










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkActa bioquímica clínica latinoamericana
versão impressa ISSN 0325-2957
Acta bioquím. clín. latinoam. v.44 n.2 La Plata mar./jun. 2010
BIOQUÍMICA CLÍNICA
Carotenoides en quimioprevención: Licopeno
Carotenoids in chemoprevention: Lycopene
Arturo Alberto Vitale1 *, Eduardo Alberto Bernatene2 *, Alicia Beatriz Pomilio3 *
1. Doctor en Ciencias Químicas (UBA).
2. Ingeniero Agrónomo (UBA).
3. Doctora de la Universidad de Buenos Aires.
* PRALIB (UBA-CONICET), Facultad de Farmacia y Bioquímica, Universidad de Buenos Aires, Junín 956, C1113AAD Ciudad Autónoma de Buenos Aires, Argentina.
Resumen
Las frutas y hortalizas son una rica fuente de carotenoides que proporcionan beneficios para la salud debido a que disminuyen el riesgo de varias enfermedades, en particular, ciertos tipos de cáncer y enfermedades cardiovasculares y oculares, lo cual está corroborado por una extensa observación epidemiológica. Los carotenoides que han sido más estudiados en este sentido son b-caroteno, licopeno, luteína y zeaxantina. La rápida y elevada cantidad de evidencias experimentales ha determinado que una dieta que contenga tomate o productos derivados del mismo, causa disminución de varios tipos de cáncer. Esto ha estimulado una amplia investigación de laboratorio y clínica, así como mucho entusiasmo comercial y público. Sin embargo, las evidencias son contradictorias. Otros trabajos de estos autores sobre compuestos antioxidantes, especialmente desde el punto de vista estructural y de bioactividad, han llevado a analizar si los efectos beneficiosos del licopeno se deben primariamente a sus propiedades antioxidantes o a otros mecanismos posibles que pueden estar involucrados, tales como la modulación de la comunicación intercelular por uniones nexus, los sistemas hormonal e inmune y las rutas metabólicas. En este trabajo se discuten aspectos estructurales, la actividad antioxidante del licopeno, su contenido en tomates, otros componentes bioactivos, mecanismos probables de acción y su influencia en cáncer de próstata, de colon, de mama y de endometrio, según estudios de casos-control, de cohortes y epidemiológicos.
Palabras clave: Carotenoides; Licopeno; Antioxidantes; Quimioprevención; Tomate
Summary
Fruits and vegetables are a rich source of carotenoids, which provide health benefits reducing the risk of several diseases, including certain cancers, cardiovascular and eye diseases, supported by extensive epidemiologic observation. Carotenoids that have been studied in this regard are b-carotene, lycopene, lutein and zeaxanthin. The rapid and large amount of experimental evidence has established that a diet containing tomato or products based on tomato results in a decrease of several types of cancer. This fact has stimulated extensive laboratory and clinical research as well as much commercial and public enthusiasm. However, evidence is controversial. The work of these authors on antioxidant compounds, especially in structural and bioactivity terms, has led to analyse whether the beneficial effects of lycopene are primarily due to its antioxidant properties or other possible mechanisms may be involved, such as the modulation of gap junctional intercellular communications, hormonal and immune systems, and metabolic pathways. In this paper, structural points of view, the antioxidant activity of lycopene, its content in tomatoes, other bioactive components, the possible mechanisms of action and their impact on prostate, colon, breast and endometrium cancer are discussed, according to case-control, cohort and epidemiological studies.
Keywords: Carotenoids; Lycopene; Antioxidants; Chemoprevention; Tomato
Introducción
En el laboratorio de los autores se continúa investigando los antioxidantes naturales, desde un punto de vista estructural, capacidad antioxidante y actividad quenching del oxígeno singulete, su distribución y su actividad biológica concomitante.
Como ejemplo de matriz acuosa, en este trabajo se ha estudiado tomate de dos cultivares diferentes que no habían sido analizados anteriormente, enfocado en el licopeno, que es su colorante natural, con una importante actividad antioxidante y de quenching del oxígeno singulete.
En el transcurso de esta investigación, surgieron otras realizadas por otros autores sobre carotenoides, en particular, licopeno y su actividad biológica, que pareció adecuado compilar.
1. ESTRUCTURA DE LOS CAROTENOIDES
Los carotenoides son tetraterpenoides que se encuentran en la naturaleza como pigmentos naturales liposolubles de numerosas frutas y verduras y se caracterizan por poseer una larga cadena poli-isoprénica. Los carotenos son los hidrocarburos de esta familia que corresponden a la fórmula C40H56, como por ejemplo: licopeno (Fig. 1), con dobles enlaces trans, si bien existen isómeros con algunas uniones cis.
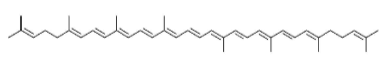
Figura 1. Estructura química del licopeno
Los carotenoides oxigenados, como alcoholes y epóxidos, se conocen como xantófilas, por ejemplo: luteína, violaxantina (Fig. 2).

Figura 2. Estructuras químicas de luteína y violaxantina
Los carotenoides se encuentran en plantas, animales y bacterias, muchas veces muy oxigenados o como cromoproteínas, siendo la parte prostética de éstas. Se suelen clasificar como: acíclicos, monocíclicos, aromáticos y ciclopentanoides, alénicos y acetilénicos, isoprenilados y degradados (1).
El consumo de una dieta rica en carotenoides se ha correlacionado epidemiológicamente con un menor riesgo de varias enfermedades, lo cual ha dado lugar a un creciente interés en el estudio de su biodisponibilidad (2)(3).
Luteína y zeaxantina (Fig. 3) son los carotenoides predominantes de la retina que actúan como fotoprotectores previniendo la degeneración retiniana. La distribución única, la localización y los altos niveles de ambos carotenoides en la mácula lútea, así como sus propiedades físico-químicas, los hace candidatos adecuados para la fotoprotección (2).

Figura 3. Estructura química de zeaxantina
El b-caroteno (Fig. 4) se utiliza como protector solar oral para la prevención de quemaduras de sol y resultó ser eficaz, ya sea solo o en combinación con otros carotenoides y vitaminas antioxidantes. Los efectos protectores se logran también con una dieta rica en licopeno (2).

Figura 4. Estructura química de b-caroteno
El licopeno es el pigmento lipofílico que confiere el color rojo característico a los tomates, y en menor cantidad, a otras frutas y verduras, como la sandía (45,3 μg/g) (4), pomelo rosado (14,2 μg/g), papaya y damasco, pero el 85% del licopeno proviene de fuentes de tomate, como la salsa de tomate en lata (287,6 μg/g) (5). El tomate fresco proporciona el 90% del licopeno necesario para el organismo, ya que es un micronutriente no sintetizado por el cuerpo humano.
A diferencia de otros carotenoides, como b-caroteno producido en gran escala por síntesis, el licopeno se obtiene fundamentalmente a partir de fuentes naturales, hongos y muy especialmente tomates. Sin embargo, los sistemas de extracción son costosos y el licopeno presenta una baja estabilidad, lo que ha limitado su utilización como colorante alimenticio.
El licopeno es un carotenoide acíclico con una cadena alifática formada por cuarenta átomos de carbono, con trece dobles enlaces de los cuales once son conjugados que le confieren la particularidad de ser muy reactivo frente al oxígeno y a los radicales libres. Por estas propiedades antioxidantes resulta probablemente eficiente como agente quimiopreventivo. Carece de un anillo de b-ionona, por lo cual no posee actividad provitamina A.
Aunque el tomate y sus productos contienen principalmente el isómero todo-trans del licopeno (79-91%), tanto el suero como los tejidos de los seres humanos y de los animales acumulan preferentemente los isómeros cis del licopeno (Fig. 5) (6) (7), correspondiendo al cis-licopeno el 79-88% del licopeno total de los tejidos benignos y malignos de próstata (6). En un estudio realizado por Stahl y Sies (8), el suero de individuos sanos contenía 50% de isómeros cis del licopeno después del consumo de jugo de tomate, que sólo contenía 20% de isómeros cis del licopeno.

Figura 5. Estructuras químicas de dos isómeros cis del licopeno.
La reactividad del licopeno se puede explicar debido a que se isomeriza rápidamente en sangre y en tejidos, desde la forma biosintética con configuración trans a una mezcla en la cual también participan los isómeros cis. Los isómeros geométricos cis (Fig. 5) presentan una mayor biodisponibilidad, probablemente porque son más solubles en las micelas del ácido biliar y por lo tanto pueden ser incorporados preferentemente en los quilomicrones (7). Aún resulta indefinido el rol biológico específico de los isómeros cis del licopeno en la salud humana.
Cuando se encuentra en la matriz de la planta o en estado sólido, el licopeno es relativamente estable, pero una vez extraído y disuelto en un solvente orgánico no polar, se vuelve inestable. En contraste con esta inestabilidad en solución en su forma pura, es estable cuando se presenta en perlas microencapsuladas dispersables en agua o en liposomas (9). Así se lo comercializa en algunos países.
Son pocos los datos acerca de la biodisponibilidad de licopeno sintético en humanos. Sin embargo, una suplementación durante 28 días con 15 mg/día de licopeno sintético (Lycovit 10% cápsulas) o con licopeno natural de tomate (Lyc-O-Mato) condujo a la misma biodisponibilidad en individuos sanos normolipémicos (10).
Se evaluó la biodisponibilidad en el intestino delgado y grueso de los principales carotenoides de frutas y hortalizas en una dieta común, usando un modelo de digestión enzimática y fermentación colónica in vitro (11). La luteína presentó mayor biodisponibilidad en el intestino delgado (79%) que el b-caroteno (27%) o el licopeno (40%). En cambio, en el intestino grueso, cantidades similares de licopeno y b-caroteno (57%) y sólo pequeñas cantidades de luteína (17%) fueron liberadas de la matriz alimentaria. Estos resultados sugieren que el 91% de b-caroteno, luteína y licopeno contenido en las frutas y hortalizas está disponible en el tracto gastrointestinal durante todo el proceso de digestión. La fermentación colónica resulta importante para la disponibilidad de carotenoides en el intestino.
Los carotenoides que son transportados por la sangre a través de las lipoproteínas, se concentran en tejidos específicos que poseen un gran número de receptores de lipoproteínas de baja densidad y una alta captación de las mismas, como ocurre en el hígado, las glándulas suprarrenales y los testículos. Si bien el licopeno es el carotenoide más abundante presente en la próstata (6), se acumulan otros carotenoides del tomate, como fitoeno, fitoaueno, ζ-, b- y γ-caroteno (Fig. 6) (12) (13). Los metabolitos del licopeno, tales como 2,6-ciclolicopen-1,5- dioles A y B, también se han encontrado en suero, leche, pulmones, mama, hígado, colon, piel y próstata de humanos (12). Sin embargo, el rol biológico específico de estos metabolitos no está claro.

Figura 6. Estructuras químicas de a-, γ-, δ-, ζ-carotenos, (15Z)-fitoeno, E-fitoeno y fitoflueno.
Se comparó la estabilidad a la auto-oxidación de los carotenoides polares, luteína y zeaxantina, con la de los carotenoides menos polares, b-caroteno y licopeno, a las concentraciones fisiológicamente relevantes patofisiológica- o fisiológicamente, de 2 y 6 μM, después de la exposición al calor o al humo de cigarrillos. Se extrajeron los carotenoides del plasma y se analizaron mediante cromatografía líquida de alta resolución (14).
La disminución de los niveles de b-caroteno y licopeno fue superior a la de los de luteína y zeaxantina. Asimismo, en los experimentos con humo de tabaco se encontraron las tasas más altas de auto-oxidación para b-caroteno y licopeno a 2 μM, pero a 6 μM luteína y zeaxantina disminuyeron en la misma medida que b-caroteno (14).
Estos datos apoyan los estudios previos que sugirieron que el estrés oxidativo degrada al b-caroteno y al licopeno más rápidamente que a la luteína y a la zeaxantina, excepto en la degradación térmica de los carotenoides disueltos en tetrahidrofurano pues se degradan más rápidamente el b-caroteno y la zeaxantina (14).
Debido a su estructura química, el licopeno es un compuesto extremadamente hidrofóbico que exhibe varias propiedades biológicas únicas y diferenciales.
Aunque los modelos de cultivos celulares permiten examinar los eventos moleculares y celulares potencialmente modulados por los compuestos fitoquímicos, se debería tener precaución al evaluar los efectos de los carotenoides in vitro ya que son a menudo oxidados y degradados como ya se indicara (15). Por lo tanto, los efectos biológicos de los carotenoides pueden ser debidos también a los productos de degradación de los carotenoides.
Su actividad biológica incluye la actividad antioxidante (quenching del oxígeno singulete y captación de radicales peroxilo), la inducción de la comunicación intercelular y el control de crecimiento, pero no la actividad de provitamina A.
2. CONTENIDO DE LICOPENO, FLAVONOIDES Y POLIFENOLES TOTALES EN EL TOMATE
Las principales fuentes de licopeno para los humanos son los tomates (cultivares y variedades de Lycopersicon esculentum) y los productos del tomate, pero la biodisponibilidad a partir de diferentes alimentos varía considerablemente. Factores como la madurez del tomate, los distintos genotipos o la forma culinaria de procesarlo influyen en la asimilación en el organismo y en el aprovechamiento del licopeno.
El consumo regular de tomates se ha asociado a través de estudios poblacionales con una disminución del riesgo de enfermedades crónico-degenerativas (16). Los resultados epidemiológicos confirmaron que los efectos observados sobre la salud se deben a la presencia de diferentes compuestos antioxidantes, como carotenoides, en particular licopeno, vitamina C, ß-caroteno y a-tocoferol, flavonoides y compuestos fenólicos (17). En este laboratorio, se estudiaron dos cultivares de extensa difusión en el Área Metropolitana de Buenos Aires (AMBA), Superman y Trafalgar, encontrándose que poseían 1,85 y 1,14 mg/100 g PF de rutina, que es un diglicósido del flavonol quercetina, mientras que contenían 9,16 y 8,05 mg/100 g PF de licopeno respectivamente.
El contenido de licopeno en varios productos de tomate varió de 42 ppm a 365 ppm. El nivel promedio diario de ingesta dietaria de licopeno resultó ser de 25,2 mg/día. Los tomates frescos representaron el 50% de la ingesta total de licopeno (16).
Es de destacar que la cantidad de licopeno presente en el tomate fresco depende del cultivar o de la variedad utilizada, las condiciones ambientales, el estado de maduración en el momento de la cosecha y el tratamiento post-cosecha previo a la llegada a la boca de expendio (18) (19). Por estos motivos, las cuantificaciones realizadas presentan diferencias y se deben especificar los parámetros indicados (Tabla I).
Tabla I. Determinación del contenido de licopeno en tomates frescos

Se ha determinado el patrón de flavonoides presentes en los tomates, como flavonoles y flavanonas, y de compuestos polifenólicos, como los ácidos fenólicos: ácido clorogénico (0,20 ± 0,03 mg/100 g) y ácido cafeico (0,03 ± 0,01 mg/100 g), las concentraciones de licopeno (valor medio de 5,38 ± 0,90 mg/100 g) y de b-caroteno (valor medio de 1,18 ± 0,40 mg/100 g) y la actividad antioxidante total de la fracción fenólica de diferentes líneas y cultivares de tomate (valor medio de 1,3 ± 0,10 mmoles/g). La caracterización se obtuvo por medio de análisis por espectrofotometría y por HPLC (16) (26). El valor medio para cada flavonoide fue de 0,68 ± 0,16 para naringenina, 0,74 ± 0,12 para rutina y 0,32 ± 0,06 para una rutina-pentósido. El contenido medio de polifenoles totales fue de 13,15 ± 1,15 mg/100 g (26).
Casi todas las líneas caracterizadas por bajo contenido de carotenoides producen altos niveles de polifenoles y por consiguiente, poseen el más poderoso potencial antioxidante.
La actividad antioxidante media de cuatro cultivares comerciales de tomate de Nueva Zelanda, cultivados bajo similares condiciones hidropónicas, medidos con el ensayo de decoloración del radical ABTS, osciló entre 2329 y 3268 μmoles TEAC/100 g de materia seca (MS) en los extractos hidrofílicos y de 178 a 303 μmoles TEAC/100 g MS en los extractos lipofílicos. Los principales componentes antioxidantes por cada 100 g MS fueron compuestos fenólicos (343-451 mg equivalentes a ácido gálico en el extracto hidrofílico y 53-74 mg equivalentes a ácido gálico en el extracto lipofílico), flavonoides (175-204 mg equivalentes a rutina), ácido ascórbico (153-195 mg) y licopeno (33-54 mg) (27).
Dado que existe un interés creciente en la capacidad de las dietas ricas en polifenoles de modular las enfermedades relacionadas con la edad y promover el envejecimiento saludable, se llevó a cabo un experimento piloto con ocho variedades de tomate para correlacionar la capacidad antioxidante total de las variantes del tomate con los flavonoides constitutivos específicos presentes. Se observó una fuerte correlación con el flavonoide rutina, pero no con otros flavonoides, tales como naringenin-chalcona o hidroxicinamatos, como ácido clorogénico, que también están presentes en el tomate (28). Para poner a prueba el rigor de esta correlación se realizó un segundo estudio con otras 37 variedades de tomate seleccionados por sus niveles bajos, medios y altos de rutina, demostrándose que la rutina contribuye en mayor medida a la capacidad antioxidante de los tomates, con lo cual este flavonoide puede ser una molécula diana útil para la sobrerregulación o desinhibición en los tomates con el fin de mejorar su estado antioxidante (28).
En el marco de programas de mejoramiento del tomate (breeding programs) con el objetivo de desarrollar genotipos nutricionales superiores, se estudiaron ocho componentes que contribuyen a la sana calidad del tomate (licopeno, b-caroteno, otros carotenoides, aavonoides, ácidos fenólicos, vitaminas C y E, residuo seco) (29). Doce líneas avanzadas de cultivo (breeding) de tomate y seis cultivares de polinización abierta se cultivaron bajo condiciones estrictamente controladas y se analizaron en cuanto a su contenido de antioxidantes. De los 18 genotipos analizados, 10 mostraron un nivel elevado de carotenoides totales, 6 alto nivel de b-caroteno, 9 alto nivel de licopeno, 15 de flavonoides y 2 una concentración relevante de vitamina E. Con estos datos y una compilación de la literatura sobre la composición del tomate, se propuso un índice, denominado índice de calidad nutricional antioxidante, como una herramienta para hacer frente a los programas de mejoramiento en la selección de genotipos del tomate con cualidades nutritivas antioxidantes (29).
Es importante destacar que los procesos industriales utilizados en la elaboración de productos derivados del tomate modifican el contenido de los metabolitos (Tabla II). Si bien existe una variada respuesta a los factores tecnológicos, en el caso del licopeno se produce un aumento en su biodisponibilidad (30) y también una disminución en su contenido (31). Los productos finales están afectados por tres factores fundamentales: genotipo del tomate, temperatura de secado y duración del mismo.
Tabla II. Contenido de licopeno e ingesta estimada en productos basados en el tomate (32)

Se estudiaron las variaciones del contenido de licopeno y de los principales antioxidantes (suma de los compuestos fenólicos totales y flavonoides totales) en tomates durante el proceso de secado con aire, teniendo en cuenta factores como cultivares del tomate, temperaturas de secado (40, 80 y 120 °C) y tiempo de secado (0 a 240 minutos). Los resultados mostraron que el contenido de licopeno aumentó a todas las temperaturas de secado durante los primeros 60 min. Sin embargo, los cultivares de tomate rojo, como HR (Holland Red), SN (Sheng-Neu) y TTL (Taur-Tay-Lang), presentaron una disminución significativa en el contenido de licopeno a 120 °C después del secado durante 75 min (31).
De acuerdo con los datos experimentales, se propuso un valor umbral de los principales antioxidantes de 500 mg/100 g de materia seca de tomate, tal que cuando el contenido de antioxidantes es inferior a este valor durante el secado con aire, el contenido de licopeno se reduce rápidamente en todos los cultivares de tomate. Además, los tomates de color amarillo, que contienen más antioxidantes totales inicialmente y los retienen más durante el secado con aire a 40 a 80 °C, resultaron ser los cultivares adecuados para el procesado térmico (31).
Capanoglu et al. (25) llegaron a la conclusión que se producen auctuaciones en el contenido de licopeno durante la elaboración de los productos basados en tomate, que están asociadas a cada una de las etapas. El resultado final es que el contenido de licopeno es inferior al de partida, probablemente debido a que la simulación del proceso en el laboratorio no provee los mismos productos finales que a escala industrial.
En los modelos animales en los que se estudió el cáncer de próstata, la suplementación con tomate provocó mayores efectos que la efectuada solamente con licopeno sobre la prevención carcinogénica y la mortalidad (33). Varios investigadores estudiaron la acción del suplemento de licopeno a través de los alimentos y encontraron una disponibilidad sistemática muy comparable con los productos de tomate procesados (34).
3. MECANISMOS DE ACCIÓN DE LOS TOMATES Y SUS COMPUESTOS FITOQUÍMICOS
Varios estudios indicaron que las personas que viven en la región del Mediterráneo tienen muy bajas tasas de enfermedades crónicas, como enfermedades cardiovasculares y cáncer, debido a la dieta mediterránea, que es rica en verduras, tomates, frutas, pescados y aceite de oliva (35).
Se propusieron e identificaron varios mecanismos de acción posibles para los compuestos fitoquímicos del tomate, en particular para el licopeno: actividad antioxidante, alteración del metabolismo xenobiótico, inhibición de la progresión del ciclo celular, inducción de apoptosis, aumento de la comunicación intercelular por uniones nexus, inhibición de transducción de señales del factor de crecimiento I tipo insulina (insulin-like growth factor-1, IGF-1) o modulación del eje IGF-1, inhibición de la activación y señalización de andrógenos, antiproliferación de células cancerígenas, actividad antimetastásica, entre otros (36-38).
Aunque variados, los mecanismos pueden ser complementarios y superpuestos, y una combinación de los mismos puede ser responsable de los efectos anticancerígenos de los compuestos fitoquímicos del tomate observados en estudios epidemiológicos, estudios con animales y en experimentos in vitro.
3.1. Actividad antioxidante y quenching del oxígeno singulete: Los carotenoides y los flavonoides presentes en los tomates poseen propiedades antioxidantes, por lo que disminuyen los radicales libres (39) (40). Más aún, los flavonoles dietarios (quercetina principalmente) protegen al ADN de los linfocitos humanos en diabéticos contra el daño oxidativo, lo cual es un efecto biológico de importancia médica potencial que parece estar asociado con la absorción de esos flavonoles dietarios (41). Tanto a-tocoferol como quercetina resultaron ser inhibidores potentes del daño al ADN (espontáneo o inducido por carcinógenos) de una manera dependiente de la dosis. Los jugos frescos de calabaza amarga y de tomate también podrían proteger del daño al ADN inducido por DMBA, pero no tan efectivamente como los agentes individuales (42). Además, el licopeno es el antioxidante más potente en inhibir (quenching) al oxígeno singulete in vitro respecto a otros carotenoides (17) (43). Individuos sanos suplementados durante 15 días con productos de tomate presentaron un aumento significativo en la demora de la oxidación ex vivo de las lipoproteínas (44) y varios estudios demostraron que el consumo de tomate y jugo de tomate llevan a una disminución del daño del ADN de los linfocitos (45). Estos estudios sugieren que el consumo de tomate puede proporcionar protección contra el daño oxidativo in vivo, lo que potencialmente previene las mutaciones asociadas con la iniciación y la progresión del cáncer.
3.2. Alteración del metabolismo de los xenobióticos: Breinholt et al. (46) encontraron que el licopeno indujo significativamente las enzimas de la fase I, como las enzimas que dependen del citocromo P450, de una manera dependiente de la dosis y aumentó al doble la quinon- reductasa (QR) hepática, una enzima de la fase II. Otros estudios han demostrado que el licopeno indujo enzimas de desintoxicación de la fase II en una variedad de modelos animales (47) (48). Esta clase de enzimas es importante para la eliminación de sustancias extrañas y de carcinógenos del cuerpo. Los flavonoides del tomate, tales como canferol, quercetina y naringenina, han demostrado ser muy potentes y selectivos en la inhibición de las isoformas del citocromo P450-1A y otros estudios demostraron que la quercetina induce a QR (39).
3.3. Modulación de los factores de crecimiento: Se sabe que las concentraciones circulantes de IGFs y varios IGFBPs están inauenciados por los hábitos dietarios y otros factores del estilo de vida (49). Un estilo de vida occidental se asocia positivamente con el riesgo de cáncer, en parte a través de efectos sobre la insulina y los IGFs (50). Tanto la insulina como IGF-I pueden estimular el crecimiento tumoral por inducir la proliferación e inhibir la apoptosis. Se considera que las proteínas de unión a IGF (IGFBPs) inhiben y estimulan la interacción de IGF-I con el receptor de IGF-I (51).
Las alteraciones en la actividad de IGF-1, que estimula la proliferación y la resistencia apoptótica en las células, se examinaron en un estudio de caso-control de 112 hombres (52). El consumo del tomate cocido se asoció con una disminución del 31,5% en los niveles séricos de IGF-1.
La administración de suplementos de licopeno se encontró que disminuía significativamente la expresión tumoral de IGF-1 en ratas (53). Se halló una tendencia significativa a menor IGF-1 en suero y mayor proteína- 3 unida al factor de crecimiento tipo insulina (insulin-like growth factor binding protein-3, IGFBP-3) con un mayor consumo semanal de ketchup y jugo de tomate en 344 hombres libres de enfermedades (54) y se encontró una disminución similar en la relación de IGF-1 a IGFBP-3 en hurones alimentados con licopeno (55). Una menor relación de IGF-1 a IGFBP-3 se considera beneficiosa dado que IGFBP-3 se une a IGF-1, impidiendo así que IGF-1 estimule la proliferación celular. Es decir que, tanto el licopeno como los flavonoides del tomate, quercetina, canferol y rutina, inter`eren con la señalización de IGF- 1 in vitro, evitando así que el factor de crecimiento estimule la proliferación celular (53) (55-60).
Estudios epidemiológicos prospectivos indicaron que las concentraciones circulantes de IGF-I total relativamente altas están asociadas con mayor riesgo de cáncer de próstata, premenopáusico de mama y colorrectal, mientras que se conocen asociaciones tanto positivas como negativas para IGFBP-3 (61).
En 3 de 6 estudios transversales (52) (54) (62-65), la mayor ingesta de tomates cocidos o procesados, o licopeno, se asoció con menor concentración de IGF-I (52), mayor concentración de IGFBP-3 (63), o bien, una relación molar inferior de IGF-I a IGFBP-3 (54).
El licopeno puede aumentar las concentraciones de IGFBP-1 sérico en mujeres y IGFBP-2 sérico en hombres, lo que puede dar lugar a una menor biodisponibilidad de IGF-I (66). Por lo tanto, puede proporcionar un medio de última instancia para reducir el riesgo de cáncer colorrectal y potencialmente los riesgos de otros importantes tipos de cáncer, como cáncer de próstata y premenopáusico de mama. Sin embargo, la variación interindividual en los efectos de IGFBP-1 y -2 fue elevada, posiblemente complicada por las diferencias en la duración del ayuno y, en consecuencia, en las concentraciones de insulina. Por lo tanto, los resultados deben confirmarse en estudios más grandes de intervención, aleatorizados, con control de la duración del ayuno (66).
3.4. Control del crecimiento celular: El licopeno indujo la detención del ciclo celular en una serie de líneas celulares de cáncer mediante el bloqueo de la transición de la fase G1 a la fase S del ciclo celular. En las células epiteliales normales de la próstata, el tratamiento con licopeno dio lugar a una disminución dependiente de la dosis en la ciclina D1, que es una proteína que regula la transición de la fase G1 a la fase S en las células (67). En las células MCF-7 y T-47d de cáncer de mama y en las células ECC-1 de cáncer de endometrio, se encontró también que el licopeno disminuye los niveles de ciclina D1 y mantiene los niveles de p27 en los complejos de ciclina E-cdk2 (68).
La p27 es una proteína inhibidora de la quinasa dependiente de ciclina (cdk) que impide la progresión celular de la fase G1 a la fase S, provocando así la detención del ciclo celular. Efectos similares del licopeno sobre los niveles de p27 en células de cáncer de próstata no se han examinado exhaustivamente.
3.5. Inducción de la comunicación intercelular: Se propuso que el licopeno y sus metabolitos aumentan la comunicación intercelular de uniones de nexo mediante el aumento de los niveles de conexina 43 (69) (70). La formación de uniones nexus permite la comunicación de célula a célula, lo cual es importante en la regulación del crecimiento celular rápido, descontrolado. Resulta de interés que, la inhibición del crecimiento por licopeno en células PC-3 de cáncer de próstata, MCF-7 de mama y KB- 1 bucal, se reflejó en el aumento de los niveles de conexina 43, mientras que el licopeno falló en inhibir el crecimiento o en aumentar los niveles de conexina 43 en células metastásicas PC-3MM2 de cáncer de próstata y células normales de la mucosa bucal (71) (72), sugiriendo así que el aumento de la conexina 43 puede ser importante para la acción del licopeno contra el cáncer.
En la Tabla III se describen varios trabajos en los que se evaluó el comportamiento del licopeno para prevenir y tratar enfermedades crónicas.
Tabla III. Algunos estudios en los cuales se analizan los efectos del licopeno

4. ACTIVIDAD ANTIOXIDANTE
La actividad antioxidante de los carotenoides y las propiedades bioquímicas que inauyen en las vías de señalización se han discutido como mecanismos básicos de prevención. El concepto ha sido cuestionado debido a los datos conaictivos de estudios de intervención con b-caroteno para prevenir diversos tipos de cáncer y enfermedades cardiovasculares. Pero el b-caroteno es distinto al licopeno, ya sea estructural como mecanísticamente. Los estudios epidemiológicos sugieren los efectos protectores del licopeno en algunos tipos de cáncer, por ejemplo: cáncer de próstata (79). Los estudios in vitro e in vivo sobre el crecimiento de células tumorales apoyan esta conclusión (8).
Debido a su sistema de dobles enlaces conjugados, el licopeno es un sistema rico en electrones, susceptible de ser atacado por parte de los reactivos electrofílicos. Por lo tanto, el licopeno puede unirse al oxígeno singulete y a los radicales libres como el radical hidroxilo (HO·) y varios radicales peróxido. Este comportamiento es la base de su acción antioxidante en los sistemas biológicos, por lo que es un eficiente agente quimiopreventivo.
