










Services on Demand
Journal

Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkActa bioquímica clínica latinoamericana
Print version ISSN 0325-2957
Acta bioquím. clín. latinoam. vol.49 no.3 La Plata Sept. 2015
INMUNOLOGÍA
Estudio del efecto de Trichinella spiralis sobre la desialización aplicando el Método de Polibrene
Study of Trichinella spiralis effect on desialylation by Polybrene Method
Estudo do efeito de Trichinella spiralis em desialylation aplicando o Método de Polibreno
Patricia Ponce de León1, Gabriel López Murúa2, Liliana Racca3
1 Doctora en Ciencias Biomédicas.
2 Bioquímico.
3 Licenciada en Estadística.
Área Parasitología, Facultad de Ciencias Bioquímicas y Farmacéuticas. Universidad Nacional de Rosario. Argentina.
CORRESPONDENCIA DRA. PATRICIA PONCE DE LEON Area Parasitologia, Facultad de Ciencias Bioquimicas y Farmaceuticas Universidad Nacional de Rosario. Suipacha 531 2000 ROSARIO, Argentina E-mail: tefu1958@hotmail.com
Resumen
Experiencias previas comunicaron que los eritrocitos (GR) incubados con larvas recién nacidas (LRN) y larvas musculares (LM) de T. spiralis presentan menor contenido de ácido siálico que los correspondientes GR Controles. El objetivo del trabajo fue estudiar el efecto de LRN y LM sobre la desialización aplicando el Método de Polibrene. Se trabajó con concentrados de LRN y LM incubados en partes iguales con GR Grupo O, en medio salino y enzimático, durante 120 min a intervalos de 30 min. Los Controles fueron incubados con igual volumen de solución salina. Se aplicó el Método de Polibrene y se calculó el valor del coeficiente CexpCASP. Para analizar el efecto del parásito, medio de incubación y tiempo de tratamiento en los valores de CexpCASP, se utilizó un ANOVA multifactorial. Los resultados mostraron que el medio enzimático y el aumento del tiempo de incubación producen la mayor disminución de carga globular, reflejada en menores valores de CexpCASP, sin diferencias significativas con el estadio larval utilizado. La experiencia sugeriría que durante su migración por el torrente circulatorio las LRN podrían captar ácido siálico eritrocitario y alterar el comportamiento hemorreológico, así como también permitiría especular que ambos estadios larvales podrían secuestrarlo de células del hospedador para interferir y/o evadir su respuesta inmune.
Palabras clave: Trichinella spiralis; Desialización; Método de Polibreno.
Summary
Previous experiences reported that erythrocytes (RBCs) incubated with newborn larvae (NL) and muscle larvae (ML) of T. spiralis present lower sialic acid than corresponding Control RBCs. The objective of this work was to study the effect of NL and ML on desialylation applying Polybrene Method. Work was performed with NL and ML concentrates, which were incubated with an equal volume of O Group RBCs in saline and enzymatic mediums, for 120 minutes at 30-minute intervals. Control RBCs were incubated with an equal volume of saline solution. Polybrene Method was applied and the value of CexpCASP coefficient was calculated. To analyze the effect of the parasite, incubation medium and treatment time on the CexpCASP values, a multifactorial ANOVA was used. The results showed that enzymatic medium and increasing the incubation time produce the greatest decrease in globular charge, which is reflected in lower coefficient values. No significant differences were observed with the larval stage used. The experience would suggest that during their migration through the bloodstream, the NL could capture erythrocyte sialic acid and alter the hemorrheologic behavior, and this also would allow to speculate that both larval stages could seize it from the cells of the host in order to interfere and/or evade the immune response.
Keywords: Trichinella spiralis; Desialylation; Polybrene Method.
