










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEl hornero
versión impresa ISSN 0073-3407versión On-line ISSN 1850-4884
Hornero vol.30 no.2 Ciudad Autónoma de Buenos Aires dic. 2015
ARTÍCULO
Cambios en la distribución altitudinal de las aves de Río Macho, Cartago, Costa Rica
Oscar Ramírez-Alán 1,4, Roberto Vargas-Masís 1,2 y Roberto A. Cordero 3
1 Escuela de Ciencias Biológicas, Universidad Nacional. Aptdo 86-3000, Heredia, Costa Rica.
2 Vicerrectoría de Investigación, Universidad Estatal a Distancia. San José, Costa Rica.
3 Laboratorio de Ecología Funcional y Ecosistemas Tropicales (LEFET), Escuela de Ciencias Biológicas, Universidad Nacional. Heredia, Costa Rica.
4 osoramirez@gmail.com
RESUMEN
Las aves poseen un alto potencial como indicadoras de los efectos del cambio climático debido a su facilidad de observación y de monitoreo continuo. Estudios previos acerca del efecto del cambio climático sobre las aves han mostrado que existen variaciones en la distribución de las especies. El objetivo de este estudio es evaluar el cambio en la distribución altitudinal de la avifauna de la Estación Biológica Río Macho y sus alrededores, en Costa Rica. Utilizando puntos de conteo, transectas, redes de niebla, grabaciones de sonido y observaciones, se registró la avifauna del área y se compararon los límites de altitud máxima previamente reportados para las especies con las altitudes registradas en este trabajo. Se registraron 197 especies, de las cuales el 14.8% fueron encontradas a altitudes mayores que el límite máximo reportado previamente. Las especies de zonas altas con mayor sensibilidad ecológica poseen un mayor riesgo de extinción, ya que no tienen la posibilidad de desplazarse a zonas más altas. Por lo tanto, se deben desarrollar estrategias para mantener poblaciones viables de especies endémicas de zonas altas.
PALABRAS CLAVE: Cambio climático; Costa Rica; Distribución altitudinal.
ABSTRACT
Changes in the altitudinal distribution of birds in Rio Macho, Cartago, Costa Rica.
Birds have a high potential as indicators of the effects of climate change, because they are easy to observe and monitoring. Previous studies on the effect of climate change on birds showed that there are significant variations in the distribution of species. Our objective was to evaluate changes in the elevational distribution of the avifauna in Río Macho, Cartago, Costa Rica. We used point counts, transects, mist nets, acoustic recordings and observations in order to record the local avifauna. The maximum elevation previously reported for these species was compared with elevations recorded during this study. We recorded 197 species, and 14.8% of them were observed at a higher elevation than the maximum previously reported. Species inhabiting higher elevations have a greater extinction risk because they cannot move to upper areas. Therefore, we must advance strategies in order to maintain viable populations for endemic species in higher elevations.
KEY WORDS: Altitudinal distribution; Climate change; Costa Rica.
Recibido 8 julio 2015
Aceptado 28 diciembre 2015
Las aves son afectadas por los cambios en la estructura de los bosques producidos por la tala, la fragmentación, los incendios y los cambios en el uso del suelo (Wiens 1995, Root et al. 2003, Crick 2004), que provocan que los sitios originales de reproducción desaparezcan o que se vuelvan vulnerables para la permanencia de sus poblaciones. En consecuencia, las aves deben modificar sus rangos altitudinales en respuesta a estos cambios (Sala et al. 2000, Lang et al. 2003). A ello se suma que la diversidad de todos los grupos de fauna es afectada por la deforestación, los cambios en el uso de suelo, la aparición de enfermedades, la sobreexplotación de recursos, la contaminación y otras actividades humanas (Lawton et al. 2001, Ray et al. 2006, Feeley y Silman 2010, Fuller et al. 2012, Sekercioglu et al. 2012).
