










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkActa bioquímica clínica latinoamericana
versión impresa ISSN 0325-2957
Acta bioquím. clín. latinoam. vol.47 no.2 La Plata abr./jun. 2013
BIOQUÍMICA CLÍNICA
Mecanismo de acción de la toxina alfa hemolisina de Escherichia coli
Action mechanism of Escherichia coli alpha-hemolysin toxin
Mecanismo de ação da toxina alfa-hemolisina de Escherichia coli
Laura Bakás1a, Sabina Maté2a, Romina Vazquez3b, Vanesa Herlax2c
1 Dra. en Ciencias Bioquímicas.
2 Dra. de la Facultad de Ciencias Exactas.
3 Bioquímica.
a Miembros de la carrera científica de la Comisión de Investigaciones Científicas de la Provincia de Buenos Aires.
b Becaria de posgrado del Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET).
c Miembro de la carrera científica de CONICET.
Instituto de Investigaciones Bioquímicas La Plata (INIBIOLP), CCT- La Plata, CONICET. Facultad de Ciencias Médicas. Universidad Nacional de La Plata. La Plata, Buenos Aires, Argentina.
CORRESPONDENCIA DRA. LAURA BAKÁS Instituto de Investigaciones Bioquímicas La Plata (INIBIOLP) Facultad de Ciencias Médicas 60 y 120, 1° piso 1900 LA PLATA, BUENOS AIRES, Argentina. E-mail: lbakas@biol.unlp.edu.ar
Resumen
Escherichia coli es una de las bacterias anaerobias facultativas más predominantes en el intestino, siendo, en la mayoría de los casos, inocua para el huésped. Existen cepas que traslocan al torrente sanguíneo causando enfermedades extraintestinales como infecciones urinarias, septicemia y meningitis. Dentro de éstas se encuentran las cepas uropatogénicas (Uropathogenic Escherichia coli: UPEC), que secretan varios factores de virulencia. Estos últimos incluyen: toxinas, sistemas de adquisición de hierro, adhesinas y antígenos capsulares. Las principales toxinas secretadas son: alfa-hemolisina (HlyA) y el factor necrotizante citotóxico 1 (CNF-1). En esta revisión se presenta una descripción exhaustiva de HlyA, incluyendo su síntesis, maduración y exportación desde la bacteria. La acilación de la proteína en dos residuos internos de lisina la convierte en una toxina muy virulenta al exponer regiones intrínsecamente desordenadas que son esenciales en diferentes pasos del mecanismo de acción de la misma. Específicamente, la exposición de estas regiones está involucrada en interacciones proteína-proteína dentro del proceso de oligomerización. La formación del oligómero es responsable de la permeabilidad inducida en las células blanco. Finalmente, basado en los conocimientos acerca de las características estructurales y funcionales de HlyA, se presentan potenciales usos de HlyA en terapias basadas en toxinas
Palabras clave: Toxinas; Oligomerización; Microdominios de membrana; Inmunotoxinas; Regiones intrínsecamente desordenadas.
Summary
Escherichia coli is one of the predominant species of facultative anaerobes in the human gut, and in the majority of the cases it is harmless to the host. Some strains of this species can translocate to blood and cause infection such as urinary infection, septicemia and meningitis. These are the uropathogenic E. coli strains (UPEC) that secrete a number of virulence factors. The latter include a number of secreted toxins, iron-acquisition systems, adhesins, and capsular antigens. Secreted toxins include HlyA, the cytotoxic necrotizing factor-1 (CNF-1). In this review an exhaustive description of the toxin has been delineated, including its synthesis, maturation, and export from the bacteria. The acylation of the protein at two internal lysine residues gives the toxin its virulence, by exposing intrinsic disordered regions that are essential in different steps of the toxin's mechanism of action. The further exposure of regions involved in the protein-protein interaction within the oligomerization process is responsG-ible for the permeability induced in all the target cells. Based on the already known structural and functional characteristics of HlyA, the potential use in toxin-based therapy is presented.
Key words: Toxins; Oligomerization; Membrane microdomains; Immunotoxins; Intrinsic disordered regions.
Resumo
Escherichia coli é uma das bactérias anaérobias facultativas mais predominantes no intestino, sendo na maioria dos casos inócua para o hóspede. Há cepas que passam ao torrente sanguíneo causando doenças extraintestinais como infecção urinária, septicemia e meningite. Dentro destas se encontram as cepas uropatogênicas (Uropathogenic Escherichia coli: UPEC) que secretam varios fatores de virulência. Estos últimos incluem: toxinas, sistemas de aquisição de ferro, adesinas e antígenos capsulares. As principais toxinas secretadas são: alfa hemolisina (HlyA) e o fator necrotizante citotóxico 1 (CNF-1). Nesta revisão apresenta-se uma descrição exaustiva de HlyA incluindo sua sintese, seu amadurecimento e exportação a partir da bactéria. A acilação da proteína em dois residuos internos de lisina a transforma numa toxina muito virulenta ao expor regiões intrinsecamente desordenadas que são essenciais em diferentes passos do mecanismo de ação da mesma. Especificamente, a exposição destas regiões esta envolvida em interações proteína-proteína dentro do processo de oligomerização. A formação do oligômero é responsável pela permeabilidade induzida nas células alvo. Finalmente, com base nos conhecimentos acerca das características estruturais e funcionais de HlyA, apresentam-se potenciais usos de HlyA em terapias baseadas em toxinas.