Resumo
Experiências anteriores relataram que os eritrócitos (GV) incubados com larvas recém-nascidas (LRN) e larvas musculares (LM) da T. spiralis apresentam menor conteúdo de ácido siálico do que os correspondentes GR Controles. O objetivo do trabalho foi estudar o efeito de LRN e LM em desialylation aplicando o Método de Polybrene. Trabalhou-se com concentrados de LRN e LM incubados com igual volume com GV Grupo O, em meio salino e enzimático, durante 120 minutos a intervalos de 30 minutos. Os Controles foram incubados com o mesmo volume de solução salina. Foi aplicado o Método de Polybrene e calculado o valor do coeficiente CexpCASP. Para analisar o efeito do parasita, meio de incubação e do tempo de tratamento, nos valores do CexpCASP foi usada uma ANOVA multifatorial. Os resultados mostraram que o meio enzimático e o aumento do tempo de incubação, produzem a maior redução de carga globular, que se reflete em menores valores de CexpCASP, sem diferenças significativas com o estágio larval utilizado. A experiência sugeriria que durante sua migração através da corrente sanguínea as LRN poderiam capturar ácido siálico eritrocitário e alterar o comportamento hemorreológico, bem como permitiria também especular que ambos os estágios larvais poderiam sequestrá-lo de células do hospedeiro, com o fim de interferir e /ou evitar a resposta imune.
Palavras-chave: T. spiralis; Desialylation; Método de Polybrene.
Introducción
El nematodo Trichinella spiralis causa una enfermedad llamada trichinellosis, la cual se trasmite de modo accidental al hombre por ingestion de carne o sus derivados crudos o insuficientemente cocidos, procedentes de animales infectados. La principal fuente de infeccion humana es el cerdo. La variabilidad y la intensidad de los sintomas de esta enfermedad dependen de la carga parasitaria que afecta al individuo, la edad, sexo, estado nutricional, hormonal e inmunologico del paciente y del tejido que ha sido invadido (1). Actualmente la trichinellosis es una enfermedad endemica en Argentina, debido principalmente a las pautas culturales por las que es habitual el consumo de alimentos que contienen carne cruda o semi-cocida, utilizandose para su elaboracion cerdos faenados y procesados en el ambito familiar, sin inspeccion veterinaria (2). Los brotes debidos a T. spiralis de la especie porcina domestica siguen siendo los mas frecuentes en Argentina, pero pueden ser causados tambien por otras especies que infectan animales tales como caballos, perros, jabalies, osos y morsas. Se ha reconocido que los reptiles tambien pueden servir como hospedadores. Estas ultimas revelaciones se han producido como consecuencia de una serie de descubrimientos sobre la sistematica del parasito, facilitada por las nuevas herramientas moleculares. Esta nueva base de conocimientos, ademas de tener una importante relevancia para las politicas de seguridad alimentaria y medidas de profilaxis, plantea interrogantes importantes sobre la filogenia de Trichinella spp, las caracteristicas ecologicas de las especies y sus historias geograficas (3).
El complejo ciclo evolutivo de T. spiralis incluye la migracion de las larvas recien nacidas por el torrente circulatorio hasta su enquistamiento en musculo. Si se considera que las larvas recien nacidas, en el recorrido por la circulacion sanguinea, establecen un intimo contacto con los eritrocitos del hospedador y que las larvas musculares se enquistan en un tejido rico en residuos sializados, se podria especular que ambos estadios parasitarios podrian captar acido sialico. El acido sialico es el principal responsable de la carga eritrocitaria, pero debido a sus multiples funciones, actualmente se reconoce su importancia biologica y su participacion en la respuesta inmune (4). En experiencias previas se ha comunicado que los globulos rojos incubados con larvas de T. spiralis presentan un aumento de agregacion en relacion a la producida cuando los mismos globulos permanecen en solucion salina, indicando que el parasito capta el acido sialico eritrocitario (5).
El unico metodo directo de cuantificacion de acido sialico en el globulo rojo es su dosaje en la membrana eritrocitaria. Debido a que la implementacion de esta tecnica no es accesible a todos los laboratorios, surgieron varios metodos que permiten estudiar de manera indirecta la carga y el contenido de acido sialico superficial del eritrocito. Uno de ellos es el que utiliza bromuro de hexadimetrina (Polibrene). Este polimero sintetico, cargado positivamente, tiene la propiedad de aglutinar los eritrocitos normales, pero no los globulos con carga negativa reducida (6). El objetivo de este trabajo fue estudiar el efecto de larvas recien nacidas y larvas musculares de T. spiralis sobre la desializacion aplicando el metodo de Polibrene.