Las aves poseen un alto potencial como indicadoras de los efectos del cambio climático sobre la biota (Tankersley y Orvis 2003, Buermann et al. 2011) debido a su facilidad de observación y de monitoreo continuo, en especial cuando se establecen investigaciones a largo plazo de su distribución. La avifauna de los bosques montanos es considerada la más vulnerable a las variaciones climáticas a pequeña y gran escala (Tankersley y Orvis 2003, Sekercioglu et al. 2008, Buermann et al. 2011, Foden et al. 2013), las cuales afectan también a otros organismos (Both et al. 2006). Estudios previos acerca del efecto del cambio climático sobre las aves han mostrado que existen variaciones en la distribución de las especies (Both et al. 2006, Hitch y Leberg 2007). Estos cambios ponen en riesgo su viabilidad y permanencia, en especial en ecosistemas boscosos (Still et al. 1999, Peh 2007, Sekercioglu et al. 2008, Marini et al. 2009). Algunas aves han modificado el inicio del periodo de reproducción (Svensson y Nilsson, 1995, Bêty et al. 2004), han disminuido su tamaño poblacional, han adelantado el periodo de migración (Gunnarsson et al. 2006) o han modificado las distancias de migración y su distribución (Feria-Arroyo et al. 2013). El propósito de este estudio es evaluar el cambio en la distribución altitudinal de la avifauna de la Estación Biológica Río Macho y sus alrededores, en Costa Rica, durante 2009-2011, como un indicador del fenómeno de cambio climático.
MÉTODOS
El estudio se realizó en 2009-2011 en dos ambientes ubicados al sur del valle de Orosí, Cartago, Costa Rica (Fig. 1). El primer ambiente es un bosque de aproximadamente 50 años de regeneración natural ubicado en las cercanías de la Estación de Biología Tropical y Acuicultura Río Macho (9°45'N, 84°51'O; 1900 msnm) expuesto a actividad ganadera durante la década de 1950. El sitio presenta una precipitación anual de 2416 mm (Riba- Hernández 1996) y una temperatura mensual promedio de 15.8 °C (Pérez-Molina y Cordero Solórzano 2012). El segundo ambiente es un bosque en regeneración ("tacotal") con unos 15 años de abandono que antiguamente fue una plantación comercial de manzana rosa (Syzigium jambos), ubicado 1 km al sureste del embalse El Llano (9°45'N, 83°50'O; 1650 msnm). Su temperatura mensual promedio es de 16.5 °C. Según Holdridge (1967) ambos ambientes corresponden a un Bosque Muy Húmedo Montano Bajo. La Estación de Biología Tropical y Acuicultura Río Macho forma parte de la Reserva Forestal Río Macho, la cual en la Cordillera de Talamanca forman el mayor corredor biológico altitudinal de Costa Rica. Ambas contribuyen con el mayor número de ecosistemas, alta diversidad de flora y fauna en peligro de extinción, así como un alto endemismo (TNC-INBio 2006). Para la caracterización de los pisos altitudinales se utilizó la información del Atlas de Costa Rica (Ortiz- Malavassi 2014) y la información contenida en las capas de ecosistemas (Kapelle et al. 2002).
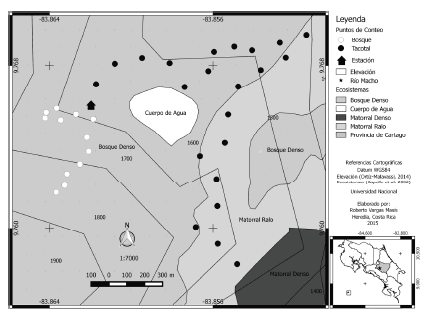
Figura 1. Ubicación de los sitios de muestreo en los ambientes de bosque y tacotal de la Estación Biológica Río Macho y sus alrededores (Cartago, Costa Rica).