Palavras chave: Toxinas; Oligomerização; Microdomínio de membrana; Imunotoxinas; Regiões intrinsecamente desordenadas.
1. Introducción
Escherichia coli es el agente responsable de, al menos, el 80% de las infecciones del tracto urinario (ITU), patología que actualmente se encuentra entre las infecciones más comunes en el mundo (1)(2). Las cepas de E. coli que causan ITU son llamadas cepas uropatogénicas (Uropathogenic Escherichia coli: UPEC). En los últimos años se ha obtenido una enorme cantidad de información proveniente del secuenciamiento de genomas de varias cepas clínicas. Estos datos, junto con informes epidemiológicos, han confirmado que diferentes patotipos de E. coli comparten muchos conocidos, así como también presuntos, factores de virulencia. Estos últimos incluyen un número de toxinas secretadas, sistemas de adquisición de hierro, adhesinas y antígenos capsulares (3). Las proteínas secretadas, entre las que se incluye la toxina proteica alfa-hemolisina (HlyA) y el factor necrotizante citotóxico 1 (CNF-1), pueden alterar las cascadas de señalización de las células huésped, alterando la respuesta inflamatoria e induciendo la muerte celular, mientras que sideroforos como aerobactina, bacteriocina y enterobactina permiten que ExPEC secuestren hierro (3)(4).
Experimentos realizados en cultivo celular han mostrado que altos niveles de HlyA pueden causar la lisis osmótica de la célula huésped, mientras que concentraciones sublíticas de la toxina pueden modular las rutas de sobrevida de la célula huésped (5-9). Tanto HlyA como CNF-1 pueden además estimular la ruptura de la barrera epitelial (10) y son los responsables de la traslocación de la bacteria desde el tracto digestivo al torrente sanguíneo para finalmente diseminarse y colonizar diferentes tejidos, como el nervioso y el tracto urinario, causando así la patología.
2. Alfa-hemolisina (HlyA) de E. coli
HlyA representa el prototipo de la primera familia de proteínas RTX (Repeat in Toxin) caracterizada por Welch (11). Producida por una gran variedad de bacterias gram-negativas, estas proteínas exhiben dos características comunes: la primera es la presencia de un nonapéptido repetido rico en residuos de glicina y ácido aspártico, localizados cerca de la porción C-terminal; la segunda es la secreción desde la bacteria por el sistema tipo I (un transportador ABC). Este grupo de proteínas RTX comprende toxinas, la mayoría de ellas con actividad formadora de poros, que se detectan por la formación de un halo hemolítico rodeando a las colonias, cuando éstas son crecidas en placas agar sangre (11)(12). Recientemente fue descubierto un subgrupo de toxinas RTX muy grandes (>3200 residuos) con múltiples actividades, como por ejemplo: actividad lipasa y proteasa, siendo la toxina de Vibrio cholerae prototipo de esta familia. En resumen, las proteínas RTX forman una amplia familia de proteínas con un amplio espectro de actividades biológicas y bioquímicas (13).
3. Síntesis y estructura de HlyA
La síntesis, maduración y secreción de HlyA está determinada por el operón hlyCABD (14-17). El gen estructural hlyA codifica para un polipéptido de 110-kDa. Este polipéptido, denominado proHlyA, es la forma inactiva de la toxina, que se convierte en lítica luego de una activación postraduccional. El pI estimado de la toxina es 4,5 que es una característica común entre las toxinas RTX. Según las predicciones estructurales realizadas, el dominio hidrofóbico N-terminal contiene 9 α-hélices anfipáticas (18), siendo ésta otra característica común de las toxinas RTX. Mediante experimentos realizados en liposomas conteniendo un reactivo fotoactivable, Hyland et al. demostraron que la región comprendida entre los residuos 177-411 es la que se inserta en la membrana de las células blanco (19). El dominio de unión a calcio consiste de 11-17 repeticiones de un nonapéptido rico en glicinas y ácido aspártico, con estructura de hoja plegada β. Aunque se asume que la interacción de HlyA con membranas ocurre principalmente a través del dominio de las α-hélices anfipáticas, recientemente se ha propuesto que ambos dominios están involucrados en la interacción con la membrana, siendo el dominio de unión a calcio el responsable de las etapas tempranas de la interacción de la toxina con la membrana de la célula blanco (20).
Uno de los aspectos más controvertidos en relación al mecanismo de acción de la toxina es la existencia de un receptor específico de alta afinidad. Cortajarena et al. observaron que una corta secuencia del dominio C-terminal (entre los residuos 914-936) se une a la glicoforina A de los eritrocitos (21); mientras que otro grupo postula que la unión a eritrocitos es inespecífica (22).
Los últimos 60 residuos aminoacídicos de la porción C-terminal consisten en dos α-hélices separadas por 8-10 residuos cargados. Este dominio está implicado en el transporte de la toxina al medio extracelular, a través del sistema de secreción tipo I compuesto por HlyA B y HlyA D (23). Como se mencionó anteriormente, estas dos proteínas también están codificadas en el mismo operón que HlyA. La Figura 1 muestra un esquema de la estructura de HlyA.