Materiales y Métodos
LARVAS RECIÉN NACIDAS DE T. Spiralis (LRN)
Se trabajo con concentrados de larvas obtenidas por incubacion de hembras gravidas en 100 μL de medio RPMI suplementado al 10% con suero fetal bovino y 250 μg/ mL de gentamicina durante 18 horas a 37 oC en atmosfera de 5% de CO2. Se realizaron los recuentos de larvas, por duplicado, en microscopio optico y se prepararon concentrados con 800 100 larvas/ mL (7).
LARVAS MUSCULARES DE T. Spiralis (LM)
Se utilizaron larvas infectantes L1 obtenidas de musculo de ratones CBi infectados y liberadas por digestion artificial con pepsina y acido clorhidrico. Las LM fueron contadas microscopicamente por duplicado y se prepararon concentrados larvales de igual cantidad de larvas a los utilizados para las LRN (LM: 800 100 larvas/ mL) (8).
GLÓBULOS ROJOS (GR) EN MEDIO SALINO Y ENZIMÁTICO DE BROMELINA
Se trabajo con suspensiones de eritrocitos frescos Grupo O. Para el tratamiento enzimatico se coloco un volumen de sedimento eritrocitario, previamente lavado en solucion salina, con un volumen y medio de bromelina (9) durante 15 min en bano termostatizado a 37 oC. A continuacion los globulos fueron lavados con solucion fisiologica.
TRATAMIENTO DE LOS ERITROCITOS
El tratamiento consistio en incubar 50 μL del sedimento globular con igual volumen del concentrado de LM/ LRN (GR Tratados). Los eritrocitos Controles, en medio salino y enzimatico, fueron incubados de la misma manera con solucion fisiologica. Para evaluar el efecto de las LRN y LM sobre la desializacion se incubaron los GR Tratados y Controles, en ambos medios, desde un tiempo inicial (Tiempo 0) hasta 120 min, a intervalos de 30 min (37 °C). Finalizado cada uno de los tiempos, los eritrocitos fueron lavados en solucion salina y se prepararon suspensiones al 20% a partir del sedimento globular. Se realizaron 10 experiencias con globulos rojos (Tratados y los respectivos Controles) en medio salino y 10 en medio enzimatico.
MÉTODO DE POLIBRENE MODIFICADO EN PLACA DE VIDRIO (6)
El metodo se aplico simultaneamente en eritrocitos no Tratados (Control) y en los mismos eritrocitos Tratados con el parasito. Se colocaron, en una placa de vidrio, 2 gotas de LIM (solucion de baja fuerza ionica), y 1 gota de la suspension eritrocitaria. Se mezclo con varilla de vidrio y se agrego 1 gota de Polibrene, volviendo a mezclar con la varilla. Se homogeinizo por movimientos circulares de la placa de vidrio durante 1 min, hasta que se produjo la agregacion entre los eritrocitos con carga negativa y el polication Polibrene. A los fines de comparar la agregacion de los globulos Tratados con la del respectivo Control, se la semicuantifico con cruces (4+, 3+; 2+: 1+; +/-; -) y se asigno un Score a cada una segun Goudemand y Marsalet (10), de acuerdo a la siguiente escala:
4 + 10 1 + 2
3 + 8 +/-1 (agregacion apenas visible)
2 + 5 - 0 (sin agregacion)
Se determino para cada tiempo de Tratamiento el Coeficiente experimental de captación de ácido siálico por el parásito usando el método de Polibrene (CexpCASP) como el cociente entre el Score de la agregacion de los eritrocitos Tratados con el parasito y el Score de la agregacion de los eritrocitos Control (11).
![]()
ANÁLISIS ESTADÍSTICOS
Se utilizo un ANOVA multifactorial para estudiar el efecto del medio (enzimatico y salino), del parasito (LM y LRN) y del tiempo de incubacion en las muestras consideradas bloques, sobre la captacion de acido sialico por medio del CexpCASP (12).
Resultados
La aplicacion de ANOVA multifactorial (medio, larva, tiempo) con bloques (muestra (n=10)) mostro que la interaccion triple no fue significativa y de las interacciones dobles la unica que debio incluirse en el modelo es la de medio (enzimatico o salino) por tiempo (p<0,05). Se verificaron los supuestos exigidos y se utilizo el test de comparaciones multiples de Tukey. Se concluyo que la captacion de acido sialico no difiere para LRN y LM a la concentracion larval estudiada (p>0,09), no existiendo el efecto larva sobre el valor del coeficiente. El valor promedio de CexpCASP fue significativamente mayor para GR en medio salino respecto del enzimatico para ambos tipos de larvas (Tabla I).
