










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkActa bioquímica clínica latinoamericana
Print version ISSN 0325-2957On-line version ISSN 1851-6114
Acta bioquím. clín. latinoam. vol.53 no.2 La Plata June 2019
TOXICOLOGÍA
Micetismos. Parte 3: Síndromes tempranos gastrointestinales
Alicia Beatriz Pomilio*1a, Stella Maris Battista2b, Ángel Alonso3c
1 Doctora de la Universidad de Buenos Aires (Ph. D.), Investigadora Superior del CONICET. Profesora de la Universidad de Buenos Aires.
2 Médica. Doctorando en la Facultad de Medicina, Universidad de Buenos Aires. Docente en la Facultad de Medicina (UBA).
3 Doctor en Medicina (Ph. D.), Médico, Facultad de Medicina, Universidad de Buenos Aires (UBA). Profesor Emérito UBA. Investigador en Histología, Microbiología e Inmunología.
a Departamento de Bioquímica Clínica, Área Hematología, Hospital de Clínicas “José de San Martín”, Universidad de Buenos Aires, Av. Córdoba 2351, C1120AAF Buenos Aires, Argentina. Tel: (+54) (11) 4814 3952. abpomilio@sinectis.com.ar; pomilio@ffyb.uba.ar
b Microbiología I, Cátedra 2 (Inmunología), Facultad de Medicina, Universidad de Buenos Aires. Correo electrónico: battistasm@yahoo.com.ar
c División Alergia e Inmunología, Hospital de Clínicas “José de San Martín”, Universidad de Buenos Aires. Correo electrónico: administracion@cientifica.org.ar
* Autor para correspondencia.
Correspondencia Prof. Dra. ALICIA B. POMILIO Departamento de Bioquímica Clínica Área: Hematología Hospital de Clínicas “José de San Martín” Universidad de Buenos Aires Av. Córdoba 2351 C1120AAF CIUDAD AUTÓNOMA DE BUENOS AIRES, República Argentina. Correo electrónico: pomilio@ffyb.uba.ar
Resumen
En esta Parte 3 de la serie de cuatro artículos sobre micetismos se analizan los síndromes tempranos con síntomas gastrointestinales que se caracterizan por presentar un período de latencia muy corto, de menos de 6 horas después de la ingestión de los macromicetos. Los restantes síndromes tempranos con sintomatología compleja serán tratados en la Parte 4 de la serie. Actualmente se conocen más de 200 especies responsables de síndromes gastrointestinales, pero en este trabajo se abordarán solamente diez ejemplos que involucran los géneros Boletus [Boletus satanas (o Rubroboletus satanas) y Boletus venenatus (o Neoboletus venenata)], Hypholoma, Agaricus (Agaricus xanthodermus), Omphalotus, Lactarius, Russula, Entoloma, Chlorophyllum (Chlorophyllum molybdetes) y Leucoprinus (Leucoprinus birnbaumii). Las toxinas involucradas en estos casos presentan gran variedad estructural, desde proteínas hasta terpenoides, en particular sesquiterpenoides y triterpenoides, vinilglicina, fenol y azocompuestos, pero todas generan la misma sintomatología. Estas sustancias y otros componentes químicos de los hongos suelen ser indigestos, con una susceptibilidad variable entre los consumidores. El tratamiento es de apoyo y es estrictamente para esos casos con cuadros más graves de deshidratación. Normalmente, los casos evolucionan favorablemente después de 12 a 48 horas. Se analizan los síntomas, las toxinas involucradas, los mecanismos de acción, cuando se conocen y las especies causantes de los micetismos.
Palabras clave: Micetismos tempranos; Síndromes gastrointestinales; Latencia; Boletus; Hypholoma; Agaricus; Omphalotus; Lactarius; Russula; Entoloma: Chlorophyllum; Leucoprinus; Toxinas; Tratamiento.
Abstract
Mushroom poisonings. Part 3: early-onset gastrointestinal syndromes
This part 3 of the series of four articles on mushroom poisoning refers to early-onset gastrointestinal syndromes, which are characterized by a very short latency period of less than 6 hours after mushroom ingestion. The remaining early-onset syndromes with complex symptoms will be treated in Part 4 of the series. Currently, more than 200 species responsible for gastrointestinal syndromes are known, but in this paper only ten examples will be addressed involving the genera Boletus [e.g., Boletus satanas (or Rubroboletus satanas), and Boletus venenatus (or Neoboletus venenata)], Hypholoma, Agaricus (e.g., Agaricus xanthodermus), Omphalotus, Lactarius, Russula, Entoloma, Chlorophyllum (e.g., Chlorophyllum molybdetes), and Leucoprinus (e.g., Leucoprinus birnbaumii). The toxins involved in these cases have a great structural variety, from proteins to terpenoids, in particular sesquiterpenoids and triterpenoids, vinylglycine, phenol, and azocompounds, but all show the same symptoms. These substances and other mushroom chemical constituents are usually indigestible, with varying consumer susceptibility. The treatment is supportive and is strictly for those cases with more severe dehydration. Usually, the cases progress favourably after 12 to 48 hours.The symptoms, toxins involved, mechanisms of action when known, and the species of mushrooms responsible for the mycetisms are analysed.
Keywords: Early-onset mycetisms; Gastrointestinal syndromes; Latency period; Boletus; Hypholoma; Agaricus; Omphalotus; Lactarius; Russula; Entoloma; Chlorophyllum; Leucoprinus; Toxins; Treatment.
Resumo
Intoxicações por cogumelos. Parte 3: Síndromes precoces gastrointestinais
Nesta parte 3 da série de quatro artigos sobre intoxicação por cogumelos são analisadas as síndromes precoces com sintomas gastrointestinais que se caracterizam por apresentar um período de latência muito curto, de menos de 6 horas, após a ingestão de cogumelos. As síndromes precoces restantes com sintomatologia complexa serão tratadas na Parte 4 da série. Atualmente, são conhecidas mais de 200 espécies responsáveis por síndromes gastrointestinais, mas neste trabalho serão abordados apenas dez exemplos que envolvem os gêneros Boletus [Boletus satanas (ou Rubroboletus satanas) e Boletus venenatus (ou Neoboletus venenata)], Hypholoma, Agaricus (Agaricus xanthodermus), Omphalotus, Lactarius, Russula, Entoloma, Chlorophyllum (Chlorophyllum molybdetes) e Leucoprinus (Leucoprinus birnbaumii). As toxinas envolvidas nestes casos têm uma grande variedade estrutural, desde proteínas até terpenóides, em particular sesquiterpenóides e triterpenóides, vinilglicina, fenol e azo compostos, mas todas apresentam a mesma sintomatologia. Essas substâncias e outros constituintes químicos dos cogumelos costumam ser indigestos, com uma suscetibilidade variável entre aqueles que os consomem. O tratamento é de suporte e é rigorosamente para esses casos com quadros mais graves de desidratação. Normalmente, os casos evoluem favoravelmente após 12 a 48 horas. São analisados os sintomas, as toxinas envolvidas, os mecanismos de ação, quando conhecidos, e as espécies de cogumelos responsáveis pelas intoxicações.
Palavras-chave: Intoxicações precoces por cogumelos; Síndromes gastrointestinais; Latência muito curta; Boletus; Hypholoma; Agaricus; Omphalotus; Lactarius; Russula; Entoloma; Chlorophyllum; Leucoprinus; Toxinas; Tratamento dos pacientes.
1. Introducción
En esta Parte 3 de la serie de cuatro artículos sobre micetismos (1)(2), se analizan los síndromes tempranos del grupo 3 que comprenden toxíndromes gastrointestinales o resinoides, es decir que la sintomatología se basa en trastornos gastrointestinales de variada gravedad, que ocurren con un tiempo de latencia muy corto, generalmente inferior a 6 horas después de la ingestión de los carpóforos de hongos superiores.
El grupo 3, Parte 3, trata sobre los siguientes micetismos tempranos:
Toxíndromes gastrointestinales o resinoides. Micetismo por irritantes gastrointestinales.
a) Glicoproteínas (bolesatina y boleverina) y lectinas tóxicas en especies del género Boletus: Boletus satanas (o Rubroboletus satanas) y Boletus venenatus (o Neoboletus venenata).
b) Fasciculoles en especies del género Hypholoma.
c) Fenol y azocompuestos en Agaricus xanthodermus.
d Triterpenos tóxicos en especies del género Hebeloma.
e) Iludinas en especies del género Omphalotus.
f) Sesquiterpenos cíclicos en especies de los géneros Lactarius y Russula.
g) Vinilglicina en Entoloma sinuatum y otras especies de Entoloma.
h) Toxina hemolítica rubescenslisina en el hongo comestible Amanita rubescens.
i) Proteína tóxica en Chlorophyllum molybdites.
j) Úlceras gástricas por consumo de Leucoprinus birnbaumii.
El grupo 3, Parte 4, se referirá a los siguientes micetismos tempranos:
1) Toxíndrome muscarínico o sudoriano. Micetismo colinérgico.
2) Toxíndrome inmunohemolítico o hemolítico. Micetismo por Paxillus.
3) Toxíndrome neumónico alérgico. Licoperdonosis o micetismo por Lycoperdon perlatum y por Pholiota nameko.
4) Toxíndrome panterínico o delirante. Micetismo por compuestos isoxazólicos o síndrome pantherina/muscaria.
5) Toxíndrome coprínico o cardiovascular. Micetismo inducido por alcohol o reacción tipo disulfiram.
6) Toxíndrome psicotrópico o alucinogénico. Micetismo por derivados indólicos.
7) Toxíndrome psicotrópico por estirilpironas y gimnopilinas. Micetismo debido al hongo alucinogénico Gymnopilus spectabilis o G. junonius.
8) Toxíndrome agudo de rabdomiólisis. Micetismo por Russula subnigricans.
9) Toxíndrome debido a hongos cianogénicos, como Marasmius oreades.
10) Toxíndrome inmunosupresor. Micetismo por tricotecenos macrocíclicos de Podostroma cornu-damae.
11) Toxíndrome debido a ostreolisina de Pleurotus ostreatus.
2. Análisis de cada síndrome del micetismo principal considerado
Grupo 3, Parte 3: Micetismos tempranos gastrointestinales
2.1. Toxíndromes gastrointestinales o resinoides. Micetismo por irritantes gastrointestinales
Se conocen varias especies de hongos que producen toxíndromes gastrointestinales debido a que poseen compuestos que son irritantes de todo el tracto gastrointestinal, que si bien tienen estructuras químicas muy distintas, todos ellos dan lugar a síntomas similares.
Es por eso que en esta sección, primeramente se indican los tiempos de latencia y las características sintomatológicas y luego se procede a tratar en subsecciones las toxinas, sus estructuras y mecanismos de acción, cuando se conocen, y las especies causantes del micetismo.
Período de latencia. Entre veinte minutos a media hora y hasta 4 horas después de la ingestión.
Síntomatología general. Los síntomas se manifiestan con náuseas, malestar, cólicos y vómitos, acompañados, en ocasiones, de dolores abdominales y diarrea. La mayoría de las veces estos micetismos se resuelven en uno o dos días y el pronóstico es bueno. La complicación más común es la deshidratación debido a la pérdida importante de líquidos en vómitos y diarrea, que es necesario tratar inmediatamente. Puede ser una experiencia muy desagradable. La respuesta personal en este tipo de intoxicaciones es muy variada, desde un simple dolor y ligeras molestias, hasta graves cuadros de diarrea y deshidratación.
Algunos hongos han sido causantes de intoxicaciones gastrointestinales más graves, como es el caso de Entoloma sinuatum, el más peligroso de todos, que se tratará en la subsección 2.1.7. y que ha provocado lesiones hepáticas graves y alteraciones nerviosas, con gran debilidad de los pacientes en el período de convalecencia, y mortalidad en ocasiones.
Tricholoma pardinum y Tricholoma filamentosum son también peligrosos y causantes de los trastornos gastrointestinales mencionados, junto a lesiones hepáticas, uremia, shock y hemorragias digestivas, y dan cuadros de intoxicaciones graves, pero raramente mortales. T. pardinum a veces se confunde con Tricholoma terreum que es más grande y se encuentra en bosques altos, pero que produce un micetismo demorado con rabdomiólisis como se ha visto en la Parte 2 de esta serie de artículos (2).
Toxinas. Los irritantes gastrointestinales son los que provocan la forma más común de intoxicación por ingestión de macromicetos. Para ciertas especies no se conocen todavía las sustancias que causan la intoxicación, pero en algunas de ellas se han aislado compuestos con estructuras muy diversas, que intervienen en este tipo de trastornos gastrointestinales y que se tratarán en las subsecciones siguientes.
2.1.1. Glicoproteínas (bolesatina y bolevenina) y lectinas tóxicas en especies del género Boletus: Boletus satanas (o Rubroboletus satanas) y Boletus venenatus (o Neoboletus venenata)
Este micetismo es causado por especies del género Boletus, que es un género que en los últimos años sufrió modificaciones taxonómicas sobre la base de consideraciones morfológicas y moleculares y pasó en parte a otros géneros como Rubroboletus, Neoboletus, Caloboletus, Imperator, entre otros.
El representante más conocido causante de este síndrome es Rubroboletus satanas (“bolete de Satanás” o “boleto del diablo”) que hasta 2014 fue conocido científicamente como Boletus satanas, descripto así por el micólogo alemán Harald Othmar Lenz en 1831, quien se sintió mal por probarlo (3)(4).
El análisis genético publicado en 2013 mostró que Boletus satanas y muchos otros boletes (pero no todos) que formaban parte del clado Dupainii debían colocarse en un nuevo género (5), por lo que fue transferido a Rubroboletus en 2014 junto con otras especies relacionadas en base a evidencia morfológica y molecular (6).