El muestreo de las aves se realizó usando una combinación de puntos de conteo y transectas, con puntos ubicados cada 150 m en el tacotal y cada 50 m en el bosque. Se identificaron y contaron todas las aves observadas o escuchadas dentro de un área circular de 50 m de radio durante 6 min en cada punto. Los muestreos se realizaron de 5:00-11:00 h. Para la identificación de las aves se utilizaron las guías de Garrigues y Dean (2007) y Stiles y Skutch (2007). Se utilizaron grabaciones de sonido para las especies que no fue posible identificar de manera visual. Para complementar la descripción de la avifauna se realizaron muestreos mensuales con redes de niebla. Se usaron ocho redes (12×2.6 m, 38 mm de malla) que fueron colocadas cada 50-100 m. Las redes fueron revisadas cada 20 min. Todas las aves fueron identificadas con anillos numerados. Además, se realizaron observaciones fuera de los muestreos para complementar el relevamiento de la riqueza de especies. Para la clasificación taxonómica de la especies se utilizó la lista oficial de las aves de Costa Rica de Obando-Calderón et al. (2013) y se consignó el estatus de residente o migratoria (todas las especies migratorias en el área de estudio son neotropicales). A cada especie de ave se le asignaron los límites histórico y actual de altitud máxima utilizando las guías de Garrigues y Dean (2007) y Stiles y Skutch (2007), la plataforma de información de BirdLife International (BirdLife International 2014) y las observaciones realizadas en este estudio en 2009-2011.
RESULTADOS
Se registró un total de 3871 individuos de aves en el área de estudio durante los tres años, pertenecientes a 197 especies y 40 familias. Se identificaron 165 especies residentes, 24 migratorias, 5 con poblaciones tanto residentes como migratorias (Cathartes aura, Buteo brachyurus, Buteo jamaicensis, Cypseloides niger y Pygochelidon cyanoleuca), 2 con poblaciones residentes reproductivas y migratorias (Legatus leucophaius y Myiodynastes luteiventris) y una con poblaciones residentes, residentes reproductivas y migratorias (Elanoides forficatus). Las 24 especies migratorias pertenecen a 9 familias, de las cuales Parulidae presentó la mayor riqueza (10 especies, entre ellas Vermivora chrysoptera, Setophaga virens y Mniotilta varia), seguida por Vireonidae (Vireo flavifrons, Vireo leucophrys y Vireo olivaceus) y Accipitridae (Accipiter striatus, Accipiter cooperii y Buteo platypterus) con 3 especies cada una. Se registraron 30 especies endémicas de las tierras altas entre Costa Rica y el oeste de Panamá, entre las que se destacan Chamaepetes unicolor, Megascops clarkii, Scytalopus argentifrons, Vireo carmioli y Zeledonia coronata. Solo una especie se puede considerar como endémica continental (Elvira cupreiceps).
Durante el estudio se evidenciaron cambios en la distribución altitudinal de las especies (Tabla 1). El 14.8% de las especies se encontraron a altitudes mayores que el límite superior reportado a nivel nacional por Garrigues y Dean (2007) y Stiles y Skutch (2007). El aumento promedio (±DE) de la altitud para estas especies con respecto a los datos de esos autores fue de 263.3±132.2 m. Las especies que mostraron mayores diferencias fueron Xiphorhynchus susurrans, Pteroglossus torquatus y Tolmomyias assimilis, que se encontraron a 745, 661 y 558 m, respectivamente, por encima de lo reportado previamente (Tabla 1). Si se comparan los nuevos registros con el límite superior indicado en la plataforma de BirdLife International (2014), el 5.6% de las especies mostraron altitudes mayores que las previamente reportadas (Tabla 1). El aumento promedio de la altitud para estas especies fue en este caso de 420.5±291.2 m. Las especies que mostraron mayores diferencias fueron Psarocolius montezuma, Electron platyrhynchum y Legatus leucophaius, que se encontraron a 966, 704 y 665 m, respectivamente, por encima de lo reportado previamente (Tabla 1).
