










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkActa bioquímica clínica latinoamericana
versión impresa ISSN 0325-2957versión On-line ISSN 1851-6114
Acta bioquím. clín. latinoam. v.40 n.1 La Plata ene./mar. 2006
INMUNOLOGÍA
La respuesta inmune innata entre los factores ambientales y el aborto. Modelo de tolerancia al lipopolisacárido durante la gestación
Valeria Dubinsky1*, Gisela Junovich1*, Gabriela Gutiérrez2*
1. Licenciada en Ciencias Biológicas de la FCEyN de la UBA
2. Doctora en Ciencias de la UBA, área Inmunología de la Reproducción
* Instituto de Estudios de la Inmunidad Humoral (IDEHU)/Cátedra de Inmunología, FFyB, Universidad de Buenos Aires, Junín 956 4P (1113), Buenos Aires, Argentina.
Resumen
El aborto inmunológico del modelo animal CBA/j x DBA/2 es dependiente de factores ambientales tipo liposacárido (LPS) y de una respuesta inflamatoria local exacerbada. Al considerar que ratones deficientes en receptores Tlr4 de la respuesta inmune innata específicos para LPS son resistentes al aborto inducido por la endotoxina, se ha estudiado el efecto de la tolerización con LPS en la prevención del aborto y en la producción de citoquinas placentarias del modelo CBA/j x DBA/2. Para ello se inocularon bajas dosis de LPS en etapas pre y post-implantatorias, y con el fin de analizar el efecto tolerizante, se cultivaron placentas y esplenocitos en presencia de una dosis abortogénica de LPS. Los resultados obtenidos muestran que la inoculación post-implantatoria de 1 µg/día de LPS disminuyó el índice de aborto, mientras que indujo una baja respuesta al desafío con altas dosis de LPS en la síntesis de IL-6, tanto en bazo como en placenta. Sin embargo, el mismo desafío incrementó los niveles de TNF-a en placenta, aunque los disminuyó en bazo. A partir de estos resultados se puede concluir que la inducción de tolerancia al LPS es dependiente del órgano y de la citoquina estudiada.
Palabras clave: aborto * respuesta immune innata * lipopolisacárido * tolerización * citoquinas
Summary
Innate Immune Response between environmental factors and abortion: lipopolysaccharide tolerization model during gestation
The exacerbated local inflammatory response that triggers immunological abortion on CBA/j x DBA/2 murine model depends on environmental factors such as LPS. Since deficient Tlr4 mice are resistant to LPS-induced abortion, the effect of LPS tolerization on the prevention of fetal losses, as well as on the synthesis of placental cytokines on CBA/j x DBA/2 model has been studied. Low doses of LPS were inoculated during pre and post-implantatory stages on CBA/j pregnant mice. In order to investigate tolerization effect, abortogenic doses of LPS were added to placental and splenocyte cultures.
Results obtained with post-implantatory inoculation of 1µg/day LPS showed a decrease in the abortion rate as well as an IL-6 hyporesponsive to LPS booster, both in placenta and splenocyte cultures. However, stimulation with abortogenic dose of LPS modified TNF-a levels by an increase in placenta but a decrease in splenocyte culture supernatants. Considering these results, it can be concluded that LPS tolerance induction depends on the organ as well as on the cytokine studied.