Rubroboletus satanas se encuentra en suelos calcáreos en bosques mixtos en todas las zonas templadas, especialmente en el sur de Europa, en el sur de Inglaterra y en algunas islas del Mar Báltico (3), en Turquía (7), en Asia menor (8), en áreas costeras de California y sudeste de los Estados Unidos (9).
Los cuerpos fructíferos, achaparrados y de colores brillantes son grandes e imponentes, con un sombrero aterciopelado de color pálido de hasta 30 cm de ancho, poros de color rojo sangre y tallos bulbosos de color rojo. La carne toma un tono grisáceo y después azul cuando se corta o se lastima. Su olor es desagradable, a carroña, más notable con la edad. Su sabor es dulce y los especímenes jóvenes tienen un agradable sabor a nueces. Es el bolete más grande que crece en Europa.
Es posible confundirlo con especies cercanas, como: a) Boletus barrowsii; b) Neoboletus luridiformis; c) Caloboletus calopus; d) Rubroboletus rhodoxanthus; e) Rubroboletus legaliae; f) Imperator torosus; g) Rubroboletus eastwoodiae; h) Suillellus luridus; i) Imperator rhodopurpureus, que tampoco tienen valor como comestibles, ya que resultan indigestos o ligeramente tóxicos.
Sintomatología. R. satanas es un hongo tóxico, que causa síntomas predominantemente gastrointestinales con náuseas y vómitos violentos, que pueden durar hasta seis horas, y que se producen si se consumen crudos o fritos (la fritura no elimina la toxina), mientras que cocido es indigesto. También otros intoxicados presentaron náuseas, debilidad, mareos, ansiedad, visión borrosa, pérdida de audición, tinnitus y vómitos violentos (4).
Muchas veces no es consumido simplemente por su aspecto extraño y por el olor pútrido que desalienta su recolección (10). No obstante, se ha consumido tradicionalmente en algunas regiones de Italia y en la antigua Checoslovaquia con una cocción completa. En 2012 se registraron casos de intoxicación en Suiza por ingestión de R. satanas, con síntomas gastrointestinales graves, como vómitos recurrentes y diarrea con sangre (11).
Toxinas. La bolesatina es una potente glicoproteína monocatenaria tóxica de 63 kDa y un punto isoeléctrico pI de 8,3 [forma un puente disulfuro intracadena (12)], que fue aislada de R. satanas (12)(13). Su secuencia de aminoácidos N-terminal se estableció como: NH2-Thr- Trp-Arg-Ille-Tyr-Leu-Asn-Asn-Gln-Thr-Val-Lys-Leu-Ala- Leu-Leu-Leu-Pro-Asn-Gly (14). Como es una proteína no sorprende que el calor la desnaturalice y pierda su actividad biológica. Se estudió la distribución de la bolesatina marcada con 14C en ratones; poco después de la administración, quedaron marcados estómago e intestinos, riñones e hígado, y un poco más tarde bazo y pulmones (15). Se ha demostrado por monitoreo de la bolesatina en fracciones subcelulares de hígado y riñón de rata que está presente en todas las fracciones: citoplasma, mitocondrias, ribosomas, microsomas, incluso en el núcleo. La bolesatina se excreta tanto por orina como por heces y el 80% se elimina en las primeras 24 horas después de la intoxicación. Ninguna proteasa común (tripsina, quimotripsina, pronasa, proteinasa K) hidroliza a la bolesatina nativa ni es desnaturalizada por calor (15).
También se aisló muscarina de los carpóforos, pero en cantidades demasiado pequeñas como para producir toxicidad (10).
La bolevenina es una proteína tóxica que se aisló del hongo tóxico Boletus venenatus de Japón, denominado Neoboletus venenata desde 2016 (16) y que presentó su actividad letal después de una inyección de 10 mg/kg en ratones (4). En SDS-PAGE, en presencia o ausencia de 2-mercaptoetanol, esta proteína mostró una única banda de ~12 kDa. Por el contrario, en base a la filtración en gel y a través de la espectrometría de masa con ionización por desorción con láser asistida por matriz y tiempo de vuelo (MALDI-TOF-MS), su masa molecular relativa se estimó en ~30 kDa y ~33 kDa respectivamente, lo que indica que la proteína consta de tres subunidades idénticas. También se determinaron su pI y su secuencia de aminoácidos N-terminal hasta 18 que resultó similar a la de bolesatina (17).
Esta toxina corresponde a una familia de isolectinas que mostraron una sola banda en SDS-PAGE, pero se dividieron después en ocho isolectinas mediante cromatografía de intercambio aniónico, con pesos moleculares muy similares a los obtenidos por MALDI-TOF MS (18). Se analizó la especificidad de unión a azúcares de cada isolectina, indicando que reconocían las cadenas de azúcar unidas a N, especialmente los residuos Galβ1 → 4GlcNAcβ1 → 4Manβ1 → 4GlcNAcβ1 → 4GlcNAc (Tipo II). Las lectinas de B. venenatus causaron toxicidad mortal en ratones por vía intraperitoneal y la ingestión provocó toxicidad fetal y diarrea en ratones y también diarrea en ratas (18).
Mecanismo de acción de la bolesatina. Se demostró que la bolesatina inhibe la síntesis de proteínas en varios sistemas in vitro e in vivo. Así, por ejemplo, en mitocondrias aisladas de ratas, después de 90 minutos de preincubación con bolesatina, se inhibió la incorporación de [14C]-leucina en las proteínas mitocondriales con una CI50 (concentración de inhibición al 50%) de 530 nM (14).
Para una mejor comprensión de su mecanismo de acción sobre la síntesis de proteínas a nivel ribosómico, se trataron ribosomas de hígado de rata con bolesatina (1 a 10 μg) agregada a los sistemas de traducción de ácido poliuridílico in vitro antes y después del lavado (19). El hecho que los ribosomas todavía estuvieran activos confirmó que la bolesatina no actúa como los inhibidores de síntesis de proteínas de origen vegetal, conocidos como proteínas inactivadoras de ribosomas. Otros estudios moleculares lo confirmaron (20).
Por estudios in vitro se demostró que la bolesatina tampoco tiene un efecto directo sobre los factores de elongación, pero hidroliza los trifosfatos de nucleósidos, GTP y ATP, con la consiguiente inhibición de la síntesis de proteínas. Por lo tanto, la bolesatina debe clasificarse como una fosfatasa de trifosfatos de nucleósidos, más que como un inhibidor directo de la síntesis de proteínas (19).
Cuando la bolesatina se administra a ratones causa una trombosis masiva (21) dado que tiene propiedades de lectina (22). Las lectinas forman un gran grupo de proteínas de origen no inmune que pueden reconocer y unir azúcares, tanto libres como unidos a proteínas (glicoproteínas) o a lípidos (glicolípidos), con un alto grado de especificidad. No catalizan ninguna reacción química, no muestran actividad enzimática. Las lectinas participan en muchos procesos que requieren reconocimiento específico (reacciones inmunológicas, contacto con tejidos, interacciones huésped-patógeno, etc.) y tienen potencial aplicación biotecnológica y biomédica. Las lectinas que reconocen unidades de azúcar en las paredes o en las membranas celulares alteran su fisiología y desencadenan cambios bioquímicos en la célula. Por lo tanto, una aplicación de las lectinas, como la bolesatina y bolevenina, es la identificación de glicanos aberrantes expresados por células neoplásicas y como agentes antitumorales mediante la inducción de apoptosis por diversos mecanismos (23). Se demostró que la bolesatina inhibe la síntesis de proteínas in vitro de una manera dependiente de la concentración en una línea celular de un linfosarcoma tímico inducido por radiación (SP2/O) (24). In vivo, en ratones Balb/c con un tumor ascítico inducido por preinyección i.p. de células SP2/O permite una remisión de 50% y 30% respectivamente. Los ratones tratados sobrevivieron 120 días después del tratamiento, es decir, 90 días después de la muerte del animal control.
La DL50 (dosis letal al 50%) de la bolesatina en ratones por vía oral es de 3,3 mg/kg (15); por vía intraperitoneal (i.p) en ratones o ratas es de 1 mg/kg y por vía intravenosa (i.v) es de 0,14 mg/kg (21). La bolesatina es ~100 veces menos tóxica que la ricina, pero con una DL50 menor de 5 mg/kg y se considera dentro de las llamadas “sustancias supertóxicas” (4).
La bolesatina, además de inhibir la síntesis de proteínas, tiene una actividad mitogénica a concentraciones muy bajas en linfocitos humanos que es unas 200 veces mayor que la de otras lectinas (25). Además, la bolesatina induce la peroxidación lipídica y activa la producción de radicales libres y contribuye así a la inducción de muerte celular programada (26). La bolesatina también causa aglutinación de glóbulos rojos y plaquetas (27), lo que conduce a la formación de trombos en la sangre.
Especies causantes del micetismo. Reino: Fungi; División: Basidiomycota; Clase: Agaricomycetes; Orden: Boletales; Familia: Boletaceae: Rubroboletus satanas (Lenz) Kuan Zhao & Zhu L. Yang (6) [Basónimo: Boletus satanas Lenz; sinónimos: Suillus satanas (Lenz) Kuntze; Tubiporus satanas (Lenz) Maire]; Boletus barrowsii Thiers & A.H. Sm.; Imperator rhodopurpureus (Smotl.) Assyov (28) [Bas.: Boletus rhodopurpureus (Smotl.)]; Imperator torosus (Fr.) A ssyov (28) [Bas.: B. torosus Fr.; sin: Dictyopus torosus (Fr.) Quél.; Boletus purpureus var. xanthocyaneus Romagn.; B. xanthocyaneus (Ramain) Romagn.]; Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini [Bas.: Boletus luridiformis Rostk.; sin.: Suillus luridiformis (Rostk.) Kuntze; B. erythropus Pers.]; Neoboletus venenatus (Nagas) G. Wu & Zhu L. Yang (16) [Bas.: Boletus venenosus Nagas]; Caloboletus calopus (Pers.) Vizzini; Rubroboletus eastwoodiae (Murrill) Arora, Schwarz & Frank (Boletus eastwoodiae Murill hasta 2015); Rubroboletus legaliae (Pilát & Dermek) Della Maggiora & Trassinelli (29) [Bas.: Boletus legaliae Pilát; sin: B. splendidus C.Martín; B. spinari Hlaváček]; Rubroboletus rhodoxanthus (Krombh.) Kuan Zhao & Zhu L. Yang (6) conocido como B. rhodoxanthus hasta 2014 [Sin: B. sanguineus var. rhodoxanthus Krombh.]; Suillellus luridus (Schaeff.) Murrill [Bas.: Boletus luridus Schaeff.; sin: B. rubeolarius Bull.; B. subvescus J.F. Gmel.; Leccinum luridum (Schaeff.) Gray; Tubiporus luridus (Schaeff.) P. Karst.; Dictyopus luridus (Schaeff.) Quél.].
2.1.2. Fasciculoles en especies del género Hypholoma
El hongo basidiomiceto Hypholoma fasciculare es el responsable de este síndrome gastrointestinal. Es tóxico, tiene sabor amargo y emite un olor desagradable, como a iodo o azufre. Suele encontrarse en grandes grupos, principalmente sobre tocones de coníferas o de árboles caducifolios. Tiene importancia forestal ya que su micelio, principalmente saprótrofo, se alimenta degradando la madera caducifolia en descomposición debido a su menor contenido de lignina en relación con la de coníferas (30). Además de la madera muerta, también infesta árboles vivos y hace que mueran en unos pocos años.
H. fasciculare es uno de los hongos más comunes en los bosques de Europa, es el más frecuente en Gran Bretaña y abundante en el norte de Europa, Irán, Turquía (7), en Asia (Japón, China y Corea), en América del Norte y en América del Sur, especialmente en la Argentina. Otras especies de Hypholoma crecen en Colombia y Brasil. Se ha utilizado con éxito en silvicultura como un tratamiento experimental para desplazar competitivamente una enfermedad fúngica común de las coníferas, Armillaria solidipes, en bosques de coníferas (31).
Hay muchas especies de Hypholoma relacionadas, como: Hypholoma capnoides, que es comestible, así como H. sublateritium tóxica e H. epixanthum tóxica.
Sintomatología. Los síntomas por ingestión de H. fasciculare pueden retrasarse de 5 a 10 horas tras el consumo, después de lo cual puede haber diarrea, náuseas, vómitos, proteinuria y colapso. Se han registrado parálisis y problemas de visión. Los síntomas generalmente se resuelven en unos pocos días.
Sus toxinas (fasciculoles) actúan sobre el estómago y el intestino delgado y provocan vómitos y diarrea; sin embargo, no ocurre daño permanente. Hace algún tiempo se creía que el hongo era mortalmente tóxico, ya que los compuestos tóxicos causaron parálisis y muerte en ratones. Hasta ahora no se ha demostrado un efecto similar en humanos.
Un extracto de H. fasciculare presentó actividad inhibidora sobre la trombina (32).
Toxinas. Este hongo es bien conocido por sus diversos componentes terpenoidales (ergosteroles, triterpenoides y sesquiterpenoides) (33-37) y particularmente por la presencia de triterpenoides con esqueleto de lanostano (lanosteroides) como componentes tóxicos (33-35)(38)(39). Estos lanostanos inhiben el crecimiento de las plántulas de repollo chino y tienen actividad antimicrobiana y de inhibición de calmodulina (33-35).
En 1983 se aislaron e identificaron los lanostanos tóxicos: fasciculol E y fasciculol F (Fig. 1) de los carpóforos de H. fasciculare o Naematoloma fasciculare, que causaron parálisis y muerte en ratones, con valores de DL50 en ratones de 50 mg/kg y 168 mg/kg (i.p.) respectivamente (33).