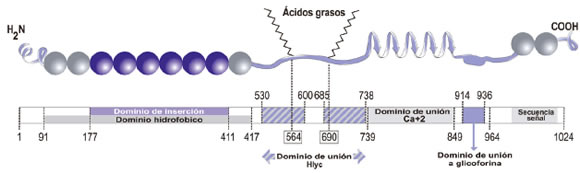
Fig. 1. Esquema de la estructura de HlyA donde se indican los dominios más relevantes para el mecanismo de acción. (Figura adaptada del capítulo "E. coli alpha hemolysin and properties" del libro Biochemistry (24))
4. Activación postraduccional de HlyA
La protoxina proHlyA, sin actividad lítica, se convierte en el citosol de la bacteria en una toxina activa mediante una acilación dirigida por HlyC, antes de su exportación al medio extracelular. Este proceso de modificación postraduccional consiste en la acilación de dos residuos internos de Lys mediante enlaces ε-amino, catalizados por la aciltransferasa Hlyc, que se expresa junto con la protoxina (25).
El mecanismo de este nuevo tipo de acilación de proteínas ha sido extensamente analizado para HlyA (15),(26). HlyC emplea residuos de ácido graso transportados por la proteína transportadora de acilos (PTA) para formar un intermediario covalente acil-HlyC, el cual transfiere los residuos de ácido graso a un grupo ε de las lisinas Lys 564 y Lys 690 de la proHlyA (27)(28). PTA transporta diversos residuos de ácido graso, incluyendo palmitato (16:0) y palmitoleato (16:1), el más común en E. coli y que puede ser empleado para la acilación in vitro de una manera eficiente como donor de acilo para modificar HlyA (15)(29). In vivo, sin embargo, HlyC presenta una alta selectividad por ácido mirístico (14:0), de manera que este ácido graso constituye aproximadamente el 68% de las cadenas de ácido graso unidas a Lys 564 y Lys 690 de HlyA nativa (30). Es importante destacar que sólo la acilación de Lys 690 es esencial para la actividad de HlyA.
La importancia de esta modificación postraduccional radica en que cambia el comportamiento de la proteína de una forma benigna a una forma netamente tóxica. Además, este tipo de modificaciones constituye un mecanismo único en procariotas, existiendo unos pocos casos en eucariotas, como por ejemplo en el receptor nicotínico de acetilcolina, receptor de insulina y en citoquinas como TNF-α, IL-1α y IL-1β (31).
5. Secreción al medio extracelular
La maduración de la proteína no es una modificación necesaria para su exportación (32). Todas las toxinas RTX son secretadas a través de ambas membranas por un sistema de exportación tipo I que emplea como sistema de reconocimiento la porción C-terminal (33)(34), y no se realiza a través de un péptido señal (14) ni de intermediarios periplásmicos (35)(36). El aparato secretor de HlyA está compuesto de HlyB (una ATPasa de membrana interna, conocida como ATP-binding cassette), HlyD (una proteína de fusión de membranas) y TolC (una proteína de membrana interna) (37-39). En E. coli y en la mayoría de los patógenos, TolC está codificada por un gen separado del operón hlyCABD. La formación de un agregado entre el complejo HlyB-HlyD y TolC en la membrana facilita la formación de un canal de exportación a través del cual pasa HlyA (40). La energía necesaria para la secreción no sólo es aportada por la hidrólisis de ATP mediada por HlyB sino también por la fuerza protón motriz ejercida sobre la membrana interna (41)(42). Si bien se asume que el sistema de secreción Tipo I involucra la traslocación de proteínas desplegadas (43), se ha reconocido recientemente que el contacto con HlyD afecta el plegado de HlyA durante el tránsito por el canal (44). En los últimos años, muchos investigadores han estado interesados en este tipo de secreción debido a su potencial aplicación en la exportación de proteínas quiméricas y en la producción de vacunas (45)(46).
A pesar de la existencia de su propia maquinaria de exportación en la bacteria, HlyA también se ha detectado en vesículas de membrana externa (VME) liberadas de cepas clínicas (47), así como de cepas de E. coli recombinantes (48)(49).
Las VME son constantemente liberadas de la superficie de bacterias gram-negativas durante el crecimiento bacteriano. Todas las bacterias gram-negativas estudiadas hasta hoy, incluyendo E. coli, producen VME, estando su liberación incrementada cuando la bacteria se encuentra expuesta a condiciones de estrés y tratamiento con antibióticos. Aún cuando la liberación de VME no pudo ser demostrada in vivo, su presencia fue detectada en el plasma de pacientes con diferentes enfermedades infecciosas (50). Las VME sirven como vehículo de proteínas y lípidos de bacterias gram-negativas jugando un rol en la colonización y transmisión de factores de virulencia en el huésped o modulando las defensas del huésped al actuar como un reservorio de factores de virulencia protegiéndolos de las proteasas extracelulares del huésped y penetrando en los tejidos más fácilmente que las bacterias de mayor tamaño (51). En el caso particular de HlyA, se ha demostrado que la toxina secretada por este mecanismo es transferida a la célula huésped de manera más concentrada, siendo por lo tanto, hemolíticamente más eficiente que la toxina libre (48)(49).
6. El mecanismo de acción de HlyA
HlyA actúa sobre una gran variedad de tipos celulares de diversas especies -ej.: glóbulos rojos, fibroblastos embrionarios y adultos, granulocitos, linfocitos y macrófagos (52)- actuando también sobre sistemas modelo de membranas (liposomas) libres de proteína (53).
