA L (2013) Almejas en golfos norpatagónicos, Argentina: disponibilidad trófica para aves playeras migratorias. Revista de Biología Marina y Oceanografía 48:219-225. [ Links ]
20. PAGNONI GO (1997) Poblamiento de la infauna en la zona intermareal del Golfo San José (Provincia del Chubut) y su importancia en la alimentación de aves migratorias. Tesis doctoral, Universidad Nacional de La Plata, La Plata. [ Links ]
21. PETRACCI P (2002) Diet of Sanderling in Buenos Aires Province, Argentina. Waterbirds 25:366-370. [ Links ]
22. PIERSMA T (1994) Close to the edge: energetic bottlenecks and the evolution of migratory pathways in knots. Tesis doctoral, Rijksuniversiteit Groningen, Groninga. [ Links ]
23. VAN GILS JA, DEKINGA A, SPAANS B, VAHL WK Y PIERSMA T (2005) Digestive bottleneck affects foraging decisions in Red Knots Calidris canutus. II. Patch choice and length of working day. Jounal of Animal Ecology 74:120-130. [ Links ]