Figura 1. Estructuras químicas de fasciculol E y fasciculol F.
Se aislaron 15 triterpenoides tipo lanostano (lanosteroides) (1-15) (Fig. 1)(Fig. 2) del extracto metanólico de H.fasciculare (39): Fasciculol J (1), fasciculol K (2), fasciculol L (3), fasciculol M (4) y fasciculol G (5) (Fig. 2), fasciculol E (6)(35), ácido fascicúlico B (7)(35), ácido fascicúlico F (8), fasciculol D (9), fasciculol F (10), fasciculol B (11), fasciculol C (12), fasciculol H (13)(38), fasciculol I (14)(38) y sublateriol C (15)(40) (Fig. 1)(Fig 2).

Figura 2. Estructuras químicas de los compuestos triterpénicos tipo lanostano 1-5, 7-9 y 11-15.
Todos los triterpenoides aislados (Fig. 1)(Fig. 2), excepto 1 y 2, mostraron actividad citotóxica contra las líneas celulares de cáncer humano A549, SK-OV-3, SK-MEL-2 y HCT-15 con valores de CI50 (concentración inhibitoria al 50%) de 2,29-9,86 μM. Los compuestos 11, 12 y 15 mostraron una considerable citotoxicidad contra estas líneas celulares, destacándose la actividad del compuesto 12 (CI50: 2,37, 2,82, 2,29 y 3,06 μM respectivamente) (39).
La presencia de un grupo 3-hidroxi-3-metilglutarilo unido a C-3 en los compuestos 1 y 2 redujo la actividad marcadamente en comparación con los compuestos 7 y 8, con la cadena lateral conectada a C-2. Los compuestos 13 y 14, con una funcionalidad de cetona en C-12, presentaron una buena citotoxicidad selectiva contra la línea celular SK-OV-3 [CI50 (13): 9,86; CI50 (14): 7,98 μM] (39).
Considerando que la progresión del cáncer está asociada con respuestas inflamatorias, también se evaluó la actividad antiinflamatoria de quince lanosteroides, examinando sus efectos inhibidores sobre la producción de óxido nítrico en una línea celular microglial murina activada por lipopolisacárido. Entre los compuestos ensayados, sólo 4 y 7 inhibieron moderadamente la producción de óxido nítrico, sin toxicidad celular (39).
También de los carpóforos de H. fasciculare se aislaron ergosterol, peróxido de ergosterol (5α,8α-epidioxi-22E-ergosta-6,22-dien-3β-ol; contenido muy alto: al menos 0,23%) (36) y tres derivados de ergosterol: (22E,24R)- ergosta-7,22-dien-3β,5α,6β,9α-tetrol, (22E,24R)-5α,8α- epidioxiergosta-6,22-dien-3β-ol 3-O-β-D-glucopiranósido y (22E,24R)-5α,8α-epidioxiergosta-6,9,22-trien-3β-ol 3-O-β- D-glucopiranósido. Se analizó su actividad citotóxica frente a cuatro líneas celulares de cáncer humano (A549, SK-OV-3, SK-MEL-2 y HCT-15) (41).
Mecanismo de acción. Varios lanosteroides detienen el ciclo celular en G1 o en G2/M, afectan a las proteínas transportadoras MDR y MRP, aumentan los niveles de p53 y Bax, disminuyen la expresión de MMP, inducen la apoptosis, o inhiben no sólo la fosforilación de Erk1/2, sino también la activación de NF-κB y AP-1 (42). Por lo tanto, parece que los compuestos activos 3-15 (Fig. 1) (Fig. 2) de H. fasciculare pueden ser valiosos en estudios posteriores abordando su mecanismo citotóxico y para experimentos in vivo.
También los fasciculoles de H. fasciculare resultaron ser inhibidores de calmodulina (34); en particular los ácidos fascicúlicos A, B y C presentaron potente actividad antagonista de calmodulina (35).
El peróxido de ergosterol y el ergosterol tienen acción antiviral y antitumoral, pero además mostraron una actividad anticomplementaria muy fuerte en la ruta clásica, con valores de CI50 de 5,0 µM y 1,0 µM respectivamente (36). El sistema del complemento consiste en más de 20 proteínas séricas, que se activan mediante un mecanismo en cascada de la vía clásica o de la vía alternativa. La inflamación y las reacciones alérgicas desempeñan un rol importante en la defensa del huésped. El peróxido de ergosterol es un esterol de C28 que presenta actividad antimicrobiana, citotóxica, inmunosupresora y otras que han sido compiladas recientemente, junto con su mecanismo de acción (43).
Especies causantes del micetismo. División: Basidiomycota; Clase: Agaricomycetes; Orden: Agaricales; Familia: Strophariaceae: Hypholoma fasciculare (Huds.) P. Kumm. [Bas: Agaricus fascicularis Huds.; sin.: Naematoloma fasciculare (Huds.) P. Karst.] crece en la Argentina (provincias de Salta, Tucumán, Entre Ríos y Buenos Aires) (44)(45) y en Brasil (46); Hypholoma epixanthum (Fr.) Quél. [Bas.: Agaricus epixanthus Fr.; sin.: Naematoloma epixanthum (Fr.) P. Karst., N. epixanthum (Fr.) P. Karst., Geophila epixantha (Fries) Quél., Hypholoma radicosum J.E. Lange, N. radicosum (J.E. Lange) Konrad & Maubl., Psilocybe radicosum (J.E. Lange) Noordel.]; Hypholoma fasciculare var. mitis Raithelh. crece en la Argentina (provincias de Neuquén y Río Negro) (45)(46); Hypholoma lateritium (Schaeff.) P. Kumm. crece en la Argentina, Chile y Colombia (44)(46)(47); Hypholoma sublateritium (Schaeff.) Quél. [Bas: Agaricus sublateritius Schaeff.; sin.: Naematoloma sublateritium (Schaeff.) P. Karst.] crece en la Argentina (Buenos Aires y Tierra del Fuego) (45)(48); Hypholoma sublateritium var. aranoides Raithelh. crece en la Argentina (45)(46).
2.1.3. Fenol y azocompuestos en Agaricus xanthodermus
Agaricus xanthodermus es un hongo saprotrófico basidiomiceto responsable de un síndrome gastrointestinal. Está extendido en el Norte de África, Asia, Europa, América del Norte y América del Sur; se encuentra en la Argentina. Aparece en pequeños grupos en jardines y en los lugares donde hay hojas en descomposición, troncos, tocones, especialmente de coníferas. Su carne es blanca y su olor es fuerte y desagradable.
Sintomatología. Causa problemas digestivos. Los síntomas pueden incluir calambres abdominales, náuseas y diarrea; menos comúnmente dolor de cabeza, mareos, sudoración y somnolencia.
Toxinas. Los esporóforos frescos de A. xanthodermus produjeron 4,4'-dihidroxiazobenceno, fenol, p-quinol (=hidroquinona) y 4,4'-dihidroxibifenilo (Fig. 3) (49). Fue la primera vez que se encontró un compuesto azo endógenamente en la naturaleza, mientras que los restantes compuestos no se habían aislado previamente de hongos superiores. El fenol está presente en A. xanthodermus en una concentración suficientemente alta como para dar cuenta de su toxicidad (49). Todos estos compuestos son tóxicos y resulta llamativa su presencia en la naturaleza.

Figura 3. Estructuras químicas de 4,4'-dihidroxiazobenceno, 4,4'-dihidroxibifenilo, hidroquinona y fenol.
Especies causantes del micetismo. División: Basidiomycota; Clase: Agaricomycetes; Orden: Agaricales; Familia: Agaricaceae: Agaricus xanthodermus Genev. [Sin.: Pratella xanthoderma (Genev.) Gillet; Psalliota xanthoderma (Genev.) Richon & Roze; Fungus xanthodermus (Genev.) Kuntze] crece en la Argentina (provincias de Buenos Aires, Tucumán, Neuquén, Río Negro, Chubut, Santa Cruz y Tierra del Fuego); también en Buenos Aires crece Agaricus xanthodermus var. croceus Raithelh (48)(50).
2.1.4. Triterpenos tóxicos en especies del género Hebeloma
Las especies de Hebeloma que producen trastornos gastrointestinales por ingestión de sus carpóforos son: Hebeloma vinosophyllum, Hebeloma crustuliniforme, Hebeloma sinapizans, Hebeloma spoliatum, Hebeloma senescens y Hebeloma versipelle (51).
H. vinosophyllum es un hongo tóxico que crece en Asia, descripto por el micólogo japonés Tsuguo Hongo en 1965, que se registró por primera vez en el sudeste asiático en los bosques vietnamitas de Pinus kesiya en 2014 (52). Es un hongo amoniacal (desarrolla carpóforos en suelos con amoníaco agregado u otros materiales que contienen nitrógeno) que tiende a crecer en los cadáveres de animales.
H. sinapizans y H. crustuliniforme son hongos tóxicos que crecen principalmente en Europa y América del Norte (30), pero H. crustuliniforme crece también en América del Sur, como por ejemplo la Argentina (44)(53). Ambas especies crecen bajo árboles frondosos, bosques caducifolios y de coníferas. Estos hongos tienen un olor y sabor parecido al rábano, aunque H. crustuliniforme es más amargo.
Hebeloma velutipes es uno de los miembros más comunes y abundantes del género y H. sinapizans es una de sus especies más antiguas y más frecuentemente registradas. Usando grandes tamaños de muestras, varios loci y el análisis de tipos, recientemente se exploró la taxonomía, los límites de las especies europeas, la distribución y la clasificación infragenérica de estas dos especies y sus parientes (54). Los resultados filogenéticos son congruentes con respecto a la delimitación de especies, pero sugieren historias evolutivas conflictivas en su filogenia. Se evaluó el respaldo filogenético para las secciones y se describieron los caracteres morfológicos utilizados para distinguir las diez especies europeas reconocidas (54).
Toxinas. Se aislaron glicósidos triterpénicos de tipo cucurbitano, llamados hebevinósidos, como compuestos tóxicos de H. vinosophyllum (55-57) y triterpenos de tipo lanostano tóxicos de H. crustuliniforme, H. sinapizans y H. spoliatum (58).
Los hebevinósidos de los carpóforos de H. vinosophyllum (55)(56), fueron identificados como: 3β,7β,16β- trihidroxicucurbita-5,24-dien-3-O-(4-O-acetil)-β-D-xilopiranósido-16-O-(6-O-acetil)-β-D-glucopiranósido [Hidrhebg-3-O- ( 4 - A c ) X i l - 1 6 -O- ( 6 - A c ) G l u ] , 3β,7β,16β-trihidroxicucurbita-5,24-dien-3-O-β-D-xilopiranósido-16-O-(6-O-acetil)-β-D-glucopiranósido [Hidrhebg-3-O-Xil-16-O-(6-Ac)Glu], 3β,7β,16β-trihidroxi-cucurbita-5,24-dien-3-O-β-D-xilopiranósido-16-O-β-D-glucopiranósido [Hidrhebg-3-O-Xil-16-O-Glu], 3β,7β,16β-t rihidroxi-cucurbita-5,24-dien-3-O-β-D-xilopiranósido-16- O-(4,6-di-O-acetil)-β-D-glucopiranósido [Hidrhebg-3-O-Xil-16-O-(4,6-diAc)Glu], 3β,7β,16β-trihidroxicucurbita- 5,24-dien-3-O-(4-O-acetil)-β-D-xilopiranósido-16-O-(4,6- di-O-acetil)-β-D-glucopiranósido [Hidrhebg-3-O-(4-Ac) Xil-16-O-(4,6-diAc)Glu] y 3β,7β,16β-trihidroxicucurbita- 5,24-dien-3-O-β-D-xilopiranósido [Hidrhebg-3-O-Xil] (Fig. 4). La aglicona común para todos ellos es 3β,7β,16β- trihidroxicucurbita-5,24-dieno (hidroxihebevinogenina: Hidrhebg) (Fig. 4).
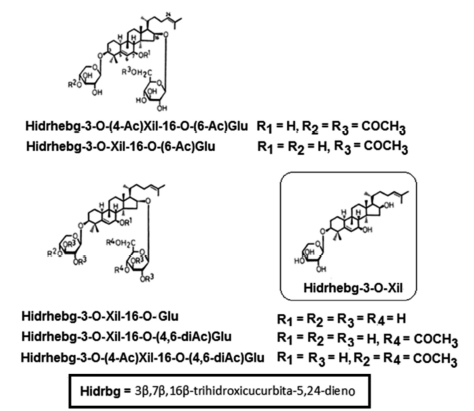
Figura 4. Estructuras químicas de los glicósidos triterpénicos con esqueleto tipo cucurbitano (hebevinósidos) de H. vinosophyllum.
Se demostró que la xilosa y la glucosa de los hebevinósidos pertenecen a la serie D. También se investigó la relación entre la estructura y la toxicidad de los hebevinósidos.
El compuesto tóxico 3β-acetil-2α-(3'-hidroxi-3'-metil) glutarilcrustulinol (ácido hebelómico) (Fig. 5) se aisló de los carpóforos de H. crustuliniforme y de H. sinapizans, el cual resultó citotóxico (59).