El microambiente en el huésped encontrado por ExPEC es extremadamente pobre en nutrientes, de allí que una de las funciones que podría tener HlyA sería la destrucción de las células blanco, facilitando de esta manera su aporte de nutrientes y otros factores críticos para su crecimiento como hierro. El mecanismo de acción lítica es un mecanismo complejo que finaliza en la lisis celular. En este proceso, se han reconocido al menos 3 etapas: unión a la membrana de la célula blanco, inserción y oligomerización.
Los estudios acerca de la unión de HlyA a membranas y la caracterización de un posible receptor específico para la toxina han arrojado resultados contradictorios. Lally et al. mostraron resultados sobre el rol de la integrina (CD11a/CD18; α1β2 integrina), actuando como receptor de HlyA en neutrófilos mononucleares (54) encontrando posteriormente que HlyA reconoce y se une a oligosacáridos de la subunidad β2 del receptor integrina (55). Estos resultados sugieren que la unión inicial de la toxina a diferentes células podría ocurrir a través del reconocimiento de componentes glicosilados de la membrana, como glicoproteínas y gangliósidos. Recientemente, Cortajarena et al. encontraron que HlyA se une a glicoforina presente en los glóbulos rojos de caballo y que esta unión se pierde al tripsinizar las proteínas de la membrana del glóbulo rojo (56). Además estos autores encontraron que la glicoforina purificada de fantasmas de estos glóbulos rojos al ser reconstituída en liposomas, incrementa significativamente la sensibilidad de los liposomas hacia la toxina. Posteriormente los mismos autores informaron que los residuos aminoacídicos 914-936 de HlyA serían responsables de la unión a este receptor (21).
Sin embargo, otros estudios indican que la unión de la toxina a las células ocurre de una manera no saturable, por lo que la toxina no se uniría a un receptor específico, ni en glóbulos rojos ni en linfocitos (22). Además, existe un gran número de resultados que muestran que HlyA produce la lisis de liposomas libres de proteínas. Resultados de Ostolaza et al. muestran que HlyA produce la liberación de solutos fluorescentes encapsulados en liposomas por un mecanismo todo o nada. Empleando vesículas unilamelares grandes de diferente composición lipídica, estos investigadores encontraron que vesículas compuestas de fosfatidilcolina, fosfatidiletanolamina y colesterol en relación molar 2:1:1 presentaron la mayor sensibilidad frente a la toxina (53). Estos resultados demuestran que la presencia de un receptor no es necesaria para que la lisis ocurra. En definitiva, existe una gran controversia respecto de la presencia o ausencia de un receptor proteico de alta afinidad, que podría deberse a que los estudios antes mencionados se han realizado en diferentes tipos celulares.
La interacción de HlyA con membranas desprovistas de receptor ocurre en dos etapas: una adsorción inicial y reversible, sensible a fuerzas electrostáticas seguida de una inserción irreversible en la membrana (57)(58). Estudios realizados con el dominio de unión a calcio aislado, proponen que la toxina se uniría a la membrana a través de este dominio, en las etapas tempranas de la interacción (20).
El paso siguiente en la etapa hemolítica sería la inserción de la toxina en la membrana. Hyland et al. demostraron que la región de HlyA que se inserta en la membrana está localizada entre los residuos 177 y 411 (19). Además, la inserción sería una etapa independiente de la lisis dado que mutantes de HlyA desprovistos de actividad lítica pueden insertarse en monocapas lipídicas (59)(60). En el medio extracelular, HlyA debe unir calcio para adoptar una conformación tal que haga que la unión sea irreversible y transforme a la toxina en una forma hemolíticamente activa (61)(60). Este segundo paso de activación es además dependiente de la acilación, ya que la capacidad de unir calcio disminuye en la proteína no acilada (62). Una vez activada por la unión de calcio, se adsorbe reversiblemente para finalmente insertarse en la membrana (57). La forma de HlyA insertada en la membrana se comporta como una proteína integral que no puede ser extraída sin el uso de detergentes (18).
Sin embargo, proHlyA, la forma no acilada de la proteína, también interactúa con membranas. Esta observación no es sorprendente ya que el análisis de la secuencia del polipéptido muestra hélices anfipáticas en la región comprendida entre los residuos aminoacídicos 250-400. Si bien la presencia de este dominio anfipático es esencial para la actividad, su sola presencia no imparte actividad lítica, ya que proHlyA es incapaz de alterar la permeabilidad de la membrana (18). Experimentos de adsorción de la proteína en la interfase aire-agua sugieren que los ácidos grasos presentes en HlyA y ausentes en proHlyA, no modifican las propiedades de adsorción en superficies, y que la principal diferencia entre el precursor y la proteína madura es que proHlyA es virtualmente incapaz de insertarse en la monocapa lipídica (59). Además, se encontró que la presencia de las dos cadenas acilo unidas a HlyA le confieren a la toxina la capacidad de unirse irreversiblemente en la membrana, evento esencial para que el proceso lítico tenga lugar (49)(63). En resumen, aunque los ácidos grasos covalentemente unidos en HlyA incrementen la afinidad de la proteína por calcio induciendo una conformación competitiva para su interacción con membranas, la ausencia de ácidos grasos no modifica su capacidad de unión con la membrana, por lo que el rol de los ácidos grasos debe ser relevante en algún paso posterior del mecanismo lítico. Se ha demostrado que el rol de los ácidos grasos es inducir un cambio conformacional que permita la exposición de regiones intrínsecamente desordenadas dentro de la secuencia de la proteína como consecuencia de la inducción de un estado del tipo glóbulo fundido (molten globule).