Las especies reactivas del oxígeno (en inglés: reactive oxygen species, ROS), como el anión radical superóxido o el radical hidroxilo, se generan en forma natural a partir del metabolismo basal, la respiración celular, los ciclos de oxidación microsomales y durante los procesos inflamatorios (80). Sin embargo, algunos tóxicos pueden sufrir ciclos de óxido-reducción que incrementan esta generación natural produciendo un desequilibrio que es el conocido estrés oxidativo. Es decir que el estrés oxidativo es la pérdida del equilibrio entre la producción de radicales libres y la acción de los sistemas antioxidantes, a favor de los primeros.
El estrés oxidativo causado por los radicales libres y ROS puede dañar a las macromoléculas, como hidratos de carbono, lípidos, proteínas y ADN, pudiendo desencadenar carcinogénesis, envejecimiento prematuro y enfermedades cardiovasculares. Los radicales libres pueden interaccionar con los lípidos insaturados de la membrana celular, provocando la peroxidación de los mismos y conduciendo finalmente a una destrucción generalizada de la membrana. Además, se pueden generar productos estables de degradación de la membrana que dañen al genoma u otras estructuras celulares (81) (82).
Es decir que los radicales libres o ROS y el daño oxidativo inducido por ellos, han sido implicados en la patogenia de varias enfermedades humanas (81) (83), como diabetes, formación de placas ateromatosas por oxidación y depósito de lipoproteínas de baja densidad (low-density lipoprotein: LDL) y carcinogénesis por genotoxicidad (44) (84) (85). También el daño fotooxidativo participa en la bioquímica patológica de varias enfermedades que afectan a la piel y a los ojos, y los carotenoides pueden proteger los tejidos expuestos a la luz (2). El b-caroteno puede tener beneficios adicionales debido a su capacidad de convertirse en vitamina A (86).
El estrés oxidativo se ha logrado medir mediante distintas técnicas; una de ellas es el análisis de la actividad de ciertas enzimas en sangre y de la capacidad antioxidante total del plasma (87).
Los sistemas antioxidantes que inactivan a los radicales libres pueden clasificarse en sistemas enzimáticos de acción intracelular como: catalasa (CAT), superóxido-dismutasa (SOD) y glutatión-peroxidasa (GSH-Px) (88) (89) y sistemas no enzimáticos, que son sustancias eliminadoras o secuestradoras de ROS como: a-tocoferol (vitamina E), ácido ascórbico, carotenos aportados por la dieta, glutatión y licopeno (90). Estos últimos están presentes en el tomate.
Dentro de los agentes antioxidantes no enzimáticos, el licopeno es uno de los antioxidantes más potentes conocidos hasta ahora, propiedad dada por su gran capacidad de secuestrar ROS in vitro (43) y, en especial, por eliminar al oxígeno singulete (17) (44). Sus propiedades químicas le confieren una potencial acción preventiva de las enfermedades cardiovasculares, protegiendo a las biomoléculas críticas como los lípidos y LDL ya que como antioxidante evita que el colesterol "malo", o LDL, se oxide y produzca daños a la membrana de las células (acción anti-estrés oxidativo). El licopeno también tiene efecto anticarcinogénico, protegiendo a las proteínas y al ADN ya que atrapa a los radicales libres que afectan a las membranas celulares y atacan el material genético de las células (17). El licopeno puede desregular el elemento de respuesta antioxidante y por lo tanto estimular la producción de enzimas celulares, como la peróxido-dismutasa, la glutatión-S-transferasa y la quinon-reductasa para proteger a las células de ROS y de otras moléculas electrofílicas.
La capacidad del licopeno para retrasar la oxidación de las células, y de este modo su multiplicación, garantiza su capacidad preventiva. Los distintos isómeros configuracionales del licopeno disminuyen los niveles del antígeno prostático específico (PSA) evitando el desarrollo de tumores secundarios.
Se ha señalado el rol del óxido nítrico (NO) en la carcinogénesis, ya que se ha detectado gran cantidad de NO en los tejidos cancerosos y se encontró que la expresión de la NO sintasa inducible (iNOS) se correlaciona con el crecimiento tumoral y la metástasis. También se encontró que NO poseía actividad de iniciación tumoral en la carcinogénesis de piel de ratón.
La supresión de la inducción iNOS también se observó en el ratón fitoeno sintasa transgénico. Tras la administración de glicerol (un promotor del tumor pulmonar), se observó menor inducción del gen de iNOS en el pulmón de los ratones que producen fitoeno, comparando con los ratones control. El uso combinado de varios tipos de antioxidantes distribuidos en los alimentos, por ejemplo: mezcla de carotenoides y flavonoides, mostró ser un método efectivo para la prevención del cáncer (91).
Reacciones fotooxidativas: La exposición de los organismos vivos a la luz ultravioleta provoca reacciones fotooxidativas. Los carotenoides son antioxidantes dietarios y muestran efectos fotoprotectores en la piel humana, atrapando eficientemente radicales peroxilo e inhibiendo la peroxidación lipídica (92).
Se usaron fibroblastos cultivados de piel humana para examinar los efectos protectores de los carotenoides, licopeno, b-caroteno y luteína sobre la peroxidación lipídica inducida por UVB. Los carotenoides fueron distribuidos en las células mediante liposomas. Las células fueron expuestas a la luz UVB durante 20 min. Licopeno, b-caroteno y luteína fueron capaces de disminuir la formación de especies reactivas al ácido tiobarbitúrico (thiobarbituric reactive species: TBARS) inducidas por UV en 1 h a niveles de 40-50% de los controles libres de carotenoides (92).
Se describió la respuesta sérica de los carotenoides y tocoferoles ante la suplementación con carotenoides de extractos naturales (a-caroteno + b-caroteno, luteína o licopeno; 15 mg/día) y/o con a-tocoferol (100 mg/día) en un estudio de intervención multicéntrico, controlado con placebo, de 400 voluntarios sanos, hombres y mujeres, entre 25-45 años de edad, procedentes de cinco regiones europeas (Francia, Irlanda del Norte, República de Irlanda, Holanda y España) (93).
No se observaron efectos secundarios significativos (excepto carotenodermia) o cambios en los índices bioquímicos o hematológicos a lo largo del estudio. Se describieron las respuestas dosis-tiempo, la distribución de los isómeros, la variabilidad de los individuos y los efectos secundarios durante la suplementación con los principales carotenoides dietarios en individuos sanos (93).
El estrés oxidativo juega también un rol importante en el desarrollo de enfermedades neurodegenerativas. Las enfermedades neurodegenerativas son un grupo de enfermedades con diferente importancia clínica y etiología, como la enfermedad neurona motora, la esclerosis lateral amiotrófica, desórdenes del cerebelo, enfermedad de Parkinson, enfermedad de Huntington, enfermedad de Alzheimer cortical destructiva y esquizofrenia.
Numerosos estudios epidemiológicos y experimentales muestran muchos factores de riesgo, como la edad avanzada, defectos genéticos, anomalías de enzimas antioxidantes, excitotoxicidad, anormalidades del citoesqueleto, autoinmunidad, deficiencias de minerales, estrés oxidativo, toxicidad metabólica, hipertensión y otros trastornos vasculares (94) (96-98).
El creciente conjunto de evidencias implica la toxicidad de los radicales libres, las mutaciones inducidas por los radicales y el deterioro oxidativo de las enzimas y la disfunción mitocondrial debido a defectos genéticos congénitos en las manifestaciones clínicas de enfermedades neurodegenerativas (94) (96-98).
Recientemente se midieron los marcadores de estrés oxidativo en muestras de sangre de 338 pacientes (965 observaciones) con Alzheimer, demencia vascular, diabetes (tipo II) superpuestas a demencias, enfermedad de Parkinson y controles (99). El estrés oxidativo sistémico se asoció significativamente (P << 0,001) con todas estas enfermedades indicando un desbalance en las respuestas periféricas/adaptativas a los desórdenes oxidativos a través de diferentes pasos metabólicos de radicales libres (99).
Se cree que los carotenoides tienen un beneficio terapéutico en el tratamiento de las enfermedades cardiovasculares a causa de sus propiedades antioxidantes. Sin embargo, ensayos aleatorios prospectivos han fallado en demostrar un beneficio consistente de b-caroteno en pacientes con riesgo de enfermedades cardiovasculares. Esto puede atribuirse a las distintas propiedades antioxidantes de varios carotenoides causantes de interacciones físico-químicas, dependientes de la estructura, con las membranas biológicas. Para probar esta hipótesis, se midieron los efectos de astaxantina, zeaxantina, luteína, b-caroteno y licopeno sobre la peroxidación lipídica utilizando membranas modelo enriquecidas con ácidos grasos poli-insaturados (100). Los efectos correlativos de estos compuestos sobre la estructura de la membrana se determinaron utilizando las metodologías de difracción de rayos X de pequeño ángulo (small-angle x-ray diffraction). Los carotenoides no-polares, licopeno y b-caroteno, desordenaron la bicapa de la membrana y estimularon la peroxidación lipídica de la membrana (>85% de aumento en los niveles de hidroperóxido lipídico), mientras que astaxantina (carotenoide polar) preservó la estructura de la membrana y exhibió una actividad antioxidante significativa (>40% de disminución en los niveles de hidroperóxido lipídico) (100).
Estos resultados sugieren que el potencial antioxidante de los carotenoides depende de sus distintas interacciones con la membrana lipídica. Esta relación de estructura y función puede explicar las diferencias en la actividad biológica informada para varios carotenoides, con importantes implicancias terapéuticas (100).
Si bien los resultados de experimentos in vitro, estudios epidemiológicos y ensayos clínicos, informan que los carotenoides pueden ser poderosos antioxidantes in vivo, hay escasos datos sobre la administración de suplementos de licopeno purificado per se en humanos. Por lo tanto, recientemente se realizó un estudio dosis-respuesta de los efectos de diferentes dosis de suplementos de licopeno purificado (0, 6,5, 15, 30 mg/día) sobre los biomarcadores de estrés oxidativo en voluntarios sanos (101). La selección de la dosis (6,5, 15 ó 30 mg de licopeno) se basó en estudios anteriores, que demostraron que a estas dosis, alcanzables a través de la dieta, el licopeno está biodisponible y ejerce protección contra el estrés oxidativo (33) (102). Este fue un ensayo de doble ciego, aleatorizado, controlado con placebo, que examinó los efectos de 8 semanas de administración de suplementos de licopeno purificado. Como resultado, los niveles de licopeno en plasma, independientemente de la dosis, aumentaron significativamente en todos los grupos suplementados con licopeno versus placebo. ANOVA reveló una disminución significativa de los daños en el ADN mediante el ensayo cometa y una disminución significativa en 8-hidroxi-2'-desoxiguanosina (8-OHdG) urinaria a las 8 semanas versus la línea basal, con 30 mg de licopeno/día. No se observaron diferencias inter- o intra-grupos para la glucosa, perfil lipídico u otros biomarcadores de peroxidación lipídica en cualquier punto de dosis/ tiempo (101).
Estos datos recientes muestran el rol exclusivo del licopeno como antioxidante celular en reducir la susceptibilidad del ADN de los linfocitos contra el daño oxidativo y 8-OHdG urinaria, en individuos sanos que consumieron 30 mg diarios de licopeno purificado. Estos efectos del licopeno pueden tener importantes consecuencias en la prevención de cáncer de próstata, pulmón y estómago, como se informó en anteriores observaciones epidemiológicas y requiere más investigación con ensayos clínicos más grandes (101).
Otro importante biomarcador del daño al ADN que se afectó por la suplementación con licopeno a 30 mg/día fue 8-OHdG urinaria (101). Se había observado anteriormente un aumento de 8-OHdG en los tejidos diana de varios modelos de cáncer en animales y en leucocitos humanos de pacientes con varias enfermedades asociadas con el estrés oxidativo (103). Bowen et al. (104) observaron una disminución significativa en 8- OHdG de leucocitos en pacientes con adenocarcinoma de próstata localizado, que consumían platos de pasta con salsa de tomate durante 3 semanas, proporcionando alrededor de 30 mg de licopeno/día.
De todos los carotenoides, el licopeno ha demostrado ser el más potente antioxidante in vitro, haciendo que muchos investigadores llegaran a la conclusión que las propiedades antioxidantes del licopeno son las responsables de la prevención de enfermedades. Sin embargo, al compilar recientemente los ensayos en humanos y en animales con licopeno o con extractos conteniendo licopeno, se observó un apoyo limitado a la función antioxidante in vivo para el licopeno (105). Más aún, los niveles de licopeno en los tejidos suelen ser demasiado bajos para desempeñar una función antioxidante significativa. Por lo tanto, se llegó a la conclusión que hay una escasez general de evidencias de apoyo a la "hipótesis antioxidante" como el principal mecanismo de acción in vivo para el licopeno. Asimismo, se postuló que los productos metabólicos del licopeno, denominados licopenoides, pueden ser bioactivos y responsables de parte de la bioactividad indicada para el licopeno (105). Los licopenoides pueden producirse a partir de la monooxigenasa de los carotenoides: CMO II (carotenoid monooxygenase II: CMO II), en forma similar a la producción de retinoides a partir de b-caroteno por CMO I (106) (107) (Fig. 7). También se cree que, debido a la similitud estructural, el fitoeno y el fitoaueno pueden también clivarse como el licopeno con CMO II para formar fitoenoides y fitoauenoides (Fig. 7). Los metabolitos de los carotenos del tomate pueden ser responsables del menor riesgo de cáncer de próstata en hombres que consumen grandes cantidades de productos del tomate.

Figura 7. Metabolismo propuesto para los carotenoides del tomate. Formación de licopenoides.
4.1. Tomate y estrés oxidativo: Los estudios clínicos han demostrado que el consumo dietario de productos del tomate, que contienen licopeno, disminuye los biomarcadores de estrés oxidativo (daño celular del ADN y biomarcadores de la oxidación de lípidos) en individuos sanos, fumadores y diabéticos tipo 2 (44) (85) (108-110). Si bien los efectos sobre la peroxidación lipídica han sido algo conaictivos (111-113), varios estudios han demostrado consistentemente una disminución significativa en el daño al ADN de los linfocitos, a raíz de la intervención dietaria con productos del tomate en voluntarios humanos sanos (110) (114). Sin embargo, estos estudios no sugieren un rol del licopeno solo, sino que otros compuestos antioxidantes del tomate, como otros carotenoides, vitaminas C y E y flavonoides, pueden interactuar para conferir un beneficio preventivo contra las enfermedades asociadas al estrés oxidativo (115).
Con respecto a la peroxidación lipídica, los estudios in vitro han mostrado una capacidad de la óleo-resina del tomate superior a la del licopeno, en la inhibición in vitro de la oxidación de LDL de hasta cinco veces. El licopeno purificado se demostró que actúa sinérgicamente con otros antioxidantes naturales como la vitamina E, el flavonoide glabridina, los compuestos fenólicos ácido rosmarínico y ácido carnósico, y ajo, en la inhibición in vitro de la oxidación de LDL (116).
Los tomates contienen una matriz con potasio, foliato, vitaminas A, C y E, otros carotenoides (a-, b- γ -caroteno, luteína, fitoeno, fitoaueno) y flavonoides. El consumo del tomate podría aumentar las lipoproteínas de alta densidad (HDL) y disminuir el estrés oxidativo a corto plazo ya que el tomate tiene una alta capacidad antioxidante debido a su alto contenido de nutrientes y compuestos fitoquímicos, que pueden actuar de manera sinérgica como antioxidantes a través de interacciones complejas (117). Los tomates también son una fuente concentrada de flavonoles, con hasta un máximo de 98% del total de flavonoles contenidos en la piel del tomate como formas conjugadas de quercetina y canferol (118) (119). La flavanona naringenina está presente en pequeñas cantidades en los tomates en su forma conjugada (120). La fibra es otro componente dietario que se asocia a la disminución de riesgo de cáncer y en los productos del tomate se encuentran cantidades apreciables. De los productos del tomate, la pasta de tomate es la que posee más fibra, 11,8 g de fibra por taza.
Se han establecido claramente los niveles séricos de licopeno y cómo éstos varían con el consumo de un derivado de tomate. Sin embargo, sus efectos beneficiosos son significativos sólo cuando se incorpora a través del tomate y no aisladamente (44) (85).
Se investigó la actividad antioxidante sinergística del licopeno en combinación con b-caroteno, vitamina E y luteína, utilizando un sistema de liposomas. Las mezclas de carotenoides fueron más eficientes en la protección de los liposomas de la oxidación que cada carotenoide individualmente. Las mezclas de licopeno y vitamina E presentaron la mayor actividad antioxidante sinergística (117).
Estudios observacionales han informado la asociación positiva entre el consumo del tomate y sus derivados y las concentraciones totales plasmáticas de licopenos y de HDL (121).
Más de 600 factores no-nutrientes de alimentos funcionales (functional food factors, FFFs) en hortalizas y frutas se consideran eficaces para la promoción de la salud y la prevención de enfermedades. Se construyó una base de datos con estos factores para estimar las clases químicas y la cantidad total de ingesta de los mismos, con el fin de facilitar la investigación nutricional. Hasta el momento, flavonoides, terpenoides, carotenoides y compuestos de azufre están incluídos en esa base de datos de FFF (122).
Se conocen datos comparativos sobre la absorción y transferencia relativas de siete carotenoides diferentes: a-caroteno, b-caroteno, licopeno, astaxantina, b-criptoxantina, luteína y zeaxantina. Estos datos indican que se absorbieron menores cantidades de carotenoides y se transfirieron de manera más eficiente que en el caso de cantidades mayores, lo cual sugiere un efecto de saturación a mayor exposición (3).
Los mecanismos mediante los cuales los carotenoides ejercen el efecto quimiopreventivo son controvertidos. La mayor parte de la evidencia acumulada se ha hecho en base a estudios efectuados in vitro o retrospectivos, siendo escasos y controversiales los estudios hasta ahora realizados con seres humanos. Por ello, Madrid et al. (123) evaluaron in vivo los efectos de los suplementos dietarios con puro jugo de tomate concentrado sobre los cambios a corto plazo en el estado oxidativo y en las lipoproteínas plasmáticas en 17 voluntarios sanos, que recibieron un suplemento de puro jugo de tomate durante 7 días. En la línea basal, al final del período de suplementación y ocho días después del final de la suplementación, se extrajo una muestra de sangre para medir la capacidad antioxidante total (total antioxidant capacity: TRAP), los antioxidantes enzimáticos (CAT y SOD), los antioxidantes no enzimáticos (licopeno y a-tocoferol) y las lipoproteínas plasmáticas (123).
Como resultado, el nivel de licopeno aumentó al principio y significativamente en comparación con los niveles basales (48%). TRAP, CAT y SOD no variaron significativamente. El colesterol-HDL aumentó significativamente en 5,6 ± 4,3 mg/dL en el segundo período de muestreo, mejorando la relación colesterol/HDL. La elevación de los valores plasmáticos de HDL resultó rápida y significativa, siendo uno de los aspectos de mayor relevancia en este estudio, ya que presenta una potencial capacidad terapéutica en pacientes con dislipidemias. Estos hallazgos concuerdan con un estudio de tipo observacional en 1.687 adultos mayores del Reino Unido que presenta una asociación positiva entre concentración de licopenos y de HDL (121).
Contrariamente a lo informado en estudios in vitro, la actividad de los antioxidantes plasmáticos enzimáticos (TRAP, CAT y SOD) no sufrieron modificaciones significativas durante el período de suplementación (123). Esta situación puede deberse a que el tiempo de exposición al jugo de tomates fue corto y los efectos en la capacidad antioxidante total y en la actividad enzimática final no alcanzaron a verificarse, debido a que estas enzimas pueden requerir un mayor tiempo para su inducción intracelular.
Los antioxidantes no enzimáticos (licopeno y a-tocoferol) sí sufrieron una modificación manifiesta con el suministro de jugo de tomate. Esto, y el mantener la capacidad antioxidante total del plasma, parece ser la expresión de un equilibrio oxidativo compensatorio fisiológico, debido a que la homeostasis de las sustancias oxidantes del organismo se obtiene mediante un control enzimático y la participación de los antioxidantes de los alimentos (124). Los niveles de a-tocoferol disminuyen, probablemente, como consecuencia de su acción antioxidante precoz; sin embargo, hay que considerar que ambos antioxidantes no enzimáticos son liposolubles, por lo que el contenido plasmático medido representa sólo una pequeña fracción del a-tocoferol y licopeno total inserto en las membranas lipídicas. Además, la diferencia verificada entre los estudios puede tener explicación en la distinta biodisponibilidad de los carotenoides en los derivados del tomate que se utilizan en ellos.
Los estudios de cohortes sugieren que los individuos con mayor ingesta de tomates y productos de tomate tienen un menor riesgo de enfermedades degenerativas. Recientemente, se intentó distinguir el efecto del licopeno del de tomate en el estrés oxidativo, mediante el uso de tomate de color amarillo, una variedad de tomate desprovista de licopeno. Para ello, se compararon los efectos de la alimentación con ninguno (control), con 16% de tomate amarillo liofilizado (TA), con 16% de tomate rojo liofilizado (TR) ó 0,05% de glóbulos de licopeno (L) en un modelo de rata con estrés oxidativo leve inducido por una dieta baja en vitamina E. Se evaluaron los parámetros fisiológicos, tales como la ingesta, peso del cuerpo, bazo e hígado, niveles de colesterol y triglicéridos (TG). Se midieron las concentraciones de licopeno y de vitamina E y los biomarcadores de estrés oxidativo en el plasma y/o tejido hepático y/o cardíaco de las ratas (125).
Como resultado, TR, TA y L no tuvieron ningún efecto sobre la ingesta de las ratas, el peso del cuerpo y del bazo. TR, TA y L no tuvieron ningún efecto sobre la concentración plasmática de colesterol. TR disminuyó significativamente el nivel de TG en comparación con el control, TA y L (125).
Las ratas alimentadas con TR o L acumularon licopeno en el plasma, en contraste con las ratas alimentadas con TA. El nivel en corazón de TBARS en ratas alimentadas con TR o TA fue significativamente menor que el del control y el de las ratas alimentadas con L (125).
A pesar de las concentraciones similares de licopeno en plasma y en hígado, las ratas alimentadas con L mostraron un nivel significativamente superior de TBARS en corazón que las ratas alimentadas con tomates. TR aumentó la actividad de superóxido-dismutasa en eritrocitos (erythrocyte superoxide dismutase: eSOD) en comparación con el nivel de L y óxido nítrico (NO) comparado con el control y L. L disminuyó significativamente el nivel de la capacidad reductora de hierro del plasma (ferric reducing ability of plasma: FRAP) en comparación con el control, TR y L (125).
Se demostró así que los tomates, contengan o no licopeno, tienen un mayor potencial que el licopeno, para atenuar y/o revertir los parámetros relacionados con el estrés oxidativo en un contexto de leve estrés oxidativo.
Como la actividad biológica de los componentes fenólicos contribuye en gran medida a los efectos beneficiosos del tomate sobre la salud humana (26), se estudió el efecto de los compuestos fenólicos en tomates frescos y cocidos sobre la expresión de COX-2 en células KB respecto a la regulación de las respuestas inaamatorias inducidas por 12-O-tetradecanoílforbol-13-acetato (TPA) (126). Estos estudios celulares mostraron que los extractos fenólicos de los tomates cocidos causaron un aumento de la supresión de la expresión de COX-2 en comparación con los de tomate fresco. La fracción que contenía taninos no-condensados (rica en flavanos no-tanínicos) de tomates frescos suprimió significativamente la expresión de COX-2 en comparación con el control negativo, pero ambas fracciones conteniendo taninos no-condensados y taninos condensados (rica en taninos condensados y compuestos fenólicos simples) de tomates cocidos mostraron supresión en la expresión de COX- 2. Estos resultados sugieren que los compuestos fenólicos del tomate pueden desempeñar un importante rol en la quimioprevención del cáncer (126).
Se realizó un estudio en humanos para investigar si el jugo de tomate, rico en licopeno natural y enriquecido con vitamina C, es capaz de reducir varios biomarcadores del estrés oxidativo y la inflamación y si el efecto se puede atribuir al licopeno, la vitamina C u otro micronutriente.
Después de 2 semanas de depleción dietaria, los voluntarios fueron asignados aleatoriamente a ingerir jugo de tomate, ya fuese con (LC) o sin (L) la fortificación con vitamina C durante 2 semanas (dosis diaria de 20,6 mg de licopeno y 45,5/435 mg de vitamina C). Se analizaron plasma y orina para carotenoides y vitamina C, estado de lípidos, capacidad antioxidante, TBARS y 8-epi- PGF2a, carbonilos de proteínas, citoquinas IL-1b y TNFa y proteína C-reactiva (C-reactive protein: CRP) (127).
El consumo de jugo de tomate condujo a una disminución significativa de los niveles de colesterol total (L: 157,6 vs. 153,2 mg/dL; LC: 153,4 vs. 147,4 mg/dL) y la de la CRP (L: 315,6 vs. 262,3 μg/L; LC: 319,2 vs. 247,1 μg/L) en ambos grupos. El jugo fortificado con vitamina C elevó un poco la capacidad antioxidante en la orina y disminuyó TBARS en plasma y orina. Todos los demás marcadores se vieron afectados en menor medida o se mantuvieron sin cambios (127). La reducción del colesterol se correlacionó significativamente con la captación de licopeno, mientras que los otros efectos no pudieron ser relacionados con micronutrientes particulares. Es decir que, cualquier efecto beneficioso del consumo de tomate para la salud humana no puede atribuirse únicamente al licopeno y, como indica la suplementación adicional con ácido ascórbico, varios antioxidantes podrían ser necesarios para optimizar la protección contra las enfermedades crónicas.
Se estudió si la suplementación con alimentos funcionales ricos en antioxidantes, basada en tomates, podía actuar como adyuvante del interferón en la terapia de erradicación de virus de la hepatitis C (VHC), en particular, reducir la anemia durante la terapia con interferón pegilado (interferón-PEG) y ribavirina en la hepatitis crónica C (128).
El interferón-PEG es una nueva presentación farmacológica del interferón que se obtiene mediante la unión fisicoquímica de una molécula de interferón-a recombinante a una molécula de polietilenglicol. La conjugación de proteínas con polietilenglicol es un método bien conocido para retrasar su eliminación del organismo, prolongar y mantener su actividad. Por consiguiente, el compuesto resultante es muy estable y tras su administración subcutánea, se produce una liberación rápida del interferón, pero con eliminación lenta, de manera que se consiguen picos de concentración prolongados, durante al menos 90 horas, con una fase ulterior de eliminación gradual a lo largo de varios días. Entonces, la acción del interferón se mantiene de forma ininterrumpida durante un período de tiempo mucho más largo, obteniéndose un efecto terapéutico sostenido con una sola inyección semanal.
El estrés oxidativo juega un rol importante en la fisiopatología de la anemia hemolítica durante la terapia con ribavirina. La eficacia de la suplementación antioxidante con vitaminas C y E como compuestos puros, es aún controvertida. Se desarrolló un alimento funcional con un alto contenido de antioxidantes naturales y con alta biodisponibilidad de carotenoides. El estudio se hizo con 92 pacientes con hepatitis C crónica, tratados con terapia estándar de combinación. Cuarenta y seis de ellos recibieron 100 g/día del alimento funcional (grupo 1) y 46 no (grupo 2). El efecto de la actividad antioxidante se evaluó mediante la comparación del cumplimiento de la dosis completa de ribavirina y los niveles de hemoglobina durante los primeros 3 meses de tratamiento (128).
Como resultado, sólo 8,7% de los pacientes en el grupo 1 tuvo que reducir su dosis diaria de ribavirina, mientras que la reducción de ribavirina fue necesaria para el 30,4% de los pacientes en el grupo 2. Los niveles de hemoglobina mostraron diferencias significativas a los 15, 30 y 90 días durante el tiempo de observación (128). Los resultados demostraron que los alimentos funcionales reducen la gravedad de la anemia relacionada con ribavirina y mejoran la tolerancia a la dosis completa de ribavirina en pacientes con hepatitis C crónica.
Recientemente, se desarrolló un alimento basado en tomates para propósitos médicos especiales (food for special medical purposes: FSMP) y se evaluó su eficacia en pacientes con VHC en ensayos clínicos abiertos, aleatorios, en ocho voluntarios sanos y 39 pacientes con VHC, con FSMP, usado como tratamiento adyuvante a la terapia farmacológica con interferón-PEG y ribavirina (129). En el estudio de biodisponibilidad, los individuos sanos consumieron 100 g/día de FSMP durante una semana y se determinó su perfil basal de carotenoides en el suero, después de la semana de administración y 7 días después. La misma cantidad de FSMP fue consumida durante 6 meses por 20 de los 39 pacientes con VHC en el ensayo clínico. Las concentraciones de transaminasa, hemoglobina (Hb) e hidroperóxido en suero durante la terapia fueron controladas en todos los pacientes (129).
Como resultado, el consumo de FSMP provocó una cuadruplicación de la concentración sérica de licopeno en los individuos sanos. También se registró un aumento significativo de los carotenoides después de un mes de consumo en pacientes con VHC. FSMP no tuvo influencia en los niveles de transaminasas y de Hb en suero, así como en la respuesta terapéutica. La disminución en hidroperóxidos séricos fue independiente del consumo de FSMP en los pacientes que respondieron a largo plazo, mientras que los pacientes que no-respondieron (NR) del grupo FSMP mostraron una mayor reducción que los pacientes NR del grupo control (129).
En conclusión, FSMP fue efectivo en mejorar el status de carotenoides en individuos sanos. En pacientes con VHC, no influyó en la respuesta terapéutica, pero impidió el agotamiento de los carotenoides séricos y fue efectivo en mejorar el status oxidativo durante la terapia antiviral en los pacientes NR.
Recientemente se compilaron los resultados de los ensayos de intervención dietaria en humanos, que emplean el ensayo cometa (electroforesis monocelular en gel: SCGE), que se han publicado en los últimos años (es decir, entre 2005 y 2008) y se describieron las nuevas tendencias y desarrollos, así como los problemas actuales relacionados con el diseño de ensayos de intervención y con la interpretación de los resultados. La mayoría de los nuevos estudios se llevaron a cabo con alimentos complejos de origen vegetal y jugos; sólo unos pocos se realizaron con componentes individuales de los alimentos (130).