Figura 5. Estructuras químicas de triterpenos de tipo lanostano de H. crustuliniforme, H. sinapizans y H. spoliatum.
Se aisló además el ácido hebelómico con otros dos triterpenos de tipo lanostano de carpóforos de H. spoliatum, como los principios tóxicos fatales y se identificaron como: 3,21-diacetil-2-(3'-hidroxi-3'-metil)glutarilcrustulinol y 3β-acetil-2α-(3'-hidroxi-3'-metil)glutarilanhidrocrustulinol (Fig. 5) (58).
También se obtuvieron triterpenos tipo lanostano de carpóforos de H. senescens (60) y de H. versipelle (61).
Ensayos en ratones con los glicósidos triterpénicos de tipo cucurbitano. Por administración i.p. por separado en ratones de 100 mg/kg de los dos glicósidos: Hidrhebg-3-O-(4-Ac)Xil-16-O-(6-Ac)Glu e Hidrhebg-3-O-Xil-16-O-(6-Ac)Glu (Fig. 4), cuatro de cinco ratones murieron tras presentar parálisis. Se demostró, inyectando diferentes glicósidos, que la sustitución de un grupo hidroxilo de la posición 7β con un grupo metoxilo aumenta la toxicidad de los hebevinósidos (58).
Al administrar i.p. 100 mg/kg de Hidrhebg-3-O-Xil- 16-O-Glu (Fig. 4), los cuatro ratones murieron tras presentar parálisis, pero, con 50 mg/kg de este glicósido, sobrevivieron los cinco; el valor de la DL50 en ratones (i.p) fue de 66 mg/kg. Al administrar i.p. 200 mg/kg de uno de los glicósidos con una unidad 3-xilosilo, un 7-metoxilo y un 16-hidroxilo, sobrevivieron los cinco ratones (58).
Por otro lado, al administrar 100 mg/kg de un glicósido parcialmente hidrolizado y desacetilado, con un 3-hidroxilo, un 7-etoxilo y un 16-glucosilo, los tres ratones murieron tras presentar parálisis. Este resultado sugiere que la presencia de un 16-glucosilo en los hebevinósidos es indispensable para que haya toxicidad.
Ensayos en ratones con los triterpenos de tipo lanostano. Una dosis de 100 mg/kg de ácido hebelómico, 3,21-diacetil-2-(3'-hidroxi-3'-metil)glutarilcrustulinol y 3β-acetil-2α-(3'-hidroxi-3'-metil)glutarilanhidrocrustulinol (Fig. 5) en ratones por vía i.p. causó la muerte tras parálisis de las extremidades inferiores (58).
Mecanismo de acción. Los triterpenos de tipo lanostano causaron la relajación del intestino delgado de ratón, contraído por tratamiento con cloruro de acetilcolina o cloruro de bario in vitro. Estos lanosteroides parecen presentar un efecto de relajación similar a la papaverina (58).
Especies causantes del micetismo. División: Basidiomycota; Clase: Agaricomycetes; Orden: Agaricales; Familia: Cortinariaceae (Hymenigastraceae o Strophariaceae según la clasificación): H. crustuliniforme (Bull.) Quél. [Bas.: Agaricus crustuliniformis Bull.] crece en Europa, Australia, América del Norte y en América del Sur en la Argentina (Buenos Aires) (44)(53); H. vinosophyllum Hongo; H. spoliatum (Fr.) Gillet; H. sinapizans (Paulet) Gillet [Bas.: Hypophyllum sinapizans Paulet; sin.: Agaricus sinapizans (Paulet) Fr.; H. sinapizans (Paulet) Sacc.; H. sinapizans (Fr.) Sacc.] crece en Europa y América del Norte; H. versipelle (Fr.) Gillet crece en Europa (51)(54).
2.1.5. Iludinas en especies del género Omphalotus
Las especies del género cosmopolita Omphalotus son tóxicas y tienen como característica la bioluminiscencia de sus láminas que da una luminosidad verdosa o azul-verdosa en la oscuridad, debida a la enzima luciferasa, que actúa sobre el compuesto luciferina, y que conduce a la emisión de luz al igual que las luciérnagas (62). Los carpóforos carnosos tienen un tallo excéntrico que hace que el sombrero, de color anaranjado con diversos tintes según la especie, tenga forma de riñón o abanico.
Las especies de Omphalotus causan una putrefacción blanca al romper la lignina en sus árboles huéspedes (63); el hongo penetra en el duramen del árbol a través de una fisura en su corteza, ya sea por una rama que cae, daños causados por insectos o por daños mecánicos de la tala. Es decir que estos hongos desempeñan un papel importante en la descomposición de la madera y el reciclaje de los nutrientes en el suelo.
Varias especies de Omphalotus con propiedades bioluminiscentes similares se encuentran en todo el mundo, todas las cuales se presumen tóxicas. Las especies más conocidas son Omphalotus olearius (“seta de olivo”, “jack-o'- lantern”) que crece como parásito o saprobiótico en tocones en descomposición, en raíces enterradas o en la base de árboles de hoja caduca (olivos, robles o castañas) en áreas boscosas de Europa, especialmente en zonas de clima mediterráneo y en Sudáfrica (64) y Omphalotus illudens (“jack-o'-lantern”) que es una especie morfológicamente similar, pero filogenéticamente distinta (63), proveniente del este de América del Norte. También se destacan Omphalotus nidiformis (“hongo fantasma”) que es tanto saprotrófico como parasitario que crece en grupos superpuestos en una amplia variedad de árboles muertos o moribundos, en especial de eucaliptos, acacias, casuarinas y pinos, del sur de Australia y de Tasmania (65) y que se registró en India en 2012 creciendo bajo un tocón de cocotero (66), Omphalotus olivascens (“jack-o'-lantern occidental”) por análisis molecular muy relacionado con O. nidiformis (63) se encuentra en el sur y centro de California y en Méjico (67) y, por último, Omphalotus japonicus (“tsukiyotake”) (68-70) que crece en la madera de haya en descomposición (71) en las regiones montañosas de Japón, en Corea (72), China (73) y el este de Rusia, y está más estrechamente relacionado con un clado compuesto por O. nidiformis, O. olivascens y Omphalotus olearius según un estudio molecular de 2004 (63). O. japonicus fue responsable del 31,6% de los casos de intoxicación en Japón entre 1996 y 2005, más que cualquier otro hongo (74).
Todas estas especies son bioluminiscentes y tóxicas, y contienen las mismas toxinas (63).
Síntomatología. Aunque no son letales, O. olearius, O. illudens, O. olivascens y O. japonicus producen cólicos, vómitos y diarrea severos. Al consumir O. olearius predominan los vértigos, mareos, parestesias, lipotimias y relajación muscular. O. nidiformis induce vómitos, que generalmente se producen entre 30 minutos y dos horas después del consumo y duran varias horas; no se presenta diarrea y los pacientes se recuperan sin secuelas duraderas. El tratamiento más común es la terapia de fluidos (75)( 76).
Toxinas. Las iludinas son una familia de sesquiterpenos muy tóxicos con una subestructura de espirociclopropano, que presentan propiedades antibióticas y antitumorales (77); son responsables de los efectos nocivos causados por varias especies de Omphalotus cuando se consumen crudas o cocidas.
En 1963, las estructuras de iludinas M y S (Fig. 6) se asignaron totalmente (78). Los compuestos estructuralmente relacionados, iludina A y B (Fig. 6), se identificaron en 1991 como antibacterianos de Clitocybe illudens (=Omphalotus olearius) (79). También del cultivo de esta especie se aisló iludinina (Fig. 6), que es un alcaloide sesquiterpenoidal (80).

Figura 6. Estructuras químicas de iludina A, B, M y S, iludinina e irofulveno.
Iludina S junto con iludina M e iludosina, han sido identificados en O. illudens (77). Los carpóforos de O. japonicus (81-83) y de O. nidiformis (84) también contienen iludinas S y M, junto con iludinas F, G y H (85).
O. nidiformis mostró toxicidad marcada en cultivos celulares de cáncer gástrico (AGS), de colon (HT-29) y de mama independiente de estrógeno (MDA-MB-231) (86).
Las iludinas S y M presentan efectos antibacterianos y antifúngicos; son comunes al género Omphalotus y no se encuentran en ningún otro hongo basidiomiceto (84).
Los micelios de los cultivos sumergidos de O. olearius produjeron también dodecapéptidos cíclicos nematicidas, denominados onfalotinas A-I (Fig. 7), que no están presentes en los carpóforos (87-89). El compuesto principal es onfalotina A (Fig. 7), que es un aceite incoloro que permanece estable durante el aislamiento y el almacenamiento; contiene una alta proporción de L-aminoácidos metilados, como sarcosina (metilglicina), metilvalina y metilisoleucina. Estas onfalotinas presentan una actividad fuerte y selectiva contra nematodos fitopatógenos como Meloidogyne incognita, de importancia económica, con valores de DL50 entre 0,5 y 2,0 μg/mL (90) (onfalotina A presentó DL50=2 μg/mL; diez veces más potente que la ivermectina). Los efectos citotóxicos fueron comparativamente débiles (87-89).

Figura 7. Estructuras químicas de las onfalotinas A-I.
Mecanismo de acción. Las iludinas son agentes alquilantes del ADN (91). Dentro de las células, la iludina S (Fig. 6) reacciona con el ADN y lo daña, lo cual bloquea el proceso de transcripción. Este bloqueo sólo puede aliviarse mediante el sistema de reparación de ADN a través de la escisión de nucleótidos. La célula deja sin reparar el daño en las áreas no transcriptas del ADN.
El efecto citotóxico de la iludina es de interés para tratar algunos cánceres, pero es demasiado tóxica para su uso directo, por lo que primero debe modificarse químicamente. Irofulveno o 6-hidroximetilacilfulveno, un derivado semisintético de la iludina S desarrollado por la compañía MGI Pharma, está pasando por la fase II de ensayos clínicos como tratamiento potencial de tumores refractarios y recidivantes, incluidos los cánceres de ovario, próstata, hepatocelular, de mama, pulmón y colon (92-94), pero debido a sus efectos secundarios hematológicos, aún debe ser aprobado para uso clínico.
Especies causantes del micetismo. División: Basidiomycota; Clase: Agaricomycetes; Orden: Agaricales; Familia: Marasmiaceae (Omphalotaceae en desuso): Omphalotus olearius (DC.) Sing. [Bas: Agaricus olearius DC.; sin.: A. olearius subsp. phosphoreus Battarra ex Pers., Dryophila phosphorea (Battarra ex Pers.) Quél., Clitocybe olearia (DC.) Maire, Clitocybe phosphorea (Battarra ex Pers.) Bohus] (95); Omphalotus illudens (Schwein.) Bresinsky & Besl [Bas.: Agaricus illudens Schwein.; sin.: Panus illudens (Schwein.) Fr., Clitocybe illudens (Schwein.) Sacc., Lentinus illudens (Schwein.) Henn., Pocillaria illudens (Schwein.) Kuntze, Monodelphus illudens (Schwein.) Earle, Omphalotus olearius var. illudens (Schwein.) A. Ortega & Esteve-Rav. (96)]; Omphalotus nidiformis (Berk.) O.K. Mill.; Omphalotus olivascens H.E.Bigelow, O.K.Mill. & Thiers; Omphalotus japonicus (Kawam.) Kirchm. & OK Mill. [Bas.: Pleurotus japonicus Kawam.; sin.: Armillaria japonica (Kawam.) S.Imai; Pleurotus harmandii Har. & Pat.; Lampteromyces japonicus (Kawam.) Singer; Omphalotus guepiniformis (Berk.) Neda].
2.1.6. Sesquiterpenos cíclicos en especies de los géneros Lactarius y Russula
Una característica general de las especies del género Lactarius (“níscalos”) es que los carpóforos contienen un látex o jugo acuoso lechoso que expulsan al cortarlos cuando están frescos y que cambia de color en contacto con el aire. Este látex fluye por un sistema de tubos lacticíferos que se ramifican por todos los tejidos del esporóforo. El color y el sabor de este látex varían entre las diferentes especies, y son de gran importancia taxonómica.
El látex contiene una variedad de sesquiterpenos que a mayor número de insaturaciones químicas darán lugar a mayor intensidad en el color. Se han aislado sesquiterpenos con esqueletos de lactarano, seco-lactarano (abierto el anillo de siete miembros) y marasmano de los carpóforos de especies de los géneros Lactarius y Russula. Estos sesquiterpenos desempeñan una función de defensa química, contra los depredadores y es por eso que presentan fuerte actividad antimicrobiana, mutagénica y antialimentaria o antifeedant, pero además son tóxicos para los humanos.
Sintomatología. Los síntomas por ingestión de Lactarius helvus aparecen dentro de los treinta minutos posteriores al consumo, con náuseas y vómitos acompañados de vértigo y escalofríos. En octubre de 1949, 418 personas se intoxicaron con L. helvus cerca de Leipzig en el este de Alemania (97).
Las toxinas se destruyen mediante ebullición profunda; L. helvus se usa en pequeñas cantidades como especia después del secado.
Lactarius torminosus se caracteriza por su sabor intensamente picante cuando se consume crudo y, en exceso, puede provocar ampollas en la lengua (98). Su toxicidad se limita a un síndrome gastrointestinal debido a la presencia en el hongo de sustancias no absorbibles, pero irritantes para el tubo digestivo. Los síntomas que se experimentan tras consumir carpóforos crudos son náuseas, diarreas severas, vómitos y dolor abdominal que aparecen entre 15 minutos y tres horas después de la ingestión. Esto puede conducir a deshidratación, espasmo muscular y colapso circulatorio. Aún así, ocupan una parte muy baja en la escala de peligrosidad; de hecho, la gastroenteritis generalmente se resuelve sin tratamiento en un par de días (99). Esta especie se consume en Finlandia, Rusia y otros países de Europa septentrional y oriental, tras una cocción inicial y varios días de remojo en salmuera.
Lactarius necator ha causado varios casos de gastroenteritis, probablemente debido a insuficiente pretratamiento, como salazón y/o sancochado (10)(64)(100).
Se considera que Lactarius rufus es un hongo comestible, después del tratamiento previo apropiado. Su consumo ha dado lugar a varios casos de gastroenteritis, probablemente debido a un pretratamiento no adecuado (10)(64)(100).
Toxinas. Son sesquiterpenos con diferente esqueleto y distinta concentración, y también según los procedimientos y el solvente de extracción; así, al usar etanol se obtuvieron éteres etílicos como compuestos no originales de los carpóforos.
Bajo condiciones bien controladas en las etapas de extracción y aislamiento, se obtuvo estearoilvelutinal (éster de ácido esteárico del sesquiterpeno velutinal muy inestable) (Fig. 8) como único sesquiterpeno de carpóforos de Lactarius vellereus (101). Cuando el hongo se lesiona o es atacado por insectos o caracoles, el estearoilvelutinal se convierte rápidamente, posiblemente por procesos enzimáticos, en los dos dialdehídos sesquiterpénicos tóxicos: (+)-isovelleral con esqueleto de marasmano, y velleral (Fig. 8) con esqueleto de lactarano. Estos dos compuestos tóxicos son luego reducidos gradualmente mediante el carpóforo a compuestos menos tóxicos como isovellerol y vellerol (Fig. 8) y, eventualmente, a vellerdiol (diol de velleral) (Fig. 8) (101). Estos compuestos constituyen el sistema de defensa química que se mencionara.