La mayoría de las regiones desordenadas de HlyA, predichas empleando el predictor de regiones naturalmente desordenadas (PONDR), están localizadas en la mitad C-terminal de la proteína (Fig. 2). Estos dominios podrían estar relacionados con diferentes etapas del mecanismo de acción desde la exportación de la toxina de la bacteria hasta la formación de poros en la célula blanco. Estas regiones son responsables de promover interacciones proteína-proteína involucradas en el proceso de oligomerización, tal como se ha encontrado en el caso de HlyA (49)(64). En contraste con esto, no se ha podido aislar un oligómero en el caso de proHlyA (49)(65).

Fig. 2. Activación de proHlyA por acilación y exposición de regiones intrínsecamente desordenadas (Figura adaptada del capítulo "E. coli alpha hemolysin and properties" del libro Biochemistry (24))
El poro formado, el cual se ha caracterizado por medidas de conductancia y tiempo de vida de la membrana empleando membranas lipídicas, es un poro proteolipídico y altamente dinámico, ya que estos parámetros son fuertemente dependientes de la composición lipídica de la membrana (66). Además, se ha encontrado que el proceso de oligomerización está facilitado por microdominios de membrana (64)(65).
Una amplia variedad de patógenos interactúan con microdominios de la membrana plasmática. Estos microdominios están enriquecidos en esfingolípidos y colesterol y probablemente existan en un estado de líquido ordenado (67). Muchas proteínas son dirigidas específicamente a estos microdominios por su asociación favorable con las cadenas ordenadas de los lípidos que los forman. Es esperable que proteínas como HlyA, que poseen grupos acilo saturados, particionen en dichos dominios. Si bien se ha encontrado que los ácidos grasos de HlyA no están implicados en la unión a membranas, sí favorecen la oligomerización de la toxina. Dicha oligomerización ocurre en los microdominios de membrana, que actuarían como plataformas para la concentración de la toxina (65). El número de moléculas de HlyA que deben asociarse para formar el poro no se ha determinado, sin embargo, a partir del análisis de la cinética del proceso de hemólisis en glóbulos rojos control y deplecionados de colesterol, es posible asumir que a altas dosis de toxina, existe una oligomerización progresiva que lleva a la fusión de pequeños poros con una rápida destrucción de la célula. En contraste, a bajas concentraciones de toxina, los poros serían de menor tamaño, de manera tal que las células, si bien están dañadas, podrían sobrevivir el tiempo suficiente como para que se activen las rutas apoptóticas. En conclusión, la fusión de los oligómeros sería el paso limitante en la formación del poro, teniendo un papel muy importante en esta etapa la integridad de los microdominios de membrana enriquecidos en colesterol.
Finalmente, las concentraciones sublíticas de HlyA, alcanzadas lejos del foco de infección, pueden ser aún más relevantes fisiológicamente. Estudios recientes han demostrado que concentraciones sublíticas de un número de toxinas formadoras de poros pueden modular una amplia variedad de rutas de transducción de señales en el huésped, tales como el incremento oscilatorio de las concentraciones de calcio, la activación de la ruta de señalización de las MAPquinasas y la alteración de los patrones de fosforilación y acetilación de histonas (68)(69). Recientemente se ha encontrado que HlyA puede estimular la inactivación de la proteínquinasa B serina/treonina (PKB), enzima que juega un rol central en la progresión del ciclo celular, metabolismo, tráfico vesicular y las vías involucradas en el proceso inflamatorio (5). Estos hechos podrían ayudar a entender las implicancias de las concentraciones sublíticas de HlyA en la inhibición de la quimiotaxis y en la estimulación de las rutas inflamatorias y apoptóticas en el huésped (7)(70-73).
7. Conclusión
Las toxinas bacterianas han sido definidas como: "sustancias solubles que pueden alterar el metabolismo normal de las células del huésped, produciendo efectos perjudiciales en el mismo" (74). Sin embargo, en la última década, los investigadores han buscado la manera de obtener beneficios a partir de la toxina, de allí el aumento de las terapias basadas en toxinas. Dentro de estas terapias se puede mencionar la producción de inmunotoxinas y de vacunas.
Las inmunotoxinas son proteínas quiméricas compuestas por una toxina fusionada a una proteína que reconoce específicamente un ligando sobreexpresado en células cancerosas. Considerando que a concentraciones sublíticas HlyA produce la inactivación de PKB, proteína clave en la sobrevida de la célula huésped (5); y que además no es necesario el ingreso de HlyA al interior celular para que produzca este efecto, HlyA podría ser un buen candidato para la producción de inmunotoxinas (24).
Muchos son los conocimientos acerca de la estructura y función de HlyA que harían posible la proyección sobre su aplicación en estas terapias. Sin embargo, sería necesario el esclarecimiento de la existencia de un receptor específico, así como también la caracterización del dominio de la proteína involucrado en el reconocimiento de la membrana.
1. Marrs CF, Zhang L, Foxman B. Escherichia coli mediated urinary tract infections: are there distinct uropathogenic E. coli (UPEC) pathotypes? FEMS Microbiol Lett 2005; 252(2): 183-90. [ Links ]
2. Foxman B, Brown P. Epidemiology of urinary tract infections: transmission and risk factors, incidence, and costs. Infect Dis Clin North Am 2003; 17(2): 227-41.
3. Wiles TJ, Kulesus RR, Mulvey MA. Origins and virulence mechanisms of uropathogenic Escherichia coli. Exp Mol Pathol 2008; 85(1): 11-9.