Key words: abortion * innate immune response * lipopolysaccharide * tolerization * cytokines
INTRODUCCIÓN
El aborto espontáneo es la más frecuente de las complicaciones de la gestación en la mujer. La mayoría de las pacientes abortadoras recurrentes presentan problemas inmunológicos: desbalance entre citoquinas inflamatorias (Th1) y antinflamatorias (Th2), presencia de autoanticuerpos, elevada acción de células NK, entre otros. Sin embargo, hasta ahora no se ha podido establecer una relación directa entre el sistema inmune innato y el aborto. Recientes estudios demuestran que el aborto inmunológico provocado por una respuesta inflamatoria exacerbada, es dependiente de factores ambientales del tipo LPS (lipopolisacárido bacteriano) (1). El LPS es un componente de la membrana externa de bacterias Gram negativas que al unirse a receptores tipo Toll, induce una respuesta inflamatoria severa mediante la iniciación de señales intracelulares múltiples, que incluyen la activación del factor nuclear kb. Esta cascada de señalización lleva a la síntesis de citoquinas Th1, incrementando la inflamación que conduce al daño celular, lo cual se traduce en un aumento del nivel de pérdidas fetales (1) (2). Los receptores tipo Toll (Tlr), entre los que se encuentra el Tlr4 específico para el LPS, son componentes esenciales del sistema immune innato que reconocen secuencias conservadas sobre la superficie de microorganismos. El desbalance de esta función inmune innata puede contribuir a ciertas complicaciones durante la preñez (3). Diferentes estudios han demostrado que la administración de endotoxinas bacterianas a ratones preñados induce la resorción (aborto) (4)(5). Ratones deficientes en Tlr4 (C3H/Hej) son resistentes al aborto inducido por LPS (1). Cambios en la expresión de Tlr4 alteran la respuesta celular al LPS, mientras que la tolerancia al LPS ha sido claramente asociada con una disminución de la expresión de Tlr4 sobre la superficie celular (6). Estudios previos han demostrado que el tratamiento in vivo con bajas dosis de LPS (no abortogénicas y subletales) es capaz de reducir la respuesta de los macrófagos a una segunda estimulación con esta molécula, y que este mecanismo es mediado por la inactivación de la vía NFkb que se produce por la disminución de la expresión del receptor Tlr4 en la superficie de estas células (6). Sin embargo, la inducción de tolerancia al LPS durante la gestación, aún no ha sido estudiada.
Para el estudio de los aspectos inmunológicos implicados en la reproducción, ha sido particularmente útil el uso de un modelo animal murino de aborto espontáneo, debido a la disponibilidad de cepas genéticamente definidas, al corto tiempo de gestación y al bajo costo de las investigaciones. La cruza murina alogeneica CBA/j x DBA/2 muestra una inapropiada expresión de citoquinas inflamatorias en la interfase materno-fetal que correlaciona con el alto índice de muerte fetal (7). Se ha demostrado que las unidades fetoplacentarias de la cruza abortadora CBA/j x DBA/2 secretan niveles menores de IL-6 con respecto a la cruza control no abortadora CBA/j x BALB/c. Más aún, la inoculación de esta citoquina en el período post-implantación fue capaz de regular los niveles fetoplacentarios de IL-6, mientras que el índice de resorción disminuyó casi un 50%(8).
A partir de estos antecedentes, se decidió investigar el efecto de la inducción de tolerancia al LPS durante la gestación del modelo abortador y su influencia sobre el índice de pérdidas fetales y los niveles de citoquinas relacionadas con el aborto recurrente.
MATERIALES Y MÉTODOS
Animales
Ratones hembras de la cepa CBA/j de alrededor de cuatro meses de edad fueron provistos por el bioterio de la Comisión Nacional de Energía Atómica. Ratones machos de la cepa DBA/2 en edad reproductiva fueron provistos por el bioterio del IDEHU, FFyB, UBA. Se realizaron cruzas alogeneicas monogámicas de hembras CBA/j con machos DBA/2 (CBA/j x DBA/2) que presentan alta tasa de resorción fetal (7).
Detección de la preñez
La preñez fue determinada por la observación de tapón vaginal, considerando como día 0,5 al de su aparición. A partir de este momento, las hembras fueron separadas en jaulas individuales.
Determinación del índice de pérdidas fetales (IR)
El IR fue determinado mediante el recuento de fetos y placentas de morfología normal y de resorciones (abortos). Para ello, hembras primíparas fueron sacrificadas el día 13,5 de la gestación, mediante dislocación cervical. Los porcentajes de preñez y resorción fueron calculados mediante la siguiente fórmula: IR=100 x (Re/Re+F) (F=fetos y placentas de morfología y tamaño normales y Re=fetos y placentas indistinguibles, inserciones de menor tamaño). Luego se procedió a la extracción de placentas y bazos para su posterior cultivo.