Tabla 1. Límites de altitud máxima (msnm) reportados por Garrigues y Dean (2007) y Stiles y Skutch (2007) (GD-SS), BirdLife International (2014) (BI) y observados en 2009-2011 en ambientes de bosque y tacotal de la Estación Biológica Río Macho y sus alrededores (Cartago, Costa Rica). Para cada especie se indica, además, su estatus migratorio.

DISCUSIÓN
El área de la Estación Biológica Río Macho y sus alrededores posee una gran riqueza de especies residentes, las cuales mantienen poblaciones que dependen de las condiciones locales para sobrevivir. Además, están presentes un número importante de especies migratorias que viajan grandes distancias en búsqueda de mejores condiciones (Woodcock y Woodcock 2007) y para las cuales el área puede cumplir un papel importante en su conservación. El sitio alberga un número importante de aves endémicas (31 especies) que, comparado con zonas de mayor protección como los bosques de Monteverde (41 especies), lo sitúan como un área importante para la conservación de las aves.
Las observaciones realizadas evidencian cambios en la distribución altitudinal de las aves residentes que podrían inicialmente atribuirse al cambio climático de las últimas décadas, ya que éste repercute directamente sobre la fenología de los bosques (Opler et al. 1980, Loiselle y Blake 1991) y provoca que las aves no cuenten con los requerimientos ecológicos necesarios. Los cambios en la distribución altitudinal pueden estar relacionados a variaciones en la estructura del hábitat (Loiselle y Blake 1991, Ramírez-Alán 2008, Bulluck et al. 2013). El área de estudio ha sufrido variaciones en el uso del suelo en las últimas décadas, pasando de una recuperación parcial del bosque a una reforestación con especies exóticas (Pérez-Molina y Cordero Solórzano 2012). Los patrones de movimiento de las especies pueden variar debido a su alta capacidad de desplazamiento y a su adaptación a los cambios de los bosques, siendo favorecidos por las conexiones entre parches boscosos (Chaves- Campos 2004, Stiles 2008).
Los cambios en la distribución altitudinal pueden indicar que el área estudiada representa una zona de transición importante para ciertos grupos de aves, como por ejemplo para las poblaciones nómades de Formicarius rufipectus que fueron observadas durante ciertas épocas del año (enero, abril y julio), así como para las especies migratorias que hacen uso de los bosques de alturas intermedias, las cuales pueden ser afectadas debido a su susceptibilidad a los cambios en la estructura del bosque y a las interacciones con otras especies (Bulluck et al. 2013), como es el caso de Vermivora chrysoptera, Setophaga virens y Mniotilta varia.
Las alteraciones en la estructura y fenología del bosque en las áreas estudiadas afectan en mayor medida a las especies residentes o endémicas con ámbitos restringidos, las cuales suelen ser más sensibles a la extinción (Shoo et al. 2005, Colwell et al. 2008, Gasner et al. 2010, Foden et al. 2013). Los procesos como la competencia y la predación, así como las enfermedades, entre otros, aumentan en respuesta a la variación temporal de los recursos, y un aumento de la temperatura favorece potencialmente la transmisión de parásitos y virus (Møller et al. 2013).