4. Guyer DM, Radulovic S, Jones FE, Mobley HL. Sat, the secreted autotransporter toxin of uropathogenic Escherichia coli is a vacuolating cytotoxin for bladder and kidney epithelial cells. Infect Immun 2002; 70(8): 4539-46.
5. Wiles TJ, Dhakal BK, Eto DS, Mulvey MA. Inactivation of host Akt/protein kinase B signaling by bacterial pore-forming toxins. Mol Biol Cell 2008; 19(4): 1427-38.
6. Jonas D, Schultheis B, Klas C, Krammer PH, Bhakdi S. Cytocidal effects of Escherichia coli hemolysin on human T lymphocytes. Infect Immun 1993; 61(5): 1715-21.
7. Cavalieri SJ, Snyder IS. Effect of Escherichia coli alpha-hemolysin on human peripheral leukocyte viability in vitro. Infect Immun 1982; 36(2): 455-61.
8. Chen M, Tofighi R, Bao W, Aspevall O, Jahnukainen T, Gustafsson LE, et al. Carbon monoxide prevents apoptosis induced by uropathogenic Escherichia coli toxins. Pediatr Nephrol 2006; 21(3): 382-9.
9. Dhakal BK, Mulvey MA. The UPEC pore forming toxin α-Hemolysin triggers proteolysis of host proteins to disrupt cell adhesion, inflammatory and survival pathways. Cell Host Microbe 2012; 11(1): 58-69.
10. Smith YC, Rasmussen SB, Grande KK, Conran RM, O'Brien AD. Hemolysin of uropathogenic Escherichia coli evokes extensive shedding of the uroepithelium and hemorrhage in bladder tissue within the first 24 hours after intraurethral inoculation of mice. Infect Immun 2008; 76(7): 2978-90.
11. Welch RA. Pore-forming cytolysins of Gram-negative bacteria. Mol Microbiol 1991; 5(3): 521-8.
12. Müller D, Hughes C, Goebel W. Relationship between plasmid and chromosomal hemolysin determinants of Escherichia coli. J Bacteriol 1983; 153(2): 846-51.
13. Linhartova I, Bumba L, Masin J, Basler M, Osicka R, Kamanova J, et al. RTX proteins: a highly diverse family secreted by a common mechanism. FEMS Microbiol Rev 2010; 34(6): 1076-112.
14. Felmlee T, Pellett S, Lee EY, Welch RA. Escherichia coli hemolysin is released extracellularly without cleavage of a signal peptide. J Bacteriol 1985; 163(1): 88-93.
15. Issartel JP, Koronakis V, Hughes C. Activation of Escherichia coli prohaemolysin to the mature toxin by acyl carrier protein-dependent fatty acylation. Nature 1991; 351 (6329): 759-61.
16. Koronakis V, Li J, Koronakis E, Stauffer K. Structure of TolC, the outer membrane component of the bacterial type I efflux system, derived from two-dimensional crystals. Mol Microbiol 1997; 23(3): 617-26.
17. Nieto JM, Bailey MJ, Hughes C, Koronakis V. Suppression of transcription polarity in the Escherichia coli hemolysin operon by a short upstream element shared by polysaccharide and DNA transfer determinants. Mol Microbiol 1996; 19(4): 705-13.
18. Soloaga A, Veiga MP, García Segura LM, Ostolaza H, Brasseur R, Goñi F M. Insertion of Escherichia coli alpha-haemolysin in lipid bilayers as a non-transmembrane integral protein: prediction and experiment. Mol Microbiol 1999; 31(4): 1013-24.
19. Hyland C, Vuillard L, Hughes C, Koronakis V. Membrane interaction of Escherichia coli hemolysin: flotation and insertion-dependent labeling by phospholipid vesicles. J Bacteriol 2001; 183(18): 5364-70.
20. Sánchez-Magraner L, Viguera AR, García-Pacios M, Garcillán MP, Arrondo JL, de la Cruz F, et al. The calcium-binding C-terminal domain of Escherichia coli alpha-hemolysin is a major determinant in the surface-active properties of the protein. J Biol Chem 2007; 282(16): 11827-35.
21. Cortajarena AL, Goñi FM, Ostolaza H. A receptor-binding region in Escherichia coli alpha-haemolysin. J Biol Chem 2003; 278(21): 19159-63.
22. Valeva A, Walev I, Kemmer H, Weis S, Siegel I, Boukhallouk F, et al. Binding of Escherichia coli hemolysin and activation of the target cells is not receptor-dependent. J Biol Chem. 2005; 280(44): 36657-63.
23. Hui D, Morden C, Zhang F, Ling V. Combinatorial analysis of the structural requirements of the Escherichia colihemolysin signal sequence. J Biol Chem 2000; 275(4): 2713-20.
24. Bakás L, Maté S, Vazquez R, Herlax V. E. coli alpha-hemolysin and properties. Chapter 4. En: Ekinci D, editor. Biochemistry. Rijeka (Croacia): InTech; 2012. p. 107-40.
25. Goebel W, Hedgpeth J. Cloning and functional characterization of the plasmid-encoded hemolysin determinant of Escherichia coli. J Bacteriol 1982; 151(3): 1290-8.
26. Stanley P, Packman LC, Koronakis V, Hughes C. Fatty acylation of two internal lysine residues required for the toxic activity of Escherichia coli hemolysin. Science 1994; 266(5193): 1992-6.