Con verduras específicas, por ejemplo: berro y repollitos de Bruselas, se observaron efectos potentes antioxidantes; también el café causó un efecto protector y es notable que fue más efectivo que el consumo de una dieta que contenía niveles elevados de frutas y hortalizas (130).
Resultan de interés los desarrollos recientes, así como el desarrollo de protocolos que permiten controlar la protección contra compuestos químicos genotóxicos contenidos en la dieta humana. Se demostró en estudios preliminares que la alteración de la actividad de las enzimas metabolizantes de drogas por factores dietarios, conduce al cambio de sensibilidad de los linfocitos contra el daño al ADN causado por determinados carcinógenos de la dieta (130).
El desarrollo de protocolos para los experimentos con células exfoliadas bucales es otra innovación potencialmente valiosa. El adecuado diseño experimental de los ensayos SCGE sigue siendo motivo de debate y la evaluación de los datos disponibles muestra que hay una necesidad urgente de desarrollar guías sobre el número de participantes, los períodos de muestreo, la duración de los ensayos, el uso de placebos y la definición de fases adecuadas de ejecución y de lavado (130).
Estudios recientes mostraron que los resultados de estudios dietarios podrían estar sesgados por factores tales como edad, sexo, índice de masa corporal y hábitos de estilos de vida y por efectos estacionales. Otro problema aún sin resolver es la interpretación de los resultados de los ensayos SCGE en lo que respecta a los efectos beneficiosos potenciales para la salud. El uso de técnicas -ómicas puede contribuir a proporcionar explicaciones mecanísticas, además de los enfoques convencionales (como mediciones de las enzimas) (130).
5. ACTIVIDAD INHIBITORIA DEL OXÍGENO SINGULETE
El licopeno es el más potente quencher de oxígeno singulete entre los carotenoides naturales (131) (132); exhibe la mayor constante de velocidad de quenching del oxígeno singulete (kq = 31 x 109 M-1s-1) y su nivel plasmático es ligeramente mayor que el de b-caroteno (kq = 14 x 109 M-1 s-1) (43). Esto es de gran interés general, ya que los carotenoides nutricionales, especialmente el b-caroteno y otros antioxidantes como a-tocoferol (kq = 0,3 x 109 M-1s-1) están implicados en la defensa contra los estados pro-oxidantes.
Se examinó la capacidad de quenching del oxígeno singulete de varios carotenoides naturales mediante la medición de la fotooxidación del ácido linoleico, sensibilizada con azul de toluidina, midiendo el consumo de oxígeno y la absorbancia ultravioleta a 235 nm (133). Se encontró que el quenching de oxígeno aumentó con el número de dobles enlaces conjugados en los carotenoides, pero el quenching varió con la estructura de la cadena y los grupos funcionales. Los carotenoides acíclicos, como el licopeno, aumentaron más el quenching que los carotenoides cíclicos. Los grupos ceto conjugados y la presencia de un anillo de ciclopentano estimularon el quenching, mientras que los grupos hidroxilo, epóxido y metoxilo mostraron menores efectos. Los carotenoides bacterianos fotosintéticos, espiriloxantina y rodopina, resultaron ser más eficaces como quenchers, seguidos por el carotenoide capsorbina (133).
Con el uso de un sistema in vitro en el cual el licopeno se une a la superficie de los linfocitos humanos, se demostró que el licopeno provee la mayor protección contra el daño celular producido por el oxígeno singulete (134).
6. COMUNICACIÓN INTERCELULAR POR UNIONES NEXUS
Para examinar la relevancia de la alteración de la comunicación intercelular por uniones nexus (gap-junctional intercellular communication, GJIC) en la actividad quimiopreventiva frente a la carcinogénesis (70), se estudiaron los efectos de licopeno, a- y b-caroteno, típicos carotenoides quimiopreventivos, sobre el acoplamiento celular vía las uniones nexus en el hígado de rata in vivo usando una técnica directa de transferencia funcional de pigmento (direct functional dye-transfer technique) (135). Se encontró que los tres compuestos ensayados en una dosis de 50 mg/kg de peso corporal por día, inhibieron GJIC, mientras que un tratamiento similar con 5 mg/kg de peso corporal causó un aumento de GJIC, especialmente en los grupos tratados con b-caroteno y licopeno. Con dosis de 0,5 mg/kg de peso corporal, los tres compuestos no tuvieron ningún efecto. Los resultados muestran que los tres carotenoides modulan diferencialmente GJIC dependiendo de la dosis, con efectos beneficiosos sobre la comunicación intercelular sólo detectados con una de las dosis. El resultado sugiere que la determinación de la dosis de productos químicos a ser usados es crucial para los estudios de intervención en humanos (135).
7. ACTIVIDAD ANTI-INFLAMATORIA DEL LICOPENO
La inflamación es una compleja serie de reacciones homeostáticas que involucra a los mecanismos inmunológicos humorales y celulares para proteger al organismo. Si esta reacción resulta exagerada o crónica, ocurren cambios patológicos. El fenómeno inflamatorio se desencadena por diferentes estímulos que inducen la liberación de mediadores proinflamatorios endógenos, que a su vez inducen la activación endotelial. El endotelio activado expresa de novo o incrementa la expresión de diferentes moléculas de adhesión, entre ellas la E-selectina (selectina E ó CD62E) e molécula de adhesión intercelular-1 (intercellular adhesion molecule 1: ICAM-1) (ó CD54) (136).
Las moléculas de adhesión son receptores funcionales que se expresan en la membrana celular y participan activamente en múltiples fenómenos fisiológicos y patológicos. Desempeñan un papel fundamental en la migración y activación de los leucocitos en la inmunovigilancia, en el desarrollo de la respuesta inflamatoria y de los mecanismos que intervienen en la respuesta inmune celular. La característica fundamental de estos receptores es la capacidad de transducir señales al interior de la célula y modular cascadas de señales inducidas por diferentes factores de crecimiento.
El licopeno ha demostrado, en estudios epidemiológicos, que previene las enfermedades cardiovasculares. Sin embargo, el mecanismo de su acción anti-inflamatoria sigue siendo poco claro.
Por ello, se estudiaron los efectos de una dieta rica en tomates sobre los marcadores de inflamación vascular. Las concentraciones plasmáticas de selectina-E, ICAM-1 y proteína C-reactiva de alta sensibilidad (high sensitivity C-reactive protein: hs-CRP) se determinaron por ELISA en 103 voluntarios aparentemente sanos. Los voluntarios fueron asignados al azar a dos grupos: 50 participantes comieron 300 g de tomates por día durante un mes y 53 participantes comieron su dieta habitual pero sin tomates durante ese período. Los marcadores de inflamación se midieron antes y después de un mes de la dieta asignada (137).
Los dos grupos dietarios presentaron características clínicas basales similares y niveles basales similares de los marcadores inflamatorios. Después de un mes de las dietas asignadas, se mantuvieron sin cambios las concentraciones de hs-CRP, selectina-E e ICAM-1 en comparación con la basal en la dieta rica en tomate. Sin embargo, la concentración de ICAM-1 se incrementó significativamente en el grupo dietario regular de 247,55±55 ng/mL a 264,71±60,42 ng/mL (137).
Estos resultados muestran que los mecanismos beneficiosos de la dieta (mediterránea) rica en tomates no están directamente relacionados con la inhibición de los marcadores de inflamación vascular. Sin embargo, recientemente se ha podido demostrar que el licopeno inhibe la expresión de ICAM-1 endotelial inducida por TNF-a y la adhesión endotelial de monocitos.
Los mediadores inflamatorios, como TNF-a e interleuquina (IL)-1b e IL-8, pueden aumentar la unión de LDL al endotelio y sobrerregular o desinhibir la expresión de las moléculas de adhesión de leucocitos sobre el endotelio durante la aterogénesis (138). Se estudió el efecto del licopeno sobre la señalización inducida por TNF-a en las células endoteliales umbilicales humanas (human umbilical endothelial cells: HUVECs). Se encontró que la expresión de ICAM-1inducida por TNF-a en HUVECs fue inhibida por el licopeno, mientras que no se afectó la expresión de COX-2 ni la expresión de la molécula de citoadhesión endotelial plaquetaria (platelet-endothelial cell adhesion molecule: PECAM-1 ó CD31). PECAM- 1 se expresa en la superficie de plaquetas, monocitos, neutrófilos, subpoblaciones de células T, uniones de las células endoteliales y en células NK, participando en la migración transendotelial de estas últimas.
Un nuevo análisis indicó que el licopeno atenuaba la fosforilación de IκB inducida por TNF-a, la expresión de NF-κB y la translocación de NF-κB p65 desde el citosol al núcleo. Además, el licopeno inhibió la formación de los complejos de NF-κB-DNA inducida por TNF-a, pero no la de AP1-ADN, según se determinó por el ensayo de desplazamiento de movilidad electroforética (electrophoretic mobility shift assay: EMSA) (138).
Por otra parte, el licopeno no afectó a p38 inducida por TNF-a ni la fosforilación de quinasa 1/2 extracelular regulada por la matriz (extracellular matrix-regulated kinase1/2: ERK1/2) y la señalización inducida por interferón-γ (IFN-γ), lo que sugiere que el licopeno afecta principalmente a la vía de señalización de NF-κB inducida por TNF-a. En un estudio funcional, el licopeno, de manera dependiente de la dosis, atenuó la adhesión de los monocitos a la monocapa endotelial, pero no esa adhesión a la matriz extracelular (138).
En conjunto, ésta es la primera evidencia que demuestra que el licopeno es capaz de inhibir la activación de NF-κB inducida por TNF-a, la expresión de ICAM-1 y la interacción endotelial de los monocitos, lo que sugiere un rol anti-inflamatorio del licopeno y, posiblemente, explica en parte porqué el licopeno puede prevenir enfermedades cardiovasculares.
8. VIAS DE SEÑALIZACION: DESREGULACIÓN DE LA VÍA AKT EN CÁNCER HUMANO
Las mutaciones en la posición q23 del cromosoma 10 se encuentran entre las más comunes observadas en cáncer humano y, particularmente prevalentes en glioblastomas, cáncer prostático avanzado y carcinoma endometrial. En 1997, dos grupos publicaron el descubrimiento de un gen supresor de tumores en este locus, al que llamaron PTEN (phosphatase and tensin homolog deleted on chromosome ten) o MMAC (mutated in multiple advanced cancers) (139). PTEN posee la capacidad de desfosforilar PIP3 (fosfatidilinositol- 3,4,5-trifosfato: PI(3,4,5)P3]. Estas moléculas son segundos mensajeros cruciales en los caminos de señalización que controlan la proliferación y la supervivencia celular. PTEN actúa como una fosfatasa lipídica in vivo.
Los fosfoinosítidos PI(3,4,5)P3 y PI(3,4)P2 se generan en respuesta a diversos estímulos, incluyendo factores de crecimiento y supervivencia, luego de la activación de la fosfatidilinositol 3-quinasa (phosphatidylinositol 3-kinase: PI3K) en la membrana plasmática.
La interacción con PIP3 o PIP2 sirve para reclutar proteínas, tales como PKB/Akt hacia la membrana, donde se activan y transducen señales promotoras de supervivencia y proliferación. Es importante hacer notar que estas señales dependientes de fosfoinosítidos, son controladas continuamente mediante la acción de PTEN, que los desfosforila en la posición 3 del anillo inositol.
Como se ve, las proteínas Akt están implicadas en muchos procesos celulares, como el metabolismo, la muerte celular programada y la proliferación. Akt (proteín- quinasa B) es una serín-/treonín-quinasa, que es un regulador central de los procesos celulares ampliamente divergentes, como proliferación, diferenciación, migración, supervivencia y metabolismo. Akt es activada por una variedad de estímulos, a través de los receptores de factor de crecimiento, de una manera dependiente de PI3K. Akt también es regulada negativamente por la fosfatasa supresora del tumor PTEN.
El producto del gen supresor de tumores (PTEN) actúa como una fosfatasa de fosfolípidos: a) En condiciones de crecimiento normal, las señales estimulatorias que salen del receptor para insulina, activan a la enzima PI3K, la que fosforila a fosfatidilinositol 4,5-bisfosfato (PIP2) para generar PIP3, una molécula lipídica de señalización. Corriente abajo, PIP3 activa diversos efectores, incluyendo al producto del protoconcogen PKB/Akt. El rol de PTEN es desfosforilar a PIP3, actuando como un control negativo en la activación de PKB/Akt. b) Si una mutación en PTEN le impide realizar la desfosforilación, PIP3 no podrá ser inactivado y, por consiguiente, continuará propagando su señal corriente abajo. Esto podría resultar en la activación continuada de PKB/Akt, que, en combinación con otros factores, podría llevar a un crecimiento celular aumentado y un posible desarrollo de un tumor.
Es decir que, generalmente, se produce una alteración de la señalización normal de Akt/PKB/PTEN en muchos cánceres humanos, lo cual juega un rol importante en el desarrollo, progresión y resistencia terapéutica del cáncer.
Numerosos estudios han puesto de manifiesto que el bloqueo de la señalización de Akt causa apoptosis e inhibición del crecimiento de las células tumorales (140). Por lo tanto, esta vía de señalización, tanto corriente arriba como corriente abajo de Akt, recientemente ha atraído considerable atención como un nuevo blanco para las estrategias terapéuticas efectivas del cáncer (141).
La regulación aberrante de la ruta de señalización de supervivencia fosfoinosítido 3-quinasa/Akt en el cáncer ha llevado a un interés en la supresión de esta vía para tratar el cáncer. Estudios previos identificaron un rol importante para fosfoinosítido 3-quinasa/Akt en la progresión de cáncer de colon.
Recientemente, se investigó el efecto inhibitorio del licopeno en la vía de señalización Akt en las células HT- 29 de cáncer de colon humano. El licopeno inhibió la proliferación celular en las células HT-29 de cáncer de colon humano con un valor IC50 de 10 μM. El tratamiento con licopeno suprimió la activación de Akt y el nivel de proteína b-catenina no fosforilada en las células humanas de cáncer de colon (142).
9. INDUCCION DE APOPTOSIS
Si bien la evidencia epidemiológica demuestra que una ingesta alta en frutas y vegetales ricos en carotenoides está asociada con un menor riesgo de cáncer, los resultados de ensayos de intervención indican que los suplementos de b-caroteno acentúan el riesgo de desarrollar incidencia y mortalidad por cáncer de pulmón entre fumadores. Un posible mecanismo que puede explicar el doble rol de los carotenoides como agentes beneficiosos y perjudiciales en el cáncer es que su exceso o deficiencia puede generar cambios en las vías moleculares implicadas en la señalización apoptótica.
Se examinó la potencia de varios carotenoides, que se encuentran comúnmente en el plasma humano, para inducir apoptosis en las células T-linfoblastos Jurkat E6.1 malignas. A una concentración de 20 μM, el orden de potencia para inducir la apoptosis después de 24 horas fue de: b-caroteno > licopeno > luteína > b-criptoxantina = zeaxantina. La cantaxantina falló en inducir apoptosis en estas condiciones. b-caroteno indujo apoptosis en una forma dependiente del tiempo y de la concentración, con una concentración efectiva más baja, de alrededor de 3 μM (143). El pre-acondicionamiento de b-caroteno durante 72 horas destruyó su actividad pro-apoptótica casi completamente, mientras que la degradación durante 6 horas o menos, no la destruyó, indicando que el b-caroteno mismo, y/o un producto temprano de la degradación de b-caroteno, son los compuestos que inducen la muerte (143).
La apoptosis inducida por b-caroteno se caracterizó por la condensación de cromatina y la fragmentación nuclear, la degradación de ADN, el clivaje de PARP y la activación de la caspasa-3.
El antioxidante BO-653 inhibió la degradación de b- caroteno in vitro e incrementó significativamente su citotoxicidad, indicando que no es probable que un efecto pro-oxidante del b-caroteno cause su actividad pro-apoptótica (143).
Recientemente se examinó el efecto anti-proliferativo y apoptótico del licopeno sobre otras líneas de células malignas. Se incubaron las células de las siguientes líneas con tres concentraciones de licopeno: 1,0, 2,0 y 4,0 μM: carcinoma de colon humano (human colon carcinoma: HuCC), leucemia linfocítica crónica B (B chronic lymphocytic leukemia: EHEB), eritroleucemia humana (human erythroleukemia: K562) y Raji, un prototipo de la línea celular de linfoma de Burkitt (78).
Los resultados mostraron que el licopeno ejerció un efecto significativo, dependiente de la dosis, sobre la capacidad de proliferación de las líneas K562, Raji y HuCC, mientras que se observó este efecto en las células EHEB sólo con la dosis más alta utilizada en el estudio (78).
Se encontró la mayor tasa de apoptosis después de la incubación de las células HuCC con licopeno 2,0 y 4,0 μM y en las células Raji tras la incubación con licopeno 2,0 μM (78). En conclusión, el efecto anti-proliferativo del licopeno sobre las células tumorales y su efecto sobre la tasa de apoptosis depende de la dosis y del tipo de células malignas.
10. EFECTO INMUNOMODULADOR DEL LICOPENO
Hace varios años se informó que el licopeno suprimió significativamente el desarrollo de tumores mamarios espontáneos en ratones y se sugirió la contribución del licopeno al mantenimiento de la homeostasis física. Se investigaron los efectos del licopeno en la subpoblación de células T intratímicas y periféricas como parámetros inmunorreguladores. En los ratones control con tumor, se observó: la inactivación de la diferenciación de las células T intratímicas, un aumento del porcentaje de las células CD4+ CD8+ y una disminución de las células CD4+CD8+ (144).
Por otra parte, en los ratones tratados con licopeno, se recuperó la diferenciación anormal de células T intratímicas, llevando a niveles no tumorales. La relación CD4/CD8 periférica aumentó parcialmente por el tratamiento con licopeno, lo cual resultó en un aumento de la subpoblación CD4+ (144).
Estas observaciones sugieren que uno de los roles inmunomoduladores del licopeno es normalizar el cambio de la diferenciación de células T intratímicas causada por tumores.
Hay evidencias que indican que el consumo regular de productos del tomate está asociado con efectos inmunomoduladores favorables. Además, se ha demostrado que los extractos de tomate poseen actividad in vitro antioxidante, anticancerígena y antitrombótica. Por eso, se examinó el efecto in vitro del licopeno en la producción de citoquinas por células mononucleares de sangre periférica (peripheral blood mononuclear cells: PBMC) en 15 individuos saludables (145).
En primer lugar, 2 x 106 PBMC suspendidas en 1 mL de medio condicionado se incubaron durante 24 y 48 h, sin o con las siguientes concentraciones de licopeno: 0,25; 0,50; 1,00; 2,00 y 4,00 μM. Se evaluó la producción de las citoquinas siguientes: IL-1b, IL-1ra, IL-2, IL-6 e IL- 10, así como TNF-a e IFN- γ (145).
El licopeno indujo un aumento dependiente de la dosis en la producción de IL1b y TNF-a y una disminución en la secreción de IL-2, IL-10 e IFN-γ, mientras que la de IL-6 e IL-1ra no se vio afectada (145). Es decir que la comprensión del rol del licopeno en la modulación del sistema inmune puede promover decisiones sobre la suplementación dietaria con licopeno para la reducción del riesgo de determinadas enfermedades.
11. LICOPENO: ACTIVIDAD ANTI-ATEROGÉNICA Y ENFERMEDADES VASCULARES
El proceso aterogénico se desarrolla en varias enfermedades, entre ellas las enfermedades autoinmunes (146). Varias evidencias sugieren que la lipoproteína de baja densidad (LDL) modificada oxidativamente es aterogénica y que, por consiguiente, la aterosclerosis puede ser atenuada con antioxidantes naturales, que inhiben la oxidación de LDL (116).
Se estudió el efecto del licopeno del tomate, solo o en combinación con otros antioxidantes naturales, sobre la oxidación de LDL. Para ello, LDL (100 μg de proteína/mL) se incubó con concentraciones en aumento de licopeno o de óleo-resina del tomate (extracto lipídico de los tomates que contiene licopeno 6%, b-caroteno 0,1%, vitamina E 1% y polifenoles), después de lo cual fue oxidado mediante la adición de CuSO4 5 μM. La óleo-resina del tomate exhibió una capacidad superior para inhibir la oxidación de LDL que el licopeno puro, por hasta cinco veces [97% vs. 22% de inhibición de formación de sustancias reactivas al ácido tiobarbitúrico (TBARS) y 93% vs. 27% de inhibición de formación de peróxidos lipídicos respectivamente]. Debido a que la óleo-resina del tomate contiene, además de licopeno, vitamina E, flavonoides y compuestos fenólicos, se ha estudiado una posible interacción cooperativa entre el licopeno y tales antioxidantes naturales. Una combinación de licopeno 5 μM con vitamina E (a-tocoferol) en el rango de concentración de 1-10 μM dio lugar a una inhibición de la oxidación de LDL inducida por iones cobre que fue significativamente mayor que las inhibiciones esperadas para cada compuesto individual (116).
El efecto antioxidante sinérgico del licopeno con la vitamina E no fue compartido por γ-tocotrienol. Cuando se agregó licopeno 5 μM a LDL, en combinación con glabridina, ácido rosmarínico, ácido carnósico, o ajo, se obtuvieron efectos antioxidantes sinergísticos contra la oxidación de LDL, inducida por los iones de cobre o por el generador de radicales AAPH (116).
También se observaron efectos similares interactivos con licopeno y b-caroteno, pero con menor sinergismo. Se estudió la relevancia in vivo del licopeno combinado con otros antioxidantes naturales. Para ello, se administró a cuatro individuos sanos una comida grasa que contenía 30 mg de licopeno en la forma de óleo-resina de tomate. Se elevó la concentración de licopeno en el plasma postprandial en un 70% en comparación con el plasma obtenido antes del consumo de comida. La LDL postprandial aislada 5 horas después del consumo de alimento mostró una susceptibilidad significativamente menor a la oxidación en un 21% (116).
Se llegó a la conclusión que el licopeno actúa sinérgicamente, como un antioxidante efectivo contra la oxidación de LDL, en combinación con varios antioxidantes naturales como la vitamina E, el flavonoide glabridina, los compuestos fenólicos ácido rosmarínico y ácido carnósico, y el ajo. Estas observaciones sugieren una característica antiaterogénica superior para la combinación de diferentes antioxidantes naturales que la de cada uno individualmente.
Se sabe que la dieta desempeña un rol complejo en el desarrollo de enfermedades cardiovasculares, principal causa de muerte en el mundo occidental. Pero el procesado de los tomates puede afectar significativamente la biodisponibilidad de sus nutrientes. La homogenización, el tratamiento térmico y la incorporación de aceite en los productos basados en tomate dan lugar a una mayor biodisponibilidad del licopeno, mientras que algunos de los mismos procesos causan una pérdida significativa de otros nutrientes. El contenido de nutrientes también se ve afectado por la variedad y el grado de madurez (147).
Muchos de estos nutrientes pueden funcionar individualmente o en conjunto, para proteger a las lipoproteínas y a las células vasculares de la oxidación. Esta hipótesis ha sido corroborada por estudios in vitro, algunos in vivo y muchos epidemiológicos que asocian la reducción del riesgo cardiovascular con el consumo de alimentos ricos en antioxidantes.
Otras funciones cardioprotectoras provistas por los nutrientes de los tomates pueden incluir la reducción del colesterol-LDL, de la homocisteína, de la agregación plaquetaria y de la presión arterial (147).
El licopeno inhibió, en un estudio in vitro y en un pequeño estudio clínico, la actividad de una enzima esencial, que participa en la síntesis de colesterol, lo cual sugiere un efecto hipocolesterolémico (148).
Sabiendo que los tomates y el licopeno son considerados potentes antioxidantes, que reducirían la mortalidad y la morbilidad cardiovascular considerablemente, se estudiaron los efectos de una dieta rica en tomates (300 g de tomates al día durante un mes) sobre el perfil lipídico (149).
Se encontró que la dieta rica en tomates (300 g al día durante un mes) aumentó el nivel de colesterol-HDL significativamente en un 15,2% (149).
Se investigó la tasa de peroxidación lipídica mediante la estimación de los niveles de malondialdehído (TBARS) de las enzimas antioxidantes, como SOD, GSH-Px, GR, GSH, perfil lipídico, que incluye el colesterol total, triglicéridos, HDL, LDL, lipoproteína de muy baja densidad y la hemoglobina glicosilada (HbA1c) en un grupo de diabéticos tipo 2 (n = 40) y un grupo control (n = 50) (150).
Se observaron niveles significativamente más bajos de las enzimas antioxidantes y una tasa muy alta de peroxidación lipídica en el grupo de diabéticos tipo 2, cuando se compararon con los controles. Asimismo, se observaron niveles significativamente más altos de perfil lipídico y HbA1c en el grupo diabético en comparación con el grupo control.
Los resultados de estudios observacionales sugieren que el estrés oxidativo y el status hiperlipidémico, que prevalecen en la hipertensión, desempeñan un rol importante en la causalidad de las complicaciones secundarias. Por lo tanto, se midió la tasa de peroxidación lipídica por medio de la medición de malondialdehído (MDA) y se estimó la actividad de las enzimas plasmáticas antioxidantes SOD, GSH-Px, GR, GSH y el perfil de lípidos en suero, que incluye el colesterol total y los triglicéridos en un grupo de hipertensos de grado I (n = 40) y un grupo control (n = 50). Después de 60 días de administración de tomates (cocidos), el grupo de hipertensos (n = 30) mostró una mejoría significativa en los niveles séricos de las enzimas antioxidantes y una disminución de la tasa de peroxidación lipídica, pero no hubo cambios significativos en el perfil lipídico (151).
En conclusión, el licopeno del tomate puede tener considerable potencial terapéutico natural como antioxidante, pero no puede ser utilizado como agente hipolipidémico en la hipertensión de grado I.
Para contribuir a establecer los mecanismos ateroprotectores involucrados en el menor riesgo de enfermedades cardiovasculares por ingesta de tomate, se investigaron recientemente los efectos sobre los lípidos plasmáticos y la oxidación de LDL provocados por un aumento de productos de tomate en la ingesta alimentaria. Para ello, en un estudio de intervención dietaria con 21 individuos sanos, se incluyó un período basal, uno de 3 semanas de dieta sin productos del tomate y una dieta de 3 semanas alta en tomate (400 mL de jugo de tomate y 30 mg de ketchup por día). La concentración de colesterol total se redujo significativamente en 5,9% y la concentración de colesterol-LDL en 12,9% con una dieta alta en tomate en comparación con la dieta baja en tomate (152).
En conclusión, una dieta alta en productos de tomate produjo efectos ateroprotectores, se redujeron significativamente los niveles de colesterol- LDL y aumentó la resistencia de LDL a la oxidación en adultos sanos normocolesterolémicos. Estas características ateroprotectoras se asociaron con los cambios en los niveles séricos de licopeno, b-caroteno y γ-caroteno (152).
También se estudiaron los efectos de los compuestos fenólicos de los tomates sobre la regulación de los lípidos en sangre. En un ensayo clínico en humanos se examinaron: la antioxidación en plasma, el status de los lípidos sanguíneos y las respuestas fenólicas después de la ingestión de tomates frescos, jugo de tomate y bebida con licopeno. Se usaron tomates frescos de distinto tamaño, después de blanquearlos o después del blanqueo y calentamiento. Los contenidos de compuestos fenólicos del tomate se incrementaron en un 34% en los tomates pequeños y en un 23% en las tomates grandes después del tratamiento de blanqueo y calentamiento a 100 °C durante 30 min. Los compuestos fenólicos mostraron una buena actividad antioxidante (57-71%) y también promovieron sinérgicamente la antioxidación (81-100%) de los carotenoides del tomate. En el estudio clínico, la capacidad antioxidante total y el contenido de compuestos fenólicos en el plasma se incrementaron después de la administración de tomate fresco y jugo de tomate, pero no se observó diferencia significativa en el consumo de bebida con licopeno. Los niveles de triglicéridos y de colesterol-LDL se redujeron después de la administración de tomate fresco y jugo de tomate y se incrementó el colesterol-HDL (153).
Para investigar los mecanismos moleculares de la pasta de tomate, que contiene aproximadamente un 0,1% de licopeno, en la regulación del metabolismo de los lípidos y de la oxidación, recientemente, se alimentaron hámsters con un 3% ó 9% de pasta de tomate, que contenía 0,2% de colesterol, y se midieron: colesterol total (CT), LDL, HDL y triglicéridos (TG). Los resultados mostraron menores tasas de CT sérico y niveles de LDL, debido a la ingestión de 9% de pasta de tomate, siendo de 14,3% y 11,3% respectivamente. Las concentraciones de 3% y el 9% de pasta de tomate después de 8 semanas de alimentación aumentaron significativamente los niveles séricos de HDL, en un 19,4% y 28,8% respectivamente. Después de la ingestión de pasta de tomate durante 8 semanas, los niveles de malondialdehído (MDA) en plasma disminuyeron significativamente, en un 80,18% y 89,33% respectivamente, indicando la actividad antioxidante potente de la pasta de tomate, conjuntamente con el aumento de las actividades de las enzimas SOD, CAT y GSH-Px (154).
El análisis por electroforesis bidimensional en gel (2- DE) reveló que la anhidrasa carbónica III (carbonic anhydrase III: CAIII) y la adenilato-quinasa 2 (AK2) pueden ser dos reguladores importantes implicados en los efectos antilipídico y antioxidante de la pasta de tomate, abriendo nuevos puntos de vista en el valor nutricional del tomate en la promoción de la salud pública (154). Con este trabajo se caracterizaron los efectos hipolipemiantes y los mecanismos antioxidantes de la pasta de tomate.
Las enfermedades cardiovasculares resultan de la acumulación sub-endotelial de células inflamatorias y de células de músculo liso. Se ha sugerido que el licopeno desempeña un rol en la prevención de las enfermedades cardiovasculares, pero su mecanismo de acción aún sigue siendo en gran medida desconocido. Se ha examinado su efecto sobre las células de músculo liso. Se encontró que la preincubación de PDGF-BB con licopeno causó una marcada inhibición del receptor-b PDGF inducido por PDGF-BB (PDGFR-b), PLCγ y en la fosforilación de ERK1/2 en células de músculo liso de rata A10 y células de músculo liso aórticas primarias cultivadas. En marcado contraste, el licopeno no influyó en la fosforilación de ERK1/2 inducida por EGF. Sorprendentemente, análisis adicionales indican que el licopeno podría unirse directamente a PDGF-BB e inhibir la interacción PDGF-BB-células de músculo liso (155).
