










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkActa bioquímica clínica latinoamericana
versión impresa ISSN 0325-2957
Acta bioquím. clín. latinoam. vol.49 no.2 La Plata jun. 2015
INMUNOLOGÍA
Estudio de la cinética de captación de ácido siálico por larvas musculares de Trichinella spiralis
Study of kinetics of sialic acid capture by muscle larvae of Trichinella spiralis
Estudo da cinética de captaçao de ácido siálico por larvas musculares da Trichinella spiralis
Gabriel López Murúa1, Liliana Racca2, Patricia Ponce de León3
1 Bioquímico.
2 Licenciada en Estadística.
3 Doctora en Ciencias Biomédicas.
Área Parasitología, Facultad de Ciencias Bioquímicas y Farmacéuticas. Universidad Nacional de Rosario. Argentina.
CORRESPONDENCIA DRA. PATRICIA PONCE DE LEÓN Área Parasitología, Facultad de Ciencias Bioquímicas y Farmacéuticas, Universidad Nacional de Rosario, Argentina. Suipacha 531 - 2000 ROSARIO, Argentina E-mail: tefu1958@hotmail.com
Resumen
Se ha comunicado que los eritrocitos (GR) incubados con larvas infectantes de Trichinella spiralis presentan mayor agregación que los GR Controles incubados con solución salina, lo cual indica que el parásito capta el ácido siálico globular. El objetivo fue estudiar la cinética de captación de ácido siálico por larvas musculares de T. spiralis aplicando Análisis Digital de Imágenes. Se trabajó con concentrados larvales de T. spiralis incubados en partes iguales con GR Grupo O, en medio salino y enzimático (GR Tratados), durante 120 minutos a intervalos de 15 minutos. Los Controles fueron incubados con igual volumen de solución salina. Se aplicó Análisis Digital de Imágenes y se calculó el valor del Coeficiente de células aisladas (CCA). Para analizar el efecto del tiempo de incubación en los valores de CCA, se utilizó un análisis de la variancia para un diseño en bloques completos aleatorizados. Los resultados mostraron que el valor promedio de CCA, en ambos medios, varió significativamente con el tiempo de incubación, lo que evidencia que el aumento del contacto de las larvas con los GR produce una disminución del ácido siálico que se refleja en mayores valores de CCA. La experiencia realizada sugeriría que durante su permanencia y viabilidad en el quiste, las larvas de T. spiralis podrían ir captando ácido siálico de la célula muscular con el objeto de interferir y/o evadir la respuesta inmune del hospedador.
Palabras claves: Trichinella spiralis; Cinética; Captación de ácido siálico.
Summary
It has been reported that erythrocytes (RBC) which were incubated with infective larvae of T. spiralis exhibit higher aggregation than RBC Controls incubated with saline solution, indicating that the parasite captures erythrocyte sialic acid. The objective of this work was to study the kinetics of sialic acid capture by muscle larvae of T. spiralis using Digital Image Analysis. The work was performed with larvae concentrates of T. spiralis, which were incubated with an equal volume of O Group RBC in saline and enzymatic mediums (Treated RBC), for 120 minutes at 15 minute intervals. Digital Image Analysis was applied and the value of Isolated Cells Coefficient (ICC) was calculated. To analyze the effect of incubation time on the ICC values, an analysis of variance was used to design randomized complete blocks. The results showed that the average value of ICC, both saline and enzymatic mediums, varied significantly with incubation time. It was shown that the increase in the contact of the larvae with erythrocytes produces the decrease of globular sialic acid, which is reflected in higher ICC values. The experience would suggest that during their stay and viability in the trichina cysts, the larvae could be capturing muscle sialic acid in order to interfere and/or evade the host immune response.
Keywords: Trichinella spiralis; Kinetics; Sialic acid capture.