27. Worsham LM, Trent MS, Earls L, Jolly C, Ernst-Fonberg ML. Insights into the catalytic mechanism of HlyC, the internal protein acyltransferase that activates Escherichia coli hemolysin toxin. Biochemistry 2001; 40(45): 13607-16.
28. Worsham LM, Langston KG, Ernst-Fonberg ML. Thermodynamics of a protein acylation: activation of Escherichia coli hemolysin toxin. Biochemistry 2005; 44(4): 1329-37.
29. Trent MS, Worsham LM, Ernst-Fonberg ML. The biochemistry of hemolysin toxin activation: characterization of HlyC, an internal protein acyltransferase. Biochemistry 1998; 37(13): 4644-52.
30. Lim KB, Walker CR, Guo L, Pellett S, Shabanowitz J, Hunt DF, et al. Escherichia coli α-hemolysin (HlyA) is heterogeneously acylated in vivo with 14-, 15-, and 17-carbon fatty acids. J Biol Chem 2000; 275(47): 36698-702.
31. Stanley P, Koronakis V, Hughes C. Acylation of Escherichia coli hemolysin: a unique protein lipidation mechanism underlying toxin fuction. Microbiol Mol Biol Rev 1998 Jun; 62(2): 309-33.
32. Ludwig A, Vogel M, Goebel W. Mutations affecting activity and transport of haemolysin in Escherichia coli. Mol Gen Genet 1987; 206(2): 238-45.
33. Nicaud JM, Mackman N, Gray L, Holland IB. The C-terminal, 23 kDa peptide of E. coli haemolysin 2001 contains all the information necessary for its secretion by the haemolysin (Hly) export machinery. FEBS Lett 1986; 204(2): 331-5.
34. Stanley P, Koronakis V, Hughes C. Mutational analysis supports a role for multiple structural features in the C-terminal secretion signal of Escherichia coli haemolysin. Mol Microbiol 1991; 5(10): 2391-403.
35. Felmlee T, Welch RA. Alterations of amino acid repeats in the Escherichia coli hemolysin affect cytolytic activity and secretion. Proc Natl Acad Sci USA 1988; 85(14): 5269-73.
36. Koronakis V, Koronakis E, Hughes C. Isolation and analysis of the C-terminal signal directing export of Escherichia coli hemolysin protein across both bacterial membranes. EMBO J 1989; 8(2): 595-605.
37. Schulein R, Gentschev I, Mollenkopf HJ, Goebel W. A topological model for the haemolysin translocator protein HlyD. Mol Gen Genet 1992; 234(1): 155-63.
38. Wandersman C, Delepelaire P. TolC, an Escherichia coli outer membrane protein required for hemolysin secretion. Proc Natl Acad Sci USA 1990; 87(12): 4776-80.
39. Wang RC, Seror SJ, Blight M, Pratt JM, Broome-Smith JK, Holland IB. Analysis of the membrane organization of an Escherichia coli protein traslocator, HlyB, a member of a large family of prokaryote and eukaryote surface transport proteins. J Mol Biol 1991; 217(3): 441-54.
40. Koronakis V, Sharff A, Koronakis E, Luisi B, Hughes C. Crystal structure of the bacterial membrane protein TolC central to multidrug efflux and protein export. Nature 2000; 405(6789): 914-9.
41. Koronakis V, Hughes C, Koronakis E. Energetically distinct early and late stages of HlyB/HlyD-dependent secretion across both Escherichia coli membranes. EMBO J 1991; 10(11): 3263-72.
42. Koronakis E, Hughes C, Milisav I, Koronakis V. Protein exporter function and in vitro ATPase activity are correlated in ABC-domain mutants of HlyB. Mol Microbiol 1995; 16(1): 87-96.
43. Young J, Holland IB. ABC transporters: bacterial exporters-revisited five years on. Biochim Biophys Acta 1999; 1461(2): 177-200.
44. Pimenta AL, Racher K, Jamieson L, Blight MA, Holland IB. Mutations in HlyD, part of the type 1 translocator for hemolysin secretion, affect the folding of the secreted toxin. J Bacteriol 2005; 187(21): 7471-80.
45. Gentschev I, Mollenkopf H, Sokolovic Z, Hess J, Kaufmann SH, Goebel W. Development of antigen-delivery systems, based on the Escherichia coli hemolysin secretion pathway. Gene 1996; 179(1): 133-40.
46. Gentschev I, Dietrich G, Goebel W. The E. coli alpha-hemolysin secretion system and its use in vaccine development. Trends Microbiol 2002; 10(1): 39-45.
47. Balsalobre C, Silvan JM, Berglund S, Mizunoe Y, Uhlin BE, Wai SN. Release of the type I secreted alpha-haemolysin via outer membrane vesicles from Escherichia coli. Mol Microbiol 2006; 59(1): 99-112.
48. Herlax V, Henning MF, Bernasconi AM, Goñi FM, Bakás L. The lytic mechanism of Escherichia coli α-hemolysin associated to outer membrane vesicles. Health 2010; 2(5): 484-92.
49. Herlax V. Interacción lípido-proteína: Mecanismo de acción de alfa hemolisina de E.coli. Tesis doctoral. La Plata: Universidad Nacional de La Plata; 2006.
50. Beveridge TJ. Structures of Gram-negative cell walls and their derived membrane vesicles. J Bacteriol 1999; 181(16): 4725-33.