Figura 8. Estructuras químicas de estearoilvelutinal, velutinal, (+)-isovelleral, velleral, isovellerol, vellerol y vellerdiol.
La toxicidad de isovelleral y de velleral es similar y es más de 10 veces mayor que la de estearoilvelutinal, isovellerol, vellerol y vellerdiol. La marcada disminución de la toxicidad de isovelleral y velleral por reducción química del grupo aldehído de C-13 indica la importancia de este último para la actividad biológica.
El hecho de que los sesquiterpenos, no ésteres, se formen cuando el hongo se lesiona y, su posterior reducción enzimática a otros compuestos mucho menos tóxicos, tendría el efecto de evitar que el hongo entre en contacto y sea atacado por estos compuestos antifúngicos de defensa.
Velleral e isovelleral ya se habían aislado de carpóforos de L. torminosus, cuyo sabor acre y picante, así como la toxicidad de los carpóforos se debe a la formación de estos dialdehídos insaturados. Velleral e isovelleral mostraron actividad antimicrobiana y antifúngica, isovellerol y vellerol mostraron actividad parcial y, estearoilvelutinal y vellerdiol, ninguna actividad (102).
Isovelleral también resultó ser un potente mutágeno de acción directa en el test de Ames, así como un antifeedant para insectos y un potente antifeedant para topos. Isovellerol también resultó ser mutagénico, pero unas diez veces menos que isovelleral.
Los derivados de lactarano, marasmano y/o seco-lactarano como (+)-isovelleral, velleral, isovellerol y vellerol no son exclusivos de L. vellereus, sino que se encuentran en muchas especies de Lactarius y Russula; como por ejemplo, especies tóxicas (pungentes) del género Lactarius: Lactarius aurantiacus, Lactarius chrysorrheus, L. helvus, L. necator, L. rufus, Lactarius scrobiculatus, Lactarius subdulcis, Lactarius trivialis, Lactarius turpis, Lactarius utilis y Lactarius volemus (103)(104). L. necator, L. rufus y L. trivialis contienen estearoilvelutinal y 6-cetoestearoilvelutinal (105). L. trivialis también contiene lactarorrufina A, blenina C y el furanolactarano furandiol (105).
Cada especie parece exhibir su propio patrón característico, con los siguientes parámetros variables: a) los ácidos grasos en los ésteres de velutinal; b) la cantidad de velutinal originalmente presente en el hongo; c) la cantidad relativa formada de isovelleral y velleral; d) la tasa de conversiones enzimáticas; e) la formación de otros sesquiterpenos, supuestamente derivados de isovelleral y velleral.
Russula queletii, no comestible, por ejemplo, que contiene menos éster de velutinal que L. vellereus, produce velleral y vellerol, pero no isovelleral e isovellerol, y además, también produce otros sesquiterpenos.
Los micólogos clasifican a los hongos según su pungencia y, como isovelleral y velleral tienen un sabor muy pungente, se debería esperar que las especies pungentes de Russulaceae produjeran isovelleral y/o velleral, a diferencia de las no pungentes. A veces el primer sabor confunde, como ocurre con Lactarius quietus que se describe como de sabor suave o ligeramente amargo, pero si se mantiene en la boca durante al menos un minuto, aparece un sabor fuerte debido a la presencia de isovelleral, acompañado por isovellerol; L. quietus no produce velleral ni vellerol.
Del extracto etanólico de los carpóforos de Lactarius blennius, que es un hongo no comestible que crece en los bosques de hayas en los Apeninos italianos, se aislaron varios sesquiterpenos, entre ellos las lactonas blenina A y su derivado 8-O-acetilado, blenina B, los furosesquiterpenos 3,8-dihidroxilactariusfurano o 3,8-dihidroxifurolactarano (o furandiol), 3-hidroxi-8-O-acetil-lactariusfurano, 8-hidroxifurolactar-2-eno, y las lactonas sesquiterpénicas lactarorrufina A y blenina C (Fig. 9) (105).

Figura 9. Estructuras químicas de blenina A y su derivado 8-O-acetilado, blenina B, 3,8-dihidroxilactariusfurano (o furandiol), 3-hidroxi-8-O-acetil-lactariusfurano, 8-hidroxifurolactar-2-eno, lactarorrufina A y blenina C.
Varios de ellos se han encontrado previamente en diferentes especies de hongos, así por ejemplo, 3,8-dihidroxifurolactarano y su derivado 8-O-acetilado, 3-hidroxi-8-acetoxilactariusfurano o 3-hidroxi-8-acetoxifurolactarano se encuentran también en L. scrobiculatus y L. necator; blenina C se aisló de L. blennius y de L. scrobiculatus; lactarorrufina A, furandiol y metoxi-furanalcohol se aislaron de Russula sardonia (10) y de Russula emetica (106).
Todos estos compuestos tienen el esqueleto de lactarano inusual y característico que parece exclusivo de los sesquiterpenos de las especies tóxicas y no comestibles de los géneros Lactarius y Russula.
Otros compuestos presentes en los carpóforos. En L. helvus el compuesto que da lugar al aroma y olor distintivo de esta especie es el sotolón (4,5-dimetil-3-hidroxi-2,5-dihidrofuran-2-ona) (Fig. 10) (107), que da también el olor característico a las semillas de hinojo, al apio de monte, al sake añejado y al tabaco tostado. Recientemente se realizó una actualización sobre los compuestos volátiles de hongos (108).

Figura 10. Estructura química de sotolón.
Los carpóforos de L. trivialis y L. rufus contienen muscarina en unos 2 mg/kg de peso fresco, que corresponden a L-(+)-muscarina (~ 40%) y sus estereoisómeros (~60%). Estas cantidades bajas no son de preocupación toxicológica para los niveles normales de consumo de estos hongos (64).
Los carpóforos frescos de L. turpis y L. necator contienen el compuesto mutagénico necatorina (Fig. 11), a concentraciones de 3 a 20 mg/kg, por lo que no se recomienda comerlos. La ebullición reduce la concentración de este compuesto, pero no lo elimina de manera efectiva (109)(110). Necatorina se aisló como cristales rojos brillantes y demostró ser muy mutagénica en el test de Ames en tres cepas de Salmonella.

Figura 11. Estructuras químicas de necatorina, necatorona, 4,4'-binecatorona, 10-desoxi-4,4'-binecatorona y 10,10'-didesoxi-4,4'-binecatorona.
La ebullición/escaldado destruye aproximadamente la mitad de la necatorina, y deja alrededor del 25% del contenido original en el hongo y ~25% en el agua (109)(110). La destrucción de necatorina pura por ebullición fue más efectiva a pH ácido (pH=5,0).
El compuesto mutagénico necatorona (5,10-dihidroxi-6H-pirido[4,3,2-kl]-acridin-6-ona) (Fig. 11) es un pigmento alcaloidal aislado de los carpóforos de L. turpis o L. necator y Lactarius atroviridis (109)(110), junto con dímeros de este alcaloide, como 4,4′-binecatorona (sólido cristalino pardo rojizo), 10-desoxi-4,4′-binecatorona (sólido cristalino pardo rojizo) y 10,10′-didesoxi-4,4′- binecatorona (sólido cristalino anaranjado) (Fig. 11), que también son mutagénicos (111).
Varias especies de Lactarius presentaron un poliisopreno correspondiente a un contenido de caucho líquido de 0,1% a 7% en peso seco (112). Se encontró que el caucho de L. volemus, L. chrysorrheus y Lactarius hygrophoroides es un homólogo del poliprenol que está compuesto de un grupo dimetilalilo, dos unidades de trans-isopreno, 160-300 unidades de cis-isopreno y un grupo terminal hidroxilo o éster (112)(113). El número de grupos terminales y de unidades trans disminuye durante el envejecimiento de los esporóforos. El caucho de Lactarius piperatus, L. vellereus y Lactarius subpiperatus es cis-poliisopreno, con cantidades muy pequeñas de grupos terminales y unidades trans (112)(113). Dada la gran cantidad de caucho natural que poseen se considera que los carpóforos de L. volemus pueden usarse para producir caucho. Es probable que el alto contenido de cis-poliisopreno pueda influir negativamente en la digestibilidad de este hongo.
L. rufus contiene una lectina con seis subunidades idénticas de ~17 kDa; el 85% de su actividad se pierde calentando el hongo a 65 °C (114). Se sabe que algunas lectinas en alimentos causan malestar gastrointestinal, especialmente si la comida no fue suficientemente tratada térmicamente.
Antes del consumo, estos hongos requieren ser pretratados (salado/tratamiento con calor) para destruir las sustancias acres y se debe descartar el agua del pretratamiento.
Los carpóforos de L. volemus contienen un compuesto con un esqueleto de hepta-nor-ergostano llamado volemolida (Fig. 12), derivado del ergosterol, que es el esterol común en hongos, que puede tener aplicación en la quimiotaxonomía fúngica (115) y volemitol (D-glicero-D-manoheptitol) (Fig. 12), un glicitol de siete carbonos, que se encuentra libre en muchas especies de plantas y de algas pardas (116). También se identificaron siete esteroles tipo ergostano, muy oxigenados, similares a los esteroles del coral blando marino y las esponjas marinas, que son raros en los hongos (117).

Figura 12. Estructuras químicas de volemolida y volemitol.
Las especies comestibles Lactarius deliciosus y Lactarius deterrimus contienen además de los sesquiterpenos con esqueleto de azuleno, compuestos indólicos y lectinas. Así, se aislaron e identificaron 5-hidroxitriptofano, serotonina y melatonina, así como sulfato de kinurenina, que es un metabolito de triptofano, de los carpóforos de L. deliciosus (118); se demostró que por ebullición de este hongo 5-hidroxitriptofano y serotonina se destruyeron completamente, mientras que se formaron triptofano, 5-metiltriptofano y triptamina. De manera similar, triptofano, triptamina y melatonina se aislaron de L. deterrimus (119). Algunos de estos compuestos pueden actuar como hormonas tisulares y neurotransmisores y altas ingestas pueden dañar al sistema nervioso (120).
L. deliciosus posee una lectina dimérica de ~37 kDa (subunidades de ~19 y 18 kDa) (121), mientras que la lectina de L. deterrimus es de ~31 kDa con dos subunidades idénticas (122).
Mecanismo de acción de las toxinas. Los sesquiterpenos descriptos actúan como irritantes en el tracto gastrointestinal de los pacientes. Se necesita ahondar más sobre la manera en que actúan.
Diferencias químicas entre hongos comestibles y tóxicos del género Lactarius.
Se encontró que los carpóforos intactos de los hongos comestibles, L. deliciosus y L. deterrimus, contienen sólo un sesquiterpeno en forma de dos ésteres de ácidos grasos: 15-O-estearoiloxidihidroazuleno (componente principal) y 15-O-linoleiloxidihidroazuleno (componente minoritario) (Fig. 13) (123). Al igual que en las especies tóxicas (pungentes) de Lactarius, si bien menos rápidamente, estos ésteres originales se convierten, enzimáticamente, en otros compuestos cuando se lesionan (se cortan) los carpóforos. No se detectaron diferencias, ni cuali ni cuantitativas, entre L. deliciosus y L. deterrimus en este aspecto. Las conversiones de los sesquiterpenos pueden ser parte de un sistema de defensa química, como se ha mencionado para otras especies (123).