Resumo
Foi relatado que os eritrócitos (GV) incubados com larvas infectantes da Trichinella spiralis apresentam maior agregação que os GV Controles incubados com soro fisiológico, indicando que o parasita capta o ácido siálico globular. O objetivo do trabalho foi estudar a cinética de captação de ácido siálico por larvas musculares da T. spiralis por Análise Digital de Imagens. O trabalho foi feito com concentrados de larvas de T. spiralis incubados em partes iguais com GV Grupo O, em meio salino e enzimático (GV Tratados), durante 120 minutos a intervalos de 15 minutos. Os controles foram incubados com igual volume de solução salina. Foi aplicada a Análise Digital de Imagens e calculado o Coeficiente de Células Isoladas (CCI). Para analisar o efeito do tempo de incubação sobre os valores do CCI foi usada uma análise da variância para um desenho em blocos completos casualizados. Os resultados mostraram que o valor médio de CCI, em ambos os meios, variou significativamente com o tempo de incubação, evidenciando que o aumento do contato das larvas com os GV produz uma diminuição do acido siálico, que se reflete em valores de CCI mais elevados. A experiência realizada sugeriria que as larvas T. spiralis durante sua permanência e viabilidade no cisto, poderiam ir captando ácido siálico da célula muscular, com o fim de interferir e /ou evitar a resposta imune do hospedeiro.
Palavras chave: Trichinella spiralis; Cinética; Captação de ácido siálico.
Introducción
Trichinella spiralis es causal de una enfermedad cosmopolita llamada triquinosis, la cual no ocasiona de manera habitual la muerte, pero disminuye considerablemente la calidad de vida del hospedador (1). La triquinosis es una enfermedad silenciosa, debido a que no presenta un cuadro clínico definido en su inicio, por lo que puede confundirse con enfermedades gastrointestinales, mientras que cuando existe una carga parasitaria importante, las manifestaciones clínicas observadas dependen del estadio de la infección (fase intestinal, sistémica o muscular); por este motivo generalmente no es considerada como un problema de salud pública, a pesar de que sigue prevaleciendo en la población por el consumo de carne de cerdo o sus derivados, proveniente de animales infectados con quistes de triquina que fueron alimentados con desperdicios o criados en contacto con basurales y roedores (2). En experiencias previas se ha comunicado que los glóbulos rojos incubados con larvas infectantes de T. spiralis presentan un aumento de agregación en relación a la producida cuando los mismos glóbulos permanecen en solución salina, indicando que el parásito capta el ácido siálico eritrocitario (3). Este hecho permitiría especular que el nematodo podría secuestrarlo a partir de otros glucoconjugados que posean residuos sializados, tal como se sugirió para Ascaris lumbricoides (4). T. spiralis presenta 3 estadios: ejemplares adultos (hembras y machos), larvas recién nacidas, y larvas infectantes, los cuales estimulan una respuesta inmunológica distinta en el hospedador, ya que presentan diferencia en la composición antigénica de la cutícula y de las secreciones, así como diferente localización en el hospedero (5).
El ácido siálico es el principal responsable de la carga eritrocitaria, pero debido a sus múltiples funciones, actualmente se reconoce su importancia biológica y su participación en la respuesta inmune (6). Muchas investigaciones han demostrado que diversos protozoos lo utilizarían para sus procesos metabólicos y/o mecanismos de evasión inmunológica (7-10), por lo que se acepta su influencia en la relación hospedero-parásito. El objetivo de este trabajo fue estudiar la cinética de captación de ácido siálico por larvas infectantes de T. spiralis aplicando Análisis Digital de Imágenes.
Materiales y Métodos
MUESTRAS
Larvas Musculares de T. spiralis: Se utilizaron larvas infectantes L1 obtenidas de músculo de ratones CBi infectados y liberadas por digestión artificial con pepsina y ácido clorhídrico (11). Las larvas fueron contadas microscópicamente por duplicado y se prepararon concentrados larvales con una cantidad de 8000 ± 500 larvas/mL. Glóbulos rojos en medio salino (GR) y enzimático de bromelina (GRB): Se trabajó con suspensiones de eritrocitos frescos Grupo O, lavados en solución salina, previa separación del plasma autólogo. Para el tratamiento enzimático se colocó un volumen de sedimento eritrocitario, previamente lavado en solución salina, con un volumen y medio de bromelina (12) durante 15 minutos en baño termostatizado a 37 ºC. A continuación los glóbulos fueron lavados con solución fisiológica.
MÉTODOS
Tratamiento de los eritrocitos: El tratamiento consistió en incubar el sedimento globular con igual volumen del concentrado larval (GR Tratados). Los eritrocitos Controles, en medio salino y enzimático, fueron incubados de la misma manera con solución fisiológica. Para evaluar el efecto de las larvas sobre la desialización se incubaron los GR Tratados y Controles desde un tiempo inicial (Tiempo 0) hasta 120 minutos, a intervalos de 15 min (37 °C). Finalizado cada uno de los tiempos, los eritrocitos fueron lavados en solución salina. Se realizaron 10 experiencias con glóbulos rojos (Tratados y los respectivos Controles) en medio salino y 10 en medio enzimático.