Tabla I. Valores de CexpCASP por medio y tipo de larva ( x ± sx )

Los resultados estadisticos mostraron que el tiempo de incubacion hizo disminuir el valor promedio de CexpCASP, el cual fue significativamente mayor al tiempo inicial que a los demas tiempos para los eritrocitos en ambos medios. El valor promedio del coeficiente obtenido para GR en medio salino al inicio, a los 60 y 120 min fue significativamente mayor que para GR en medio enzimatico, mientras que a los 30 y a los 90 min de incubacion no existieron diferencias significativas en la captacion de acido sialico en ambos medios (Fig. 1). En el medio salino la captacion de acido sialico eritrocitario no difirio significativamente entre los 30 y 60 min, ni entre 90 y 120 min pero si entre estos dos intervalos de tiempo. En el medio enzimatico la disminucion del coeficiente fue estadisticamente significativa entre 30 min y los demas tiempos, asi como tambien entre 60 y 120 min, pero su valor a los 90 min no difirio del valor a los 60 y 120 min de incubacion (Fig. 2).

Figura 1. Valor promedio de CexpCASP de eritrocitos en medio salino y enzimático para cada tiempo de incubación.
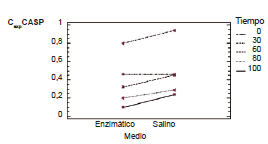
Figura 2. Valor promedio de CexpCASP por el tiempo de incubación para eritrocitos en medio enzimático y salino
Discusión y Conclusiones
De todas las relaciones que existen entre los seres vivos, el parasitismo es una de las mas complejas. El parasito es un organismo que vive a expensas de otro de distinta especie de quien obtiene sus nutrientes y morada y al cual puede eventualmente producir dano. Una clave para entender las infecciones parasitarias es el estudio de las interacciones que se establecen entre los dos organismos involucrados: el hospedador y el parasito (13). La infeccion producida por el nematodo T. spiralis es un problema mundial porque constituye una de las principales zoonosis de America latina. En Argentina se la considera una enfermedad endemica con picos epidemicos y esta incluida dentro de las enfermedades alimentarias de notificacion medica obligatoria por ley nacional desde el ano 1960 (14). La trichinellosis no solo representa un problema para la salud humana sino tambien para la salud animal, ocasionando una merma economica en terminos de productividad (15).
A pesar de la alta prevalencia global de los helmintos intestinales en humanos, y del dano economico causado por estos en animales de interes productivo, el conocimiento de la relacion hospedador-parasito es aun incompleto. La susceptibilidad a muchas enfermedades se origina en la accion reciproca de factores ambientales y geneticos del huesped, mientras que la susceptibilidad a infecciones parasitarias esta determinada por una ecuacion mas compleja que resulta de la interaccion del ambiente, el hospedador y el parasito (16). La supervivencia de los parasitos en sus hospedadores se ve permanentemente amenazada por la respuesta inmunitaria de los mismos y es por ello que se vieron obligados a generar mecanismos precisos que les permitan evadir la inmunidad y en este contexto han desarrollado numerosas estrategias de evasion (17); particularmente los helmintos han adquirido la notable habilidad de atenuar la respuesta inmune del hospedador, protegiendose de la eliminacion y minimizando la patologia producida en el huesped (18). Los acidos sialicos se situan en el extremo de las moleculas de las que forman parte y fueron hallados en vertebrados, algunos tipos de bacterias, virus e invertebrados, pero no han sido identificados en vegetales (19). Presentan importantes funciones biologicas, algunas diametralmente opuestas, entre las cuales se encuentran las debidas a su carga negativa, a su influencia sobre la estructura de macromoleculas y sobre la especificidad de antigenos, a su papel protector frente a ataques enzimaticos, a la caracteristica de tener sitios de reconocimiento y a la vez efecto de enmascaramiento (19)(20). Estos azucares juegan un papel destacado en la respuesta inmune regulando los procesos de reconocimiento celular y molecular, dado que en diversas situaciones son componentes esenciales de receptores implicados en la transduccion de senales. (17), asi como tambien participan inhibiendo la via alternativa del complemento (21-23). Actualmente se reconoce la existencia de mas de 50 estructuras de acidos sialicos y no solo se estan identificando nuevos, sino que ademas se esta profundizando en el estudio de sus funciones biologicas (4).