En los estudios funcionales, el licopeno inhibió la proliferación y la migración de células de músculo liso, inducidas por PDGF-BB, en concentraciones de 2 a 10 μM. Por el contrario, el licopeno no inhibió la migración de las células endoteliales inducida por bFGF y VEGF. La zimografía en gelatina demostró que el efecto del licopeno sobre la migración de células de músculo liso no se debía a la inhibición de las metaloproteinasas de la matriz. Estos resultados, en conjunto, proporcionan la primera evidencia que demuestra que el licopeno inhibe la señalización de proliferación y migración inducidas por PDGF-BB, en ratas A10 y células de músculo liso aórticas (155). Uno de los mecanismos de acción es aquel mediante el cual el licopeno es capaz de unirse a PDGF-BB e inhibir su interacción con células de músculo liso, que es muy diferente a los antagonistas PDGFR-b anteriormente desarrollados. Estos resultados ayudan a comprender mejor los efectos beneficiosos del licopeno en la prevención de las enfermedades cardiovasculares.
Por el contrario, el licopeno no resultó efectivo como agente hipolipidémico en la cardiopatía coronaria (156). Esto surgió de investigar la tasa de peroxidación lipídica por medio de la estimación de MDA, los niveles de las enzimas séricas antioxidantes SOD, GSH-Px, glutatión-reductasa, glutatión reducido y perfil lipídico, que incluye el colesterol total, triglicéridos, HDL, LDL y lipoproteína de muy baja densidad en un grupo de pacientes con cardiopatía coronaria y un grupo control. Después de 60 días de administración de suplementos de tomate al grupo de pacientes cardíacos, se observó una mejoría significativa en los niveles séricos de las enzimas antioxidantes y disminuyó significativamente la tasa de peroxidación lipídica, pero no hubo cambios significativos en el perfil lipídico (156). Estos hallazgos sugieren que el licopeno del tomate puede tener considerable potencial terapéutico como antioxidante, pero no puede ser utilizado como agente hipolipidémico en la cardiopatía coronaria.
12. EFECTO ANTI-ASMA
Las dietas ricas en antioxidantes también están asociadas con una reducción de la prevalencia del asma. Recientemente, se investigaron los cambios en el asma y en la inflamación de las vías respiratorias como consecuencia de una dieta baja en antioxidantes y la subsiguiente utilización de tratamientos ricos en licopeno. Para ello, adultos asmáticos (n = 32) consumieron una dieta baja en antioxidantes durante 10 días y luego se inició un estudio aleatorizado, transversal, involucrando 3 x 7 días de tratamiento [placebo, extracto de tomate (45 mg de licopeno/día) y jugo de tomate (45 mg licopeno/día)]. Con el consumo de una dieta baja en antioxidantes, disminuyeron las concentraciones plasmáticas de carotenoides, empeoró el Score Control del Asma, disminuyeron %FEV(1) y %FVC y aumentó el porcentaje de neutrófilos en el esputo (157).
El tratamiento con extracto de jugo y extracto de tomate redujo el inaujo de neutrófilos en las vías respiratorias y también redujo la actividad de elastasa de neutrófilos en el esputo. En conclusión, los tratamientos ricos en licopeno modifican la inflamación no-eosinofílica de las vías respiratorias en el asma. El consumo de antioxidantes dietarios modifica los resultados clínicos del asma. Menor ingesta de antioxidantes dietarios contribuye al aumento de prevalencia del asma. Los suplementos ricos en licopeno deberían investigarse más a fondo como una intervención terapéutica (157).
13. ANTI-CANDIDIASIS
Recientemente se investigó la actividad antifúngica del licopeno y su modo de acción. El licopeno mostró potentes efectos antifúngicos en hongos patógenos, ensayados de una manera independiente de la energía, con bajos efectos hemolíticos contra los eritrocitos humanos (158).
Para confirmar esta actividad del licopeno, se investigaron sus efectos sobre el dimorfismo de Candida albicans inducido por suero bovino fetal, que desempeña un rol clave en la patogénesis de una invasión al huésped. El licopeno ejerció una potente actividad antifúngica sobre los micelios de C. albicans inducidos en el suero. Para entender el modo de acción del licopeno, se examinó su acción contra las membranas celulares fúngicas mediante el análisis FACS y la prueba de liberación de glucosa y trehalosa (158). El licopeno causó daños significativos en la membrana citoplasmática y muerte celular en Candida albicans. Es decir que el licopeno tiene una considerable actividad antifúngica, que merece una mayor investigación para aplicaciones clínicas.
14. LICOPENO CONTRA EL CANCER
Se ha visto que el licopeno presenta una serie de actividades, pero se hizo popular igual que el tomate por disminuir el riesgo de cáncer de próstata (159) y por sus efectos quimiopreventivos en algunos tipos de cáncer, avalado por estudios epidemiológicos (8). Una revisión sistemática de 72 estudios epidemiológicos informó una relación inversa consistente entre la ingesta de tomates o los niveles plasmáticos de licopeno y el cáncer de próstata, pulmón y estómago (84).
El licopeno se caracteriza por presentar actividad anticancerígena en glándula mamaria, hígado, piel y pulmón en modelos de ratón y también por inhibir el desarrollo de focos crípticos aberrantes de colon en ratas (91). Ningún estudio indicó que un mayor consumo de tomate o niveles de licopeno en sangre aumentará significativamente el riesgo de cáncer (160).
14.1. Cáncer de próstata
La próstata, la mayor glándula accesoria masculina, es una fuente potencial de graves desórdenes que afectan la salud y la calidad de vida de los hombres mayores. El tamaño de dicha glándula es similar al de una nuez que envuelve a la uretra que está justo adyacente a la vejiga y otras condiciones patológicas pueden provocar la obstrucción en el flujo de orina con el consiguiente daño a la vejiga y a los riñones.
Los desórdenes prostáticos incluyen inflamación (prostatitis), hiperplasia prostática benigna (HPB) y cáncer de próstata. Mientras que la prostatitis crónica puede afectar a cualquier hombre adulto a la edad de setenta años, más del 40% de los hombres desarrollan un crecimiento del tejido prostático debido a una HPB y, al alcanzar la edad de ochenta años, muchos padecerán cáncer de próstata.
El tratamiento de cáncer involucra radiación, varios procedimientos quirúrgicos, terapia hormonal anti-androgénica con posibles efectos colaterales de incontinencia urinaria e impotencia. Estas perspectivas provocan, tanto en los pacientes como en los médicos, el esfuerzo en desarrollar estrategias de "espera cautelosa", ya que el cáncer de próstata es de lento crecimiento, especialmente en hombres de mayor edad (161).
La incidencia del cáncer de próstata varía considerablemente entre regiones geográficas, con elevadas tasas localizadas en los Estados Unidos y Escandinavia y con menor incidencia en China y otros países de Asia. Estas diferencias son causadas fundamentalmente por la susceptibilidad genética, la exposición a factores ambientales (como la dieta) o a una combinación de las mismas (162) (163).
En los países desarrollados, el cáncer de próstata se encuentra en el segundo puesto entre los diagnósticos de cáncer de mayor frecuencia, y a su vez, es la tercera causa más común de muerte por cáncer en los hombres occidentales. Los hombres afroamericanos se encuentran entre los grupos étnicos que presentan las tasas más altas de cáncer de próstata (0,275%) en el mundo, siendo mayores a las que presentan los caucásicos (0,173%) y finalmente los de origen hispano (0,1276%) o asiático (164).
Con el advenimiento del ensayo PSA (del inglés, prostatic specific antigen), el porcentaje de pacientes con la enfermedad ha disminuído de manera notable (165).
Además del rol de la dieta en disminuir la progresión del tumor, también se han usado agentes hormonales como los estrógenos y los inhibidores de la 5-a- reductasa como agentes preventivos terciarios (166) (167).
Clínicamente, su diagnóstico puede ser local o avanzado y el tratamiento a aplicar puede consistir en una vigilancia cuidadosa del paciente hasta un radical tratamiento local o un tratamiento de privación androgénica. Este último tratamiento reduce los síntomas de cáncer de próstata en aproximadamente un 70% a 80%, principalmente en los pacientes que presentan un estadío avanzado de la enfermedad. Pero muchos tumores vuelven a generarse en un lapso de dos años, transformándose en un estado independiente de los andrógenos (hormone refractory prostate cancer: HRPC), que si bien ha sido incurable hasta hace unos años, recientemente ha comenzado a ser el blanco terapéutico de drogas en ensayos clínicos de fase II y III (168).
La disparidad existente entre la incidencia y la tasa de mortalidad puede hacer inferir que sólo una pequeña proporción de los diagnósticos de bajo riesgo de cáncer de próstata puede conducir a una enfermedad que amenaza la vida del paciente (169).
Dependiendo del diseño del estudio, gran número de individuos deben ser inscriptos en el ensayo de quimio-prevención y se requieren largos intervalos de seguimiento, por lo que estos ensayos son de gran complejidad y de costo elevado (160).
A pesar de intensos esfuerzos, poco se sabe acerca de la etiología del cáncer de próstata y el tratamiento de las formas avanzadas de la enfermedad ha tenido un éxito limitado. Sin embargo, los estudios epidemiológicos combinados con modelos animales y experimentos in vitro indican que la dieta y la actividad física pueden desempeñar roles importantes, tanto en la incidencia como en la progresión de la enfermedad (170). La obesidad puede aumentar el riesgo primario y bioquímico (aumento de PSA) o la recurrencia clínica. Entre los grupos individuales de alimentos/nutrientes un alto consumo de grasa total, grasas saturadas, carne, productos lácteos y calcio se relacionan con un mayor riesgo. Los productos del tomates, soja, licopeno, selenio, ácidos grasos ω-3 marinos y vitamina E en fumadores pueden estar inversamente asociados con cáncer de próstata (171) y servir como agentes quimiopreventivos (172) (173).
La epidemiología descriptiva del cáncer de próstata sugiere que es una enfermedad previsible. La prevención tiene la ventaja teórica de no sólo salvar vidas, sino también reducir la morbilidad de la terapia radical del cáncer de próstata. Recientemente, se llevó a cabo una compilación sobre la prevención del cáncer de próstata y las estrategias utilizadas, como la reducción de las grasas dietarias y la suplementación con vitaminas D y E, y selenio (174). Se analizó la ingesta dietaria de soja, té verde, tomate y productos ricos en tomate (licopeno), así como los datos relativos a la intervención farmacológica con inhibidores de la ciclooxigenasa, antiestrógenos y, en particular, inhibidores de la 5a-reductasa (174). Por último, se presentaron varios diseños de ensayos, así como ensayos actuales de fase 2. Las respuestas de`nitivas a los ensayos pivotales de fase 3 estarán disponibles en los próximos 2 a 7 años (174).
La ingestión de licopeno (principalmente en productos de tomate) se asoció con un menor riesgo de desarrollo de cáncer de próstata, lo cual está sustentado por estudios in vivo e in vitro que muestran que el licopeno incorporado oralmente está biodisponible, acumulándose en el tejido prostático y específicamente, en el núcleo de las células epiteliales de la próstata (175) (176). El efecto inhibitorio de una dieta con licopeno no se observó en modelos de carcinogénesis de próstata en ratas (177).
La adenosíndesaminasa (ADA) es una enzima del grupo de las hidrolasas, involucrada en el catabolismo de las bases purínicas. Es un marcador de inmunidad celular, cuya actividad plasmática está aumentada en enfermedades donde la respuesta inmune tiene una mediación celular. Se sabe que ciertas sustancias que poseen potenciales características inhibidoras de ADA pueden desempeñar un rol muy importante en la represión y/o involución de tumores. Se estudiaron los efectos del jugo de tomate sobre la actividad de ADA utilizando tejido prostático de diez pacientes con probada patología de cáncer de próstata. Se observó un importante efecto inhibitorio de la actividad de ADA, con y sin preincubación, a medida que los tejidos eran sometidos a concentraciones crecientes de jugo de tomate (Tabla IV).
Tabla IV. Concentración del jugo de tomate y actividad de ADA (73).

Otros componentes pueden también contribuir a la prevención del cáncer. Varias observaciones apuntan al procesado del tomate como una posible fuente de contribuyentes, aún no identificados, para la quimioprevención del cáncer. En primer lugar, durante el procesado, el contenido de licopeno, ascorbato y otros antioxidantes naturales disminuye como resultado de la degradación térmica y de la isomerización (178) (179). Sin embargo, la actividad antioxidante total del jugo de tomate procesado térmicamente o de otros productos, puede no disminuir y, de hecho, a menudo aumenta (30) (178) (179). En segundo lugar, el consumo de productos procesados de tomate, en lugar de tomates frescos, se asoció epidemiológicamente con una disminución en el riesgo de cáncer de próstata (176). En tercer lugar, el consumo de tomate en polvo inhibió la carcinogénesis de próstata en ratas (180), lo que sugiere que, además del licopeno, los productos del tomate contienen otros compuestos que incrementan la supervivencia de los animales de experimentación. La mayoría de los ensayos clínicos con productos del tomate también sugieren una acción sinérgica del licopeno con otros nutrientes (38) (181).
Durante la elaboración de alimentos, como es sabido, se producen varias reacciones químicas, entre ellas la reacción de Maillard, que es principalmente responsable del pardeamiento no enzimático y sabor específicos de los alimentos horneados, asados y secos (182). En la etapa inicial de la reacción de Maillard se produce la condensación de un azúcar reductor, como la glucosa, con los grupos amino de los aminoácidos libres o de las proteínas, para formar glucosilaminas inestables, también conocidas como bases de Schiff. Los ácidos orgánicos débiles catalizan la transformación de las glucosilaminas, a través del reordenamiento de Amadori, en cetosaminas (Fig. 8) más estables que se conocen también como productos de reordenamiento de Amadori. Los frutos secos y hortalizas, como el tomate en polvo procesado, contienen cantidades significativas de cetosaminas, principalmente en forma de conjugados no-digeribles de fructosa- aminoácidos, que son parcialmente absorbidos en la circulación, pero son excretados sin cambios (38).

Figura 8. Estructura química de una cetosamina.
Las cetosaminas ingeridas son absorbidas parcialmente, pero no se metabolizan y pueden tener un potencial de prevención contra la carcinogénesis de próstata. Anteriormente, se estableció el potencial antitumoral de las cetosaminas sintéticas en modelos de melanoma, cáncer de mama y cáncer de próstata (183) (184). Los estudios mecanísticos in vitro y ex vivo sugirieron que estos compuestos pueden actuar como inhibidores de la adhesión homotípica y heterotípica de las células de cáncer, posiblemente teniendo como blanco los hidratos de carbono antígenos y las lectinas asociados al tumor (38) (185).
Algunos estudios publicados recientemente mostraron la ventaja de combinar el tomate dietario con otros agentes quimiopreventivos dietarios, tales como brócoli en polvo (186) o vitamina E/selenio (187), para la reducción del crecimiento del adenocarcinoma de próstata implantado en rata, o ajo (188) (189) para la quimioprevención de otros cánceres.
Los experimentos in vitro mostraron que FruHis es en realidad un poderoso protector de la degradación del ADN por ROS en condiciones en las que otros antioxidantes solubles en agua provenientes del tomate no son tan eficaces. El efecto protector de FruHis contra la degradación oxidativa del ADN puede estar parcialmente relacionado, no sólo con su potencial reductor, como se demuestra por los ensayos de Folin-Ciocalteau y FRAP, sino también con sus excelentes propiedades quelantes de cobre (190).
La actividad antioxidante es el mecanismo que prevalece sugerido para el licopeno, selenio, vitamina E y otros agentes de quimioprevención que se están probando actualmente en ensayos clínicos para el cáncer de próstata (191). Con la edad, la próstata gradualmente pierde la capacidad antioxidante, que se manifiesta a través de la pérdida de la expresión de glutatión S-transferasa y la subsiguiente generación de glutatión en la forma reducida (192). Se piensa que esto es una condición que hace que el epitelio de la próstata sea sensible a la mutación inducida por estrés oxidativo, dando lugar finalmente a las transformaciones neoplásicas (193). El consumo de una dieta rica en licopeno, en base a salsa de tomate, sin embargo, dio lugar a una disminución en el daño oxidativo del ADN en pacientes con adenocarcinoma localizado en la próstata (33).
Una serie de antioxidantes que atrapan ROS mostraron un potencial de protección contra la carcinogénesis experimental de próstata, confirmando así que el estrés oxidativo es un objetivo válido para la quimioprevención del cáncer de próstata (166) (192). Muchos de estos antioxidantes, como el licopeno, también actúan como inhibidores de la proliferación de las células de cáncer, lo que indica que poseen el potencial terapéutico (194).
Hasta ahora, se reconocían generalmente tres clases principales de moléculas antioxidantes, estructuralmente definidas, en los productos del tomate: (a) ácido ascórbico (230 mg/100 g de tomate fresco; mucho menos en tomate procesado) y otras vitaminas, como tocoferol; (b) carotenoides, como licopeno y caroteno (hasta 60 mg/100 g de tomate fresco; 5% a 25% menos en tomate procesado); (c) compuestos fenólicos (hasta 550 mg/100 g), como conjugados de los ácidos cafeico y cumárico, flavonoides como quercetina y canferol, etc. (195). Las cetosaminas, con su contenido total de hasta 8.000 mg/100 g de materia seca, pueden representar un nuevo tipo de antioxidantes potenciales dietarios, aunque específicos para los alimentos preparados a partir de productos reconstituidos del tomate deshidratado. Las cetosaminas pueden interactuar con el licopeno contra la carcinogénesis de próstata y la proliferación de células tumorales (38).
Los estudios intervencionales con suplementos de productos de tomate y selenio mostraron un retraso en la progresión de la enfermedad (171).
Actualmente se ha progresado en la determinación de los factores nutricionales y hormonales que aumentan la probabilidad de aparición del cáncer de próstata. La prevención dietaria del cáncer de próstata requiere un régimen nutricional especial combinado con suplementos nutricionales (alimentos funcionales, nutraceúticos, selenio).
El 85% de la ingesta de licopeno proviene de tomates frescos y de productos basados en el tomate (22). Por lo tanto, es necesario considerar las interacciones aditivas o sinérgicas que se producen entre los componentes bioactivos.
Debido a que se sabe que el licopeno dietario se concentra en la próstata humana, se investigó su captación así como la localización subcelular en el ambiente controlado del cultivo de células utilizando las líneas celulares LNCaP, PC-3 y DU145 de cáncer de próstata en humanos. Después de 24 horas de incubación con licopeno 1,48 μM, las células LNCaP acumularon 126,6 pmoles de licopeno/millón de células, que fue 2,5 veces superior a las células PC-3 y 4,5 veces superior a las células DU145 (196).
Entre estas líneas de células, sólo las células LNCaP expresan al antígeno prostático específico y completamente al receptor funcional de andrógenos. Los niveles de antígeno prostático específico secretado al medio de incubación por las células LNCaP se redujeron en un 55% como resultado del tratamiento con licopeno 1,48 μM (196).
Se llevó a cabo la unión del licopeno al dominio unido al ligando del receptor humano de andrógenos, pero no se encontró que el licopeno fuera un ligando de este receptor. A continuación, se realizó el fraccionamiento subcelular de las células LNCaP expuestas al licopeno mediante centrifugación seguida por análisis cuantitativo mediante cromatografía líquida-espectrometría de masas en tándem (CL-EM-EM) para determinar las localizaciones celulares específicas del licopeno. La mayoría del licopeno (55%) se localizó en las membranas nucleares, seguido por un 26% en la matriz nuclear y, a continuación, 19% en los microsomas. No se detectó licopeno en el citosol (196). Estos datos sugieren que la rápida captación del licopeno por las células LNCaP podría facilitarse mediante un receptor o una proteína de unión y que el licopeno se almacena selectivamente en el núcleo de las células LNCaP (196).
Se evaluó la combinación de tomate y brócoli en el modelo Dunning R3327-H de adenocarcinoma de próstata en ratas macho Copenhague (n = 206). Las ratas fueron alimentadas con dietas que contenían 10% de tomate, 10% de brócoli, 5% de tomate más 5% de brócoli (combinación 5:5), 10% de tomate más 10% de brócoli (combinación 10:10) en polvo, o licopeno (23 o 224 nmol/g de dieta) durante aproximadamente 22 semanas, desde un mes antes de recibir implantes s.c. tumorales. Se compararon los efectos de la dieta con la castración quirúrgica (2 semanas antes de la terminación) o finasterida (5 mg/kg de peso corporal por vía oral, 6 días/semana). La castración redujo significativamente el peso de la próstata, las áreas de tumor y el peso del tumor (62%), mientras que la finasterida redujo el peso de la próstata, pero no tuvo ningún efecto sobre el área o peso tumoral. El licopeno a 23 ó 224 nmol/g de la dieta redujo no significativamente el peso del tumor en un 7% o 18% respectivamente, mientras que el tomate redujo significativamente el peso del tumor en un 34%. El brócoli disminuyó significativamente el peso del tumor en un 42%, mientras que la combinación 10:10 causó una disminución significativa del 52% (186).
Las reducciones del crecimiento del tumor estuvieron asociadas con una reducción de la proliferación y el aumento de la apoptosis, según la cuantificación realizada mediante inmunohistoquímica de la proliferación de núcleos de células de antígeno y el ensayo ApopTag. La combinación de tomate y brócoli fue más efectiva que el tomate o el brócoli individualmente para frenar el crecimiento tumoral y concuerda con las recomendaciones de salud pública de aumentar la ingesta de una variedad de componentes de vegetales (186).
También se demostró en estudios de régimen dietario en pacientes que el efecto preventivo del tomate es más potente que el del licopeno solo (125). Se compararon los efectos de tomate rojo, tomate amarillo (desprovisto de licopeno) y licopeno sobre la expresión de conexina 43 (Cx43), que es una proteína que regula el crecimiento celular, sobre una línea celular de cáncer de próstata que expresa el receptor androgénico.
Las células fueron incubadas con suero proveniente de ratas alimentadas con una dieta control (C), control con una dieta suplementada con tomate rojo (TR), control con suplemento de tomate amarillo (TA) o cápsulas de licopeno (L). Posteriormente a la exposición de las células a TR o TA durante 48 horas, la expresión de Cx43 aumentó significativamente en comparación con las células expuestas a C, mientras que el efecto L no fue significativamente diferente. Las células incubadas con TR y L tuvieron niveles similares de licopeno, mientras que aquellas incubadas con TA no contenían licopeno (77) (125).
Estos datos muestran primeramente que el suero nutricionalmente enriquecido con tomates rojos y amarillos podría sobrerregular o desinhibir el turn-over de Cx43 en las células PC3AR, independientemente del nivel de licopeno. Según el enfoque fisiológico utilizado en este estudio, se llega a la conclusión que hay otros compuestos distintos al licopeno que contribuyen significativamente al efecto preventivo de los tomates (77) (125).
En efecto, se había demostrado anteriormente que las células epiteliales humanas de próstata cultivadas en un medio suplementado con factor de crecimiento eran también sensibles a la inhibición del crecimiento por los flavonoides (58). Genisteína, quercetina, canferol y biochanina A presentaron inhibición del crecimiento, dependiente de la dosis, con una concentración inhibitoria del 50% (IC50) entre 25 y 40 μM, mientras que rutina y daidzeína fueron menos potentes, con un IC50 > 60 μM.
14.2. Cáncer de colon
La quimioprevención del cáncer colorrectal se ha convertido en esencial en el mundo moderno industrializado, ya que el cáncer del intestino grueso es una de las principales causas de mortalidad por cáncer, sólo superada por el cáncer de pulmón (197).
El cáncer de colon integra factores de estilo de vida y múltiples alteraciones genéticas, y sin la intervención preventiva, una parte sustancial de la población es probable que desarrolle cáncer colorrectal en algún momento de sus vidas. La dieta y la nutrición juegan claramente un rol en su etiología.
Se examinaron los efectos quimiopreventivos de los carotenoides, como fucoxantina, licopeno y luteína, así como curcumina y su derivado, tetrahidrocurcumina (THC), sobre el desarrollo de focos crípticos aberrantes (FCA), que son las más tempranas lesiones preneoplásicas rectocolónicas, en ratones, iniciados con dihidrocloruro de 1,2-dimetilhidrazina (DMH). Se evaluó también la influencia sobre la proliferación de las células epiteliales crípticas colónicas en términos de la incorporación de 5-bromo-2'-desoxiuridina (BrdU) (198).
Los ratones B6C3F1, machos, de cinco semanas de edad se dividieron en tres grupos, a los grupos 1 y 2 se les dio DMH (20 mg/kg de peso corporal, s.c.) dos veces a la semana durante 3 semanas. Los animales del grupo 1, fueron tratados con uno de los compuestos: licopeno (0,005% y 0,0025%) o fucoxantina (0,01%) en el agua para beber y luteína (0,05%), curcumina (0,5%) o THC (0,5% y 0,2%) en la dieta a partir de las semanas 5-12. El grupo 2 sirvió como un control del carcinógeno solo y a los ratones del grupo 3 se les dieron los compuestos a probar solamente. Todos los animales fueron sacrificados en la semana 12 (198).
Los números de FCA/ratón en el grupo 1 tratado con fucoxantina (47,1 ± 13,7), luteína (42,6 ± 19,6) o 0,5% de THC (46,6 ± 17,7) disminuyeron significativamente en comparación con el valor del grupo 2 control (63,3 ± 19,4). Los números de criptas aberrantes/ratón también fueron significativamente menores después del tratamiento con luteína (79,9 ± 34,7) o 0,5% de THC (81,8 ± 32,5) que en el grupo control (115,1 ± 37,1). Fucoxantina, licopeno y luteína disminuyeron el número de focos crípticos aberrantes de colon en ratones (198).
Los índices de marcación con BrdU en ratones tratados con luteína y 0,5% de THC disminuyeron significativamente en ambos compartimentos de la mitad superior e inferior de las criptas colónicas en comparación con los controles, especialmente en los datos de la mitad superior correspondientes a la reducción de ACs/ratón. Los resultados sugieren que fucoxantina, luteína y THC pueden tener potencial como agentes quimiopreventivos contra la carcinogénesis colónica.
El efecto inhibitorio de los cuatro carotenoides frecuentes en la sangre y los tejidos humanos contra FCA colónicos se examinó en ratas Sprague-Dawley. Las ratas recibieron tres dosis intrarrectales de N-metilnitrosourea en la semana 1 y una sonda diaria de dosis escalonadas de los carotenoides durante las semanas 2 y 5. Licopeno, luteína, a-caroteno y carotenos de palma (una mezcla de a-caroteno, b-caroteno y licopeno) inhibieron cuantitativamente el desarrollo de FCA en la semana 6, pero el b-caroteno no. Los resultados sugirieron que el licopeno y la luteína en pequeñas dosis podrían prevenir potencialmente la carcinogénesis colónica (199).
Se investigó la inhibición de la carcinogénesis de colon por el licopeno y el jugo de tomate en ratas F344/NSlc hembras de siete semanas de edad, las cuales recibieron una dosis intrarrectal de 2 mg (experimento I) o de 4 mg (experimento II) de N-metilnitrosourea 3 veces a la semana durante 3 semanas y tuvieron libre acceso a una de las 4 bebidas: agua común (grupo control), solución de 17 ppm de licopeno en agua (grupo L) y jugo de tomate diluído que contenía 17 ppm (grupo jT) o 3,4 ppm (grupo jt) de licopeno, a lo largo de los experimentos (200). La incidencia de cáncer de colon en la semana 35 fue significativamente menor en el grupo jT, pero no en el grupo L, que en el grupo control: 21% y 33% vs 54% en el experimento I. Fue significativamente inferior en el grupo de jT que en el jt y grupos control, 40% vs 72% y 84% en el experimento II. Se detectó una cantidad apreciable de licopeno (0,02 μg/g) en la mucosa del colon de ratas en el grupo jT, pero no en el grupo de jt. Finalmente, el efecto inhibitorio del jugo de tomate rico en licopeno (17 ppm) se observó en el modelo de carcinogénesis de colon en ratas (200).
Los resultados sugieren que el jugo de tomate rico en licopeno puede tener un efecto protector contra la carcinogénesis colónica.
Cuando se administra un carcinógeno específico para colon, como azoximetano (AOM), iNOS y COX-2 están sobreexpresadas en los tumores de colon en humanos y en ratas. iNOS puede regular la producción de COX-2 de las prostaglandinas proinaamatorias, que se sabe que desempeñan un rol fundamental en el desarrollo del tumor de colon (102).
Los experimentos fueron diseñados para evaluar las propiedades potenciales quimiopreventivas de inhibidores muy selectivos de iNOS, administrados individualmente y en combinación con un inhibidor selectivo de COX-2, sobre el desarrollo de FCA colónicos inducidos por AOM. Ratas F344 fueron alimentadas con dietas experimentales que contenían uno de los siguientes: 0, 10, 30 o 100 ppm del inhibidor selectivo de iNOS: tetrazolamida de L-N6-(1-iminoetil)lisina (SC-51), 1800 ppm del menos potente inhibidor selectivo de iNOS: aminoguanidina (AG), 500 ppm del inhibidor de COX-2: celecoxib, 320 ppm del antiinflamatorio no esteroideo: sulindac (control positivo) o 30 ppm de SC-51 con 500 ppm de celecoxib y 100 ppm de SC-51 con 500 ppm de celecoxib (102).
Una y dos semanas después, las ratas recibieron inyecciones s.c. de AOM en una dosis de 15 mg/kg de peso corporal. A las 17 semanas de edad, todas las ratas fueron sacrificadas. Se evaluó cada colon por FCA y las mucosas colónicas fueron ensayadas respecto a las actividades enzimáticas de las isoformas de COX y NOS (102).