Distribución de agregados eritrocitarios por Análisis Digital de Imágenes (13-15): Los GR Tratados y Controles, tanto en medio salino como enzimático, fueron suspendidos en plasma autólogo al 0,13% y mantenidos en reposo durante 5 minutos para permitir la agregación. Cada suspensión fue examinada en un portaobjeto cóncavo con microscopio óptico invertido (Union Optical, Japón). Las imágenes de las diferentes poblaciones de agregados para cada muestra, fueron obtenidas por duplicado (objetivo: 40X; cámara digital Canon Powershot A640). Se calculó en las imágenes de cada muestra, el porcentaje de células individuales y se promediaron los valores obtenidos con los cuales se determinó el valor del Coeficiente de células aisladas (CCA) definido como la diferencia entre el número de células individuales iniciales (Control) y finales (Tratados) en relación al número de células aisladas observado antes del tratamiento (13)(14). Cuando el valor de CCA es igual a 0 indica que no hubo variación en el número de células aisladas por acción del tratamiento, y cuando es igual a 1 señala la ausencia de células aisladas (agregación total).
ANÁLISIS ESTADÍSTICOS
Para analizar el efecto del tiempo de incubación en los valores del coeficiente CCA en medio salino y enzimático, se utilizó un análisis de la variancia para un diseño en bloques completos aleatorizados previa verificación de los supuestos exigidos por la técnica y las comparaciones múltiples de Tukey a los fines de detectar en cuál/cuáles tiempos difiere el valor de CCA (16).
Resultados
El valor promedio de CCA, tanto en medio salino como enzimático, varió significativamente con el tiempo de incubación (p<0,0001). Los resultados mostraron que si bien en ambos casos el coeficiente alcanzó valores semejantes a las 2 horas de incubación, los cambios significativos en la desialización se produjeron a distintos tiempos de incubación según se trabajó con GR o GRB, observándose el primer cambio significativo en medio enzimático a los 15 minutos, mientras que en medio salino fue a los 45 minutos. Las comparaciones múltiples determinaron que el aumento registrado en medio salino no fue significativo entre los 0 y 15 minutos, como tampoco difirieron entre sí los valores de CCA obtenidos a los 15 y 30 min ni los correspondientes a los 75 y 90 minutos (p<0,01). En medio enzimático el aumento en los valores de CCA no difirió significativamente entre los 45 y 60 minutos, entre los 60 y 75 min, así como tampoco entre los 75 y 90 min ni entre los 105 y 120 min (p<0,01). La descripción de los datos de las experiencias realizadas se muestra en las Tablas (I y II) y en las Figuras (1 y 2).
Tabla I. Media aritmética ( - X) y desvío estándar (Sx) de los valores de CCA en las 10 suspensiones de GR en los 9 tiempos estudiados.

Tabla II. Media aritmética ( - X) y desvío estándar (Sx) de los valores de CCA en las 10 suspensiones de GRB en los 9 tiempos estudiados.

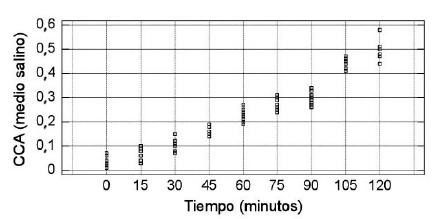
Figura 1. Distribución de los valores de CCA en medio salino, por tiempo de incubación.

Figura 2. Distribución de los valores de CCA en medio enzimático, por tiempo de incubación.
Los estudios estadísticos concluyeron que el valor promedio de CCA en medio salino y enzimático es dependiente del tiempo de tratamiento (p<0,0001), así como también se verificó que las diferentes suspensiones de eritrocitos son un factor de variación que debe tenerse en cuenta, ya que el efecto bloque (suspensión eritrocitaria) es estadísticamente significativo (p<0,05). Los resultados mostraron que el aumento del tiempo de contacto de las larvas con los eritrocitos produce una disminución del ácido siálico, que se traduce en mayores valores de CCA. La desialización ocurre más rápidamente en medio enzimático que en el salino, si bien la captación de ácido siálico en el tiempo final es semejante en ambos.