Sobrenadantes de cultivos de placenta
Las placentas obtenidas por disección abdominal fueron separadas de la decidua, fraccionadas y cultivadas individualmente en medio Roswell Park Memorial Institute (RPMI 1640, GIBCO, Carlsbad, California, EEUU) adicionado con 10% de suero fetal bovino (SFB, GIBCO) y con 1% de penicilina/estreptomicina (GIBCO). Luego de 48 h de cultivo a 37 °C en atmósfera húmeda controlada al 5% de CO2, los SCP fueron recolectados, centrifugados a 5000 rpm durante 5 min, luego a 10.000 rpm durante 10 min y conservados a –70 ºC para su posterior análisis.
Sobrenadante del cultivo de esplenocitos
La obtención de la suspensión celular de esplenocitos se realizó en condiciones de esterilidad y mediante el vaciado mecánico del bazo con el uso de tijeras. Luego de efectuar lavados con medio RPMI, la suspensión obtenida fue centrifugada a 3000 rpm por 10 min. El recuento celular y la viabilidad se determinaron por coloración de exclusión con Azul Tripán (0,2% P/V en H2O destilada). Los esplenocitos fueron cultivados en una concentración de un millón de células/mL en medio RPMI adicionado con 10% de suero fetal bovino (SFB, GIBCO) y con 1% de penicilina/estreptomicina (GIBCO) durante 48 h a 37 °C en atmósfera húmeda controlada al 5% de CO2. Los sobrenadantes del cultivo fueron recolectados, centrifugados a 5000 rpm durante 5 min, luego a 10.000 rpm durante 10 min y conservados a –70 ºC para su posterior análisis.
Determinación de los niveles de citoquinas
La cuantificación de los niveles de citoquinas (IL-6, TNF-a) fue realizada utilizando ensayos enzimáticos de captura en fase sólida (ELISA) tipo sandwich, según el método de Butler y cols (9), utilizando reactivos comerciales (R&D, Minneápolis, MN, EEUU). Los anticuerpos utilizados fueron los siguientes, anticuerpo monoclonal de captura anti IL-6 o anti TNF-a y anticuerpo de detección biotinilado anti IL-6 o TNF-a.
Lipopolisacárido bacteriano (LPS)
Lipopolisacárido bacteriano purificado de E. coli, lote 05:B55 de SIGMA fue utilizado para los ensayos de tolerización in vivo y para otros ensayos realizados in vitro.
Tolerización con LPS
Hembras CBA/j preñadas de 4 - 5 meses de edad, fueron tratadas vía ip con 1 ó 2 µg de LPS. Se ensayaron protocolos de 3 - 4 días de duración en etapas pre y pos-implantación (días 0, 1 y 2,5 de preñez o días 6, 7, 8 y 9,5, respectivamente).
Desafío con LPS
Placentas y esplenocitos de los animales tolerizados y no tolerizados fueron desafiados in vitro con una dosis abortogénica de 5 µg/mL de LPS (10). Los sobrenadantes de cultivo fueron recolectados, centrifugados a 5000 rpm durante 5 min, luego a 10.000 rpm durante 10 min y conservados a –70 ºC para su posterior análisis.
Análisis Estadístico
Para el cálculo estadístico de los resultados se utilizaron los métodos de análisis de varianza tipo t-test o one way ANOVA, según correspondía. Para determinar las diferencias entre las medias se utilizaron los métodos de Mann-Whitney y Newman-Keuls de comparaciones múltiples.
RESULTADOS
Efecto del tratamiento con LPS sobre el índice de resorción
Habiéndose demostrado que el aborto provocado por un exceso de citoquinas inflamatorias en el modelo CBA/j x DBA/2 es dependiente de la presencia de LPS ambiental, los autores se propusieron inducir tolerancia inmunológica al LPS durante la preñez de estas hembras abortadoras. Para ello se inocularon dosis no abortogénicas de la endotoxina, en diferentes etapas gestacionales. Los resultados demostraron que la dosis de 2 µg/día de LPS ip no fue capaz de modificar el índice de pérdidas fetales, independientemente del tipo de tratamiento realizado (pre o post-implantación). Sin embargo, la inoculación post-implantacional con 1 µg de LPS disminuyó el índice de pérdidas fetales, aunque las diferencias encontradas con el número de animales analizados hasta ahora, no han sido significativas (Figura 1).