El Metodo de Polibrene, sencillo y rapido, permite evaluar de manera indirecta la carga y el contenido de acido sialico superficial del eritrocito. Mollison ha descripto esta tecnica en tubo (9) y Lim en placa (6), utilizando ambos autores la misma concentracion de polimero (0,1%). La aplicacion de este metodo en la experiencia realizada, permitio observar la disminucion en la cantidad de agregado formado en los globulos Tratados con respecto al correspondiente Control, indicando que el acido sialico eritrocitario disminuyo al contacto con las larvas LRN y LM. El analisis estadistico concluyo que no difirio la captacion para ambos estadios parasitarios a la concentracion larval utilizada (800 100 larvas/ mL). En experiencias previas, aplicando Analisis Digital de Imagenes tambien se observo que tanto las LM como las LRN, alteraban la agregacion eritrocitaria, pero el efecto fue mas significativo en el tratamiento con LRN (5). Debido a que se comunico que el aumento de la agregacion era dependiente de la concentracion larval (5), la diferencia con los resultados obtenidos en este trabajo podria deberse a que al realizar el estudio por Analisis Digital de Imagenes se emplearon mayores concentraciones larvales.
El valor promedio de CexpCASP fue significativamente mayor para GR en medio salino respecto del enzimatico para ambos tipos de larvas en concordancia con los resultados obtenidos previamente (5). El contacto de los eritrocitos con el parasito produjo mayor disminucion de acido sialico en globulos deficientes de carga, sugiriendo que la accion de T. spiralis sobre la desializacion seria mas relevante en infecciones que cursen simultaneamente con otra patologia que produzca deficit de acido sialico eritrocitario, como son diabetes o hipertension, tal como fuera comunicado para A. lumbricoides (11). La captacion de acido sialico producida por LRN y LM, en ambos medios, vario significativamente con el tiempo de incubacion, en concordancia con las observaciones realizadas con T. spiralis (24) y A. lumbricoides (11)(25).
Los resultados obtenidos in vitro muestran que el aumento del tiempo de contacto de las larvas con los eritrocitos y el medio enzimatico, producen la mayor disminucion de carga globular, la cual se refleja en menores valores de CexpCASP, sin diferencias significativas entre el estadio larval utilizado. La experiencia realizada permite concluir que durante su migracion por el torrente circulatorio las larvas recien nacidas podrian captar acido sialico eritrocitario y alterar el comportamiento hemorreologico, asi como tambien permitiria especular que ambos tipos de estadios larvales podrian secuestrarlo de celulas del hospedador con el objeto de interferir y/o evadir su respuesta inmune.
1. Chávez Guajardo EG, Saldivar Elías S, Muñoz Escobedo JJ, Moreno García. Trichinellosis una zoonosis vigente. REDVET 2006; 7(5) Disponible en: http://www.veterinaria.org/revistas/redvet/n060606.html (Fecha de acceso: 11 de mayo de 2015). [ Links ]
2. Caracostantogolo J, Martínez ML. Epidemiología de la trichinellosis y situación en la Argentina. Vet Argent 2009; 26(257). Disponible en: http://www.produccionanimal.com.ar/sanidad_intoxicaciones_metabolicos/parasitarias/parasitarias_cerdos/08-Trichinellosis.pdf (Fecha de acceso: 11 de mayo de 2015). [ Links ]
3. Pozio E, Murrell KD. Systematics and epidemiology of Trichinella. Adv Parasitol 2006; 63: 367-439. [ Links ]
4. Cabezas Fernández del Campo JA. Influencia de la sialilación y de la "pegilación" de la molécula de ciertos medicamentos en su actividad. An R Acad Nac Farm 2008; 74 (3): 409-29. [ Links ]