Figura 13. Estructuras químicas de lactaroviolina, 15-hidroxidihidroazuleno,15-O-estearoiloxidihidroazuleno, 15-O-linoleiloxidihidroazuleno, lactarazuleno, lactarofulveno, 11,12-dihidrolactaroviolina, delicial, deterrol y estearoildeterrol.
Lactaroviolina, 15-hidroxidihidroazuleno, su éster 15-estearoiloxidihidroazuleno, lactarazuleno y deterrol (Fig. 13) han sido aislados de especímenes europeos de L. deliciosus. En cambio, lactarofulveno se aisló de especímenes californianos de L. deliciosus (123), mientras que 11,12-dihidrolactaroviolina (Fig. 13) se aisló de L. deterrimus de la India (124). Una explicación de esta aparente diferencia entre especímenes de diferentes continentes puede ser la existencia de subespecies de L. deliciosus y L. deterrimus.
El éster de ácido esteárico de deterrol (estearoildeterrol) (Fig. 13) se aisló previamente de carpóforos de Lactarius indigo. Este éster parece ser el principal sesquiterpenoide presente originalmente en los carpóforos de esta especie y es responsable del color azul profundo de su látex.
La inestabilidad de los derivados de azuleno, como el alcohol 15-hidroxidihidroazuleno y el delicial, favoreció la formación, durante el manejo y el almacenamiento de los extractos, de pequeñas cantidades de otros compuestos que no estaban originalmente en los carpóforos. Uno de esos compuestos resultó ser un dímero de dos sesquiterpenos guayanos. En cambio, la lactaroviolina y el deterrol, presentan cierta estabilidad; se forman lentamente y se acumulan en el tejido de los hongos durante las primeras horas después de la lesión (123).
L. deliciosus (Fr.) Gray es una especie micorrícica comestible que crece bajo pinos; en particular en la Argentina este hongo está asociado a plantaciones de Pinus elliottii introducidas en el nordeste argentino (provincia de Misiones) (125).
El látex de los carpóforos de L. deliciosus y L. deterrimus primero es de color anaranjado, pero lentamente (en minutos) se oscurece y finalmente se vuelve verde; esta característica hace que muchos dejen de lado estas exquisitas especies. Se demostró que estos colores en L. deliciosus y L. deterrimus se deben a los sesquiterpenos del tipo guayano, que por tener dobles enlaces conjugados, dan colores intensos en la zona visible. El color verde que el látex asume con el tiempo se debe a la formación de compuestos color violeta y azul (lactaroviolina y deterrol) y su mezcla con los compuestos amarillos (15-hidroxidihidroazuleno, 15-estearoiloxidihidroazuleno, 15-linoleiloxidihidroazuleno y delicial) ya presentes o también formados (123).
Los sesquiterpenos de las especies L. deliciosus y L. deterrimus tienen únicamente esqueleto de guayano, que no se encuentra en las especies tóxicas (pungentes) de Lactarius.
Especies causantes del micetismo. División: Basidiomycota; Clase: Agaricomycetes; Orden: Russulales; Familia: Russulaceae: Lactarius alnicola A.H. Sm. crece en micorrizas con coníferas y especies de hoja caduca como roble y aliso, en los bosques nativos del oeste de Estados Unidos, Méjico (Baja California), India y en las plantaciones patagónicas de Pinus ponderosa y abeto de Douglas (especies del género Pseudotsuga) en la Argentina (126); L.torminosus (Schaeff.) Gray [Bas.: Agaricus torminosus Schaeff.; sin.: Galorrheus torminosus (Schaeff.) P. Kumm.; L. torminosus f. torminosus (Schaeff.) Gray; L. torminosus var. sublateritius Kühner & Romagn.; Lactifluus torminosus (Schaeff.) Kuntze] crece en Europa formando micorrizas con abedules; Lactarius helvus (Fr.) Fr. [Bas.: Agaricus helvus Fr.; sin.: Galorrheus helvus (Fr) P. Kumm., Lactifluus helvus (Fr.) Kuntze]; L. rufus (Scop.) Fr. [Bas.: Agaricus rufus Scop.] crece en zonas templadas de Europa, e incluye zonas subárticas/ subalpinas de los países escandinavos, y en América del Norte (norte de California) forma micorrizas con pinos, abedules o abetos (103); Lactarius trivialis (Fr.) Fr. [Bas: Agaricus trivialis Fr.] se encuentra en América del Norte (EE. UU., Canadá, México) y Europa (Gran Bretaña e Irlanda, países escandinavos, algunas regiones de Alemania, Prusia Oriental y en Rusia blanca y noroccidental, pero se vuelve mucho más raro en el sur y suroeste de Europa); Lactarius turpis Fr. [Sin: Agaricus turpis Weinm., Galorrheus turpis (Weinm.) P. Kumm., Lactifluus turpis (Weinm.) Kuntze] crece en Europa y Siberia, y se ha introducido en Australia y Nueva Zelanda. Por lo general, pero no siempre, por un problema de nomenclatura, se consideran sinónimos de la misma especie: Lactarius turpis Fr., L. necator (Bull.: Fr.) Karsten y L. plumbeus (Bull.: Fr.) S. F. Gray; en Europa occidental se clasifica este hongo como no comestible; pero en países escandinavos, Siberia y el este de Europa se consumen después de hervirlos y se conservan en sal; Lactarius necator (Bull.) Pers. [Sin.: L. necator (Bull.: Fr.) Karsten] no es comestible, forma micorrizas con pícea (Picea) y abedules (Betula) en zonas templadas-hemiboreales en los países nórdicos, excepto en Islandia, donde es muy raro (103); Lactarius utilis (Weinm.) Fr. [Bas.: Agaricus utilis Weinm.; sin.: Lactifluus utilis (Weinm.) Kuntze] está muy relacionado con L. trivialis (hay diferencias en el píleo y en el látex) y se distribuye en la zona boreal del norte de Europa y forma micorrizas con píceas o abedules; Lactarius vellereus (Fr.) Fr. [Bas.: Agaricus vellereus Fr.; sin.: Agaricus vellereus var. vellereus Fr., Galorrheus vellereus (Fr.) P. Kumm., Lactarius albivellus Romagn., L. vellereus var. vellereus (Fr.) Fr., L. vellereus var. velutinus (Bertill.) Bataille, L. velutinus Bertill., Lactifluus vellereus (Fr.) Kuntze]; Lactarius volemus (Fr.) Kuntze [Bas.: Agaricus volemus Fr.; sin: L. volemus (Fr.) Fr., Agaricus lactifluus L., Agaricus oedematopus Scop., L. oedematopus (Scop.) Fr., Galorrheus volemus (Fr.) P. Kumm, Lactarius lactifluus (L.) Quél., Lactifluus oedematopus (Scop.) Kuntze, L. wangii H.A. Wen & J.Z. Ying] se considera un hongo comestible si bien contiene cis-poliisoprenos; forma micorrizas en bosques caducifolios y de coníferas; Russula emetica (Schaeff.) Pers. [Bas.: Agaricus emeticus Schaeff.; sin.: A. russula Scop., A. ruber (Lam.) DC., A. linnaei var. emeticus (Schaeff.) Fr., Amanita rubra Lam., Russula rubra (Lam.) Fr., Melanoleuca russula (Scop.) Murrill]; Russula queletii Fr.; Russula sardonia Fr.
2.1.7. Vinilglicina en Entoloma sinuatum y otras especies de Entoloma
Esta intoxicación es provocada por varios miembros del gran género Entoloma, que contiene alrededor de 1.500 especies. Muchas especies de Entoloma son tóxicas, como Entoloma sinuatum (9) en Europa y Entoloma rhodopolium (127-129) en Japón, si bien algunas otras son comestibles, como Entoloma abortivum y Entoloma sarcopum.
E. sinuatum posee el mayor número de registros de intoxicaciones por su consumo en Europa y América del Norte (30)(130)(131). Algunas guías sobre hongos se refieren a esta especie por sus nombres científicos más antiguos de Entoloma lividum o Rhodophyllus sinuatus. También Entoloma nidorosum y Entoloma vernum producen este tipo de micetismo.
E. rhodopolium es un hongo tóxico, pero no letal, de Europa y Asia. Es uno de los tres hongos más comúnmente implicados en intoxicaciones en Japón (132). Su carne es clara y endeble, con olor muy desagradable parecido al del amoníaco o la lejía (lavandina). Los estudios filogenéticos moleculares, realizados en 2017, han mostrado que los clados-I, -II y -III de E. rhodopolium japoneses son distintos de los de E. rhodopolium europeo, E. aff. rhodopolium de América del Norte, E. nidorosum y otras especies conocidas de Entoloma (133). El resultado indica que los clados japoneses de E. rhodopolium son especies nuevas, que denominaron Entoloma lacus, Entoloma subrhodopolium y Entoloma pseudorhodopolium
respectivamente. Esto indica que hay una diversidad morfológica dentro de una especie (133).
Período de latencia. Los síntomas aparecen de 30 minutos a 2 horas después del consumo y duran hasta 48 horas. En cambio, en el caso de E. sinuatum, el período de latencia es algo más largo, ya que los síntomas aparecen después de unas 4 horas.
Sintomatología. E. sinuatum es muy tóxico; causa intoxicación con náuseas, vómitos, gastroenteritis y hepatopatía leve, dolor de cabeza y diarrea severa que puede conducir a la deshidratación, un conjunto de síntomas denominado “síndrome resinoideo severo”, así como enrojecimientos en algunos lugares del cuerpo.
La pérdida importante de líquidos va generalmente acompañada por pérdida de electrolitos (magnesio, potasio, calcio), que causa calambres musculares y, a veces, trastornos circulatorios potencialmente mortales. Además, la suposición de una amenaza de vida a menudo conduce a la ansiedad con sudoración y sensación de frío. Generalmente se resuelven después de 1 a 2 días, sin secuelas (75).
En resumen, los trastornos gastrointestinales pueden ser graves, en ciertos casos mortales, a veces con un daño leve parenquimatoso hepático. También puede producirse toxicidad hepática aguda y síntomas psiquiátricos, como alteración del estado de ánimo o delirio. El delirio y la depresión son secuelas poco comunes. En raras ocasiones, los síntomas de depresión pueden durar meses (10).
La intoxicación generalmente pasa después de 5 a 6 días. Sin embargo, puede ser fatal en personas debilitadas, en adultos mayores y niños.
En cambio, E. nidorosum es una especie tóxica, cuyo olor muy desagradable desalienta su consumo y por lo tanto limita el número de pacientes intoxicados. Además, la intoxicación debida a la ingestión de E. nidorosum es más leve que la provocada por E. sinuatum; sólo causa náuseas, vómitos y diarreas, con una evolución sin casos fatales y la recuperación se produce en unas 12 horas.
La intoxicación por ingestión de E. vernum presenta como síntomas dolores abdominales con diarrea profusa, no hay casos fatales y los pacientes se recuperan, sin secuelas.
Los síntomas provocados por E. rhodopolium son predominantemente de naturaleza gastrointestinal (diarrea, vómitos, dolor gástrico, dolor de cabeza), pero se han descripto también signos de tetania (134), pérdida de equilibrio y astenia (135) y sudores fríos. Si bien E. rhodopolium contiene muscarina, muscaridina y colina, estas sustancias deben estar en cantidades pequeñas pues no se observaron síntomas de síndrome muscarínico.
Casos de intoxicación. E. sinuatum ha sido citado como responsable del 10% de todos los micetismos en Europa (136). Así, por ejemplo, 70 personas necesitaron tratamiento hospitalario sólo en Ginebra en 1983 (130), y el hongo representó 33 de 145 casos de micetismos en un período de cinco años en un solo hospital en Parma (131).
En 1974 se describió la intoxicación de una pareja debida a ingestión de E. vernum, que tuvo que ser internada (137). De 24 intoxicaciones debidas a la ingestión de E. rhodopolium y/o E. nidorosum, 21 pacientes sufrieron una intoxicación colectiva por E. rhodopolium (134). Posteriormente se registró una intoxicación colectiva de 23 personas en Japón (135), otros 5 casos y dos intoxicaciones en Francia en 1982. E. rhodopolium es una de las tres principales especies responsables de los 2.096 incidentes de micetismos que involucraron a 10.924 pacientes de 1959 a 1988 en Japón (138), que correspondieron a tasas respectivas de pacientes totales y muertes de 90,3% y 10,7% para intoxicaciones con síntomas tipo cólera, así como 90,2% y 0,2% para los de irritación gastrointestinal (138).
Toxinas. Se considera que la toxina es la vinilglicina (ácido 2-amino-3-butenoico) (Fig. 14); se aisló de Entoloma nidorosum (10) y de E. lividoalbum (139). Se han registrado casos de intoxicación (132). Si bien este aminoácido con propiedades tóxicas ha sido aislado, nada prueba que sea el único responsable de los trastornos.
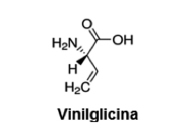
Figura 14. Estructura química de vinilglicina.
También se han encontrado alcaloides en los carpóforos de E. sinuatum (140).
Un estudio de oligoelementos en hongos de la región oriental del Mar Negro en Turquía encontró que E. sinuatum tenía los niveles registrados más altos de cobre (64,8±5,9 μg/g de carpóforos secos; insuficiente para ser tóxico) y de cinc (198 μg/g) (141). Los sombreros y los tallos ensayados en un área con altos niveles de mercurio en el sudeste de Polonia demostraron que éste bioacumulaba niveles mucho más altos de mercurio que otros hongos. Este elemento también se encontró en niveles altos en el sustrato rico en humus (142). E. sinuatum también acumula compuestos que contienen arsénico. De ~ 40 μg de arsénico presentes por gramo de tejido fresco de hongo, ~8% era arsenito y el otro 92% era arseniato (143).
Además, el hongo contiene lectinas, que específicamente producen la aglutinación de los hematíes del grupo sanguíneo A. También actúa como insecticida para las moscas de la fruta.
En 1985 se aislaron colina, muscarina y muscaridina (Fig. 15) de extractos alcohólicos del carpóforo de E. rhodopolium (144). En 1987 se demostró que una fracción de un extracto acuoso responsable de hemorragia intestinal, diarrea y muerte en ratones, tenía propiedades hemolíticas (127). Luego de ser purificada en 1988 y 1990 resultó ser una hemolisina de 40 kDa, relativamente termolábil (128)(129), cuyo rol en la intoxicación humana es desconocido. La actividad hemolítica de la hemolisina purificada se destruyó por calentamiento a 60 °C durante 10 minutos y se redujo parcialmente por pepsina, papaína y 2-mercaptoetanol. La hemolisina fue inestable por debajo de pH 7,0, pero estable a pH 8,0. El pH óptimo para la hemólisis fue 6,0. La hemólisis no se produjo por debajo de 4 °C a pesar de que la hemolisina se uniera al eritrocito. Los eritrocitos de ratón, pollo, rata, caballo y humanos fueron sensibles en este orden, pero los eritrocitos de oveja y vaca no fueron lisados por la toxina (129).