Discusión y Conclusiones
La triquinosis es una parasitosis zoonótica intestinal y tisular causada por el nematodo Trichinella, el cual infecta a una gran variedad de mamíferos, entre ellos al hombre. La forma infectante es la larva 1 (L1), que se alberga en el músculo. El miocito, rico en residuos sializados, se desarrolla como "célula nodriza", una estructura funcional diferente de cualquier otra célula de mamíferos, la cual pierde sus características de célula muscular estriada, adquiere una cápsula de colágeno, evidencia un alto grado de angiogénesis y modifica la actividad de sus organelas. Este complejo larva-célula nodriza modula de alguna manera la respuesta inmune del hospedero y puede permanecer viable durante años o eventualmente, sufrir calcificación (5)(17). El quiste se constituye por una membrana interna procedente de la fibra muscular invadida y una membrana externa homogénea y hialina, derivada del sarcolema, y sobre la superficie se desarrolla una red capilar que completa el enquistamiento de las larvas (18).
La experiencia realizada mostró que al aumentar el tiempo de contacto de las larvas musculares con los eritrocitos se produce la disminución del ácido siálico en ambos medios. Si bien el primer cambio significativo en la captación ocurrió más rápidamente en el medio deficiente en ácido siálico (enzimático) que en el medio salino, en ambos casos hubo un aumento semejante de los valores de CCA a las 2 horas de incubación. La captación del ácido siálico eritrocitario por las larvas infectantes, permitiría suponer que éstas poseen la capacidad de obtenerlo a partir de residuos sializados presentes en las células, tal como se ha comunicado para Tripanosoma cruzi (19) y se ha sugerido para A. lumbricoides (4). Los ácidos siálicos pertenecen a una familia de azúcares que se localizan en las posiciones terminales de una gran variedad de glucoconjugados. Naturalmente muestran una inmensa diversidad de estructuras que reflejan su participación en importantes procesos biológicos de crecimiento y desarrollo, tanto normal como patológico (20), participando en procesos de reconocimiento celular, vida media de células y proteínas plasmáticas, modulación del Sistema Inmune y apoptosis. También actúan como ligandos de moléculas de adhesión, como por ejemplo, selectinas o sialoadhesinas, mediando adhesión celular o transducción de señales e inhiben la vía alterna del complemento (21).
Se ha comunicado que los microorganismos que modifican sus superficies celulares con ácidos siálicos frecuentemente no son reconocidos como agentes extraños por el hospedador por lo que son capaces de evitar o inhibir la respuesta inmune. Se han descrito cuatro mecanismos de sialilación de la superficie celular en microorganismos: Síntesis de novo, Donor scavenging, Trans-sialidasa y Precursor scavenging (21)(22). Considerando que in vivo, el enquistamiento de la larva viene acompañado de una intensa respuesta inmunológica (21), que la célula muscular donde ocurre este proceso, es rica en ácido siálico y está rodeada de capilares sanguíneos (5), y que este azúcar inhibe la vía alternativa del complemento y participa activamente en los fenómenos inmunológicos (22), la experiencia realizada in vitro sugeriría que durante su permanencia y viabilidad en el quiste, las larvas de Trichinella podrían ir captando ácido siálico del músculo con el objeto de interferir y/ o evadir la respuesta inmune del hospedador.