Figura 1.
Efecto del tratamiento con LPS sobre la síntesis de IL-6 en hembras abortadoras
Considerando que tanto el exceso como la deficiencia de IL-6 están asociados al aborto, se analizó el efecto del tratamiento con 1 µg de LPS sobre los niveles de producción de esta citoquina tanto en bazo como en placenta. Los resultados obtenidos demuestran que el tratamiento evita el aumento de IL-6 provocado por una exposición a altas dosis de LPS en el cultivo tanto de bazo (1,6 ± 0,2 vs. 1,4 ± 0,1 µg/mL en animales tratados y 0,9 ± 0,5 vs. 1,2 ± 0,2 µg/mL en animales no tratados) como de placenta (Figuras 2 y 3).
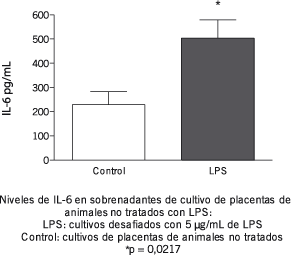
Figura 2.
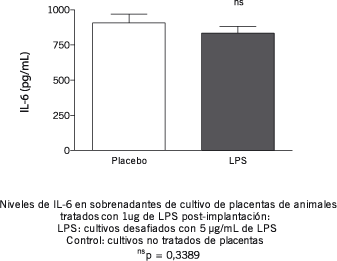
Figura 3.
Efecto del tratamiento con LPS sobre la síntesis de TNF-a en hembras abortadoras
Ya que ha sido publicado que el tratamiento con bajas dosis de LPS induce la inactivación del factor de transducción de señales NFkb provocando la supresión de la síntesis de citoquinas dependientes de esta vía (6), se ha estudiado el efecto del tratamiento sobre la síntesis de TNF-a. Los resultados en cultivos de esplenocitos se muestran en la Figura 4, donde puede observarse que el tratamiento in vivo con bajas dosis del lipopolisacárido fue capaz de inducir un cambio en la respuesta a un segundo desafío con dosis mayores de esta molécula, el cual puede observarse como una disminución significativa en la síntesis de TNF-a. Sin embargo, cuando se estudiaron los niveles de producción de TNF-a en las placentas de hembras tratadas con LPS, el estímulo con 5 µg/mL del lipopolisacárido indujo un aumento significativo en la producción de TNF- a in vitro, el cual puede observarse en la Figura 5.
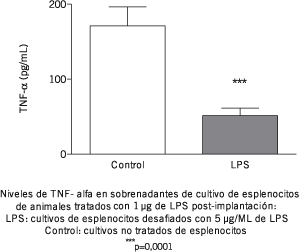
Figura 4.
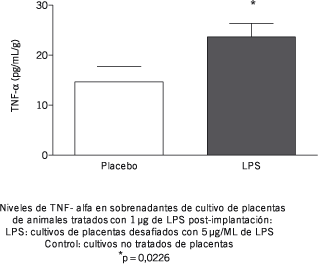
Figura 5.
DISCUSIÓN Y CONCLUSIONES
Se ha sugerido que los factores ambientales son capaces de desencadenar un desequilibrio en el balance de citoquinas Th1/Th2 durante la preñez, generando daños en la unidad fetoplacentaria que llevarían a la pérdida fetal. Entre estos factores ha sido recientemente identificado el lipopolisacárido bacteriano LPS, capaz de unirse a receptores del tipo Toll y desencadenar la síntesis de citoquinas Th1, vía el factor nuclear kb.