Figura 15. Estructura química de muscarina, colina y muscaridina.
Especies causantes del micetismo. División: Basidiomycota; Clase: Agaricomycetes; Orden: Agaricales; Familia: Entolomataceae: especies del género Entoloma (53). Entoloma sinuatum (Bull.) P. Kumm. [Bas.: Agaricus lividus Bull.; sin.: Agaricus sinuatus Pers., Entoloma sinuatum (Pers.) P. Kumm., E. eulividum Noordel., E. lividum (Bull.) Quél., Rhodophyllus lividus (Bull.) Quél., R. sinuatus Singer, R. sinuatus (Bull.) Quél.] forma micorrizas con árboles de hoja caduca principalmente haya, roble y, con menor frecuencia, abedul, en suelos arcillosos o calcáreos o en parques cercanos, a veces en forma de anillos de hadas, en Europa (desde Francia en el oeste hasta Estonia y Bielorrusia en el este, Alemania en el centro y sur hasta España, Italia, Bulgaria y Rumania y al norte Dinamarca y el sur de Noruega), América del Norte (Canadá, EE. UU.) y Asia (Turquía, Asia Menor, China y Japón); son saprotróficos, si bien esta especie forma una relación ectomicorrízica con el sauce (Salix) (145). Entoloma nidorosum (Fr.) Quél. [Bas.: Agaricus nidorosus Fr.; sin.: Rhodophyllus nidorosus (Fr.) Quél., Entoloma rhodopolium f. nidorosum (Fr.) Noordel., E. rhodopolium var. nidorosum (Fr.) Krieglst.]; Entoloma niphoides Noordel.; Entoloma rhodopolium (Fr.) P. Kumm. [Bas.: Agaricus rhodopolius Fr.; sin.: Entoloma nidorosum (Fr.) Quél., variedades de E. nidorosum, variedades de E. rhodopolium, Agaricus nidorosus Fr., variedades de A. rhodopolius Fr., Rhodophyllus nidorosus (Fr.) Quél., R. rhodopolius (Fr.) Quél., variedades de R. rhodopolius] se encuentra en bosques de frondosas (raramente aparece bajo coníferas) en Europa y Asia (Japón); Entoloma vernum Lundell [Sin.: Rhodophyllus cucullatus J. Favre, Nolanea cucullata (J. Favre) P. D. Orton, Nolanea verna (S. Lundell) Kotl. & Pouzar] (146).
2.1.8. Toxina hemolítica rubescenslisina en el hongo comestible Amanita rubescens
Amanita rubescens es una especie muy común en toda Europa y expandida a los otros continentes, si bien se cree que la encontrada en EE. UU. es diferente a la europea.
Se presenta en variantes de color de sombrero (rojizo, marrón y plata metálica brillante) y también puede asumir distintas apariencias, por ejemplo, hay una variante blanca pura. También varía el tamaño del sombrero; se han encontrado ejemplares de A. rubescens completamente maduros con sombreros de no más de 2,5 cm de diámetro, mientras que otros alcanzan casi diez veces ese tamaño. Tanto su nombre científico como los comunes derivan de la propensión de su carne a volverse rosada al cortarla o golpearla (103).
A. rubescens generalmente se considera comestible si está bien cocida; pero, dado que se sabe que contiene una toxina hemolítica en estado crudo que causa anemia y que, aún con un shock térmico para destruirla, los hongos resultan indigestos, la mayoría de las personas optan por no consumirlos.
Además, debe cuidarse su identificación porque se presta a confusión con Amanita pantherina, que es muy tóxica.
Toxinas. La toxina es la rubescenslisina, que es una proteína hemolítica, rompe la membrana citoplásmica de los leucocitos y de los eritrocitos humanos (103). En todos los tejidos investigados, los efectos de la rubescenslisina comenzaban en unos pocos minutos, eran dependientes de la dosis y prácticamente irreversibles (147).
Se detectaron pequeñas diferencias en la sensibilidad entre varios órganos y especies. Las observaciones indican que la toxina es relativamente inespecífica en su ataque a los componentes de las membranas celulares.
Ensayos en animales y en órgano aislado. En los corazones aislados de ratas y cobayos causó una contractura sistólica, que fue precedida por un flujo de salida de potasio y, a veces, por un efecto inotrópico positivo transitorio. En la aurícula izquierda (atrio izquierdo) de cobayo, estimulada eléctricamente, mostró al principio un efecto positivo, seguido de un efecto inotrópico negativo; en la aurícula derecha, que latía espontáneamente, produjo un efecto transitorio positivo seguido de efectos inotrópicos y cronotrópicos negativos. Los atrios fueron menos sensibles que los corazones intactos. En la preparación aislada de nervio frénico-diafragma de rata produjo una contractura, que se asoció con la reducción de la contractilidad indirecta y directa. En el íleon aislado de cobayo produjo una contracción lenta seguida de taquifilaxis. Como la excitabilidad disminuyó debido a rubescenslisina, también lo hizo la excitabilidad por acetilcolina y potasio. La atropina y la feniramina tuvieron sólo efectos antagonistas débiles, pero la papaverina fue más poderosa.
En los hepatocitos de rata aislados, la rubescenslisina provocó una salida rápida de potasio y protusiones de células gruesas, mientras que más tarde las células se volvieron teñibles con azul tripán. En el hígado de rata aislado y perfundido produjo una salida rápida de potasio y de enzimas citoplasmáticas y mitocondriales, y un flujo de salida más lento de β-glucuronidasa lisosómica, acompañado de un aumento en la relación lactato/piruvato y una disminución en la producción de bilis. En el riñón de rata perfundido aislado causó una salida de enzimas citoplásmicas y mitocondriales, junto con proteinuria masiva y una restricción grave de la reabsorción de sodio y potasio y de la producción de orina (147).
Especies causantes del micetismo. División: Basidiomycota; Clase: Agaricomycetes; Orden: Agaricales; Familia: Amanitaceae: Amanita rubescens (Pers.: Fr.) Gray [Bas.: Amanita rubescens Pers.; sin.: Agaricus rubescens (Pers.) Fr., Agaricus rubescens var. rubescens (Pers.) Fr., Amanita rubescens var. rubescens Pers., Amanita rubescens var. alba Coker, Amplariella rubescens (Pers.) E.-J. Gilbert, Limacium rubescens (Pers.) J. Schröt.] forma micorrizas con árboles de madera dura y de madera blanda, crece en pinares y en brezales; en muchos bosques de coníferas en suelos ácidos pobres, donde se encuentran en pequeños grupos, más que individualmente. Se encuentra en Europa (desde el Reino Unido y Francia en el oeste, pasando por toda Alemania, hasta Bielorrusia y Rusia en el este, y desde Islandia y Fenoscandia en el norte hasta España, Italia y Bulgaria en el sur; también en las Islas Canarias), en África (Marruecos y Argelia, Sudáfrica), y en Asia (Israel, Asia Menor, Siberia, Corea y Japón), en América del Norte y Central (103).
2.1.9. Proteína tóxica en Chlorophyllum molybdites
Chlorophyllum molybdites es el hongo que causa la mayor cantidad de intoxicaciones anualmente en la ciudad de Buenos Aires y alrededores (Argentina) (148)(149) y también en ciudades de América del Norte (150). Se considera que la causa de las intoxicaciones en ciudades se debe a su gran tamaño y a su hábito de crecer en áreas cercanas a zonas pobladas. Además, puede confundirse con Macrolepiota bonaerensis Speg. y con el hongo parasol comestible Macrolepiota procera.
Es un hongo imponente con un sombrero de hasta 40 cm de diámetro, de color blanquecino con escamas gruesas de color marrón. Las láminas son libres y blancas y usualmente se vuelven oscuras y verdes con la madurez; esporada verde. El estípite puede tener hasta 25 cm de altura y lleva un anillo.
Chlorophyllum molybdites, (“parasol falso”; en inglés: “false parasol”, “green-spored Lepiota” y “vomiter”) es un hongo muy tóxico que se suele confundir con las especies del complejo Chlorophyllum (“shaggy parasol”) que se encuentran en Europa, América del Norte y América del Sur, Australia y Sudáfrica, o con Coprinus comatus (O.F. Müll.) Pers. (“shaggy mane”). El complejo Chlorophyllum comprende tres especies del género Chlorophyllum que están estrechamente relacionadas: Chlorophyllum rachodes (o rhacodes), Chlorophyllum olivieri y Chlorophyllum brunneum, que se encuentran en Europa, África meridional, EE.UU. y también en la Argentina y Australia. Son hongos apreciados como comestibles de elección, pero contienen toxinas que pueden causar problemas gastrointestinales cuando se consumen crudos o poco cocidos y algunas personas presentan una fuerte respuesta alérgica incluso después de la cocción.
Recientemente, se llevaron a cabo estudios taxonómicos sobre la base de diferencias morfológicas y análisis filogenéticos moleculares de Chlorophyllum, y se reconocieron seis clados bien respaldados y 17 especies filogenéticas. Además, se propone una nueva clasificación infragenérica de Chlorophyllum, en la que el género se divide en seis secciones (151).
Período de latencia. Es muy corto, de 1-3 horas después del consumo (10).
Sintomatología. Los síntomas son predominantemente de naturaleza gastrointestinal, raramente fatales, pero graves (10)(152), con vómitos, diarrea y cólicos, heces con sangre, escalofríos, dolores intestinales, hemorragia gastrointestinal y, en algunos casos, convulsiones (153)(154). Se han detallado los síntomas ocasionados por esta especie en la Argentina (148).
Casos de intoxicaciones. Si bien se considera que este hongo es un irritante gastrointestinal benigno, puede producir toxicidad severa y los pacientes pediátricos tienen una capacidad limitada para perder líquidos antes de volverse hipovolémicos. Se menciona a modo de ejemplo, el caso de una niña de 6 años que desarrolló un shock hipovolémico por intoxicación con C. molybdites (153).
Se describieron dos casos de intoxicación por C. molybdites en adultos, con síntomas gastrointestinales graves, que requirieron hospitalización (154). Se revisaron la distribución geográfica de los casos, los signos y síntomas de la intoxicación y su tratamiento, los principios tóxicos, las esporas como alergenos, las poblaciones particularmente susceptibles y las variaciones en la respuesta asociada con cocinar estos hongos y con idiosincrasias individuales. Se indicó además la aparición ocasional de hongos estériles que carecían de sus características basidiosporas verdes (154).
También se registró la intoxicación de un hombre sano de 43 años que ingirió tres hongos de C. molybdites en la isla francesa de Reunión, situada en el océano Índico al este de Madagascar, y casi una hora y media después, presentó dolor abdominal intenso y diarrea sanguinolenta. El tratamiento sintomático y la reanimación con líquidos en la unidad de emergencia permitió que fuera dado de alta 48 horas después, sin secuelas (155). Recientemente, se registraron casos de intoxicación por C. molybdites en el estado de Kerala, India (156).
Antes de comer los carpóforos de C. molybdites se deben hervir y decantar el líquido antes de cocinarlos.
Toxina. En 1974 Eilers y Nelson demostraron que el componente tóxico de C. molybdites es de naturaleza proteica y realizaron la purificación parcial y caracterización de la proteína tóxica (157) de alto peso molecular, termolábil, que resultó tóxica al administrarla por vía intraperitoneal a animales de laboratorio (157); también se demostró que la toxina puede afectar no sólo a humanos sino también a perros y gallinas (157).
En 2012, Yamada et al. (158) aislaron una proteína tóxica de 23 kDa, que denominaron molibdofilisina, de C. molybdites al seguir su efecto letal en ratones (158). Estos hallazgos parecen estar en contraposición con los de Eilers y Nelson, que informaron que el tóxico de este hongo es una proteína polimérica con una masa molecular superior a 400 kDa, compuesta de monómeros de 40-60 kDa (157). Molibdofilisina es una proteína relativamente pequeña y en consecuencia, puede corresponder al monómero informado por Eilers y Nelson.
El análisis de la secuencia de los fragmentos trípticos de molibdofilisina mostró que esta proteína era altamente homóloga a varias metaloendopeptidasas (MEPs), como GFMEP, POMEP y AMMEP. Estas proteínas contienen un motivo de unión al cinc conocido como aspzincina. Por lo tanto, la molibdofilisina es un miembro de la familia de las deuterolisinas de las proteasas de cinc (158).
Las metaloendopeptidasas se encuentran en muchos hongos y bacterias, pero las descriptas hasta ahora no han presentado toxicidad. En este sentido, molibdofilisina es única y es de gran interés aclarar la relación entre su toxicidad y su actividad MEP. Con ese objetivo, se están realizando las determinaciones de la secuencia completa y la estructura tridimensional de molibdofilisina. Se encontró que la molibdofilisina tiene actividad proteolítica, que se mantuvo a temperaturas de hasta 60 °C con un pH óptimo de 7,0 (158).
También se aislaron de carpóforos de C. molybdites los alcaloides lepiotina A y lepiotina B (Fig. 16) (159), varios esteroides de ergostano con citotoxicidad contra las células Kato III (Fig. 16) (160), una lectina de 32 kDa con especificidad de unión más fuerte hacia el ácido N-glicolilneuramínico (161) y un glicoxiuracilo con otros compuestos conocidos (162).

Figura 16. Estructuras químicas de lepiotina A, lepiotina B, ergostanos libres y un glucopiranósido de ergostano.
Los extractos metanólicos de C. molybdites presentaron actividad antibacteriana contra tres bacterias gram positivas (Staphylococcus aureus, Bacillus subtilis, Micrococcus flavus) y dos bacterias gram negativas (Escherichia coli, Pseudomonas aeruginosa) y exhibieron una considerable actividad antifúngica contra seis patógenos fúngicos humanos (Candida albicans, Candida krusei, Aspergillus fumigatus, Mucor sp., Microsporum gypseum, Trichophyton mentagrophytes) y contra un hongo patógeno no humano (Candida maltosa); también mostró fuertes efectos antioxidantes (163).
Especies causantes del micetismo. División: Basidiomycota; Clase: Agaricomycetes; Subclase: Homobasidiomycetidae; Orden: Agaricales; Familia: Agaricaceae: Chlorophyllum molybdites (G. Mey.) Massee [Bas.: Agaricus molybdites G. Mey.; sin.: Agaricus morganii Peck, A. glaziovii Berkeley, A. guadelupensis Patouillard, Annularia camporum Speg., Chlorophyllum esculentum Massee, C. morganii (Peck) Massee, Lepiota camporum (Speg.) Speg., L. esculenta (Massee) Saccardo & P. Sydow, L. molybdites (G. Mey.) Sacc., L. morganii (Peck) Sacc., L. ochrospora Cooke & Massee, Leucocoprinus molybdites (G. Meyer) Patouillard, Macrolepiota molybdites (G. Meyer) G. Moreno, Bañares & Heykoop, Mastocephalus molybdites (G. Meyer) Kuntze, M. morganii (Peck) Kuntze, Pholiota glaziovii (Berkeley) Sacc.] es gregario, forma “anillos de hadas” en zonas abiertas de bosques de latifoliadas, crece en prados y parques en la ciudad de Buenos Aires y alrededores, provincia de Buenos Aires, Tucumán y Córdoba (Argentina) (48)(164). También crece en Escocia, Australia, Chipre, India, Japón, en el este de América del Norte y California, así como en regiones templadas y subtropicales de todo el mundo (10); Chlorophyllum rachodes (Vittad.) Vellinga [Bas.: Agaricus rachodes Vittad.; sin.: Lepiota rachodes (Vittad.) Quél.; Macrolepiota rachodes (Vittad.) Singer] (“shaggy parasol”) crece generalmente en bosques de coníferas en Tucumán y en Buenos Aires (Argentina) (48)(164). La especie también se escribe rhacodes; Chlorophyllum brunneum (Farl. & Burt) Vellinga [Bas.: Lepiota brunnea Farl. & Burt]; Chlorophyllum olivieri (Barla) Vellinga [Bas.: Lepiota olivieri Barla].
2.1.10. Úlceras gástricas por consumo de Leucocoprinus birnbaumii
Leucocoprinus birnbaumii es un hongo saprotrófico que vive en materia vegetal muy descompuesta (humus o compost) de regiones tropicales y subtropicales; suele crecer en invernaderos y macetas.
Sintomatología. Trastornos gastrointestinales. Las toxinas de Leucoprinus birnbaumii pueden causar úlceras gástricas si se ingieren (165).
Toxinas. Este frágil hongo, que es inofensivo para las plantas, debe su color amarillo a los inusuales alcaloides indólicos tóxicos birnbauminas A y B (Fig. 17), en los que se une una amida del ácido N-hidroxiindol- 3-glioxílico a N-hidroxioxamidina a través de una unidad de tetrametileno. Las N-hidroxioxamidinas fueron descriptas por primera vez en 2005 (166).