Estos agentes fueron analizados en muestras de sangre venosa, recogidas en diversos puntos de tiempo. SC- 51, administrado solo, demostró una inhibición dependiente de la dosis de la incidencia de FCA de colon. Las dosis más altas de SC-51 (100 ppm) y AG (1800 ppm) suprimieron significativamente la incidencia de FCA colónica y la multiplicidad de criptas en cuanto al número de criptas/focos aberrantes. Es importante destacar que la combinación de cualquier dosis baja o alta efectiva de SC-51 (30 o 100 ppm) y celecoxib (500 ppm) suprimió significativamente la formación de FCA colónicos inducidos por AOM y redujo significativamente la multiplicidad de cuatro o más criptas/focos aberrantes en mayor medida que lo hicieron estos agentes administrados individualmente. Como era de esperar, sulindac inhibió significativamente la formación de FCA de colon y redujo significativamente la multiplicidad de cuatro o más criptas aberrantes a aproximadamente el 45%. Las actividades enzimáticas de COX-2 e iNOS fueron inducidas en animales tratados con AOM y la administración de inhibidores de iNOS, SC-51 y AG inhibió significativamente las actividades de ambos, iNOS y COX-2, en la mucosa colónica. La administración combinada de SC-51 y celecoxib inhibió la actividad de COX-2 en mayor medida que lo hizo cualquiera de estos agentes administrados solos (102).
Los inhibidores selectivos de iNOS mostraron propiedades quimiopreventivas y la co-administración con un inhibidor selectivo de COX-2 resultó con mayor potencial quimiopreventivo.
Los estudios epidemiológicos sugieren que los carotenoides, como b-caroteno y luteína, desempeñan un rol importante en la reducción del riesgo de varios tipos de cáncer, pero mostraron ambigüedad respecto al rol de estos compuestos en el cáncer de colon.
Se evaluó entonces la eficacia quimiopreventiva de b- caroteno y luteína en niveles de dosis de aproximadamente 4 y 8% de la dosis tolerada (DT) de 2.500 ppm en ratas F344, machos, y también aproximadamente 40 y 80% de la DT sobre la carcinogénesis de colon inducida por AOM, usando FCA como un biomarcador sustituto para el cáncer de colon (201).
La administración de 100 ó 200 ppm de b-caroteno inhibió significativamente la formación colónica total de FCA, inducida por AOM, en un 24% y 36% respectivamente, mientras que la luteína a 200 ppm produjo una inhibición del 27%, pero no tuvo efecto significativo en el nivel de dosis de 100 ppm. Sorprendentemente, la administración de 1.000 ó 2.000 ppm de b-caroteno y luteína aumentó la formación de FCA de colon en una manera dependiente de la dosis, por ejemplo: a 124% y 144% para el primero y, 110% y 159% para el segundo (201).
Tomados en conjunto, estos datos indican que la actividad quimiopreventiva de b-caroteno y luteína contra la carcinogénesis de colon depende de la dosis.
Como la inflamación crónica del tracto gastrointestinal se considera como un factor de riesgo para la formación de tumor, se evaluó el efecto de la suplementación dietaria con quercetina o b-caroteno sobre la carcinogénesis de colon y la respuesta inaamatoria en ratas alimentadas con una dieta alta en grasas rica en ácidos grasos ω-6. La suplementación con quercetina o b-caroteno redujo signi`cativamente el número de FCA sólo en los animales alimentados con dieta alta en grasas; sin embargo, no se encontró diferencia significativa alguna en la incidencia del tumor (202). En la semana 33, se redujo la expresión de iNOS mediante quercetina sin significación estadística y la expresión de COX-2 se redujo ligeramente en ratas con suplementación de b-caroteno. No se observaron cambios en los niveles de PGE2. Por consiguiente, los antioxidantes dietarios se consideran supresores efectivos de la formación de lesiones precancerosas en el colon expuesto a dieta de alto riesgo (202).
Recientemente, se evaluó en los FCA la actividad protectora de sulfuro de dialilo, licopeno y teaflavina, importantes antioxidantes del ajo, tomate y té negro respectivamente, durante la carcinogénesis del colon (197). Como la inhibición de las actividades de COX-2 y de iNOS se correlaciona con la prevención del cáncer de colon, el estudio continuó con la determinación del cambio en la expresión de estas proteínas. Al seguir el tratamiento, se observó reducción significativa en la incidencia de los FCA (en 43,65% por sulfuro de dialilo, 57,39% por licopeno y 66,08% por teaflavinas), lo cual estuvo de acuerdo con la reducción de la expresión de COX-2 y de iNOS. El efecto del alimento intacto resultó ser más pronunciado que el de sus componentes por separado (197).
La concentración plasmática de IGF-I disminuyó significativamente en alrededor de 25% tras la administración de extracto de licopeno de tomate a pacientes con cáncer de colon (n = 56), candidatos a colectomía, reclutados pocos días a unas pocas semanas antes de la cirugía, en comparación con el grupo tratado con placebo. No se observó cambio significativo alguno en la proteína- 3 unida a IGF-I o a IGF-II, mientras que la relación molar de IGF-I/IGFBP-3 (de IGF-I) disminuyó significativamente (203).
Considerando que los altos niveles plasmáticos de IGF-I se habían sugerido en estudios epidemiológicos como un factor de riesgo para varios tipos de cáncer, entre ellos cáncer de colon, los resultados de este estudio aleatorizado de doble-ciego muestran que el extracto de licopeno de tomate tiene un rol en la prevención del cáncer de colon y posiblemente, de otros tipos de cáncer.
Asimismo, tomates maduros fueron sometidos a una digestión simulada in vitro y la fracción acuosa obtenida fue proporcionada a las células HT-29 y HCT-116 de adenocarcinoma de colon. La cantidad de licopeno liberada durante la digestión y transferida a la fracción acuosa fue 10 veces inferior a la presente en el homogenato de tomate antes de la digestión. El carotenoide se acumuló en las células del adenocarcinoma de colon, en forma dependiente de la dosis, después de la adición del digesto de tomate (20-100 mL/L) durante 24 hs. El digesto de tomate inhibió el crecimiento de las células HT-29 y HCT-116, en forma dependiente de la dosis, debido a la detención de la progresión del ciclo celular en la fase G0/G1 y por inducción de la apoptosis. También se observó una inhibición de la ciclina D1, Bcl-2 y la expresión de Bcl-xL, sin aparentes cambios en p53, p21, p27 y Bax (204).
Estos datos demuestran que el digesto de tomate es capaz de inhibir el crecimiento de las células del cáncer de colon mediante la modulación de la expresión de los reguladores del ciclo celular y la apoptosis.
La regulación aberrante de la ruta de señalización fosfoinosítido 3-quinasa/Akt de supervivencia en el cáncer ha llevado a un interés en la supresión de esta vía para tratar el cáncer. Recientemente, se demostró que el licopeno inhibía la proliferación celular en células HT-29 de cáncer de colon humano con un valor IC50 de 10 μM, por medio de la supresión de la activación de Akt y del nivel de proteína b-catenina no fosforilada en las células humanas de cáncer de colon. Los resultados inmunocitoquímicos indicaron que el licopeno aumentó la forma fosforilada de las proteínas b-cateninas. Estos efectos también se asociaron con una reducción de la actividad promotora y de la expresión de la proteína de la ciclina D1. Además, el licopeno aumentó significativamente la abundancia de p27kip nuclear inhibidor de quinasa y dependiente de ciclina, e inhibió la fosforilación de la proteína supresora del tumor retinoblastoma en células humanas de cáncer de colon (142).
Recientemente se efectuó una digestión in vitro de los tomates licopeno b-ciclasa (tlcy-b) y se analizaron los efectos del digesto sobre la proliferación celular de células HT-29 de adenocarcinoma de colon humano, crecidas en monocapas. Estos tomates tlcy-b, obtenidos mediante modulación de la carotenogénesis vía ingeniería genética, contienen una gran cantidad de b-caroteno, dando así tomates de intenso color anaranjado. Los tomates digeridos fueron diluidos (20 mL, 50 mL y 100 mL/L) en un medio de cultivo y se agregaron a las celdas para diferentes tiempos de incubación (24, 48 y 72 h) (205).
La inhibición del crecimiento celular por el digesto de tomate fue dependiente de la dosis y se debió a la detención de la progresión del ciclo celular en la fase G0/G1 y G2/M y a la inducción de apoptosis. Se observó una inhibición o subregulación de la expresión de ciclina D1, Bcl-2 y Bcl-x1. También se encontró que el tratamiento térmico de las muestras antes de la digestión aumentó la liberación de b-caroteno y, por lo tanto, la inhibición del crecimiento celular. Para inducir la misma inhibición del crecimiento celular con b-caroteno purificado solubilizado en tetrahidrofurano, se requiere una mayor cantidad del carotenoide, lo que sugiere que el b- caroteno micelarizado durante la digestión es utilizado más eficientemente por las células, pero también pueden estar presentes otras moléculas del tomate que se hicieron disponibles durante la digestión y cooperar con el b- caroteno en la detención del crecimiento celular (205).
Como se demostrara anteriormente, el consumo de pescado que contiene ácido eicosapentaenoico (eicosapentaenoic acid, EPA) se correlacionó con menor riesgo de cáncer; lo mismo para el licopeno. Por eso, recientemente, se demostró que una baja concentración de licopeno y EPA podría inhibir sinérgicamente la proliferación de células HT-29 de cáncer de colon humano mediante la supresión de la vía de señalización de fosfatidilinositol 3-quinasa/Akt (35).
Además, el tratamiento de licopeno y EPA también bloqueó sinérgicamente la activación corriente abajo de la molécula mTOR. Los resultados inmunocitoquímicos de tinción revelaron que licopeno y EPA también podrían desinhibir o sobrerregular la expresión de las proteínas apoptóticas, tales como Bax y ligando Fas para suprimir la supervivencia celular (35).
Estos nuevos resultados sugieren que el licopeno y EPA inhibieron sinérgicamente el crecimiento de las células HT-29 de cáncer de colon humano, incluso a baja concentración. Estos efectos inhibitorios de ambos compuestos se asociaron, en parte, con la inhibición o subregulación de la vía de señalización PI-3K/Akt/mTOR.
14.3. Cáncer de pulmón y otros cánceres
La administración de licopeno en el agua potable inhibió la incidencia y la multiplicidad de adenomas y carcinomas de pulmón en el modelo de carcinogénesis de múltiples órganos en ratones machos, pero no en hembras (206). El licopeno (50 ppm en agua potable) tuvo efecto inhibitorio sobre la carcinogénesis de pulmón en ratones machos (207).
Los ensayos de intervención con suplementos de b-caroteno no han dado ningún efecto o bien un efecto adverso sobre la incidencia de cáncer de pulmón en fumadores y en trabajadores expuestos a asbestos. Se analizó la asociación entre el riesgo de cáncer de pulmón y la ingesta de carotenoides específicos utilizando los datos primarios de siete estudios de cohorte en América del Norte y Europa. La ingesta de carotenoides se estimó a partir de los cuestionarios administrados en la etapa basal en cada estudio. Los modelos multivariados incluyeron tabaquismo y otros factores potenciales de riesgo (208).
Durante el seguimiento de hasta 7-16 años a través de los estudios, se diagnosticaron 3.155 casos incidentales de cáncer de pulmón entre los 399.765 participantes. La ingesta de b-caroteno no estuvo significativamente asociada con el riesgo de cáncer de pulmón, mientras que la ingesta de b-criptoxantina mostró una asociación inversa (208).
Estos resultados no cambiaron después del ajuste por ingesta de vitamina C (con o sin suplementos), foliato (con o sin suplementos) y otros carotenoides, y uso de multivitaminas. Aunque el tabaquismo es el principal factor de riesgo de cáncer de pulmón, una mayor ingesta de alimentos con alto contenido en b-criptoxantina, como frutas cítricas, puede disminuir el riesgo en forma modesta (208).
Recientemente, se llevó a cabo una revisión sistemática de la asociación entre los carotenoides y el cáncer de pulmón. El diseño se basó en búsquedas en bases de datos electrónicas de los artículos publicados hasta septiembre de 2007. Se analizaron seis ensayos clínicos aleatorios examinando la eficacia de los suplementos de b- caroteno y 25 estudios observacionales prospectivos evaluando la asociación entre los carotenoides y el cáncer de pulmón mediante el uso de metanálisis de efectos aleatorios (209).
Se demostró que la suplementación con b-caroteno no está asociada con una disminución en el riesgo de desarrollar cáncer de pulmón. Los resultados de estudios prospectivos de cohortes sugirieron asociaciones inversas entre los carotenoides (b-criptoxantina, a-caroteno, licopeno y luteína-zeaxantina) y el cáncer de pulmón; sin embargo, la disminución en el riesgo fue generalmente pequeña y no significativa. Estas asociaciones inversas pueden ser el resultado de la función de las mediciones de los carotenoides como un marcador de un estilo de vida más saludable (mayor consumo de frutas y hortalizas) o de confusión residual por el tabaco (209).
14.4. Cáncer de mama
El efecto protector de los carotenoides no ha sido concluyente en la quimioprevención del cáncer de mama. Por ello, se llevó a cabo un estudio caso-control en China durante 2004-2005 para investigar si la ingesta de licopeno, a-caroteno, b-caroteno, b-criptoxantina y luteína/zeaxantina está inversamente asociada con el riesgo de cáncer de mama, con 122 pacientes, cuya edad estaba entre los 24-87 años, con cáncer de mama confirmado histopatológicamente y 632 mujeres sanas de edad correspondiente al grupo anterior. Se registró la ingesta dietaria habitual y el estilo de vida mediante entrevistas personales usando cuestionarios adecuados y validados. Se utilizó la base de datos de composición de nutrientes de USDA (210).
a-Caroteno y luteína/zeaxantina no mostraron asociación con el cáncer de mama, mientras que una ingesta más alta de licopeno, b-caroteno y b-criptoxantina resultó asociada a un menor riesgo de cáncer de mama entre las mujeres chinas (210).
Como IGF-I es un importante factor de crecimiento asociado con un mayor riesgo de cáncer de mama premenopáusico, se estudiaron recientemente los efectos del licopeno sobre el sistema de IGF-1 en sobrevivientes del cáncer de mama en mujeres premenopáusicas y mujeres con alto riesgo familiar de cáncer de mama (211).
Este ensayo controlado aleatorizado demostró que dos meses de suplementación con licopeno no tuvo efecto alguno sobre IGF-I total sérico en las sobrevivientes de cáncer de mama. En cambio, se observaron efectos beneficiosos en mujeres sanas con alto riesgo (disminución significativa de IGF-I libre en la población con historia familiar: 7,6%).
Los agentes dietarios con potencial quimioprotector, como genisteína derivada de la soja y licopeno derivado del tomate, han demostrado que alteran la expresión de los genes, tal que pueden promover o inhibir potencialmente los procesos carcinogénicos. Para explorar los mecanismos por los que estos agentes pueden estar actuando, se examinó la metilación del ADN modulando la capacidad de genisteína o licopeno en varios genes relevantes del cáncer de mama. Se encontró que una concentración baja, no tóxica, de genisteína (3,125 μM, resuplementada cada 48 horas durante 1 semana) o una dosis única de licopeno (2 μM) provoca la desmetilación parcial del promotor del gen GSTP1 supresor tumoral en la línea celular MDA-MB-468 de cáncer de mama. Los estudios de RT-PCR confirmaron la falta de expresión de GSTP1 en MDA-MB-468 no tratadas, con la restauración de la expresión de GSTP1 después del tratamiento con genisteína o licopeno. El gen RARb2, sin embargo, no fue desmetilado por genisteína ni licopeno en ninguna de estas líneas celulares de cáncer de mama. Pero, el licopeno (2 μM, una vez por semana durante 2 semanas) no indujo desmetilación de los genes RARb2 y del HIN-1 en las células fibrocísticas MCF10A no-cancerígenas de mama (212).
Estos datos muestran por primera vez que el licopeno del tomate tiene actividad desmetilante directa del ADN. Tanto la genisteína como el licopeno, a concentraciones muy bajas, relevantes dietariamente, pueden mitigar los procesos tumorígenos vía la modulación del promotor de la metilación de la expresión génica.
14.5. Cáncer de ovarios
Para investigar si la ingesta de a-caroteno, b-caroteno, b-criptoxantina, luteína y zeaxantina, y licopeno está inversamente asociada con el riesgo de cáncer de ovario, se llevó a cabo un estudio caso-control en China durante 1999-2000 con 254 pacientes con cáncer epitelial de ovario confirmado histológicamente y 652 controles de edad correspondiente al grupo anterior. Se registró la ingesta dietaria habitual y el estilo de vida mediante entrevistas personales usando cuestionarios validados y la base de datos de composición de nutrientes del Departamento de Agricultura de EE.UU. (USDA) para calcular la ingesta de los carotenoides específicos (213).
Se usaron los análisis de regresión logística incondicional para estimar ORs y los intervalos de confianza 95% (IC), correspondientes a la edad, localidad, educación, índice de masa corporal (TMC), tabaquismo, bebedores de té, paridad, uso de anticonceptivos orales, terapia de reemplazo hormonal, estado menopáusico, historia familiar de cáncer de ovario, actividad física e ingesta de energía. Los resultados mostraron que una mayor ingesta de carotenoides reduce significativamente los riesgos de cáncer epitelial de ovario (213).
14.6. Cáncer de vejiga
El jugo de tomate conteniendo licopeno disminuyó el número, pero no la incidencia, de carcinomas de células transicionales de vejiga urinaria en ratas macho. Se estudiaron los efectos de drogas anti-inflamatorias no esteroideas (DAINE) como piroxicam y los carotenoides licopeno y b-caroteno, solos o en combinación, sobre el desarrollo de carcinomas super`ciales de vejiga urinaria inducidos por N-butil-N-(4-hidroxibutil) nitrosamina (BBN) en ratas Fischer 344, machos, de 6 semanas de edad, que recibieron 0,05% BBN en el agua potable durante 8 semanas, seguido por la administración de piroxicam (0,0075% en la dieta), licopeno (0,0025% en el agua potable) y/o b-caroteno (0,0025% en el agua) durante 12 semanas. Luego fueron sacrificadas para el análisis histológico de las lesiones de la vejiga urinaria (214).
Se analizó el potencial de proliferación celular mediante tinción inmunohistoquímica del antígeno nuclear de proliferación celular (proliferative cell nuclear antigen: PCNA). Piroxicam solo, piroxicam + licopeno y piroxicam + licopeno + b-caroteno, todos disminuyeron significativamente la incidencia y el número de carcinomas celulares de transición (transitional cell carcinomas: TCCs), pero la combinación de piroxicam con carotenoides no causó una clara mejoría en el potencial preventivo de piroxicam (214).
Piroxicam + b-caroteno también causó una reducción significativa y licopeno sólo una disminución ligera, pero no significativa, en el número de TCCs. En cambio, b-caroteno solo y licopeno + b-caroteno no tuvieron inauencia inhibitoria sobre ninguna de las categorías de lesión examinadas y el segundo aumentó significativamente la proporción de TCCs de alto grado. Sin embargo, todos los agentes quimiopreventivos, ya sea solos o en combinación, disminuyeron significativamente el índice de TCC PCNA, extendiéndose el efecto al epitelio de alrededor en los grupos de piroxicam + licopeno y piroxicam + licopeno + b-caroteno (214).
Estos resultados indican que el DAINE piroxicam puede ser un agente quimiopreventivo más efectivo que licopeno y b-caroteno para la carcinogénesis superficial de vejiga urinaria.
Se continuó con estudios sobre los efectos del jugo de tomate sobre la carcinogénesis de vejiga urinaria en ratas Fischer 344 machos, inducida por BBN. En forma análoga al diseño anterior, las ratas recibieron BBN durante 8 semanas, seguido por jugo de tomate diluido durante 12 semanas y fueron sacrificadas a las 20 semanas de iniciado el experimento. La concentración de licopeno fue elevada en el hígado de las ratas que recibieron jugo de tomate. El número, pero no la incidencia de los TCCs de células de vejiga urinaria disminuyó en el grupo que recibió el jugo de tomate, según el análisis histopatológico de las lesiones de vejiga urinaria. No se notó influencia en la incidencia de hiperplasias simples y nodulopapilarias, ni invasión ni diferenciación de TCC (215).
Estos resultados indican que el jugo de tomate, presumiblemente el licopeno que contiene y otros antioxidantes en combinación, ejerce un efecto inhibitorio sobre el desarrollo de TCCs de vejiga urinaria en rata.
14.7. Hepatocarcinogénesis
La mezcla de carotenoides resultó ser muy eficaz en la actividad anticarcinogénica; así, el desarrollo de hepatoma en un grupo de alto riesgo de cáncer de hígado fue suprimido significativamente por el tratamiento con mezcla de carotenoides naturales (91). Sin embargo, los resultados en relación con el potencial quimioproventivo del licopeno en la hepatocarcinogénesis en ratas son ambiguos.
Se investigaron los efectos de los carotenoides: a-caroteno, b-caroteno, licopeno, b-criptoxantina, zeaxantina, luteína, cantaxantina y astaxantina, sobre la invasión de células de ascitis de hepatoma AH109A en ratas mediante el co-cultivo de las células de hepatoma con células mesoteliales (células M) derivadas del mesenterio de rata. Todos los carotenoides examinados inhibieron la invasión AH109A en una forma dependiente de la dosis, hasta 5 μM. Las células cancerosas previamente cultivadas con hipoxantina (HX) y xantina oxidasa (XO) mostraron una actividad muy invasiva. Los carotenoides, 5 μM de b-caroteno y astaxantina, suprimieron la capacidad invasiva potenciada por esta especie reactiva de oxígeno mediante el tratamiento simultáneo de las células AH109A con los carotenoides, HX y XO (216).
Estos resultados sugieren que las propiedades antioxidantes de estos carotenoides pueden estar involucrados en su acción anti-invasiva.
Se ha demostrado que la administración dietaria de licopeno o de la medicina herbaria Sho-saiko-to (TJ-9) tiene actividad anticarcinogénica, aunque no es bien conocido el mecanismo por el cual estos productos protegen contra la carcinogénesis. Por ello, se investigaron los resultados de administrar licopeno y TJ-9 sobre la ocurrencia de neoplasia hepática en ratas Long-Evans Cinnamon (LEC), que es un modelo bien caracterizado de hepatocarcinogénesis espontánea. Se administró una dieta con 0,005% de licopeno (originalmente el producto de la óleo-resina de tomate contenía 13% de licopeno) y 1% TJ-9 (extractos crudos de 7 hierbas: raíz de bupleurum, tubérculo pinellia, raíz de Scutellaria, fruta de jinjolero, raíces de ginseng, raíz de Glycyrrhiza y rizoma de jengibre) desde las 6 semanas de edad hasta que las ratas fueron sacrificadas a las 76 semanas de edad, momento en el que la mayoría de los animales no-tratados tenían carcinoma hepatocelular. Se analizó histológicamente el desarrollo del carcinoma hepatocelular en los grupos tratados mediante la comparación con los controles sin tratar. La forma placentaria de glutatión S-transferasa (GST-P) fue analizada por un método de inmunohistoquímica. Se analizaron las concentraciones de cobre, hierro y cinc, que parecen desempeñar un rol en la hepatocarcinogénesis en ratas LEC (217).
El número de áreas GST-P-positivas dentro de las lesiones por carcinoma hepatocelular fue significativamente mayor en el grupo TJ-9 que en el control o el grupo de licopeno. El estudio también demostró una menor concentración de hierro en el hígado del grupo de licopeno que en el grupo control. No hubo diferencias en los niveles de a-fetoproteína en suero o en las tasas acumulativas de supervivencia entre los grupos (217).
En conclusión, la administración a largo plazo (6 a 76 semanas de edad) de una dieta con 0,005% de licopeno o TJ-9 no redujo el riesgo de hepatocarcinogénesis en un modelo de carcinogénesis espontánea en hígado de ratas (217).
Recientemente se investigaron los efectos antigenotóxicos y anticarcinogénicos de la óleo-resina del tomate dietario ajustados a la concentración de licopeno de 30, 100 ó 300 ppm (administrados 2 semanas antes y durante 8 semanas después de la exposición al carcinógeno) en el hígado de ratas Wistar, machos, tratadas con una sola dosis i.p. de 20 o 100 mg/kg de dietilnitrosamina (DEN) respectivamente. Se utilizaron como criterios de valoración: el nivel del daño al ADN en las células hepáticas y el desarrollo de hepatocitos putativos preneoplásicos únicos, minifocos y focos de hepatocitos alterados (FHA) positivos para GST-P. La reducción significativa del daño al ADN se detectó cuando se administró la mayor concentración de licopeno antes y durante la exposición a DEN (20 mg/kg) (218).
Tomados en conjunto, los datos sugieren un efecto quimiopreventivo de la óleo-resina del tomate contra el daño al ADN inducido por DEN, pero inefectividad clara en las fases de iniciación o promoción de la hepatocarcinogénesis en ratas.
14.8. Cáncer de endometrio
Se analizó la relación entre la ingesta de carotenoides y retinol y el cáncer de endometrio, ya que los datos disponibles eran inconsistentes. Se realizó un estudio de casos- control multicéntrico en diferentes zonas de Italia entre 1992 y 2006 sobre 454 mujeres con cáncer de endometrio incidental, histológicamente confirmado y 908 controles admitidos en la misma red de hospitales de casos para condiciones agudas, no neoplásicas. La ingesta de carotenoides y retinol fue computada a partir de un cuestionario validado de frecuencia de alimentos (219).
Como resultado, al comparar el cuartil de ingesta más alto con el cuartil más bajo, la disparidad o razón de probabilidades (odds ratio: OR) de cáncer de endometrio fueron 0,69 para b-caroteno, 0,65 para b-criptoxantina y 0,59 para la ingesta de luteína plus zeaxantina. No surgió asociación alguna con retinol (OR = 1,31), a-caroteno (OR = 0,94) y licopeno (OR = 0,95) (219).
En conclusión, estos resultados apoyan un rol favorable de los carotenoides dietarios seleccionados sobre el riesgo de cáncer de endometrio.
15. EFECTOS ADVERSOS DE LOS CAROTENOIDES
Resulta importante tener en cuenta los riesgos que existen al realizar la suplementación con algunos carotenoides y la comparación con el aporte a través de la dieta.
Como se ha visto en este trabajo, los carotenoides poseen propiedades antioxidantes y varias otras que contribuyen a la acción anticarcinogénica que se les ha atribuido a lo largo del tiempo. Sin embargo, se conocen estudios y ensayos clínicos que han mostrado efectos inesperados y paradojales para algunos carotenoides, en especial para b-caroteno.
En la década de 1980, los resultados de estudios observacionales sugirieron que los suplementos dietarios con los antioxidantes b-caroteno y vitamina A (también llamada retinol) podrían reducir la incidencia de cáncer, en particular del cáncer de pulmón. Muchos estudios sugirieron que las personas que comen muchas frutas y verduras tienen significativamente menos probabilidad de contraer cáncer (220) (221), señalando a los carotenos como los ingredientes activos. Parecía que un alto consumo de caroteno alimenticio podría reducir significativamente el riesgo de cáncer de pulmón (222), cáncer de vejiga (223), cáncer de mama (224) (225), cáncer de esófago (226) y cáncer de estómago (226) (227). Sin embargo, estudios observacionales no pueden probar la causa y efecto. Siempre es posible que las personas que consumen grandes cantidades de carotenos en su dieta sean diferentes en otros aspectos; por ejemplo: podrían tener diferentes estilos de vida, algunos más saludables que otros. Aunque las personas que consumen alimentos altos en b-caroteno parecen obtener cierta protección frente a enfermedades cardíacas y cáncer, cuando los participantes recibieron suplementos de b-caroteno, no se observó efecto protector alguno.
Se desarrollaron ensayos clínicos, como ATBC (a-Tocopherol, b-Carotene Cancer Prevention) y CARET (b-Carotene and Retinol Efficacy Trial), para probar si los suplementos de estos retinoides podrían prevenir el cáncer de pulmón en personas con alto riesgo de la enfermedad, como fumadores desmedidos, ex-fumadores y/o trabajadores expuestos a asbestos. El ensayo ATBC fue un ensayo de prevención del cáncer llevado a cabo por el Instituto Nacional del Cáncer de EE.UU. (NCI) y el Instituto Nacional de Salud Pública de Finlandia (National Public Health Institute of Finland) desde 1985 a 1993. El propósito del estudio fue determinar si los suplementos de ciertas vitaminas podrían prevenir el cáncer de pulmón y otros cánceres en un grupo de 29.133 hombres fumadores en Finlandia. Los participantes de 50 a 69 años de edad tomaron una píldora diariamente durante cinco a ocho años, que contenía uno de los siguientes: 50 mg de a-tocoferol (una forma de vitamina E), 20 mg de b-caroteno (precursor de la vitamina A; esta cantidad es más de 10 veces la necesaria para proporcionar el requerimiento diario de vitamina A), ambos, o un placebo (píldora inactiva con la apariencia de la vitamina A). El estudio se realizó en Finlandia debido a las altas tasas de cáncer de pulmón en hombres en ese país debido principalmente al consumo de cigarrillos. Las mujeres finlandesas no se incluyeron en el estudio porque su tasa anual de cáncer de pulmón (8 casos por cada 100.000 mujeres) fue sustancialmente menor que la de los hombres finlandeses (67 casos por cada 100.000 hombres).
La vitamina E y el b-caroteno fueron elegidos en este ensayo debido a que los estudios epidemiológicos han relacionado la ingesta dietaria alta y los niveles séricos elevados de estos micronutrientes con un menor riesgo de cáncer, especialmente cáncer de pulmón. Ambos son antioxidantes, compuestos que pueden prevenir que los agentes carcinógenos dañen el ADN y otros sistemas celulares.
En 1994, los resultados del estudio ATBC (228-230) mostraron que los suplementos de b-caroteno no previnieron el cáncer pulmonar, sino que incrementaron en un 18% el riesgo de contraerlo y se registró un 8% de aumento de mortalidad global en aquellos que recibieron los suplementos de b-caroteno.
La vitamina E no tuvo efecto sobre la incidencia de cáncer de pulmón o la mortalidad global. Los hombres que tomaron ambos suplementos tuvieron resultados similares a los del b-caroteno solo (229). Los efectos adversos del b-caroteno aparecieron más fuertemente en hombres con un consumo de alcohol relativamente modesto (más de 11 g por día; 15 g de alcohol equivale a una bebida) y en los que fumaban por lo menos 20 cigarrillos diarios (231). Los participantes que tomaron vitamina E tuvieron un 32% menos de casos de cáncer de próstata y 41% menos de muertes por cáncer de próstata. La muerte por accidente cerebrovascular hemorrágico también se incrementó en un 50% en los hombres que tomaron suplementos de a-tocoferol; el aumento se produjo principalmente entre los hombres con hipertensión (232) (233).