1. Murray PR, Rosenthal K, Pfaller MA. Microbiología Médica. 5° ed. Madrid: Gea Consultoría Editorial; 2006. [ Links ]
2. Reveles Hernández R, Saldivar Elías S, Maldonado Tapia C, Muñoz Escobedo J, Moreno García M. Evaluación de la infección de Trichinella spiralis en cerdos gonadectomizados, Zacatecas, México. Acta Med Peruana 2011; 28(4): 211-6. [ Links ]
3. Ponce de León P, López Murúa G, Bertorini G, Vasconi MD, Riquelme B. Efecto biorreológico de larvas de Trichinella spiralis sobre la agregación eritrocitaria mediante Análisis Digital de Imágenes. Ciencia y Tecnología. Rosario: UNR Editora; 2013. [ Links ]
4. Ponce de León P, Di Vita S, Racca L, Biondi C, Valverde J. Extractos de Ascaris lumbricoides: alteración de la carga eritrocitaria utilizando el Método de Azul Alcian. Rev Cubana Med Trop 2011; 63(3): 263-7. [ Links ]
5. Riva E, Steffan P E, Fiel CA. Trichinellosis: aspectos múltiples de una zoonosis global. En: Mejoramiento del control de la trichinellosis. FAO América Latina y el Caribe, Roma. Organización de las Naciones Unidas para la Agricultura y la Alimentación, pp: 94-109. [ Links ]
6. Cabezas Fernández del Campo JA. Influencia de la sialilación y de la "pegilación" de la molécula de ciertos medicamentos en su actividad. An R Acad Nac Farm 2008; 74 (3): 409-29. [ Links ]
7. Nok AJ, Humprey C, Zelibe N, Yako SK. Sialidase: surface localization, properties and hydrolysis of ghost red blood cells and brain cells-identification in tripanosomiasis. Zentrabl Natur 2003; 58: 504-601. [ Links ]
8. Colli W. Trans-sialidase: a unique enzyme activity discovered in the protozoan Trypanosoma cruzi. FASEB J 1993; 7: 1257-64. [ Links ]
9. Colli W, Hemphil A, Ross CA. Flagellum-mediated adhesion of Trypanosoma congolense to bovine aorta endotelial cells. Parasitol Res 1995; 81: 412-20. [ Links ]
10. Colli W, Pasvol G, Wainscoat JS, Weatherall DJ. Erythrocytes deficient in glycophorin resist invasion by the malarial parasite Plasmodium falciparum. Nature 1982; 297: 64-6. [ Links ]
11. Shore García L, Ash L. Diagnóstico parasitológico-Manual de laboratorio clínico. 2º ed. Buenos Aires: Médica Panamericana; 1983. [ Links ]
12. Mollison PL. Transfusión de Sangre en Medicina Clinica. Madrid: Ed Reverté SA; 1987. [ Links ]
13. Foresto P, D'Arrigo M, Carrera L, Etcheparre R, Valverde J, Rasia R. Evaluation of red blood cell aggregation in diabetes by computerized image analysis. Medicina 2000; 60: 570-2. [ Links ]
14. Ponce de León P, Valverde J, Riquelme B. Acción biorreológica de extractos de Ascaris lumbricoides en eritrocitos humanos. RSVM 2012; 32: 62-9. [ Links ]
15. Ponce de León P, Del Balzo G, Riquelme B. Biorheological action of Ascaris lumbricoides larvae on human erythrocytes. Cell Biochem Biophys 2013; 65(2): 237-42. [ Links ]
16. Wackerly D, Mendenhall W, Scheaffer R. Estadística matemática con aplicaciones. 7° ed. México: Thomson Learning; 2010. [ Links ]
17. Uribarren Berrueta T. Trichinelosis o Triquinelosis. Departamento de Microbiología y Parasitología. Recursos en Parasitología. UNAM, 2013. Disponible en: http://www.facmed.unam.mx/deptos/microbiologia/parasitologia/trichinelosis.html. Fecha de acceso: 16 de noviembre de 2014. [ Links ]
18. Ramírez Valenzuela M. Epidemiología de la Trichinellosis. Ciencia Veterinaria 1981; 3: 1-58. [ Links ]
19. Souto Padron T. The surface charge of trypanosomatids. An Acad Bras Ciênc 2002; 74: 649-75. [ Links ]
20. Lehmann F, Tiralongo E, Tiralongo J. Sialic acid-specific lectins: occurrence, specificity and function. Cell Mol Life Sci 2006; 63: 1331-54. [ Links ]
21. Vimr ER, Lichtensteiger C. To sialylate, or not sialylate: that is the question. Trends Microbiol 2002; 10: 254-7. [ Links ]
22. Reglero A, Bravo I, Fernandez-Martinez V. Ácidos siálicos: distribución, metabolismo y función biológica. An R Acad Nac Farm 2007; 73(4): 833-71. [ Links ]
23. Previato L, Todeschini AR, Previato JO. Enfermedad de Chagas. Macromoléculas: Carbohidratos, lípidos y glucoproteínas. Disponible en:http://www.fiocruz.br/chagas_esp/cgi/cgilua.exe/sys/start.htm?sid=79. Fecha de acceso: 2 de mayo de 2014. [ Links ]
24. Alfonso I, Papazian O, Valencia P. Hipotonía neonatal generalizada. Rev Neurol 2003; 37 (3): 228-39. [ Links ]
25. Norris KA, Cooper NR, So M. Characterization of a Trypanosoma cruzi C3 binding protein with functional and genetic similarities to human complement regulatory protein, decay-accelerating factor. J Immunol 1991; 147 (7): 2240-7. [ Links ]
Recibido: 30 de septiembre de 2014
Aceptado: 2 de diciembre de 2014