El área de Río Macho tiene influencia de las montañas del Caribe y aproximadamente el 30% de las especies residentes reproductivas migran altitudinalmente (Stiles 1983). Cerca del 85% de estas especies se movilizan hacia las partes más altas de las montañas para reproducirse como una estrategia para evitar la predación, que disminuye con la altitud (Boyle 2008). Se requieren más datos y tiempo para entender mejor la dinámica de estos bosques, ya que se desconoce la disponibilidad de recursos para las especies migratorias altitudinales (Loiselle y Blake 1991, Solórzano et al. 2000, Chaves-Campos 2004, Boyle 2008). El desplazamiento altitudinal podría ocasionar que algunas especies no se adapten rápido a los cambios en la temperatura, lo que les impediría soportar los fuertes cambios climáticos que podrían ocurrir (Devictor et al. 2012). El desplazamiento de grupos faunísticos ya ha sido detectado en otras regiones (Martín et al. 2012) y no se considera un evento reciente. En Costa Rica ya se han reportado registros accidentales de aves que no corresponden a su distribución original en el trópico centroamericano (Obando-Calderón et al. 2013). El cambio climático es un aspecto clave para la conservación de los bosques tropicales en Costa Rica, por lo cual deben adoptarse compromisos internacionales y medidas bilaterales para el mantenimiento y aumento de la conectividad entre los remanentes de bosque en las matrices actuales de paisaje. Las estrategias incluyen mantener un monitoreo de las aves, generar conectividad entre los ecosistemas y restaurar zonas forestales (Sayre et al. 2002).
Este es el primer estudio que intenta documentar los cambios en la distribución altitudinal de las aves de la Estación Biológica Río Macho. Las especies de zonas altas con mayor sensibilidad ecológica poseen un mayor riesgo de extinción, ya que no tienen la posibilidad de desplazarse a zonas más altas, como es el caso de las especies endémicas. Los modelos propuestos por Gasner et al. (2010) proyectan una declinación de hasta el 50% de las especies en respuesta a un cambio climático. Por lo tanto, se deben desarrollar estrategias para mantener poblaciones viables de especies endémicas de zonas altas. Por otra parte, es necesario investigar más sobre los recursos que utilizan estas especies, así como las interacciones. Para entender la dinámica de las poblaciones es importante mantener registros históricos de los cambios en su distribución con el fin de analizar sus posibles causas. En consecuencia, se recomienda continuar con la observación de estas aves en la zona, en donde se están registrando cambios en la distribución de las especies, con el fin de plantear estrategias de conservación a corto, mediano y largo plazo.
AGRADECIMIENTOS
Agradecemos a los estudiantes que colaboraron en el proyecto de la Escuela de Ciencias Biológicas. Este trabajo fue financiado con fondos de proyectos FEES-CONARE, específicamente el proyecto "Análisis ecosistémico para la evaluación de la restauración forestal y sus implicaciones en el secuestro de carbono en un bosque nublado". Además, agradecemos a los revisores anónimos por las sugerencias finales.
BIBLIOGRAFÍA CITADA
1. BÊTY J, GIROUX JF Y GAUTHIER G (2004) Individual variation in timing of migration: causes and reproductive consequences in greater snow geese (Anser caerulescens atlanticus). Behavioral Ecology and Sociobiology 57:1-8 [ Links ]