En los Estados Unidos se llevó a cabo el ensayo CARET (234) (235) en el que se compararon los efectos de b-caroteno más vitamina A (palmitato de retinilo) con los de placebo en 18.314 hombres y mujeres de edad entre 45 y 74 años, entre los cuales había fumadores, ex-fumadores y/o trabajadores expuestos a asbestos. Se observaron un 28% mayor de incidencia de cáncer de pulmón y 17% más de mortalidad global en el grupo que tomó la suplementación vitamínica. Es decir que el ensayo CARET confirmó los resultados del ensayo ATBC. Tanto CARET como el ensayo ATBC fueron de doble ciego, lo que significa que ni los participantes ni sus médicos supieron, hasta después, quien recibió los suplementos y quién recibió el placebo.
Una vez concluídos ambos ensayos, se continuó con el seguimiento de los participantes para conocer los efectos a largo plazo de los suplementos sobre las tasas de cáncer de pulmón y las tasas de mortalidad general. Los datos adicionales sobre la incidencia de cáncer y la mortalidad relacionada con causas específicas se adquirieron hasta abril de 1999 (seis años después del final del ensayo) y para la mortalidad total hasta abril de 2001 (ocho años después del final del ensayo).
Los resultados demostraron que los participantes del CARET que habían tomado suplementos de b-caroteno y vitamina A tenían un 12% más de riesgo de desarrollar cáncer de pulmón durante el período de seguimiento post-ensayo que los que habían tomado placebo. La gente en el grupo con suplemento también tenía un riesgo un 8% mayor de morir por cualquier causa que los del grupo placebo. Estos riesgos mayores persistieron después de seis años de seguimiento post-ensayo, aunque por entonces no fueron estadísticamente significativos, es decir, que podrían haber ocurrido por casualidad.
Los que tomaron el suplemento no estuvieron en riesgo elevado de muerte por enfermedad cardíaca o accidente cerebrovascular durante el período de post-ensayo.
En los ocho años de seguimiento post-ensayo que realizara el grupo de ATBC (236), los participantes que tomaron b-caroteno experimentaron un 7% más de mortalidad global que los hombres con placebo. El aumento del riesgo de cáncer de pulmón se redujo poco después que los participantes dejaron de tomar los suplementos de b-caroteno. Durante los dos últimos años, las tasas de mortalidad general fueron comparables con la de los participantes que no recibieron b-caroteno. En el grupo que recibió b-caroteno, la mayor mortalidad durante el ensayo se debió a enfermedades cardiovasculares y cáncer de pulmón. En contraste, el aumento de la mortalidad durante el período posterior a la intervención se debió sólo a enfermedades cardiovasculares. Las tasas de menor incidencia de cáncer de próstata en los participantes que tomaron suplementos de a-tocoferol durante el ensayo volvieron a la normalidad poco después de que finalizara el ensayo, pero por debajo de las tasas del grupo placebo a lo largo del período de seis años posteriores a la intervención. Estos resultados contrastan con el seguimiento post-ensayo del estudio CARET (237) que muestra que los suplementos de b-caroteno pueden causar daños más prolongados en algunas personas con alto riesgo de cáncer de pulmón. Esto último podría deberse a los cambios celulares causados por el b-caroteno - cambios que persisten incluso después que los niveles del nutriente en sangre retornan a la normalidad.
Los resultados tanto del ensayo como del seguimiento post-ensayo del estudio ATBC, en conjunción con los resultados del estudio CARET, proporcionan una fuerte evidencia del efecto adverso del uso de suplementos de b-caroteno sobre la incidencia de cáncer de pulmón en fumadores (238). Los posibles efectos preventivos del a- tocoferol sobre el cáncer de próstata requieren confirmación en otros ensayos en curso.
De estos grandes estudios clínicos surge que no se ha demostrado que los suplementos de b-caroteno sean perjudiciales para los no-fumadores (239). Esto se basa en parte en otro gran ensayo desarrollado casi al mismo tiempo, el Physicians' Health Study (Estudio de Salud en Médicos), de 12 años de duración con 22.000 médicos de sexo masculino, que encontró que 50 mg de b-caroteno (cerca de 25 veces la cantidad necesaria para proporcionar el requerimiento diario de vitamina A) tomados cada tres días, no tenía efecto (ni beneficio ni daño) sobre el riesgo de cáncer o enfermedad cardíaca en hombres sanos. En este estudio, el 11% de los participantes fueron fumadores y el 39% ex-fumadores (240-243). De manera similar, otro estudio de los complementos de b- caroteno fracasó en encontrar efecto alguno sobre el riesgo de cáncer en mujeres (244).
El Estudio sobre la Salud de los Médicos II (Physicians' Health Study II), dirigido por investigadores de la Harvard Medical School en Boston, evaluó la vitamina E, vitamina C, b-caroteno y un multivitamínico para la prevención primaria de enfermedades cardiovasculares, cáncer total y cáncer de próstata entre 15.000 médicos de sexo masculino de 50 años o más. El ensayo comenzó en 1997 y concluyó en diciembre de 2007. También se evaluó el efecto de estos agentes en pólipos de colon y cáncer de colon, cataratas (245), degeneración macular y deterioro cognitivo temprano (246) y dentro de este estudio se realizó un ensayo aleatorio de los suplementos de b- caroteno y la función cognitiva en hombres (247).
Asimismo, se efectuó un estudio aleatorizado, de doble ciego, de Fase III de los ensayos mediante la suplementación oral de b-caroteno en mujeres con alto grado de neoplasia intraepitelial cervical (248) y se demostró en un ensayo clínico controlado que la suplementación con a-tocopherol (vitamina E) y b-caroteno no afectaba el riesgo de gran aneurisma aórtico abdominal (249).
No se registró ningún efecto por la suplementación con b-caroteno sobre el riesgo de cáncer de piel no-melanoma entre hombres con bajos niveles plasmáticos de b-caroteno (250).
Seis ensayos clínicos aleatorios que examinaron la eficacia de los suplementos de b-caroteno y 25 estudios observacionales prospectivos determinando la asociación entre los carotenoides y el cáncer de pulmón fueron analizados mediante el uso de meta-análisis con efectos aleatorios (209), llegando a la conclusión que los suplementos de b-caroteno no se asociaron con una disminución en el riesgo de desarrollar cáncer de pulmón. Los resultados de los estudios prospectivos de cohorte sugirieron una asociación inversa entre los carotenoides y el cáncer de pulmón; sin embargo, la disminución en el riesgo es generalmente pequeña y no es estadísticamente significativa. Estas asociaciones inversas pueden ser el resultado de la función de las mediciones de carotenoides como un marcador de un estilo de vida más sana (mayor consumo de frutas y vegetales) o de la confusión residual por tabaquismo.
Se registraron los resultados de nueve ensayos controlados aleatorios. La suplementación con b-caroteno no presentó ningún efecto beneficioso en la prevención del cáncer, como cáncer de páncreas, cáncer colorrectal, cáncer de próstata, cáncer de mama, melanoma y cáncer de piel no melanoma. Por el contrario, se asoció con mayor riesgo no sólo de cáncer de pulmón, sino también de cáncer gástrico, en dosis de 20-30 mg/día de b-caroteno, en fumadores y trabajadores con asbestos (251).
Un meta-análisis para examinar los estudios sobre la suplementación en la prevención primaria o terciaria no mostró ningún beneficio en la supervivencia global, independientemente del tipo de tumor estudiado o del anti-oxidante evaluado (252). Los encargados de evaluar el efecto de los antioxidantes no médicos tomados durante los tratamientos contra el cáncer (quimioterapia o radioterapia) indicaron que si el objetivo de reducir los efectos secundarios a veces se consigue, el riesgo de la progresión tumoral y de aumentar la mortalidad, no deben desestimarse (252).
El meta-análisis de ensayos controlados aleatorios indicó que no hay evidencia clínica para apoyar el efecto general de prevención primaria y secundaria de estos suplementos antioxidantes sobre el cáncer. Más aún, el uso de éstos podría ser perjudicial para algunas personas (253).
Como hemos visto, los resultados de los ensayos clínicos han demostrado las limitaciones de los suplementos de b-caroteno, no de una dieta rica en b-caroteno y otros carotenoides (239). Muchos estudios apuntan a los beneficios en la salud por tal dieta, ya que comer frutas y verduras parece reducir el riesgo de cáncer en un 32- 44% (254) (255).
El estudio ATBC, el estudio CARET, el ensayo de Prevención Polip de antioxidantes (Antioxidant Polyp Prevention trial) y el estudio E3N brindaron evidencia que los efectos adversos de la suplementación con b-caroteno se correlacionan con la condición de fumador de los participantes del estudio. En contraste, el estudio de salud de médicos, el ensayo Linxian y un análisis conjunto de siete estudios epidemiológicos de cohorte no han apoyado esta evidencia. El modelo de cáncer de pulmón de hurón y ratón A/J se ha utilizado para investigar el mecanismo de interacción de b-caroteno con carcinógenos en el pulmón. Ambos modelos tienen ventajas y desventajas específicas. Hay una serie de hipótesis sobre la interacción de b-caroteno/ humo de tabaco, incluidas las alteraciones del metabolismo de los retinoides y las vías de señalización y la interacción con las enzimas CYP y la pro-oxidación/oxidación del DNA (256).
Los modelos animales demuestran consistentemente efectos negativos sólo en el hurón y después de la administración de b-caroteno en aceite de maíz en dosis farmacológicas. No se observaron efectos o incluso efectos de protección contra el humo o la exposición a carcinógenos cuando se aplicó b-caroteno en dosis fisiológicas o en combinación con las vitaminas C y E, ya sea como una mezcla o en una formulación estable. En conclusión, los estudios en humanos y en animales han demostrado que las circunstancias específicas, entre ellas fuerte tabaquismo, parecen influir en el efecto de la alta ingesta de b-caroteno. En poblaciones normales y saludables, que no fuman, hay evidencias de efectos beneficiosos (256).
Hay varias explicaciones posibles para estos resultados aparentemente contradictorios. Pues, como ya se mencionara, es posible que el consumo de caroteno como tal no esté relacionado al cáncer y que algunos factores no relacionados comunes a personas con una dieta alta en caroteno sean la causa de los beneficios observados en ensayos observacionales.
Otra posibilidad es que el b-caroteno por sí solo no sea efectivo y que los otros carotenos encontrados en frutas y verduras puedan ser más importantes para prevenir el cáncer que el b-caroteno. Se ha sugerido que el tomar suplementos de b-caroteno disminuye la captación en el cuerpo de otros carotenos benéficos y de ese modo provoca un efecto dañino (257). Como soporte de esta teoría, un estudio grande encontró que el consumo de frutas y verduras generalmente está asociado con un menor riesgo de cáncer pulmonar, pero cuando se toma b-caroteno, este efecto preventivo desaparece (254).
MECANISMOS POSIBLES
El b-caroteno mismo puede actuar como un anticancerígeno, pero sus productos oxidados pueden facilitar la carcinogénesis (258).
La respuesta carcinogénica a la suplementación con b-caroteno en altas dosis informada en los ensayos de intervención humana mencionados podría estar relacionada con la inestabilidad de la molécula de b-caroteno en el medio ambiente rico en radicales libres en los pulmones de fumadores de cigarrillos. Esto es especialmente posible porque el humo del cigarrillo también causa una disminución en los niveles tisulares de otros antioxidantes, como el ascorbato y el a-tocoferol, que normalmente tienen un efecto estabilizante de la forma no-oxidada del b-caroteno. La intervención nutricional mediante una combinación de antioxidantes (b-caroteno, a-tocoferol y vitamina C) como agentes anticarcinogénicos podría ser una forma adecuada de reducir el riesgo de cáncer de una manera racional y realista.
El potencial de los carotenoides para modular las enfermedades crónicas relacionadas con el tabaquismo está actualmente bajo investigación y se ha puesto un renovado interés en lograr un mejor entendimiento del mecanismo(s) de acción de los carotenoides en los sistemas biológicos expuestos al humo del tabaco. Los datos disponibles actualmente indican que, mientras que los carotenoides por sí solos no son dañinos, su interacción con el humo puede pasar de beneficiosa a perjudicial dependiendo de la dosis, del tipo de carotenoide, así como del medio ambiente biológico en el que actúan. Varios mecanismos se han propuesto para explicar ese cambio, como: a) cambios en el estado oxidativo de la célula, los cuales desequilibran la balanza de antioxidante-prooxidante del b-caroteno hacia un estado de prooxidante, b) modulación de los niveles de las proteínas claves involucradas en la regulación de la proliferación celular y la apoptosis, c) reducción del paso de la señal del ácido retinoico que desactiva la expresión RARb y estimula a AP-1, d) interferencia con la absorción de otros nutrientes en el mejor perfil antioxidante, e) formación de determinados productos específicos de oxidación de los carotenoides (259) (260).
Es importante tener en cuenta las evidencias disponibles en cultivos de células, modelos animales y humanos para una acción moduladora de los carotenoides en los efectos peligrosos del humo, así como las principales vías moleculares implicadas en este proceso (259) (260).
a) Factores relacionados con la proliferación de células epiteliales:
Estudios en modelos animales como el hurón, cuya absorción de b-caroteno es más similar a la de los seres humanos que la de la rata, han demostrado que altas dosis de suplementos de b-caroteno, particularmente en combinación con la exposición al humo de cigarrillos, desactivan al gen supresor de tumores RARb y estimulan a c-Fos, c-Jun y ciclina D1, que inducen la proliferación celular (261) (262) (263). El efecto neto de estos sistemas de modelos animales ha sido una fuerte respuesta proliferativa y la evidencia de carcinogénesis pulmonar (261). En particular, mientras que dosis altas de b-caroteno pueden activar la expresión de estas proteínas reguladoras del ciclo celular, los flavonoides son inhibidores (264). Estudios in vitro (265), en modelos de animales (264) y de alimentación en humanos (266) todos apoyan un rol importante para los flavonoides en la transducción de señales y en la regulación del ciclo celular, además de sus funciones antioxidantes ya indicadas, y funciones reguladoras de las enzimas de la fase I y II. Los suplementos CARET pudieron haber promovido la activación de c-Fos, c-Jun y ciclina D1, tal que incluso altos niveles de frutas y verduras ricas en flavonoides (por ejemplo: cebollas, frutos rojos) no fueron capaces de anular las fuertes señales proliferativas. Por otra parte, Salgo et al. (267) observaron que b-caroteno, su epóxido y derivados cetónicos, aumentaron la formación del aducto benzo(a)pireno in vitro.
b) Acción del b-caroteno sobre la actividad de las enzimas de la fase I y II:
Los estudios experimentales en modelos animales mostraron que dosis altas de suplementos de b-caroteno modulan la actividad de las enzimas de la fase I metabolizante de xenobióticos. Se informó una estimulación en 2-15 veces de la actividad de CYP1 A1 y CYP3 A1 en los pulmones de ratas tratadas con 500 mg de b-caroteno por kg de peso corporal, en comparación con ratas que recibieron placebo (268). CYP1 A1 y CYP3 A1 son enzimas de la fase I que convierten los numerosos agentes cancerígenos de los cigarrillos, incluidos los hidrocarburos aromáticos policíclicos (PAH: Polycyclic Aromatic Hydrocarbons), en electrófilos altamente reactivos, lo cual facilita su conjugación con moléculas hidrosolubles en la preparación para la eliminación del cuerpo. En el metabolismo normal, estos compuestos intermedios, que suelen ser más citotóxicos que los compuestos originales, son descontaminados y eliminados por las enzimas de la fase II que están activadas por las verduras crucíferas y frutas y hortalizas ricas en flavonoides (269) (270) (271). Si la actividad enzimática de la fase I y la subsiguiente concentración bioactivada de los carcinógenos se incrementaron varias veces por los suplementos CARET, entonces es concebible que una carga de sustrato excesivamente grande sature los sistemas enzimáticos de desintoxicación, permitiendo así la acumulación de compuestos intermedios carcinogénicos, aún frente a altos niveles de consumo de alimentos que activen a las enzimas de la fase II. Este escenario podría explicar los resultados nulos en el grupo de intervención de CARET incluso ante alto consumo de verduras crucíferas y frutos rojos.
c) Vitamina A/retinoides de señalización en el pulmón humano:
La vitamina A se utiliza como un término genérico para todos sus derivados con actividad biológica tipo retinol. El retinol, que da origen a la vitamina A, deriva de los carotenoides (provitamina A) y también directamente de la vitamina A pre-formada contenida en la dieta. El término "retinoide" es un descriptor genérico de compuestos estructuralmente relacionados con la vitamina A y los análogos sintéticos del retinol, con o sin actividad biológica. El ácido retinoico es el catabolito celular activo (272).
Tanto la vitamina A como los retinoides poseen aparentemente funciones preventivas del cáncer y se utilizaron en ensayos clínicos para reducir la incidencia de cáncer de pulmón en individuos de alto riesgo. Los resultados obtenidos han estado en contradicción con estudios prometedores tanto in vivo como in vitro. Por tanto, parece necesario desarrollar una mejor comprensión de las vías de señalización de vitamina A/retinoides en el pulmón (272). Con este objetivo, se resumen los conocimientos relevantes centrados en el pulmón.
d) El rol de los receptores retinoicos en la carcinogénesis pulmonar:
Varios estudios in vitro e in vivo han examinado los efectos positivos y negativos de los retinoides (análogos de la vitamina A) en las lesiones premalignas y malignas. Los retinoides se han utilizado como agentes quimiopreventivos y anticancerígenos debido a su función reguladora pleiotrópica en la diferenciación celular, el crecimiento, la proliferación y la apoptosis mediante la interacción con dos tipos de receptores nucleares: los receptores del ácido retinoico (RARs: RARA, RARB y RARG) y los receptores X retinoides (RXRs: RXRA, RXRB y RXRG). Se ha esclarecido gradualmente la función de los retinoides y sus vías de señalización y pueden explicar el fracaso de los estudios quimiopreventivos anteriores. Recientemente se han recopilado los conocimientos básicos sobre el rol de los receptores retinoides en la carcinogénesis pulmonar (273). La investigación de los receptores retinoides es importante y puede contribuir a establecer nuevas estrategias en la quimioprevención en pacientes de alto riesgo y en el tratamiento del cáncer de pulmón.
e) Efecto del b-caroteno sobre los otros carotenoides dietarios:
Así como hay una interacción entre b-caroteno y tabaquismo y evidencias de que los suplementos de b-caroteno podrían provocar que la enfermedad hepática alcohólica se desarrolle más rápidamente en personas que abusan del alcohol (274) (275), también el b-caroteno pudo haber afectado la biodisponibilidad de los otros carotenoides debido a que, como es sabido, los carotenoides compiten por su incorporación dentro de las micelas y la absorción posterior. Los resultados publicados anteriormente tanto del CARET como del ATBC no mostraron efectos adversos del b-caroteno en las concentraciones séricas de los otros carotenoides y ningún exceso de cualquier incidencia de cáncer de pulmón o mortalidad total entre los participantes en el quintil más alto de la concentración sérica basal de b-caroteno (231) (234) (276). Los estudios adicionales para examinar los efectos de los compuestos fitoquímicos dietarios, las dosis farmacológicas de los micronutrientes, como b-caroteno y los agentes conocidos causantes del cáncer de pulmón sobre la regulación del ciclo celular pueden brindar una mayor comprensión de estos mecanismos biológicos complejos y su relación con el riesgo de enfermedad (259).
Otros pocos estudios han tenido un número suficiente de casos de cáncer de pulmón estratificados mediante histología tumoral; dos de las tres investigaciones mostraron que los factores dietarios pueden tener diferentes asociaciones según los tipos histológicos celulares (277) (278) (279). Los factores dietarios pueden tener la capacidad protectora discriminante entre varios tipos de tumores.
Conclusión
Un gran número de estudios epidemiológicos observacionales han demostrado consistentemente que las personas que comen más frutas y verduras, que son ricas en carotenoides y las personas que tienen más altos niveles séricos de b-caroteno tienen menor riesgo de cáncer, especialmente cáncer de pulmón. En contraste con estas observaciones, los dos estudios de intervención humana ATBC y CARET que hemos mencionado, que usaron altas dosis de suplementos de b-caroteno registraron un aumento en el riesgo de cáncer de pulmón entre los fumadores. Otros estudios in vitro e in vivo han arrojado luz sobre este dilema con respecto a la actividad quimiopreventiva potencial del b-caroteno.
En particular, si se compara el licopeno con el b-caroteno, este último es un retinoide que posee en su estructura dos ciclos de 6 carbonos en sus extremos y sus productos de oxidación dan origen a sistemas cíclicos insaturados que finalmente conducen a sistemas aromáticos carcinogénicos del tipo PAH (hidrocarburo aromático policíclico), que como es conocido estabilizan los radicales libres, por lo que son causantes de varios tipos de cáncer. Esto no sucede con el licopeno, ya que debido a su estructura lineal, no tiene posibilidad de formar precursores de PAH.
Por todo lo descripto en este ítem se puede concluir que no siempre natural es sinónimo de salud o saludable. Como con cualquier sustancia química, el riesgo de intoxicación siempre está presente. Para aquellas sustancias de origen dietario esto es mucho más importante, particularmente cuando la imagen que generan es de inocuidad y de beneficio sin riesgo asociado.
AGRADECIMIENTOS
Los autores agradecen a CONICET y a la Universidad de Buenos Aires por el apoyo financiero; a MINCYT por la biblioteca electrónica. ABP y AAV son Investigadores de CONICET; EAB es Profesional Adjunto de CONICET.
CORRESPONDENCIA
PROF. DR. ARTURO A. VITALE
Av. Santa Fe 1970, 6 E,
C1123AAO CIUDAD AUTÓNOMA DE BUENOS AIRES, ARGENTINA
1. Gros EG, Pomilio AB, Seldes AM, Burton G. Introducción al estudio de los Productos Naturales. Serie Química, Monografía N° 30, Secretaría General de la Organización de Estados Americanos (OEA), Programa Regional de Desarrollo Científico y Tecnológico, 1985. [ Links ]
2. Stahl W, Sies H. Bioactivity and protective effects of natural carotenoids. Biochim Biophys Acta 2005; 1740 (2): 101-7. [ Links ]
3. O'Sullivan L, Ryan L, O'Brien N. Comparison of the uptake and secretion of carotene and xanthophyll carotenoids by Caco-2 intestinal cells. Br J Nutr 2007; 98 (1): 38-44. [ Links ]
4. Tadmor Y, King S, Levi A, Davis A, Meir A, Wasserman B, et al. Comparative fruit colouration in watermelon and tomato. Food Res Int 2005; 38 (8-9): 837-41. [ Links ]
5. Levy J, Sharoni Y. The functions of tomato lycopene and its role in human health. Herbal Gram 2004; (62): 49-56. [ Links ]
6. Clinton SK, Emenhiser C, Schwartz SJ, Bostwick DG, Williams AW, Moore BJ, et al. cis-trans Lycopene isomers, carotenoids, and retinol in the human prostate. Cancer Epidemiol Biomarkers Prev 1996; 5 (10): 823- 33. [ Links ]
7. Boileau AC, Merchen NR, Wasson K, Atkinson CA, Erdman Jr JW. cis-Lycopene is more bioavailable than translycopene in vitro and in vivo in lymph-cannulated ferrets. J Nutr 1999; 129 (6): 1176-81. [ Links ]
8. Stahl W, Sies H. Lycopene: a biologically important carotenoid for humans? Arch Biochem Biophys 1996; 336 (1): 1-9. [ Links ]
9. van Breemen RB, Pajkovic N. Multitargeted therapy of cancer by lycopene. Cancer Lett 2008; 269 (2): 339-51. [ Links ]
10. Hoppe PP, Kramer K, van den Berg H, Steenge G, van Vliet T. Synthetic and tomato-based lycopene have identical bioavailability in humans. Eur J Nutr 2003; 42 (5): 272-8. [ Links ]
11. Goñi I, Serrano J, Saura-Calixto F. Bioaccessibility of b- carotene, lutein, and lycopene from fruits and vegetables. J Agric Food Chem 2006; 54 (15): 5382-7. [ Links ]
12. Khachik F, Carvalho L, Bernstein PS, Muir GJ, Zhao DY, Katz NB. Chemistry, distribution, and metabolism of tomato carotenoids and their impact on human health. Exp Biol Med 2002; 227 (10): 845-51. [ Links ]
13. Campbell JK, Engelmann NJ, Lila MA, Erdman JW. Phytoene, phytobuene, and lycopene from tomato powder differentially accumulate in tissues of male Fisher 344 Rats. Nutr Res 2007; 27 (12): 794-801. [ Links ]
14. Hurst JS, Contreras JE, Siems WG, Van Kuijk FJ. Oxidation of carotenoids by heat and tobacco smoke. Biofactors 2004; 20 (1): 23-35. [ Links ]
15. Nara E, Hayashi H, Kotake M, Miyashita K, Nagao A. Acyclic carotenoids and their oxidation mixtures inhibit the growth of HL-60 human promyelocytic leukemia cells. Nutr Cancer 2001; 39 (2): 273-83. [ Links ]
16. Rao AV, Agarwal S. Bioavailability and in vivo antioxidant properties of lycopene from tomato products and their possible role in the prevention of cancer. Nutr Cancer 1998; 31 (3): 199-203. [ Links ]
17. Sies H, Stahl W. Lycopene: Antioxidant and biological effects and its bioavailability in the human. Proc Soc Exp Biol Med 1998; 218 (2): 121-4. [ Links ]
18. Toor RK, Savage GP. Changes in major antioxidant components of tomatoes during post-harvest storage. Food Chemistry 2006; 99 (4): 724-7. [ Links ]
19. Fraser PD, Enfissi EMA, Bramley PM. Genetic engineering of carotenoid formation in tomato fruit and the potential application of systems and synthetic biology approaches. Arch Biochem Biophys 2009; 483 (2): 196-204. [ Links ]
20.Markovic K, Hruškar M, Vahcic N. Lycopene content of tomato products and their contribution to the lycopene intake of Croatians. Nutr Res 2006; 26 (11): 556-60.