Dado que la muerte fetal recurrente de la cruza CBA/j x DBA/2 se produce por isquemia debido a un proceso inflamatorio exacerbado y dependiente de la presencia de LPS ambiental (1), y en base a estudios previos que han demostrado que el tratamiento con bajas dosis de LPS es capaz de reducir la respuesta de los macrófagos a una segunda estimulación (6), los autores se han propuesto investigar el efecto de una tolerancia al lipopolisacárido sobre la prevención del aborto y la producción de IL-6 y TNF-a en bazo y placenta de este modelo murino. Para ello, se analizaron dos dosis diferentes de LPS de 1 y 2 µg /día y en dos protocolos desde la etapa pre o post-implantatoria, ya que ha sido demostrado que cada citoquina tiene diferentes roles en la gestación, dependiendo de la dosis y de la ventana en que se la estudia (11). Los resultados obtenidos muestran que el tratamiento con 2 µg /día de LPS no produjo ningún efecto sobre el índice de resorción o sobre las citoquinas analizadas, independientemente de la etapa estudiada. Sin embargo, la dosis post-implantación de 1 µg /día fue capaz de reducir el índice de pérdidas fetales, aunque para el número de animales analizados hasta ahora, las diferencias no han sido estadísticamente significativas (Figura 1). Este mismo tratamiento fue capaz de prevenir el aumento de los niveles de IL-6 en respuesta a una dosis abortogénica de LPS, tanto en bazo (datos no mostrados) como en placenta (Figuras 2 y 3). Este efecto podría ser explicado por la disminución de la expresión del receptor Tlr4 de la respuesta innata provocada por la inducción de tolerancia al LPS, que lleva a la inactivación del NFkb (6), aunque serán necesarios otros estudios para demostrarlo. Futuras investigaciones serán destinadas a medir el ARNm del Tlr4 antes y después del tratamiento tolerizante de las hembras preñadas.
Por otra parte, se debe considerar que la principal población celular y fuente de IL-6 placentaria es el trofoblasto, y que si bien la expresión del Tlr4 ya ha sido demostrada en estas células, sería útil determinar si existen diferencias en los niveles de expresión entre la cruza abortadora CBA/j x DBA/2, y la cruza normal CBA/j x Balb/c. Si esto se confirmara podría hipotetizarse que a través de la sobre-expresión de esta molécula en las placentas CBA/j x DBA/2, la interfase materno-fetal se vuelve más sensible a dosis ambientales de LPS que llevan al aborto asociado a un aumento de citoquinas Th1, entre las que se encuentran el TNF-a. En base a esta hipótesis, se decidió estudiar la producción de esta citoquina en los cultivos de bazo y placentas de los animales tratados con 1 µg de LPS, y se encontró que ambos órganos mostraban un patrón diferente de respuesta al tratamiento y al posterior desafío in vitro. La exposición a altas dosis del lipopolisacárido de los esplenocitos del animal tolerizado, fue capaz de disminuir significativamente los niveles de producción de TNF-a (Figura 4), pero en la placenta produjo el resultado inverso (Figura 5). Esto lleva a concluir que el efecto del tratamiento tolerizante dependería tanto del órgano como de la dosis estudiada. Más aún, teniendo en cuenta que la regulación de la expresión de Tlr4 es dosis y tiempo- dependiente (6) y que su expresión se modifica según la ontogenia de cada órgano (12), el mismo protocolo de tolerización puede tener diferentes efectos dependiendo de la población celular y de la etapa gestacional estudiada. Por otro lado, a pesar de que la IL-6 y el TNF-a son sintetizados a partir de la activación del mismo factor NFkb, posiblemente los niveles finales relativos de cada citoquina dependan de las diferentes poblaciones celulares según expresen o no Tlr4. Más aún, cabe destacar que en la placenta la principal población leucocitaria no la constituyen los macrófagos sino las células NK, fuente de TNF-a. Si bien estas células expresan Tlr4, en la placenta un 70% de las NK son de un fenotipo especial llamado uNK (Natural Killer uterinas) en las cuales aún no ha sido estudiada la expresión del Tlr4. Teniendo en cuenta que el aborto inducido por LPS en ratones se previene al neutralizar la actividad de las µNK o del TNF-a (13) y que el efecto del tratamiento de tolerización produjo una disminución no significativa en el índice de resorción de la cruza abortadora , la expresión de Tlr4 en células µNK de mujeres abortadoras en comparación con mujeres fértiles será objeto de futuras investigaciones.