2. BIRDLIFE INTERNATIONAL (2014) Data zone. BirdLife International, Cambridge (URL: http://datazone.birdlife.org/home) [ Links ]
3. BOTH C, BOUWHUIS S, LESSELLS CM Y VISSER ME (2006) Climate change and population declines in a longdistance migratory bird. Nature 441:81-83 [ Links ]
4. BOYLE WA (2008) Can variation in risk of nest predation explain altitudinal migration in tropical birds? Oecologia 155:397-403 [ Links ]
5. BUERMANN W, CHAVES JA, DUDLEY R, MCGUIRE JA, SMITH TB Y ALTSHULER DL (2011) Projected changes in elevational distribution and flight performance of montane Neotropical hummingbirds in response to climate change. Global Change Biology 17:1671-1680 [ Links ]
6. BULLUCK L, BUEHLER D, VALLENDER R Y ROBERTSON JR (2013) Demographic comparison of Golden-Winged Warbler (Vermivora chrysoptera) populations in northern and southern extremes of their breeding range. Wilson Journal of Ornithology 125:479-490 [ Links ]
7. CHAVES-CAMPOS J (2004) Elevational movements of large frugivorous birds and temporal variation in abundance of fruits along an elevational gradient. Ornitología Neotropical 15:433-445 [ Links ]
8. COLWELL RK, BREHM G, CARDELUS CL, GILMAN AC Y LONGINO JT (2008) Global warming, elevational range shifts, and lowland biotic attrition in the wet tropics. Science 322:258-261 [ Links ]
9. CRICK HPQ (2004) The impact of climate change on birds. Ibis 146:48-56 [ Links ]
10. DEVICTOR V, VAN SWAAY C, BRERETON T, BROTONS L, CHAMBERLAIN D, HELIÖLä J, HERRANDO S, JULLIARD R, KUUSSAARI M, LINDSTRÖM Å, REIF J, ROY DB, SCHWEIGER O, SETTELE J, STEFANESCU C, VAN STRIEN A, VAN TURNHOUT C, VERMOUZEK Z, WALLIS DE VRIES M, WYNHOFF I Y JIGUET F (2012) Differences in the climatic debts of birds and butterflies at a continental scale. Nature Climate Change 2:121-124 [ Links ]
11. FEELEY K Y SILMAN M (2010) Land-use and climate change effects on population size and extinction risk of Andean plants. Global Change Biology 16:3215-3222 [ Links ]
12. FERIA-ARROYO TP, SÁNCHEZ-ROJAS G, ORTIZ-PULIDO R, BRAVO-CADENA J, CALIXTO-PÉREZ E, DALE JM, DUBERSTEIN JN, LLOLDI-RANGEL P, LARA C Y VALENCIA- HERVERTH J (2013) Estudio del cambio climático y su efecto en las aves en México: enfoques actuales y perspectivas futuras. Huitzil 14:47-55 [ Links ]
13. FODEN WB, BUTCHART SHM, STUART SN, VIE JC, AKÇAKAYA HR, ANGULO A, LYNDON D, GUTSCHE A, TURAK E, CAO L, DONNER S, KATARIYA V, BERNARD R Y MACE G (2013) Identifying the World's most climate change vulnerable species: a systematic trait-based assessment of all birds, amphibians and corals. PLoS One 8:e65427 [ Links ]
14. FULLER T, BENSCH S, MüLLER I, NOVEMBRE J, PÉREZ-TRIS J, RICKLEFS RE, SMITH TB Y WALDENSTRÖM J (2012) The ecology of emerging infectious diseases in migratory birds: an assessment of the role of climate change and priorities for future research. Ecohealth 9:80-88 [ Links ]
15. GARRIGUES R Y DEAN R (2007) The birds of Costa Rica: a field guide. Comstock, Ithaca [ Links ]
16. GASNER MR, JANKOWSKI JE, CIECKA AL, KYLE KO Y RABENOLD KN (2010) Projecting the local impacts of climate change on a Central American montane avian community. Biological Conservation 143:1250-1258 [ Links ]