21. Periago MJ, Rincón F, Agüera MD, Ros G. Mixture approach for optimizing lycopene extraction from tomato and tomato products. J Agric Food Chem 2004; 52 (19): 5796-802. [ Links ]
22. Campbell JK, Canene-Adams K, Lindshield BL, Boileau TW-M, Clinton SK, Erdman Jr JW. Tomato phytochemicals and prostate cancer risk. J Nutr 2004; 134 (12 Suppl): 3486S-92S. [ Links ]
23. Abushita AA, Daood HG, Biacs PA. Change in carotenoids and antioxidant vitamins in tomato as a function of varietal and technological factors. J Agric Food Chem 2000; 48 (6): 2075-81. [ Links ]
24. Sahlin E, Savage GP, Lister CE. Investigation of the antioxidant properties of tomatoes after processing. J Food Compos Anal 2004; 17 (5): 635-47. [ Links ]
25. Capanoglu E, Beekwilder J, Boyacioglu D, Hall R, de Vos R. Changes in antioxidant and metabolite profiles during production of tomato paste. J Agric Food Chem 2008; 56 (3): 964-73. [ Links ]
26.Minoggio M, Bramati L, Simonetti P, Gardana C, Iemoli L, Santangelo E, et al. Polyphenol pattern and antioxidant activity of different tomato lines and cultivars. Ann Nutr Metab 2003; 47 (2): 64-9. [ Links ]
27. Toor RK, Lister CE, Savage GP. Antioxidant activities of New Zealand-grown tomatoes. Int J Food Sci Nutr 2005; 56 (8): 597-605. [ Links ]
28. Spencer JPE, Kuhnle GGC, Hajirezaei M, Mock HP, Sonnewald U, Rice-Evans C. The genotypic variation of the antioxidant potential of different tomato varieties. Free Radic Res 2005; 39 (9): 1005-16. [ Links ]
29. Frusciante L, Carli P, Ercolano MR, Pernice R, Di Matteo A, Fogliano V, et al. Antioxidant nutritional quality of tomato. Mol Nutr Food Res 2007; 51 (5): 609-17. [ Links ]
30. Dewanto V, Wu X, Adom KK, Liu RH. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant capacity. J Agric Food Chem 2002; 50 (10): 3010-4. [ Links ]
31. Chang C-H, Liu Y-C. Study on lycopene and antioxidant contents variations in tomatoes under air-drying process. J Food Sci 2007; 72 (9): E532-40. [ Links ]
32. Hwang E-S. Tomato-based products and lycopene in the prevention of cancer: Bioavailability and antioxidant properties. Cancer Prev Res 2005; 10 (2): 81-8. [ Links ]
33. Chen L, Stacewicz-Sapuntzakis M, Duncan C, Sharifi R, Ghosh L, van Breemen R, et al. Oxidative DNA damage in prostate cancer patients consuming tomato saucebased entrees as a whole-food intervention. J Natl Cancer Inst 2001; 93 (24): 1872-9. [ Links ]
34. Richelle M, Bortlik K, Liardet S, Hager C, Lambelet P, Baur M, et al. A food-based formulation provides lycopene with the same bioavailability to humans as that from tomato paste. J Nutr 2002; 132 (3): 404-8. [ Links ]
35. Tang FY, Cho HJ, Pai MH, Chen YH. Concomitant supplementation of lycopene and eicosapentaenoic acid inhibits the proliferation of human colon cancer cells. J Nutr Biochem 2009; 20 (6): 426-34. [ Links ]
36. Sharoni Y, Danilenko M, Dubi N, Ben-Dor A, Levy J. Carotenoids and transcription. Arch Biochem Biophys 2004; 430 (1): 89-96. [ Links ]
37.Wertz K, Siler U, Goralczyk R. Lycopene: modes of action to promote prostate health. Arch Biochem Biophys 2004; 430 (1): 127-34. [ Links ]
38.Mossine VV, Chopra P, Mawhinney TP. Interaction of tomato lycopene and ketosamine against rat prostate tumorigenesis. Cancer Res 2008; 68 (11): 4384-91. [ Links ]
39. Birt DF. Phytochemicals and cancer prevention: From epidemiology to mechanism of action. J Am Diet Assoc 2006; 106 (1): 20-1. [ Links ]
40. Venkateswaran V, Klotz LH, Ramani M, Sugar LM, Jacob LE, Nam RK, et al. A combination of micronutrients is beneficial in reducing the incidence of prostate cancer and increasing survival in the lady transgenic model. Cancer Prev Res 2009; 2 (5): 473-83. [ Links ]
41. Lean ME, Noroozi M, Kelly I, Burns J, Talwar D, Sattar N, et al. Dietary bavonols protect diabetic human lymphocytes against oxidative damage to DNA. Diabetes 1999; 48 (1): 176-81. [ Links ]
42. De S, Ganguly C, Das S. Natural dietary agents can protect against DMBA genotoxicity in lymphocytes as revealed by single cell gel electrophoresis assay. Teratog Carcinog Mutagen 2003; (Suppl 1): 71-8. [ Links ]
43. Di Mascio P, Kaiser S, Sies H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch Biochem Biophys 1989; 274 (2): 532-8. [ Links ]
44. Hadley CW, Clinton SK, Schwartz SJ. The consumption of processed tomato products enhances plasma lycopene concentrations in association with a reduced lipoprotein sensitivity to oxidative damage. J Nutr 2003; 133 (3): 727-32. [ Links ]
45. Porrini M, Riso P. Lymphocyte lycopene concentration and DNA protection from oxidative damage is increased in women after a short period of tomato consumption. J Nutr 2000; 130 (2): 189-92. [ Links ]
46. Breinholt V, Lauridsen ST, Daneshvar B, Jakobsen J. Dose-response effects of lycopene on selected drug-metabolizing and antioxidant enzymes in the rat. Cancer Lett 2000; 154 (2): 201-10. [ Links ]
47. Bhuvaneswari V, Velmurugan B, Nagini S. Induction of glutathione-dependent hepatic biotransformation enzymes by lycopene in the hamster cheek pouch carcinogenesis model. J Biochem Mol Biol Biophys 2002; 6 (4): 257-60. [ Links ]
48. Velmurugan B, Bhuvaneswari V, Burra UK, Nagini S. Prevention of N-methyl-N'-nitro-N-nitroso-guanidine and saturated sodium chloride-induced gastric carcinogenesis in Wistar rats by lycopene. Eur J Cancer Prev 2002; 11 (1): 19-26. [ Links ]
49. Voskuil DW, Vrieling A, van't Veer LJ, Kampman E, Rookus MA. The insulin-like growth factor system in cancer prevention: potential of dietary intervention strategies. Cancer Epidemiol Biomarkers Prev 2005; 14 (1): 195-203. [ Links ]
50. Kaaks R, Lukanova A. Energy balance and cancer: the role of insulin and IGF-I. Proc Nutr Soc 2001; 60 (1): 91-106. [ Links ]
51. Jones JI, Clemmons DR. Insulin-like growth factors and their binding proteins: biological actions. Endocr Rev 1995; 16 (1): 3-34. [ Links ]
52.Mucci LA, Tamimi R, Lagiou P, Trichopoulou A, Benetou V, Spanos E, et al. Are dietary inbuences on the risk of prostate cancer mediated through the insulin-like growth factor system? BJU Int 2001; 87 (9): 814-20. [ Links ]
53. Siler U, Barella L, Spitzer V, Schnorr J, Lein M, Goralczyk R, et al. Lycopene and vitamin E interfere with autocrine/paracrine loops in the Dunning prostate cancer model. FASEB J 2004; 18 (9): 1019-21. [ Links ]
54. Gunnell D, Oliver SE, Peters TJ, Donovan JL, Persad R, Maynard M, et al. Are diet-prostate cancer associations mediated by the IGF axis? A cross-sectional analysis of diet, IGF-I and IGFBP-3 in healthy middle-aged men. Br J Cancer 2003; 88 (11): 1682-6. [ Links ]
55. Liu C, Lian F, Smith DE, Russell RM, Wang XD. Lycopene supplementation inhibits lung squamous metaplasia and induces apoptosis via up-regulating insulin-like growth factor-binding protein 3 in cigarette smoke-exposed ferrets. Cancer Res 2003; 63 (12): 3138-44. [ Links ]
56. Kanagaraj P, Vijayababu MR, Ravisankar B, Anbalagan J, Aruldhas MM, Arunakaran J. Effect of lycopene on insulin- like growth factor-I, IGF binding protein-3 and IGF type-I receptor in prostate cancer cells. J Cancer Res Clin Oncol 2007; 133 (6): 351-9. [ Links ]
57. Karas M, Amir H, Fishman D, Danilenko M, Segal S, Nahum A, et al. Lycopene interferes with cell cycle progression and insulin-like growth factor I signaling in mammary cancer cells. Nutr Cancer 2000; 36 (1): 101-11. [ Links ]
58.Wang S, DeGroff VL, Clinton SK. Tomato and soy polyphenols reduce insulin-like growth factor-I-stimulated rat prostate cancer cell proliferation and apoptotic resistance in vitro via inhibition of intracellular signaling pathways involving tyrosine kinase. J Nutr 2003; 133 (7): 2367-76. [ Links ]
59. Herzog A, Siler U, Spitzer V, Seifert N, Denelavas A, Hunziker PB, et al. Lycopene reduced gene expression of steroid targets and inbammatory markers in normal rat prostate. FASEB J 2005; 19 (2): 272-4. [ Links ]
60. Ivanov NI, Cowell SP, Brown P, Rennie PS, Guns ES, Cox ME. Lycopene differentially induces quiescence and apoptosis in androgen-responsive and -independent prostate cancer cell lines. Clin Nutr 2007; 26 (2): 252- 63. [ Links ]
61. Renehan AG, Zwahlen M, Minder C, O'Dwyer ST, Shalet SM, Egger M. Insulin-like growth factor (IGF)-I, IGF binding protein-3, and cancer risk: systematic review and meta-regression analysis. Lancet 2004; 363 (9418): 1346-53. [ Links ]
62. Signorello LB, Kuper H, Lagiou P, Wuu J, Mucci LA, Trichopoulos D, et al. Lifestyle factors and insulin-like growth factor 1 levels among elderly men. Eur J Cancer Prev 2000; 9 (3): 173-8. [ Links ]
63. Holmes MD, Pollak MN, Willett WC, Hankinson SE. Dietary correlates of plasma insulin-like growth factor I and insulin-like growth factor binding protein 3 concentrations. Cancer Epidemiol Biomarkers Prev 2002; 11 (9): 852-61. [ Links ]
64. Vrieling A, Voskuil DW, Bueno de Mesquita HB, Kaaks R, van Noord PA, Keinan-Boker L, et al. Dietary determinants of circulating insulin-like growth factor (IGF)- I and IGF binding proteins 1, -2 and -3 in women in the Netherlands. Cancer Causes Control 2004; 15 (8): 787-96. [ Links ]
65. Tran CD, Diorio C, Berube S, Pollak M, Brisson J. Relation of insulin-like growth factor (IGF) I and IGF-binding protein 3 concentrations with intakes of fruit, vegetables, and antioxidants. Am J Clin Nutr 2006; 84 (6): 1518-26. [ Links ]
66. Vrieling A, Voskuil DW, Bonfrer JM, Korse CM, van Doorn J, Cats A, et al. Lycopene supplementation elevates circulating insulin-like growth factor binding protein- 1 and -2 concentrations in persons at greater risk of colorectal cancer. Am J Clin Nutr 2007; 86 (5): 1456-62. [ Links ]
67. Obermüller-Jevic UC, Olano-Martin E, Corbacho AM, Eiserich JP, van der Vliet A, Valacchi G, et al. Lycopene inhibits the growth of normal human prostate epithelial cells in vitro. J Nutr 2003; 133 (11): 3356-60. [ Links ]
68. Nahum A, Hirsch K, Danilenko M, Watts CK, Prall OW, Levy J, et al. Lycopene inhibition of cell cycle progression in breast and endometrial cancer cells is associated with reduction in cyclin D levels and retention of p27Kip1 in the cyclin E-cdk2 complexes. Oncogene 2001; 20 (26): 3428-36. [ Links ]
69. Stahl W, von Laar J, Martin HD, Emmerich T, Sies H. Stimulation of gap junctional communication: comparison of acyclo-retinoic acid and lycopene. Arch Biochem Biophys 2000; 373 (1): 271-4. [ Links ]
70. Aust O, Ale-Agha N, Zhang L, Wollersen H, Sies H, Stahl W. Lycopene oxidation product enhances gap junctional communication. Food Chem Toxicol 2003; 41 (10): 1399-407. [ Links ]
71. Forbes K, Gillette K, Sehgal I. Lycopene increases urokinase receptor and fails to inhibit growth or connexin expression in a metastatically passaged prostate cancer cell line: a brief communication. Exp Biol Med 2003; 228 (8): 967-71. [ Links ]
72. Livny O, Kaplan I, Reifen R, Polak-Charcon S, Madar Z, Schwartz B. Lycopene inhibits proliferation and enhances gap-junction communication of KB-1 human oral tumor cells. J Nutr 2002; 132 (12): 3754-9. [ Links ]
73. Durak I, Biri H, Avci A, Sözen S, Devrim E. Tomato juice inhibits adenosine deaminase activity in human prostate tissue from patient with prostate cancer. Nutr Res 2003; 23 (9): 1183-8. [ Links ]
74. van Breemen RB, Xu X, Viana MA, Chen L, Stacewicz- Sapuntzakis M, Duncan C, et al. Liquid chromatography-mass spectrometry of cis- and all-trans-lycopene in human serum and prostate tissue after dietary supplementation with tomato sauce. J Agric Food Chem 2002; 50 (8): 2214-9. [ Links ]
75. Cohn W, Thürmann P, Tenter U, Aebischer C, Schierle J, Schalch W. Comparative multiple dose plasma kinetics of lycopene administered in tomato juice, tomato soup or lycopene tablets. Eur J Nutr 2004; 43 (5): 304-12. [ Links ]
76. Burgess LC, Rice E, Fischer T, Seekins JR, Burgess TP, Sticka SJ, et al. Lycopene has limited effect on cell proliferation in only two of seven human cell lines (both cancerous and noncancerous) in an in vitro system with doses across the physiological range. Toxicol In Vitro 2008; 22 (5): 1297-300. [ Links ]
77. Gitenay D, Lyan B, Talvas J, Mazur A, Georgé S, Caris- Veyrat C, et al. Serum from rats fed red or yellow tomatoes induces Connexin43 expression independently from lycopene in a prostate cancer cell line. Biochem Biophys Res Commun 2007; 364 (3): 578-82. [ Links ]
78. Salman H, Bergman M, Djaldetti M, Bessler H. Lycopene affects proliferation and apoptosis of four malignant cell lines. Biomed Pharmacother 2007; 61 (6): 366-9. [ Links ]
79. Clinton SK. Tomatoes or lycopene: a role in prostate carcinogenesis? J Nutr 2005; 135 (Suppl): 2057S-9S. [ Links ]
80. Boveris DL, Boveris A. Oxygen delivery to the tissues and mitochondrial respiration. Front Biosci 2007; 12 (3): 1014-23. [ Links ]
81. Valdez LB, Lores Arnaiz S, Bustamante J, Alvarez S, Costa LE, Boveris A. Free radical chemistry in biological systems. Biol Res 2000; 33 (2): 65-70. [ Links ]
82. Valdez LB, Boveris A. Mitochondrial nitric oxide synthase, a voltage-dependent enzyme, is responsible for nitric oxide diffusion to cytosol. Front Biosci 2007; 12 (3): 1210-9. [ Links ]
83. Khan N, Afaq F, Mukhtar H. Cancer chemoprevention through dietary antioxidants: progress and promise. Antiox Redox Signal 2008; 10 (3): 475-510. [ Links ]
84. Giovannucci E. Tomatoes, tomato-based products, lycopene, and cancer: Review of the epidemiologic literature. J Natl Cancer Inst 1999; 91 (4): 317-31. [ Links ]
85. Upritchard J, Sutherland W, Mann J. Effect of supplementation with tomato juice, vitamin E, and vitamin C on LDL oxidation and products of inflammatory activity in type 2 diabetes. Diabetes Care 2000; 23 (6): 733-8. [ Links ]
86. Krinsky NI, Johnson EJ. Carotenoid actions and their relation to health and disease. Mol Aspects Med 2005; 26 (6): 459-516. [ Links ]
87. Böhm V, Bitsch R. Intestinal absortion of lycopene from different matrices and interactions to other carotenoids, the lipid status, and the antioxidant capacity of human plasma. Eur J Nutr 1999; 38 (3): 118-25. [ Links ]
88. Fridovich I. Superoxide radical and superoxide dismutases. Annu Rev Biochem 1995; 64: 97-112. [ Links ]
89.Muradiana KK, Utko NA, Fraifeld V, Mozzhukhina TG, Pishel IN, Litoshenko AY. Superoxide dismutase, catalase and glutathione peroxidase activities in the liver of young and old mice: linear regression and correlation. Arch Gerontol Geriatr 2002; 35 (3): 205-11. [ Links ]
90. Navarro A, Gómez C, Sánchez-Pino MJ, González H, Bández MJ, Boveris AD, et al. Vitamin E at high doses improves survival, neurological performance, and brain mitochondrial function in aging male mice. Am J Physiol- Regul Integr Comp Physiol 2005; 289 (5): R1392-9. [ Links ]
91. Nishino H, Tokuda H, Satomi Y, Masuda M, Osaka Y, Yogosawa S, et al. Cancer prevention by antioxidants. Biofactors 2004; 22 (1-4): 57-61. [ Links ]
92. Eichler O, Sies H, Stahl W. Divergent optimum levels of lycopene, b-carotene and lutein protecting against UVB irradiation in human fibroblasts. Photochem Photobiol 2002; 75 (5): 503-6. [ Links ]
93. Olmedilla B, Granado F, Southon S, Wright AJ, Blanco I, Gil-Martinez E, et al. A European multicentre, placebo-controlled supplementation study with a-tocopherol, carotene-rich palm oil, lutein or lycopene: analysis of serum responses. Clin Sci (Lond) 2002; 102 (4): 447-56. [ Links ]
94. Rao AV, Balachadran B. Role of oxidative stress and antioxidants in neurodegenerative diseases. Review. Nutr Neurosci 2002; 5 (5): 291-309. [ Links ]
95. Lam PY, Yin F, Hamilton RT, Boveris A, Cadenas E. Elevated neuronal nitric oxide synthase expression during ageing and mitochondrial energy production. Free Radic Res 2009; 43 (5): 431-9. [ Links ]
96. Valdez LB, Zaobornyj T, Boveris A. Functional activity of mitochondrial nitric oxide synthase. Methods Enzymol 2005; 396: 444-55. [ Links ]
97. Navarro A, Boveris A. Mitochondrial nitric oxide synthase, mitochondrial brain dysfunction in aging, and mitochondria- targeted antioxidants. Review. Adv Drug Deliv Rev 2008; 60 (13-14): 1534-44. [ Links ]
98. Navarro A, Boveris A, Bández MJ, Sánchez-Pino MJ, Gómez C, Muntané G, et al. Human brain cortex: mitochondrial oxidative damage and adaptive response in Parkinson disease and in dementia with Lewy bodies. Free Radic Biol Med 2009; 46 (12): 1574-80. [ Links ]
99. Serra JA, Domínguez RO, Marschoff ER, Guareschi EM, Famulari AL, Boveris A. Systemic oxidative stress associated with the neurological diseases of aging. Neurochem Res 2009 Jun 2 [Epub ahead of print]. PMID: 19488852. [ Links ]
100.McNulty H, Jacob RF, Mason RP. Biologic activity of carotenoids related to distinct membrane physicochemical interactions. Am J Cardiol 2008; 101 (10A): 20D-9D. [ Links ]
101. Devaraj S, Mathur S, Basu A, Aung HH, Vasu VT, Meyers S, et al. A dose-response study on the effects of purified lycopene supplementation on biomarkers of oxidative stress. J Am Coll Nutr 2008; 27 (2): 267-73. [ Links ]
102. Rao CV, Indranie C, Simi B, Manning PT, Connor JR, Reddy BS. Chemopreventive properties of a selective inducible nitric oxide synthase inhibitor in colon carcinogenesis, administered alone or in combination with celecoxib, a selective cyclooxygenase-2 inhibitor. Cancer Res 2002; 62 (1): 165-70. [ Links ]
103. Farinati F, Cardin R, Degan P, De Maria N, Floyd RA, Van Thiel DH, et al. Oxidative DNA damage in circulating leukocytes occurs as an early event in chronic HCV infection. Free Radic Biol Med 1999; 27 (11-12): 1284-91. [ Links ]
104. Bowen P, Chen L, Stacewicz-Sapuntzakis M, Sharifi R, Ghosh L, van Breemen R, et al. Tomato sauce supplementation and prostate cancer: lycopene accumulation and modulation of biomarkers of carcinogenesis. Exp Biol Med 2002; 227 (10): 886-93. [ Links ]
105. Erdman JW Jr, Ford NA, Lindshield BL. Are the health attributes of lycopene related to its antioxidant function? Arch Biochem Biophys 2009; 483 (2): 229-35. [ Links ]
106. Hu K-Q, Liu C, Ernst H, Krinsky NI, Russell RM, Wang X-D. The biochemical characterization of ferret carotene-9',10'-monooxygenase catalyzing cleavage of carotenoids in vitro and in vivo. J Biol Chem 2006; 281 (28): 19327-38. [ Links ]
107. Lindshield BL, Canene-Adams K, Erdman Jr JW. Lycopenoids: are lycopene metabolites bioactive? Arch Biochem Bioph 2007; 458 (2): 136-40. [ Links ]
108. Steinberg FM, Chait A. Antioxidant vitamin supplementation and lipid peroxidation in smokers. Am J Clin Nutr 1998; 68 (2): 319-27. [ Links ]
109. Visioli F, Riso P, Grande S, Galli C, Porrini M. Protective activity of tomato products on in vivo markers of lipid oxidation. Eur J Nutr 2003; 42 (4): 201-6. [ Links ]
110. Porrini M, Riso P, Brusamolino A, Berti C, Guarnieri S, Visioli F. Daily intake of a formulated tomato drink affects carotenoid plasma and lymphocyte concentrations and improves cellular antioxidant protection. Br J Nutr 2005; 93 (1): 93-9. [ Links ]
111. Hininger IA, Meyer-Wenger A, Moser U, Wright A, Southon S, Thurnham D, et al. No significant effects of lutein, lycopene or b-carotene supplementation on biological markers of oxidative stress and LDL oxidizability in healthy adult subjects. J Am Coll Nutr 2001; 20 (3): 232-8. [ Links ]
112. Briviba K, Schnäbele K, Rechkemmer G, Bub A. Supplementation of a diet low in carotenoids with tomato or carrot juice does not affect lipid peroxidation in plasma and feces of healthy men. J Nutr 2004; 134 (5): 1081-3. [ Links ]
113. Riso P, Visioli F, Erba D, Testolin G, Porrini M. Lycopene and vitamin C concentrations increased in plasma and lymphocytes after tomato intake. Effects on cellular antioxidant protection. Eur J Clin Nutr. 2004; 58 (10): 1350-8. [ Links ]
114. Porrini M, Riso P, Oriani G. Spinach and tomato consumption increases lymphocyte DNA resistance to oxidative stress but this is not related to cell carotenoid concentrations. Eur J Nutr 2002; 41 (3): 95-100. [ Links ]
115. Canene-Adams K, Campbell JK, Zaripheh S, Jeffery EH, Erdman JW. The tomato as a functional food. J Nutr 2005; 135 (5): 1226-30. [ Links ]
116. Fuhrman B, Volkova N, Rosenblat M, Aviram M. Lycopene synergistically inhibits LDL oxidation in combination with vitamin E, glabridin, rosmarinic acid, carnosic acid, or garlic. Antiox Redox Signal 2000; 2 (3): 491-506. [ Links ]
117. Shi J, Kakuda Y, Yeung D. Antioxidative properties of lycopene and other carotenoids from tomatoes: synergistic effects. Biofactors 2004; 21 (1-4): 203-10. [ Links ]
118. Stewart AJ, Bozonnet S, Mullen W, Jenkins GI, Lean MEJ, Crozier A. Occurrence of flavonols in tomatoes and tomato-based products. J Agric Food Chem 2000; 48 (7): 2663-9. [ Links ]
119. Bovy A, Schijlen E, Hall RD. Metabolic engineering of flavonoids in tomato (Solanum lycopersicum): the potential for metabolomics. Metabolomics 2007; 3 (3): 399-412. [ Links ]
120. Torres CA, Davies NM, Yañez JA, Andrewa K. Disposition of selected bavonoids in fruit tissues of various tomato (Lycopersicon esculentum Mill.) genotypes. J Agric Food Chem 2005; 53 (24): 9536-43. [ Links ]
121. Re R, Mishra GD, Thane CW, Bates CJ. Tomato consumption and plasma lycopene concentration in people aged 65 and over in a British National Survey. Eur J Clin Nutr 2003; 57 (12): 1545-54. [ Links ]
122.Watanabe S, Zhuo XG, Kimira M. Food safety and epidemiology: new database of functional food factors. Biofactors 2004; 22 (1-4): 213-9. [ Links ]
123.Madrid A E, Vásquez Z D, Leyton A F, Mandiola C, Escobar F JA. El consumo de Lycopersicum esculentum podría aumentar lipoproteínas de alta densidad (HDL) y disminuir el estrés oxidativo a corto plazo. Rev Méd Chile 2006; 134 (7): 855-62. [ Links ]
124. Periago MJ, Garcia-Alonso J, Jacob K, Olivares AB, Bernal MJ, Iniesta MD, et al. Bioactive compounds, folates and antioxidant properties of tomatoes (Lycopersicum esculentum) during vine ripening. Int J Food Sci Nutr 2008 Mar 12: 1-15. [Epub ahead of print]. PMID: 18608536 [ Links ]
125. Gitenay D, Lyan B, Rambeau M, Mazur A, Rock E. Comparison of lycopene and tomato effects on biomarkers of oxidative stress in vitamin E deficient rats. Eur J Nutr 2007; 46 (8): 468-75. [ Links ]
126. Shen YC, Chen SL, Zhuang SR, Wang CK. Contribution of tomato phenolics to suppression of COX-2 expression in KB cells. Food Sci 2008; 73 (1): C1-10. [ Links ]
127. Jacob K, Periago MJ, Böhm V, Berruezo GR. Influence of lycopene and vitamin C from tomato juice on biomarkers of oxidative stress and inflammation. Br J Nutr 2008; 99 (1): 137-46. [ Links ]
128.Morisco F, Vitaglione P, Carbone A, Stingo S, Scarpati S, Ascione A, et al. Tomato-based functional food as interferon adjuvant in HCV eradication therapy. J Clin Gastroenterol 2004; 38 (6 Suppl): S118-20. [ Links ]
129. Vitaglione P, Fogliano V, Stingo S, Scalfi L, Caporaso N, Morisco F. Development of a tomato-based food for special medical purposes as therapy adjuvant for patients with HCV infection. Eur J Clin Nutr 2007; 61 (7): 906-15. [ Links ]
130. Hoelzl C, Knasmüller S, Misík M, Collins A, Dusinská M, Nersesyan A. Use of single cell gel electrophoresis assays for the detection of DNA-protective effects of dietary factors in humans: recent results and trends. Mutat Res 2009; 681 (1): 68-79. [ Links ]
131. Conn PF, Schalch W, Truscott TG. The singlet oxygen and carotenoid interactions. J Photochem Photobiol B Biol 1991; 11 (1): 41-7. [ Links ]
132.Miller NJ, Sampson J, Candeias LP, Bramley PM, Rice- Evans CA. Antioxidant activities of carotenes and xanthophylls. FEBS Lett 1996; 384 (3): 240-2. [ Links ]
133. Hirayama O, Nakamura K, Hamada S, Kobayasi Y. Singlet oxygen quenching ability of naturally occurring carotenoids. Lipids 1994; 29 (2): 149-50. [ Links ]
134. Rao AV, Agarwal S. Role of lycopene as antioxidant carotenoid in the prevention of chronic diseases: a review. Nutr Res 1999; 19 (2): 305-23. [ Links ]
135. Krutovskikh V, Asamoto M, Takasuka N, Murakoshi M, Nishino H, Tsuda H. Differential dose-dependent effects of a-, b-carotenes and lycopene on gap-junctional intercellular communication in rat liver in vivo. Jpn J Cancer Res 1997; 88 (12): 1121-4. [ Links ]
136. Abraham CM. Moléculas de adhesión. Importancia en la respuesta inmune e inflamatoria. Rev Cubana Hematol Inmunol Hemoter 2006; 22 (2): 1-15. [ Links ]
137. Blum A, Monir M, Khazim K, Peleg A, Blum N. Tomatorich (Mediterranean) diet does not modify inflammatory markers. Clin Invest Med 2007; 30 (2): E70-4. [ Links ]
138. Hung CF, Huang TF, Chen BH, Shieh JM, Wu PH, Wu WB. Lycopene inhibits TNF-a-induced endothelial ICAM-1 expression and monocyte-endothelial adhesion. Eur J Pharmacol 2008; 586 (1-3): 275-82. [ Links ]
139. Li J, Yen C, Liaw D, Podsypanina K, Bose S, Wang SI, et al. PTEN, a putative protein tyrosine phosphatase gene mutated in human brain, breast, and prostate cancer. Science 1997; 275 (5308): 1943-7. [ Links ]
140. Sarker D, Reid AH, Yap TA, de Bono JS. Targeting the PI3K/AKT pathway for the treatment of prostate cancer. Clin Cancer Res 2009; 15 (15): 4799-805. [ Links ]
141. Tokunaga E, Oki E, Egashira A, Sadanaga N, Morita M, Kakeji Y, et al. Deregulation of the Akt pathway in human cancer. Curr Cancer Drug Targets 2008; 8 (1): 27-36. [ Links ]
142. Tang FY, Shih CJ, Cheng LH, Ho HJ, Chen HJ. Lycopene inhibits growth of human colon cancer cells via suppression of the Akt signaling pathway. Mol Nutr Food Res 2008; 52 (6): 646-54. [ Links ]
143.Müller K, Carpenter KL, Challis IR, Skepper JN, Arends MJ. Carotenoids induce apoptosis in the T-lymphoblast cell line Jurkat E6.1. Free Radic Res 2002; 36 (7): 791-802. [ Links ]
144. Kobayashi T, Iijima K, Mitamura T, Toriizuka K, Cyong JC, Nagasawa H. Effects of lycopene, a carotenoid, on intrathymic T cell differentiation and peripheral CD4/CD8 ratio in a high mammary tumor strain of SHN retired mice. Anticancer Drugs 1996; 7 (2): 195-8. [ Links ]
145. Bessler H, Salman H, Bergman M, Alcalay Y, Djaldetti M. In vitro effect of lycopene on cytokine production by human peripheral blood mononuclear cells. Immunol Invest 2008; 37 (3): 183-90. [ Links ]
146. Espondaburu OR, Fara Hunt VA, Ocampo LI. El proceso aterogénico y su desarrollo en las enfermedades autoinmunes. Acta Bioquím Clín Latinoam 2004; 38 (2): 181-92. [ Links ]
147.Willcox JK, Catignani GL, Lazarus S. Tomatoes and cardiovascular health. Review. Crit Rev Food Sci Nutr 2003; 43 (1): 1-18. [ Links ]
148. Blum A, Monir M, Wirsansky I, Ben-Arzi S. The beneficial effects of tomatoes. Eur J Int Medicine 2005; 16 (6): 402-4. [ Links ]
149. Blum A, Merei M, Karem A, Blum N, Ben-Arzi S, Wirsansky I, et al. Effects of tomatoes on the lipid profile. Clin Invest Med 2006; 29 (5): 298-300. [ Links ]
150. Bose KS, Agrawal BK. Effect of long term supplementation of tomatoes (cooked) on levels of antioxidant enzymes, lipid peroxidation rate, lipid profile and glycated haemoglobin in Type 2 diabetes mellitus. West Indian Med J 2006; 55 (4): 274-8. [ Links ]
151. Bose KS, Agrawal BK. Effect of lycopene from tomatoes (cooked) on plasma antioxidant enzymes, lipid peroxidation rate and lipid profile in grade-I hypertension. Ann Nutr Metab 2007; 51 (5): 477-81. [ Links ]