Es necesario destacar que esta es la primera vez que se genera un modelo de tolerancia al LPS durante la gestación, el cual cobra importancia a la hora de investigar los mecanismos subyacentes entre la respuesta inmune innata y la respuesta inmune adquirida, y entre la respuesta inmune inflamatoria y antinflamatoria, durante la gestación.
Agradecimientos
Los autores agradecen a la Dra. Isabel Piazzon, Laboratorio de Leucemia experimental, Instituto de Investigaciones Hematológicas, Academia Nacional de Medicina por su colaboración en este proyecto.
Correspondencia
LIC. VALERIA DUBINSKY
Instituto de Estudios de la Inmunidad Humoral (IDEHU)
Cátedra de Inmunología, FFyB, Universidad de Buenos Aires,
Junín 956 4P
1113 ciudad autónoma de BUENOS AIRES - Argentina
E-mail: gabgutie@ffyb.uba.ar
Referencias bibliográficas
1. Clark DA, Chaouat G, Gorczynski R. Thinking outside the box: mechanism of environmental selective pressures on the outcome of the materno-fetal relationship. Am J Reprod Immunol 2002;47 (5): 275-82
2. Ishida I, Kubo H, Suzuki S, Suzuki T, Akashi S, Inoue K, et al. Hipoxia diminishes Toll-like receptor 4 expresion through reactive oxygen species generated by mitochondria in endothelial cells. J Immunol 2002; 169: 2069-75.
3. Vikki MA. Toll-like receptors in the cycling female reproductive tract and during pregnancy. Current Women´s Health Rev 2005; 1: 35-42.
4. Chaouat, G. Synergy of lipopolysaccharide and inflammatory cytokines in murine pregnancy: Alloimmunization prevents abortion but does not affect the induction of preterm delivery. Cell Immunol 1994; 157: 328-40.
5. Silver RM. Bacterial lipopolysaccharide-mediated fetal death. J Clin Invest 1995; 95: 725-31.
6. Normura F, Akashi S, Sakako Y, Sato S, Kawai T, Matsumoto M, et al. Cutting edge: endotoxin tolerance in mouse peritoneal macrophages correlates with down-regulation of surface Toll-like receptor 4 expression. J Immunol 2000; 164: 3476.
7. Clark DS, McDermott MR, Szewczuk MR. Impairment of host-versus-graft reaction in pregnant mice. II. Selective suppression of cytotoxic T-cell generation correlates with soluble suppressors activity and with successful allogeneic pregnancy. Cell Immunol 1980; 52: 106-18.
8. Gutiérrez G, Sarto A, Berod L, Canellada A, Gentile T, Pascualini S, et al. Regulation of interleukin-6 fetoplacental levels could be involved in the protective effects of low-molecular weight heparin treatment on murine spontaneous abortion.. Am J Reprod Immunol 2004; 51: 160-5.
9. Suter M, Butler JE. The immunochemistry of sandwich ELISAs. II. A novel system prevents the denaturation of capture antibodies. Immunol Lett; 13(6): 313-6.
10. DW Dresser. The potentiating effect of pregnancy on humoral immune responses of mice. J Reprod Immunol 1991; 20: 253-66.
11. Chaouat G, Zourbas S, Ostojic S, Lappree-Delage G, Dubanchet S, Ledee N, et al. A brief review of recent data on some cytokine expressions at the materno-foetal interface which might challenge the classical Th1/Th2 dichotomy. Am J Reprod Immunol 2002; 53: 241-56.
12. Harju K, Glumoff V, Hallman M. Ontogeny of Toll-like receptors Tlr2 and Tlr4 in mice. Pediatr Res 2001; 49: 81-3.
13. Murphy SP, Fast LD, Hanna NN, Sharma S. Uterine NK cells mediate inflammation-induced fetal demise in IL-10-null mice. J Immunol 2005; 175: 4084-90. [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ]
Aceptado para su publicación el 10 de marzo de 2006