17. GUNNARSSON TG, GILL JA, ATKINSON PW, GÉLINAUD G, POTTS PM, CROGER RE, GUDMUNDSSON GA, APPLETON GF Y SUTHERLAND WJ (2006) Population-scale drivers of individual arrival times in migratory birds. Journal of Animal Ecology 75:1119-1127 [ Links ]
18. HITCH AT Y LEBERG PL (2007) Breeding distributions of North American bird species moving north as a result of climate change. Conservation Biology 21:534-539 [ Links ]
19. HOLDRIDGE LR (1967) Life zone ecology. Edición revisada. Tropical Science Center, San José [ Links ]
20. KAPELLE M, CASTRO M, ACEVEDO H, GONZÁLEZ L Y MONGE H (2002) Ecosistemas del Área de Conservación Osa (Acosa). Instituto Nacional de Biodiversidad, Heredia [ Links ]
21. LANG I, GORMLEY LHL, HARVEY CA Y SINCLAIR FL (2003) Composición de la comunidad de aves en cercas vivas de Río Frío, Costa Rica. Agroforestería en las Américas 10:39-40 [ Links ]
22. LAWTON RO, NAIR US, PIELKE RA Y WELCH RM (2001) Climatic impact of tropical lowland deforestation on nearby montane cloud forests. Science 294:584-587 [ Links ]
23. LOISELLE BA Y BLAKE JG (1991) Temporal variation in birds and fruits along an elevational gradient in Costa Rica. Ecology 72:180-193 [ Links ]
24. MARINI MA, BARBET-MASSIN M, ESTEVES-LÓPEZ L Y JIGUET F (2009) Predicted climate-driven bird distribution changes and forecasted conservation conflicts in a Neotropical savanna. Conservation Biology 23:1558-1567 [ Links ]
25. MARTÍN JL, BETHENCOURT J Y CUEVAS-AGULLÓ E (2012) Assessment of global warming on the island of Tenerife, Canary Islands (Spain). Trends in minimum, maximum and mean temperatures since 1944. Climatic Change 114:343-355 [ Links ]
26. MØLLER AP, MERINO S, SOLER JJ, ANTONOV A, BADÁS EP, CALERO-TORRALBO MA, DE LOPE F, EEVA T, FIGUEROLA J, FLENSTED-JENSEN E, GARAMSZEGI LZ, GONZÁLEZ-BRAOJOS S, GWINNER H, HANSSEN SA, HEYLEN D, ILMONEN P, KLARBORG K, KORPIMäKI E, MARTÍNEZ J, MARTÍNEZ-DE LA PUENTE J, MARZAL A, MATTHYSEN E, MATYJASIAK, P, MOLINA-MORALES M, MORENO J, MOUSSEAU TA, NIELSEN JT, PAP PL, RIVERODE AGUILAR J, SHURULINKOV P, SLAGSVOLD T, SZÉP T, SZÖLLÓSI E, TÖRÖK J, VACLAV R, VALERA F Y ZIANE N (2013) Assessing the effects of climate on host-parasite interactions: a comparative study of European birds and their parasites. PLoS One 8:e82886 [ Links ]
27. OBANDO-CALDERÓN G, CHAVES-CAMPOS J, GARRIGUES R, MONTOYA M, RAMÍREZ O Y ZOOK J (2013) Lista oficial de las aves de Costa Rica. Actualización 2013. Zeledonia 17:44-59 [ Links ]
28. OPLER PA, FRANKIE GW Y BAKER HG (1980) Comparative phenological studies of treelet and shrub species in tropical wet and dry forests in the lowlands of Costa Rica. Journal of Ecology 167-188 [ Links ]
29. ORTIZ-MALAVASSI E (2014) Atlas digital de Costa Rica 2014. Escuela de Ingeniería Forestal, Instituto Tecnológico de Costa Rica, Cartago [ Links ]
30. PEH KSH (2007) Potential effects of climate change on elevational distributions of tropical birds in southeast Asia. Condor 109:437-441 [ Links ]
31. PÉREZ-MOLINA JP Y CORDERO SOLÓRZANO RA (2012) Recuperación de tres coberturas forestales de altura media en Costa Rica: análisis de los oligoquetos, el mantillo y suelo. Revista de Biología Tropical 60:1431-1443 [ Links ]
32. RAMÍREZ-ALÁN OA (2008) Uso de hábitat, abundancia relativa del quetzal (Pharomachrus mocinno costaricensis) y análisis de paisaje en el sector Volcán Barva, Parque Nacional Braulio Carrillo, Costa Rica. Mesoamericana 12:21-32 [ Links ]