152. Silaste ML, Alfthan G, Aro A, Kesäniemi YA, Hörkkö S. Tomato juice decreases LDL cholesterol levels and increases LDL resistance to oxidation. Br J Nutr 2007; 98 (6): 1251-8. [ Links ]
153. Shen YC, Chen SL, Wang CK. Contribution of tomato phenolics to antioxidation and down-regulation of blood lipids. J Agric Food Chem 2007; 55 (16): 6475-81. [ Links ]
154. Hsu YM, Lai CH, Chang CY, Fan CT, Chen CT, Wu CH. Characterizing the lipid-lowering effects and antioxidant mechanisms of tomato paste. Biosci Biotechnol Biochem 2008; 72 (3): 677-85. [ Links ]
155. Lo HM, Hung CF, Tseng YL, Chen BH, Jian JS, Wu WB. Lycopene binds PDGF-BB and inhibits PDGF-BB-induced intracellular signaling transduction pathway in rat smooth muscle cells. Biochem Pharmacol 2007; 74 (1): 54-63. [ Links ]
156. Bose KS, Agrawal BK. Effect of lycopene from cooked tomatoes on serum antioxidant enzymes, lipid peroxidation rate and lipid profile in coronary heart disease. Singapore Med J 2007; 48 (5): 415-20. [ Links ]
157.Wood LG, Garg ML, Powell H, Gibson PG. Lycopene-rich treatments modify noneosinophilic airway inflammation in asthma: proof of concept. Free Radic Res 2008; 42 (1): 94-102. [ Links ]
158. Sung WS, Lee IS, Lee DG. Damage to the cytoplasmic membrane and cell death caused by lycopene in Candida albicans. J Microbiol Biotechnol 2007; 17 (11): 1797-804. [ Links ]
159. Giovannucci E, Ascherio A, Rim EB, Stampfer MJ, Colditz GA, Willet WC. Intake of carotenoids and retinol in relation to risk of prostate cancer. J Natl Cancer Inst 1995; 87 (23): 1767 -76. [ Links ]
160. Schmitz-Dräger BJ, Eichholzer M, Beiche B, Ebert T. Nutrition and prostate cancer. (Review). Urol Int 2001; 67 (1): 1-11. [ Links ]
161. Stacewicz-Sapuntzakis M, Bowen PE. Role of lycopene and tomato products in prostate health. Biochim Biophys Acta 2005; 1740 (2): 202-5. [ Links ]
162. Kamangar F, Dores GM, Anderson WF. Patterns of cancer incidence, mortality, and prevalence across five continents: defining priorities to reduce cancer disparities in different geographic regions of the world. J Clin Oncol 2006; 24 (14): 2137-50. [ Links ]
163.Mosher CE, Sloane R, Morey MC, Snyder DC, Cohen HJ, Miller PE, et al. Associations between lifestyle factors and quality of life among older long-term breast, prostate, and colorectal cancer survivors. Cancer 2009 Jul 27. [Epub ahead of print]. PMID: 19637244. [ Links ]
164.Moore SC, Peters TM, Ahn J, Park Y, Schatzkin A, Albanes D, et al. Age-specific physical activity and prostate cancer risk among white men and black men. Cancer 2009 Jul 30. [Epub ahead of print]. PMID: 19645029. [ Links ]
165. Freedland SJ, Isaacs WB, Explaining racial differences in prostate cancer in the United States: sociology or biology? Prostate 2005; 62 (3): 243-52. [ Links ]
166. Pathak SK, Sharma RA, Mellon JK. Chemoprevention of prostate cancer by diet-drived antioxidant agents and hormonal manipulation. Int J Oncol 2003; 22 (1): 5-13. [ Links ]
167. Goetzl MA, Holzbeierlein JM, Finasteride as a chemopreventive agent in prostate cancer: Impact of the prostate cancer prevention trial on urologic practice. Nat Clin Pract Urol 2006; 3 (8): 422-9. [ Links ]
168. van Weerden WM, Bangma C, de Wit R, Human xenograft models as useful tools to assess the potential of novel therapeutics in prostate cancer. Br J Cancer 2009; 100 (1): 13-8. [ Links ]
169. Damber J-E, Aus G. Prostate cancer. Lancet 2008; 371 (9625): 1710-21. [ Links ]
170. Chan JM, Gann PH, Giovannucci EL. Role of diet in prostate cancer development and progression. J Clin Oncol 2005; 23 (32): 8152-60. [ Links ]
171. Theobald S. Ernährung und Prostatakrebs. Was ist wissenschaftlich gesichert? [Nutrition and prostate cancer- what is the scientific evidence?]. Med Monatsschr Pharm 2006; 29 (10): 371-7. [ Links ]
172. Cohen LA. Nutrition and prostate cancer: a review. Ann N Y Acad Sci 2002; 963: 148-55. [ Links ]
173. Cohen LA. A review of animal model studies of tomato carotenoids, lycopene, and cancer chemoprevention. Exp Biol Med (Maywood) 2002; 227 (10): 864-8. [ Links ]
174. Fleshner N, Zlotta AR. Prostate cancer prevention: past, present, and future. Review. Cancer 2007; 110 (9): 1889-99. [ Links ]
175. Giovannucci E, Clinton SK. Tomatoes, lycopene, and prostate cancer. Proc Soc Exp Biol Med 1998; 218 (2): 129-39. [ Links ]
176. Giovannucci E. A review of epidemiologic studies of tomatoes, lycopene, and prostate cancer. Exp Biol Med 2002; 227 (10): 852-9. [ Links ]
177. Imaida K, Tamano S, Kato K, Ikeda Y, Asamoto M, Takahashi S, et al. Lack of chemopreventive effects of lycopene and curcumin on experimental rat prostate carcinogenesis. Carcinogenesis 2001; 22 (3): 467-72. [ Links ]
178. Nikoli MC, Anese M, Parpinel MT, Francesci S, Lerici CR. Loss and/or formation of antioxidants during food processing and storage. Cancer Lett 1997; 114 (1-2): 71-4. [ Links ]
179. Gahler S, Otto K, Böhm V. Alterations of vitamin C, total phenolics, and antioxidant capacity as affected by processing tomatoes to different products. J Agric Food Chem 2003; 51 (27): 7962-8. [ Links ]
180. Boileau TW-M, Liao Z, Kim S, Lemeshow S, Erdman JW, Clinton SK. Prostate carcinogenesis in N-methyl-N-nitrosourea (NMU)-testosterone-treated rats fed tomato powder, lycopene, or energy-restricted diets. J Natl Cancer Inst 2003; 95 (21): 1578-86. [ Links ]
181. Basu A, Imrhan V. Tomatoes versus lycopene in oxidative stress and carcinogenesis: conclusions from clinical trials. Eur J Clin Nutr 2007; 61 (3): 295-303. [ Links ]
182. Finot P-A. Historical perspective of the Maillard reaction in food science. Ann N Y Acad Sci 2005; 1043: 1-8. [ Links ]
183. Glinsky GV, Price JE, Glinsky VV, Mossine VV, Kiriakova G, Metcalf JB. Inhibition of human breast cancer metastasis in nude mice by synthetic glycoamines. Cancer Res 1996; 56 (23): 5319-24. [ Links ]
184. Glinsky VV, Glinsky GV, Glinskii OV, Huxley VH, Turk JR, Mossine VV, et al. Intravascular metastatic cancer cell homotypic aggregation at the sites of primary attachment to the endothelium. Cancer Res 2003; 63 (13): 3805-11. [ Links ]
185. Glinsky GV, Mossine VV, Price JE, Bielenberg D, Glinsky VV, Ananthaswamy HN, et al. Inhibition of colony formation in agarose of metastatic human breast carcinoma and melanoma cells by synthetic glycoamine analogs. Clin Exp Metastasis 1996; 14 (3): 253-67. [ Links ]
186. Canene-Adams K, Lindshield BL, Wang S, Jeffery EH, Clinton SK, Erdman JW Jr. Combinations of tomato and broccoli enhance antitumor activity in dunning r3327- h prostate adenocarcinomas. Cancer Res 2007; 67 (2): 836-43. [ Links ]
187. Venkateswaran V, Fleshner NE, Sugar LM, Klotz LH. Antioxidants block prostate cancer in lady transgenic mice. Cancer Res 2004; 64 (16): 5891-6. [ Links ]
188. Sengupta A, Ghosh S, Das S. Modulatory inbuence of garlic and tomato on cyclooxygenase-2 activity, cell proliferation and apoptosis during azoxymethane induced colon carcinogenesis in rat. Cancer Lett 2004; 208 (2): 127-36. [ Links ]
189. Bhuvaneswari V, Abraham SK, Nagini S. Combinatorial antigenotoxic and anticarcinogenic effects of tomato and garlic through modulation of xenobiotic-metabolizing enzymes during hamster buccal pouch carcinogenesis. Nutrition 2005; 21 (6): 726-31. [ Links ]
190.Mossine VV, Mawhinney TP. N-(1-Deoxy-D-fructos-1-yl)- L-histidine ("D-fructose-L-histidine"): a potent copper chelator from tomato powder. J Agric Food Chem 2007; 55 (25): 10373-81. [ Links ]
191. Bosland MC, McCormick DL. Antioxidant supplementation and cancer prevention. J Amer Med Assoc 2009; 301 (18): 1877-8. [ Links ]
192. DeWeese TL, Hruszkewycz AM, Marnett LJ. Oxidative stress in chemoprevention trials. Urology 2001; 57 (4A): 137-40. [ Links ]
193. Nelson WG, DeWeese TL, DeMarzo AM. The diet, prostate inflammation, and the development of prostate cancer. Cancer Metastasis Rev 2002; 21 (1): 3-16. [ Links ]
194. Siddiqui IA, Adhami VM, Saleem M, Mukhtar H. Beneficial effects of tea and its polyphenols against prostate cancer. Mol Nutr Food Res 2006; 50 (2): 130-43. [ Links ]
195. Vinson JA, Hao Y, Su X, Zubik L. Phenol antioxidant quantity and quality in foods: vegetables. J Agric Food Chem 1998; 46 (9): 3630-4. [ Links ]
196. Liu A, Pajkovic N, Pang Y, Zhu D, Calamini B, Mesecar AL, et al. Absorption and subcellular localization of lycopene in human prostate cancer cells. Mol Cancer Ther 2006; 5 (11): 2879-85. [ Links ]
197. Sengupta A, Ghosh S, Das RK, Bhattacharjee S, Bhattacharya S. Chemopreventive potential of diallylsulfide, lycopene and theaflavin during chemically induced colon carcinogenesis in rat colon through modulation of cyclooxygenase-2 and inducible nitric oxide synthase pathways. Eur J Cancer Prev 2006; 15 (4): 301-5. [ Links ]
198. Kim JM, Araki S, Kim DJ, Park CB, Takasuka N, Baba- Toriyama H, et al. Chemopreventive effects of carotenoids and curcumins on mouse colon carcinogenesis after 1,2-dimethylhydrazine initiation. Carcinogenesis 1998; 19 (1): 81-5. [ Links ]
199. Narisawa T, Fukaura Y, Hasebe M, Ito M, Aizawa R, Murakoshi M, et al. Inhibitory effects of natural carotenoids, a-carotene, b-carotene, lycopene and lutein, on colonic aberrant crypt foci formation in rats. Cancer Lett 1996; 107 (1): 137-42. [ Links ]
200. Narisawa T, Fukaura Y, Hasebe M, Nomura S, Oshima S, Sakamoto H, et al. Prevention of N-methylnitrosourea-induced colon carcinogenesis in F344 rats by lycopene and tomato juice rich in lycopene. Jpn J Cancer Res 1998; 89 (10): 1003-8. [ Links ]
201. Raju J, Swamy MV, Cooma I, Patlolla JM, Pittman B, Reddy BS, et al. Low doses of b-carotene and lutein inhibit AOM-induced rat colonic FCA formation but high doses augment FCA incidence. Int J Cancer 2005; 113 (5): 798-802. [ Links ]
202. Choi SY, Park JH, Kim JS, Kim MK, Aruoma OI, Sung MK. Effects of quercetin and b-carotene supplementation on azoxymethane-induced colon carcinogenesis and inflammatory responses in rats fed with high-fat diet rich in omega-6 fatty acids. Biofactors 2006; 27 (1-4): 137-46. [ Links ]
203.Walfisch S, Walfisch Y, Kirilov E, Linde N, Mnitentag H, Agbaria R, et al. Tomato lycopene extract supplementation decreases insulin-like growth factor-I levels in colon cancer patients. Eur J Cancer Prev 2007; 16 (4): 298-303. [ Links ]
204. Palozza P, Serini S, Boninsegna A, Bellovino D, Lucarini M, Monastra G, et al. The growth-inhibitory effects of tomatoes digested in vitro in colon adenocarcinoma cells occur through down regulation of cyclin D1, Bcl- 2 and Bcl-xL. Br J Nutr 2007; 98 (4): 789-95. [ Links ]
205. Palozza P, Bellovino D, Simone R, Boninsegna A, Cellini F, Monastra G, et al. Effect of b-carotene-rich tomato lycopene b-cyclase (tlcy-b) on cell growth inhibition in HT- 29 colon adenocarcinoma cells. Br J Nutr 2009; 102 (2): 207-14. [ Links ]
206. Kim DJ, Takasuka N, Kim JM, Sekine K, Ota T, Asamoto M, Murakoshi M, Nishino H, Nir Z, Tsuda H. Chemoprevention by lycopene of mouse lung neoplasia after combined initiation treatment with DEN, MNU and DMH. Cancer Lett 1997; 120 (1): 15-22 [ Links ]
207. Kim DJ, Takasuka N, Nishino H, Tsuda H. Chemoprevention of lung cancer by lycopene. Biofactors 2000; 13 (1-4): 95-102. [ Links ]
208.Männistö S, Smith-Warner SA, Spiegelman D, Albanes D, Anderson K, van den Brandt PA, et al. Dietary carotenoids and risk of lung cancer in a pooled analysis of seven cohort studies. Cancer Epidemiol Biomarkers Prev 2004; 13 (1): 40-8. [ Links ]
209. Gallicchio L, Boyd K, Matanoski G, Tao XG, Chen L, Lam TK, et al. Carotenoids and the risk of developing lung cancer: a systematic review. Am J Clin Nutr 2008; 88 (2): 372-83. [ Links ]
210. Huang JP, Zhang M, Holman CD, Xie X. Dietary carotenoids and risk of breast cancer in Chinese women. Asia J Clin Nutr 2007; 16 (Suppl 1): 437-42. [ Links ]
211. Voskuil DW, Vrieling A, Korse CM, Beijnen JH, Bonfrer JM, van Doorn J, et al. Effects of lycopene on the insulin- like growth factor (IGF) system in premenopausal breast cancer survivors and women at high familial breast cancer risk. Nutr Cancer 2008; 60 (3): 342-53. [ Links ]
212. King-Batoon A, Leszczynska JM, Klein CB, Modulation of gene methylation by genistein or lycopene in breast cancer cells. Environ Mol Mutagen 2008; 49 (1): 36-45. [ Links ]
213. Zhang M, Holman CD, Binns CW. Intake of specific carotenoids and the risk of epithelial ovarian cancer. Br J Nutr 2007; 98 (1): 187-93. [ Links ]
214. Okajima E, Ozono S, Endo T, Majima T, Tsutsumi M, Fukuda T, et al. Chemopreventive efficacy of piroxicam administered alone or in combination with lycopene and b-carotene on the development of rat urinary bladder carcinoma after N-butyl-N-(4-hydroxybutyl)nitrosamine treatment. Jpn J Cancer Res 1997; 88 (6): 543-52. [ Links ]
215. Okajima E, Tsutsumi M, Ozono S, Akai H, Denda A, Nishino H, et al. Inhibitory effect of tomato juice on rat urinary bladder carcinogenesis after N-butyl-N-(4- hydroxybutyl)nitrosamine initiation. Jpn J Cancer Res 1998; 89 (1): 22-6. [ Links ]
216. Kozuki Y, Miura Y, Yagasaki K. Inhibitory effects of carotenoids on the invasion of rat ascites hepatoma cells in culture. Cancer Lett 2000; 151 (1): 111-5. [ Links ]
217.Watanabe S, Kitade Y, Masaki T, Nishioka M, Satoh K, Nishino H. Effects of lycopene and Sho-saiko-to on hepatocarcinogenesis in a rat model of spontaneous liver cancer. Nutr Cancer 2001; 39 (1): 96-101. [ Links ]
218. Scolastici C, Lopes GA, Barbisan LF, Salvadori DM. Tomato oleoresin inhibits DNA damage but not diethylnitrosamine- induced rat hepatocarcinogenesis. Exp Toxicol Pathol 2008; 60 (1): 59-68. [ Links ]
219. Pelucchi C, Dal Maso L, Montella M, Parpinel M, Negri E, Talamini R, et al. Dietary intake of carotenoids and retinol and endometrial cancer risk in an Italian casecontrol study. Cancer Causes Control 2008; 19 (10): 1209-15. [ Links ]
220. Ziegler RG. A review of epidemiologic evidence that carotenoids reduce the risk of cancer. J Nutr 1989; 119 (1): 116-22. [ Links ]
221. Steinmetz KA, Potter JD. Vegetables, fruit, and cancer prevention: A review. J Am Diet Assoc 1996; 96 (10): 1027-39. [ Links ]
222. Flagg EW, Coates RJ, Greenberg RS. Epidemiologic studies of antioxidants and cancer in humans. J Am Coll Nutr 1995; 14 (5): 419-27. [ Links ]
223. Vena JE, Graham S, Freudenheim J, Marshall J, Zielezny M, Swanson M, et al. Diet in the epidemiology of bladder cancer in western New York. Nutr Cancer 1992; 18 (3): 255-64. [ Links ]
224. Rock CL, Saxe GA, Ruffin MT 4th, August DA, Schottenfeld D. Carotenoids, vitamin A, and estrogen receptor status in breast cancer. Nutr Cancer 1996; 25 (3): 281-96. [ Links ]
225.Wang C, Baumgartner RN, Yang D, Slattery ML, Murtagh MA, Byers T, et al. No evidence of association between breast cancer risk and dietary carotenoids, retinols, vitamin C and tocopherols in Southwestern Hispanic and non-Hispanic White women. Breast Cancer Res Treat 2009; 114 (1): 137-45. [ Links ]
226. Zheng W, Sellers TA, Doyle TJ, Kushi LH, Potter JD, Folsom AR. Retinol, antioxidant vitamins, and cancer of the upper digestive tract in a prospective cohort study of postmenopausal women. Am J Epidemiol 1995; 142 (9): 955-60. [ Links ]
227. Larsson SC, Bergkvist L, Naslund I, Rutegard J, Wolk A. Vitamin A, retinol, and carotenoids and the risk of gastric cancer: A prospective cohort study. Am J Clinical Nutr 2007; 85 (2): 497-503. [ Links ]
228. The a-Tocopherol, b-Carotene (ATBC) Cancer Prevention Study Group. The a-tocopherol, b-carotene lung cancer prevention study: design, methods, participant characteristics, and compliance. Ann Epidemiol 1994; 4 (1): 1-10. [ Links ]
229. The a-Tocopherol, b-Carotene (ATBC) Cancer Prevention Study Group. The effect of vitamin E and b-carotene on the incidence of lung cancer and other cancers in male smokers. N Engl J Med 1994; 330 (15): 1029-35. [ Links ]
230. Albanes D, Heinonen OP, Huttunen JK, Taylor PR, Virtamo J, Edwards BK, et al. Effects of a-tocopherol and b-carotene supplements on cancer incidence in the a- Tocopherol b-Carotene Cancer Prevention Study. Am J Clin Nutr 1995; 62 (6 suppl): 1427S-30S. [ Links ]
231. Albanes D, Heinonen OP, Taylor PR, Virtamo J, Edwards BK, Rautalahti M, et al. a-Tocopherol and b- carotene supplements and lung cancer incidence in the a-Tocopherol, b-Carotene Cancer Prevention Study: Effects of base-line characteristics and study compliance. J Natl Cancer Inst 1996; 88 (21): 1560-70. [ Links ]
232. Heinonen OP, Albanes D, Virtamo J, Taylor PR, Huttunen JK, Hartman AM, et al. Prostate cancer and supplementation with a-tocopherol and b-carotene: Incidence and mortality in a controlled trial. J Natl Cancer Inst 1998; 90 (6): 440-6. [ Links ]
233. Leppälä JM, Virtamo J, Fogelholm R, Huttunen JK, Albanes D, Taylor PR, et al. Controlled trial of a-tocopherol and b-carotene supplements on stroke incidence and mortality in male smokers. Arteroscler Thromb Vasc Biol 2000; 20 (1): 230-5. [ Links ]
234. Omenn GS, Goodman GE, Thornquist MD, Balmes J, Cullen MR, Glass A, et al. Risk factors for lung cancer and for intervention effects in CARET, the b-Carotene and Retinol Efficacy Trial. J Natl Cancer Inst 1996; 88 (21): 1550-9. [ Links ]
235. Omenn GS, Goodman GE, Thornquist MD, Balmes J, Cullen MR, Glass A, Keogh JP, Meyskens FL, Valanis B, Williams JH Jr, Barnhart S, Hammar S. Effects of a combination of b-carotene and vitamin A on lung cancer and cardiovascular disease. N Engl J Med 1996; 334 (18): 1150-5. [ Links ]
236. The ATBC Study Group. Incidence of Cancer and Mortality Following a-Tocopherol and b-Carotene Supplementation: A postintervention follow-up. JAMA 2003; 290 (4): 476-85. [ Links ]
237. Goodman GE, Thornquist MD, Balmes J, Cukken MR, Meyskens FL Jr, Omenn GS, et al. The b-Carotene and Retinol Efficacy Trial (CARET): Incidence of lung cancer and cardiovascular disease mortality during 6-year follow-up after stopping b-carotene and retinol supplements. J. Natl Cancer Inst 2004; 96 (23): 1743-50. [ Links ]
238. Omenn GS. Chemoprevention of lung cancers: lessons from CARET, The b-Carotene and Retinol Efficacy Trial, and prospects for the future. Eur J Cancer Prev 2007; 16 (3): 184-91. [ Links ]
239. Albanes D. b-Carotene and lung cancer: a case study. Am J Clin Nutr 1999; 69 (6): 1345S-50S. [ Links ]
240. Hennekens CH, Buring JE. Methodologic considerations in the design and conduct of randomized trials: The U.S. Physicians' Health Study. Controlled Clin Trials 1989; 10 (4 Suppl): 142S-50S. [ Links ]
241. Satterfield S, Greco PJ, Goldhaber SZ, Stampfer MJ, Swartz SL, Stein EA, et al. Biochemical markers of compliance in the Physicians' Health Study. Am J Prev Med 1990; 6 (5): 290-4. [ Links ]
242. Hennekens CH, Buring JE, Manson JE, Stampfer M, Roser B, Cook NR, et al. Lack of effect of long-term supplementation with b-carotene on the incidence of malignant neoplasms and cardiovascular disease. N Engl Med 1996; 334 (18): 1145-9. [ Links ]
243. Frieling UM, Schaumberg DA, Kupper TS, Muntwyler J, Hennekens CH. A randomized, 12-year primary-prevention trial of b-carotene supplementation for nonmelanoma skin cancer in the Physicians' Health Study. Arch Dermatol 2000; 136 (2): 179-84. [ Links ]
244. Lee I-M, Cook NR, Manson JE, Buring JE, Hennekens CH. b-Carotene supplementation and incidence of cancer and cardiovascular disease: The Women's Health Study. J Natl Cancer Inst 1999; 91 (24): 2102-6. [ Links ]
245. Vitale S, West S, Hallfrisch J, Alston C, Wang F, Moorman C, et al. Plasma antioxidants and risk of cortical and nuclear cataract. Epidemiology 1993; 4 (3): 195-203. [ Links ]
246. Christen WG, Gaziano JM, Hennekens CH. Design of Physicians' Health Study II: A randomized trial of b- carotene, vitamins E and C, and multivitamins, in prevention of cancer, cardiovascular disease, and eye disease, and review of results of completed trials. Ann Epidemiol 2000; 10 (2): 125-34. [ Links ]
247. Grodstein F, Kang JH, Glynn RJ, Cook NR, Gaziano JM. A randomized trial of b-carotene supplementation and cognitive function in men: The Physicians' Health Study II. Arch Intern Med 2007; 167 (20): 2184-90. [ Links ]
248. Keefe KA, Schell MJ, Brewer C, McHale M, Brewster W, Chapman JA, et al. A randomized, double blind, Phase III trial using oral b-carotene supplementation for women with high-grade cervical intraepithelial neoplasia. Cancer Epidemiol Biomarkers Prev 2001; 10 (10): 1029-35. [ Links ]
249. Tornwall ME, Virtamo J, Haukka JK, Albanes D, Huttunen JK. a-Tocopherol (vitamin E) and b-carotene supplementation does not affect the risk for large abdominal aortic aneurysm in a controlled trial. Atherosclerosis 2001; 157 (1): 167-73. [ Links ]
250. Schaumberg DA, Frieling UM, Rifai N, Cook N. No effect of b-carotene supplementation on risk of nonmelanoma skin cancer among men with low baseline plasma b-carotene. Cancer Epidemiol Biomarkers Prev 2004; 13 (6): 1079-80. [ Links ]
251. Druesne-Pecollo N, Latino-Martel P, Norat T, Barrandon E, Bertrais S, Galan P, Hercberg S. b-Carotene supplementation and cancer risk: a systematic review and meta-analysis of randomized controlled trials. Int J Cancer. 2009 Oct 28. [Epub ahead of print] PMID: 19876916 [ Links ]
252. Rodrigues MJ, Bouyon A, Alexandre J. Place en oncologie des compléments et suppléments en antioxydants en sus d'un régime équilibre; revue générale de la littérature. Bull Cancer 2009; 96 (6): 677-84. [ Links ]
253. Myung S-K, Kim Y, Ju W, Choi HJ, Bae WK. Effects of antioxidant supplements on cancer prevention: meta-analysis of randomized controlled trials. Ann Oncol 2010; 21 (1): 166-79. [ Links ]
254. Neuhouser ML, Patterson RE, Thornquist MD, Omenn GS, King IB, Goodman GE. Fruits and vegetables are associated with lower lung cancer risk only in the placebo arm of the b-carotene and retinol efficacy trial (CARET). Cancer Epidemiol Biomarkers Prev 2003; 12 (4): 350-8. [ Links ]
255. Lam TK, Gallicchio L, Lindsley K, Shiels M, Hammond E, Tao X, et al. Cruciferous vegetable consumption and lung cancer risk: A systematic review. Cancer Epidemiol. Biomarkers Prev 2009; 18 (1): 184 -95. [ Links ]
256. Goralczyk R. b-Carotene and lung cancer in smokers: review of hypotheses and status of research. Nutr Cancer 2009; 61 (6): 767-74. [ Links ]
257. White WS, Stacewicz-Sapuntzakis M, Erdman JW Jr, Bowen PE. Pharmacokinetics of b-carotene and canthaxanthin after ingestion of individual and combined doses by human subjects. J Am Coll Nutr 1994; 13 (6): 665-71. [ Links ]
258. Wang XD, Russell RM. Procarcinogenic and anticarcinogenic effects of b-carotene. Nutr Rev 1999; 57 (9 Pt 1): 263-72. [ Links ]
259. Brambilla D, Mancuso C, Scuderi MR, Bosco P, Cantarella G, Lempereur L, et al. The role of antioxidant supplement in immune system, neoplastic, and neurodegenerative disorders: a point of view for an assessment of the risk/benefit profile. Nutr J 2008; 7: 29-37. [ Links ]
260. Palozza P, Simone R, Mele MC. Interplay of carotenoids with cigarette smoking: implications in lung cancer. Curr Med Chem 2008; 15 (9): 844-54. [ Links ]
261. Wang X-D, Liu C, Bronson RT, Smith DE, Krinsky NI, Russell RM. Retinoid signaling and activator protein-1 expression in ferrets given b-carotene supplements and exposed to tobacco smoke. J Natl Cancer Inst 1999; 91 (1): 60-6. [ Links ]
262. Liu C, Wang X-D, Bronson RT, Smith DE, Krinsky NI, Russell RM. Effects of physiological versus pharmacological b-carotene supplementation on cell proliferation and histopathological changes in the lungs of cigarette smoke-exposed ferrets. Carcinogenesis 2000; 21 (12): 2245-53. [ Links ]
263. Witschi H, Espiritu I, Suffia M, Pinkerton KE. Expression of cyclin D1/2 in the lungs of strain A/J mice fed chemopreventive agents. Carcinogenesis 2002; 23 (2): 289-94. [ Links ]
264. Yang K, Lamprecht SA, Liu Y, Shinozaki H, Fan K, Leung D, et al. Chemoprevention studies of the flavonoids quercetin and rutin in normal and azoxymethane-treated mouse colon. Carcinogenesis 2000; 21 (9): 1655-60. [ Links ]
265. Choi JA, Kim JY, Lee JY, Kang CM, Kwon HJ, Yoo YD, et al. Induction of cell cycle arrest and apopotosis in human breast cancer cells by quercetin. Int J Oncol 2001; 19 (4): 837-44 [ Links ]
266. Morrow DM, Fitzsimmons PEE, Chopra M, McGlynn H. Dietary supplementation with the anti-tumour promoter quercetin: Its effects on matrix matalloproteinase gene regulation. Mutat Res 2001; 480-481: 269-76. [ Links ]
267. Salgo MG, Cueto R, Winston GW, Pryor WA. b-Carotene and its oxidation products have different effects on microsome mediated binding of benzo[a]pyrene to DNA. Free Radic Biol Med 1999; 26 (1-2): 162-73. [ Links ]
268. Paolini M, Antelli A, Pozzetti L, Spetlova D, Perocco P, Valgimigli L, et al. Induction of cytochrome P450 enzymes and over-generation of oxygen radicals in b- carotene supplemented rats. Carcinogenesis 2001; 22 (9): 1483-95. [ Links ]
269. Obermeier MT, White RE, Yang CS. Effects of bioflavonoids on hepatic P450 activities. Xenobiotica 1995; 25 (6): 575-84. [ Links ]
270. Rahden-Staron I, Czeczot H, Szumilo M. Induction of rat liver cytochrome P450 isoenzymes CYP 1A and CYP 2B by different fungicides, nitrofurans, and quercetin. Mutat Res 2001; 498 (1-2): 57-66 [ Links ]
271. Valerio LG Jr, Kepa JK, Pickwell GV, Quattrochi LC. Induction of human NAD(P)H: quinone oxidoreductase (NQO1) gene expression by the flavonol quercetin. Toxicol Lett 2001; 119 (1): 49-57. [ Links ]
272. Poulain S, Evenou F, Carré MC, Corbel S, Vignaud JM, Martinet N. Vitamin A/retinoids signalling in the human lung. Lung Cancer 2009; 66 (1): 1-7. [ Links ]
273. Bogos K, Renyi-Vamos F, Kovacs G, Tovari J, Dome B. Role of retinoic receptors in lung carcinogenesis. J Exp Clin Cancer Res 2008; 27 (1): 18-25. [ Links ]
274. Leo MA, Lieber CS. Alcohol, vitamin A, and b-carotene: adverse interactions, including hepatotoxicity and carcinogenicity. Am J Clin Nutr 1999; 69 (6): 1071-85. [ Links ]
275. Ni R, Leo MA, Zhao J, Lieber CS. Toxicity of b-carotene and its exacerbation by acetaldehyde in HepG2 cells. Alcohol Alcohol 2001; 36 (4): 281-5. [ Links ]
276. Albanes D, Virtamo J, Taylor PR, Rautalahti M, Pierinen P, Heinonen OP. Effects of supplemental b-carotene, cigarette smoking, and alcohol consumption on serum carotenoids in the a-Tocopherol, b-Carotene Cancer Prevention Study. Am J Clin Nutr 1997; 66 (2): 366-72. [ Links ]
277. Bandera EV, Freudenheim JL, Marshall JR, Zielezny M, Priore RL, Brasure J, et al. Diet and alcohol consumption and lung cancer risk in the New York State Cohort (United States). Cancer Causes Control 1997; 8 (6): 828-40. [ Links ]
278. Feskanich D, Ziegler RG, Michaud DS, Giovannucci EL, Speizer FE, Willett WC, et al. Prospective study of fruit and vegetable consumption and risk of lung cancer among men and women. J Natl Cancer Inst 2000; 92 (22): 1812-23. [ Links ]
279. Michaud DS, Feskanich D, Rimm EB, Colditz GA, Speizer FE, Willett WC, et al. Intake of specific carotenoids and risk of lung cancer in 2 prospective US cohorts. Am J Clin Nutr 2000; 72 (4): 990-7. [ Links ]
Aceptado para su publicación el 14 de marzo de 2010