5. Ponce de León P, López Murúa G, Bertorini G, Vasconi MD, Riquelme B. Efecto biorreológico de larvas de Trichinella spiralis sobre la agregación eritrocitaria mediante Análisis Digital de Imágenes. Ciencia y Tecnología. Rosario: UNR Editora; 2013. [ Links ]
6. Lin M. A safe, simple and efficient cross matching using the slide polybrene method. Transfusion Today 2006; 66: 28. [ Links ]
7. Luebke RW. Nematodes as host resistance models for detection of immunotoxicity. Methods 41 (1): 38-47. [ Links ]
8. Shore García L, Ash L. Diagnóstico parasitológico - Manual de laboratorio clínico. 2a ed. Buenos Aires: Médica Panamericana; 1983. [ Links ]
9. Mollison PL. Transfusión de sangre en Medicina Clínica. Barcelona: Reverté; 1987. [ Links ]
10. Goudemand M, Marsalet ID. Elements d´immuno-hématologie. Paris: Flammarion; 1967. [ Links ]
11. Ponce de León P, Racca L, Menendez M, Biondi C, Valverde J. Acción de Ascaris lumbricoides sobre la carga aniónica de eritrocitos y eritrocitos desializados. Acta Bioquim Clin Latinoam 2012; 46(2): 247-56. [ Links ]
12. Devore J. Probabilidad y Estadística para ingeniería y ciencias. 7a ed. México: Editorial Latinoamericana; 2013. [ Links ]
13. Atías A. Parasitología Médica. 3a ed. Santiago: Publicaciones Técnicas Mediterráneo Ltd; 1999. [ Links ]
14. Ribicich M, Gamble HR, Rosa A, Bolpe J, Franco A. Trichinellosis in Argentina: an historical review. Vet Parasitol 2005; 132 (1-2): 137-42. [ Links ]
15. Calcagno M, Teixeira C, Forastiero MA, Constantino SN, Venturiello S. Aspectos clínicos, serológicos y parasitológicos de un brote de triquinelosis humana en Villa Mercedes, San Luis, Argentina. Las fases aguda y crónica de la infección. Medicina 2005; 65(4): 302-6. [ Links ]
16. Nino Incani R, Morales G, Cesari IM. Parasite and vertebrate host genetic heterogeneity determine the outcome of infection by Schistosoma mansoni. Parasitol Res 2001; 87:131. [ Links ]
17. Fainboim L. Introducción a la Inmunología Humana. 6ª ed. Buenos Aires: Médica Panamericana; 2011. [ Links ]
18. Maizels RM, Yazdanbakhsh M. Immune regulation by helminth parasites: cellular and molecular mechanisms. Nat Rev Immunol 2003; 3: 733. [ Links ]
19. Silva JM, Saldahna C. Biorreologia e Hemorreologia- Origens e Evolução. SPHM 2005; 20(3): 5-11. [ Links ]
20. Cabezas JA. Neuraminic acid. - IV. Gas-Liquid Chromatography of N-Acetyl- and N-Glycolylneuraminic Acids. Rev Esp Fisiol 1965; 21: 125-30.
21. Buscaglia C. Trans- sialidasa de Tripanosoma cruzi: un blanco potencial para el tratamiento de la enfermedad de Chagas. Rev Hosp Mat Inf Ramón Sardá 2002; 21(1): 24-7. [ Links ]
22. Chabalgoity J, Pereira M, Rial A. Inmunidad contra los agentes infecciosos. Disponible en: http://www.higiene.edu.uy/cefa/2008/Inmunidadcontralosagentesinfecciosos.pdf (Fecha de acceso 11 de mayo de 2015). [ Links ]
23. Murray PR, Rosenthal K, Pfaller MA. Microbiología Médica. 2° ed. Madrid: Gea Consultoría Editorial; 2006. [ Links ]
24. López Murúa G, Racca L, Ponce de León P. Estudio de la cinética de captación de ácido siálico por larvas musculares de Trichinella spiralis. Acta Bioquím Clín Latinoam 2015; 49 (2): 267-72. [ Links ]
25. Ponce de León P, Juárez Matamoros K, Biondi C, Valverde J. Alteración de la carga aniónica superficial de glóbulos rojos y glóbulos rojos desializados por Ascaris lumbricoides. Rev Cubana Med Trop 2011; 63 (1): 87-90. [ Links ]
Recibido: 24 de abril de 2015
Aceptado: 11 de mayo de 2015