51. Kuehn MJ, Kesty NC. Bacterial outer membrane vesicles and the host-pathogen interaction. Genes Dev 2005; 19(22): 2645-55.
52. Cavalieri SJ, Bohach GA, Synder IS. Escherichia coli alpha-hemolysin: characteristics and probable role in pathogenicity. Microbiol Rev 1984; 48(4): 326-43.
53. Ostolaza H, Bartolomé B, Ortiz de Zárate I, de la Cruz F, Goñi FM. Release of lipid vesicle contents by the bacterial protein toxin alpha-haemolysin. Biochim Biophys Acta 1993; 1147(1): 81-8.
54. Lally ET, Kieba IR, Sato A, Green CL, Rosenbloom J, Korostoff J, et al. RTX toxins recognize a beta2 integrin on the surface of human target cells. J Biol Chem 1997; 272(48): 30463-9.
55. Morova J, Osicka R, Masin J, Sebo P. RTX cytotoxins recognize beta2 integrin receptors through N-linked oligosaccharides. Proc Natl Acad Sci USA 2008; 105(14): 5355-60.
56. Cortajarena A, Goñi F, Ostolaza H. Glycophorin as a receptor for Escherichia coli α-hemolysin in erythrocytes. J Biol Chem 2001; 276(16): 12513-9.54.
57. Bakás L, Ostolaza H, Vaz WL, Goñi FM. Reversible adsorption and nonreversible insertion of Escherichia coli alpha-hemolysin into lipid bilayers. Biophys J 1996; 71(4): 1869-76.
58. Ostolaza H, Bakás L, Goñi FM. Balance of electrostatic and hydrophobic interactions in the lysis of model membranes by E. coli alpha-haemolysin. J Membr Biol 1997; 158(2): 137-45.
59. Sánchez-Magraner L, Cortajarena AL, Goñi FM, Ostolaza H. Membrane insertion of Escherichia coli alpha-hemolysin is independent from membrane lysis. J Biol Chem 2006; 281(9): 5461-7.
60. Bakás L, Veiga MP, Soloaga A, Ostolaza H, Goñi FM. Calcium-dependent conformation of E. coli alpha-haemolysin. Implications for the mechanism of membrane insertion and lysis. Biochim Biophys Acta 1998; 1368(2): 225-34.
61. Ostolaza H, Goñi FM. Interaction of the bacterial protein toxin alpha-haemolysin with model membranes: protein binding does not always lead to lytic activity. FEBS Lett 1995; 371(3): 303-6.
62. Soloaga A, Ostolaza H, Goñi F, de la Cruz F. Purification of Escherichia coli pro-haemolysin, and a comparison with the properties of mature alpha-haemolysin. Eur J Biochem 1996; 238(2): 418-22.
63. Herlax V, Bakás L. Acyl chains are responsible for the irreversibility in the Escherichia coli alpha-hemolysin binding to membranes. Chem Phys Lipids 2003; 122(1-2):185-90.
64. Herlax V, Bakás L. Fatty acids covalently bound to alpha-hemolysin of Escherichia coli are involved in the molten globule conformation: implication of disordered regions in binding promiscuity. Biochemistry 2007; 46 (17): 5177-84.
65. Herlax V, Mate S, Rimoldi O, Bakás L. Relevance of fatty acid covalently bound to Escherichia coli alpha-hemolysin and membrane microdomains in the oligomerization process. J Biol Chem 2009; 284(37): 25199-210.
66. Bakás L, Chanturiya A, Herlax V, Zimmerberg J. Paradoxical lipid dependence of pores formed by the Escherichia coli alpha-hemolysin in planar phospholipid bilayer membranes. Biophys J 2006; 91(10): 3748-55.
67. Brown DA, London E. Structure and origin of ordered lipid domains in biological membranes. J Membr Biol 1998; 164(2): 103-14.
68. Hamon MA, Batsche E, Regnault B, Tham TN, Seveau S, Muchardt C, et al. Histone modifications induced by a family of bacterial toxins. Proc Natl Acad Sci USA 2007; 104(33): 13467-72.
69. Ratner AJ, Hippe KR, Aguilar JL, Bender MH, Nelson AL, Weiser JN. Epithelial cells are sensitive detectors of bacterial pore-forming toxins. J Biol Chem 2006; 281(18): 12994-8.
70. Koschinski A, Repp H, Unver B, Dreyer F, Brockmeier D, Valeva A, et al. Why Escherichia coli α-hemolysin induces calcium oscillations in mammalian cells-the pore is on its own. FASEB J 2006; 20(7): 973-5.
71. Månsson LE, Kjäll P, Pellett S, Nagy G, Welch RA, Bäckhed F, et al. Role of the lipopolysaccharide-CD14 complex for the activity of hemolysin from uropathogenic Escherichia coli. Infect Immun 2007; 75(2): 997-1004.
72. TranVan Nhieu G, Clair C, Grompone G, Sansonetti P. Calcium signalling during cell interaction with bacterial pathogens. Biol Cell 2004; 96(1): 93-101.
73. Uhlén P, Laestadius A, Jahnukainen T, Söderblom T, Bäckhed F, Celsi G, et al. Alpha-haemolysin of uropathogenic E. coli induces Ca2+ oscillations in renal epithelial cells. Nature 2000; 405(6787): 694-7.
74. Schlessinger D, Schaechter M. Bacterial toxins. 2nd ed. Baltimore: Williams and Wilkins; 1993.
Aceptado para su publicación el 8 de marzo de 2013