33. RAY DK, NAIR US, LAWTON RO, WELCH RM Y PIELKE RA (2006) Impact of land use on Costa Rican tropical montane cloud forests: sensitivity of orographic cloud formation to deforestation in the plains. Journal of Geophysical Research. Atmospheres 111:1984-2012 [ Links ]
34. RIBA-HERNÁNDEZ P (1996) Notas sobre los murciélagos de la Estación Biológica Río Macho, Costa Rica. Brenesia 45:183-186 [ Links ]
35. ROOT TL, PRICE JT, HALL KR, SCHNEIDER SH, ROSENZWEIGK C Y POUNDS JA (2003) Fingerprints of global warming on wild animals and plants. Nature 421:57-60 [ Links ]
36. SALA OE, CHAPIN III FS, ARMESTO JJ, BERLOW R, BLOOMFIELD J, DIRZO R, HUBER-SANWALD E, HUENNEKE LF, JACKSON RB, KINZIG A, LEEMANS R, LODGE D, MOONEY HA, OESTERHELD M, POFF ML, SYKES MT, WALKER BH, WALKER M Y WALL DH (2000) Global biodiversity scenarios for the year 2100. Science 287:1770-1774 [ Links ]
37. SAYRE R, ROCA E, SIDAGHATKISSH G, YOUNG B, KEEL S, ROCA R Y SHEPPARD S (2002) Un enfoque en la naturaleza. Evaluaciones ecológicas rápidas. The Nature Conservancy, Arlington [ Links ]
38. SEKERCIOGLU ÇH, PRIMACK RB Y WORMWORTH J (2012) The effects of climate change on tropical birds. Biological Conservation 148:1-18 [ Links ]
39. SEKERCIOGLU ÇH, SCHNEIDER SH, FAY JP Y LOARIE SR (2008) Climate change, elevational range shifts and bird extinctions. Conservation Biology 22:140-150 [ Links ]
40. SHOO LP, WILLIAMS SE Y HERO JM (2005) Climate warming and the rainforest birds of the Australian wet tropics: using abundance data as a sensitive predictor of change in total population size. Biological Conservation 125:335-343 [ Links ]
41. SOLÓRZANO S, CASTILLO S, VALVERDE T Y ÍVILA L (2000) Quetzal abundance in relation to fruit availability in a cloud forest in southeastern Mexico. Biotropica 32:523-532 [ Links ]
42. STILES FG (1983) Birds. Pp. 502-530 en: JANZEN DH (ed) Costa Rican natural history. University of Chicago Press, Chicago [ Links ]
43. STILES FG (2008) Ecomorphology and phylogeny of hummingbirds: divergence and convergence in adaptations to high elevations. Ornitología Neotropical 19:511-519 [ Links ]
44. STILES FG Y SKUTCH AF (2007) Guía de aves de Costa Rica. Cuarta edición. Instituto Nacional de Biodiversidad, Heredia [ Links ]
45. STILL CJ, FOSTER PN Y SCHNEIDER SH (1999) Simulating the effects of climate change on tropical montane cloud forests. Nature 398:608-610 [ Links ]
46. SVENSSON E Y NILSSON JA (1995) Food-supply, territory quality and reproductive timing in the blue tit (Parus caeruleus). Ecology 76:1804-1812 [ Links ]
47. TANKERSLEY R JR Y ORVIS K (2003) Modeling the geography of migratory pathways and stopover habitats for neotropical migratory birds. Conservation Ecology 7:e7 [ Links ]
48. TNC-INBIO (2006) Revisión de los objetos de conservación y análisis de integridad ecológica del sitio Tapantí- Macizo de la Muerte, Costa Rica. Instituto Nacional de Biodiversidad, San José [ Links ]
49. WIENS JA (1995) Habitat fragmentation: island vs. landscape perspectives on bird conservation. Ibis 137 (Suppl):97-104 [ Links ]
50. WOODCOCK J Y WOODCOCK M (2007) Diversidad de especies, fidelidad al sitio de migración, y ecología de aves migratorias terrestres en los manglares de Costa Rica. Zeledonia 11:2-13 [ Links ]