Figura 17. Estructuras químicas de birnbaumina A y birnbaumina B.
Especie causante del micetismo: División: Basidiomycota; Clase: Agaricomycetes; Orden: Agaricales; Familia: Agaricaceae: Leucoprinus birnbaumii (Corda) Sing. [Bas.: Agaricus birnbaumii Corda; sin.: Lepiota lutea (Bolton) Godfrin, Leucocoprinus luteus (Bolton) Locq., Lepiota flos-sulphuris (Schnizl.) Mattir., Leucocoprinus flos-sulphuris (Schnizl.) Cejp; L. luteus (Bolton) Locq., Lepiota coprinoides Beeli] crece en la Argentina (provincias de Tucumán, Santa Fe, Buenos Aires y Córdoba) (44)(48)(164) y en EE. UU. (Texas, Indiana, Florida, Oregon, New York) (165).
3. Tratamiento
El tratamiento de las intoxicaciones gastrointestinales suele ser de apoyo, especialmente en los cuadros de deshidratación grave. Se administran fluidos y electrolitos por vía intravenosa, con especial atención en niños y ancianos (10). Se puede usar metoclopramida en casos de vómitos recurrentes una vez que se han vaciado los contenidos gástricos (10).
Los medicamentos antiespasmódicos pueden disminuir los cólicos abdominales y el carbón activado ayuda en las primeras horas de internación para remover la toxina, evitando así su captación.
La intoxicación debida a la ingestión de E. nidorosum y/o E. rhodopolium requiere tratamiento sintomático.
4. Conclusiones
En esta Parte 3 de la serie de cuatro artículos sobre micetismos se han analizado los toxíndromes gastrointestinales o resinoides dentro de los llamados síndromes tempranos, por presentar períodos de latencia menores de 6 horas. Durante mucho tiempo se les restó importancia, y aún hoy en varios trabajos suele indicarse que las toxinas específicas no se conocen. Sin embargo, en realidad, como se ha visto en el presente artículo, las sustancias responsables son muy distintas estructuralmente, desde proteínas hasta terpenoides, vinilglicina, fenol y azocompuestos, pero provocan sintomatología muy similar. En este trabajo se seleccionaron diez ejemplos, si bien existen varios más.
Una gran cantidad de hongos superiores provocan síntomas gastrointestinales de diferente gravedad, lo que depende también de la susceptibilidad del consumidor. Como muchos de los irritantes gastrointestinales son termolábiles, se recomienda proceder a una cocción intensa antes de ingerir cualquier macromiceto o bien hervir los carpóforos descartando el agua utilizada antes de su cocción definitiva.
En cuanto a los tratamientos para los micetismos presentados se debe enfatizar la importancia de realizar un diagnóstico rápido y preciso, con la identificación de los carpóforos por servicios micológicos especializados. En la primera etapa de internación, es fundamental proceder a mejorar el cuadro de deshidratación mediante fluidos y electrolitos; el curso de este tipo de intoxicaciones suele ser favorable.
Es necesario realizar más investigaciones en el área, complementarias de la micológica, tanto desde el punto de vista químico, como bioquímico y médico; el conocimiento del tipo de micetismo, sus toxinas y mecanismos y tratamientos posibles de la intoxicación contribuyen a la adopción de las mejores prácticas clínicas. En los centros médicos se requieren protocolos de intervención claros y de fácil acceso para todos los profesionales de la salud.
El problema que subsiste es que, las intoxicaciones por ingestión de hongos superiores, aunque menos prevalentes que otras patologías, son también menos conocidas, y una estrategia equivocada o tardía puede ser fatal para el paciente.
Agradecimientos
Al Hospital de Clínicas y a la Universidad de Buenos Aires por la infraestructura brindada; a la actual Secretaría de Ciencia, Tecnología e Innovación Productiva (ex Mincyt; Argentina) por las facilidades de acceso a la biblioteca electrónica. A.B.P. es Investigadora Superior del Consejo Nacional de Investigaciones Científicas y Técnicas de la Argentina (CONICET).
1. Pomilio AB, Battista SM, Alonso A. Micetismos. Parte 1: Síndromes con período de latencia tardía. Acta Bioquím Clín Latinoam 2018; 52: 459-87.
2. Pomilio AB, Battista SM, Alonso A. Micetismos. Parte 2: Síndromes con sintomatología demorada y latencia muy larga. Acta Bioquím Clín Latinoam 2019; 53: 79- 107.
3. Nilson S, Persson O. Fungi of Northern Europe 1: larger fungi (excluding gill-fungi). New York: Penguin; 1977. [ Links ]
4. Patočka J. Bolesatine, a toxic protein from the mushroom Rubroboletus satanas. Mil Med Sci Lett (Voj Zdrav Listy) 2018; 87: 14-20. [ Links ]
5. Nuhn ME, Binder M, Taylor AFS, Halling RE, Hibbett DS. Phylogenetic overview of the Boletineae. Fungal Biol 2013; 117: 479-511. [ Links ]
6. Zhao K, Wu G, Yang ZL. A new genus, Rubroboletus, to accommodate Boletus sinicus and its allies. Phytotaxa 2014; 188: 61-77. [ Links ]
7. Sesli E. Preliminary checklist of macromycetes of the East and Middle Black Sea regions of Turkey. Mycotaxon 2007; 99: 71-4. [ Links ]
8. Biketova AYu, Kosakyan A, Wasser SP, Nevo E. New, noteworthy, and rare species of the genus Boletus in Israel. Plant Biosyst 2016; 150: 876-86. [ Links ]
9. Ammirati JA, Traquair JA, Horgen PA. Poisonous mushrooms of the Northern United States and Canada. Minneapolis: University of Minnesota Press, 1985. [ Links ]
10. Benjamin DR. Mushrooms: Poisons and panaceas — a handbook for naturalists, mycologists and physicians. New York: WH Freeman and Company, 1995.
11. Schenk-Jaeger KM, Rauber-Lüthy C, Bodmer M, Kupferschmidt H, Kullak-Ublick GA, Ceschi A. Mushroom poisoning: a study on circumstances of exposure and patterns of toxicity. Eur J Int Med 2012; 23: e85-e91. [ Links ]
12. Kretz O, Creppy EE, Dirheimer G. Characterization of bolesatine, a toxic protein from the mushroom Boletus satanas Lenz and it's effects on kidney cells. Toxicology 1991; 66: 213-24. [ Links ]
13. Kretz O, Creppy EE, Boulanger Y, Dirheimer G. Purification and some properties of bolesatine, a protein inhibiting in vitro protein synthesis, from the mushroom Boletus satanas Lenz (Boletaceae). Arch Toxicol Suppl 1989; 13: 422-7. [ Links ]
14. Kretz O, Reinbolt J, Creppy EE, Dirheimer G. Properties of bolesatine, a translational inhibitor from Boletus satanas Lenz. Amino-terminal sequence determination and inhibition of rat mitochondrial protein synthesis. Toxicol Lett 1992; 64-65: 763-6. [ Links ]
15. Kretz O, Creppy EE, Dirheimer G. Disposition of the toxic protein, bolesatine, in rats: its resistance to proteolytic enzymes. Xenobiotica 1991; 21: 65-73. [ Links ]
16. Wu G, Zhao K, Li YC, Zeng NK, Feng B, Halling R, et al. Four new genera of the fungal family Boletaceae. Fungal Divers 2016; 81: 1-24 [ Links ]
17. Matsuura M, Yamada M, Saikawa Y, Miyairi K, Okuno T, Konno K, et al. Bolevenine, a toxic protein from the Japanese toadstool Boletus venenatus. Phytochemistry 2007; 68: 893-8. [ Links ]
18. Horibe M, Kobayashi Y, Dohra H, Morita T, Murata T, Usui T, et al. Toxic isolectins from the mushroom Boletus venenatus. Phytochemistry 2010; 71: 648-57. [ Links ]
19. Ennamany R, Lavergne JP, Reboud JP, Dirheimer G, Creppy EE. Mode of action of bolesatine, a cytotoxic glycoprotein from Boletus satanas Lenz. Mechanistic approaches. Toxicology 1995; 100: 51-5. [ Links ]
20. Kretz O, Barbieri L, Creppy EE, Dirheimer G. Inhibition of protein synthesis in liver and kidney of mice by bolesatine: mechanistic approaches to the mode of action at the molecular level. Toxicology 1992; 73: 297-304. [ Links ]
21. Ennamany R, Bingen A, Creppy EE, Kretz O, Gut JP, Dubuisson L, et al. Aspirin and heparin prevent hepatic blood stasis and thrombosis induced by the toxic glycoprotein bolesatine in mice. Hum Exp Toxicol 1998; 17: 620-4. [ Links ]
22. Licastro F, Morini MC, Kretz O, Dirheimer G, Creppy EE, Stirpe F. Mitogenic activity and immunological properties of bolesatine, a lectin isolated from the mushroom Boletus satanas Lenz. Int J Biochem 1993; 25: 789-92. [ Links ]
23. Sarup Singh R, Preet Kaur H, Rakesh Kanwar J. Mushroom lectins as promising anticancer substances. Curr Protein Pept Sci 2016; 17: 797-807. [ Links ]
24. Basset L, Ennamany R, Portail JP, Kretz O, Deffieux G, Badoc A, et al. Effects of bolesatine on a cell line from the SP2/O thymic lymphosarcoma. Toxicology 1995; 103: 121-5. [ Links ]
25. Ennamany R, Kretz O, Creppy EE. Effect of bolesatine on phospholipid/calcium dependent protein kinase in Vero cells and in rat thymus. Arch Toxicol 1995; 69: 624-30. [ Links ]
26. Ennamany R, Marzetto S, Saboureau D, Creppy EE. Lipid peroxidation induced by bolesatine, a toxin of Boletus satanas: implication in m5dC variation in Vero cells related to inhibition of cell growth. Cell Biol Toxicol 1995; 11: 47-354. [ Links ]
27. Gachet C, Ennamany R, Kretz O, Ohlmann P, Krause C, Creppy EE, et al. Bolesatine induces agglutination of rat platelets and human erythrocytes and platelets in vitro. Hum Exp Toxicol 1996; 15: 26-9. [ Links ]
28. Assyov B, Bellanger JM, Bertéa P, Courtecuisse R, Koller G, Loizides M, et al. Nomenclatural novelties. Index Fungorum 2015; (243). [ Links ]
29. Della Maggiora M. Nomenclatural novelties. Index Fungorum 2015; (246). [ Links ]
30. Nilsson S, Persson O. Fungi of Northern Europe 2: gill-fungi. New York: Penguin; 1977. [ Links ]
31. Chapman B, Xiao G, Myers S. Early results from field trials using Hypholoma fasciculare to reduce Armillaria ostoyae root disease. Can J Bot 2004; 82: 962-9. [ Links ]
32. Doljak B, Stegnar M, Urleb U, Kreft S, Umek A, Ciglarič M, et al. Screening for selective thrombin inhibitors in mushrooms. Blood Coagul Fibrinolysis 2001; 12: 123-8. [ Links ]
33. Suzuki K, Fujimoto H, Yamazaki M. The toxic principles of Naematoloma fasciculare. Chem Pharm Bull (Tokyo) 1983; 31: 2176-8. [ Links ]
34. Kubo I, Matsumoto A, Kozuka M, Wood WF. Calmodulin inhibitors from the bitter mushroom Naematoloma fasciculare (Fr.) Karst. (Strophariaceae) and absolute configuration of fasciculols. Chem Pharm Bull (Tokyo) 1985; 33: 3821-5. [ Links ]
35. Takahashi A, Kusano G, Ohta T, Ohizumi Y, Nozoe S. Fasciculic acids A, B and C as calmodulin antagonists from the mushroom Naematoloma fasciculare. Chem Pharm Bull (Tokyo) 1989; 37: 3247-50. [ Links ]
36. Kim DS, Baek NI, Oh SR, Jung KY, Lee IS, Kim JH, et al. Anticomplementary activity of ergosterol peroxide from Naematoloma fasciculare and reassignment of NMR data. Arch Pharm Res 1997; 20: 201-5. [ Links ]
37. Chen H-P, Liu J-K. Secondary metabolites from higher fungi. En: Kinghorn AD, Falk H, Gibbons S, Kobayashi S (eds). Progress in the Chemistry of Organic Natural Products. Volumen 106, Cham, Suiza: Springer, 2017; pp. 1-201. [ Links ]
38. Shi XW, Li XJ, Gao JM, Zhang XC. Fasciculols H and I, two lanostane derivatives from Chinese mushroom Naematoloma fasciculare. Chem Biodivers 2011; 8: 1864-70. [ Links ]
39. Kim KH, Moon E, Choi SU, Kim SY, Lee KR. Lanostane triterpenoids from the mushroom Naematoloma fasciculare. J Nat Prod 2013; 76: 845-51. [ Links ]
40. Yaoita Y, Matsuki K, Iijima T, Nakano S, Kakuda R, Machida K, et al. New sterols and triterpenoids from four edible mushrooms. Chem Pharm Bull (Tokyo) 2001; 49: 589-94. [ Links ]
41. Kim KH, Choi SU, Noh HJ, Zee O, Lee KR. Cytotoxic ergosterol derivatives from the mushroom Naematoloma fasciculare. Nat Prod Sci 2014; 20: 76-9. [ Links ]
42. Ríos JL, Andújar I, Recio MC, Giner RM. Lanostanoids from fungi: a group of potential anticancer compounds. J Nat Prod 2012; 75: 2016-44. [ Links ]
43. Merdivan S, Lindequist U. Ergosterol peroxide: a mushroom-derived compound with promising biological activities - a review. Int J Med Mushrooms 2017; 19: 93-105. [ Links ]
44. Wright JE, Albertó E. Guía de hongos de la región pampeana. I. Hongos con laminillas. Buenos Aires: Editorial LOLA; 2002. [ Links ]
45. Niveiro N, Albertó E. Checklist of the Argentine Agaricales 2. Coprinaceae and Strophariaceae. Mycotaxon 2012; 120: 505. [ Links ]
46. Coimbra VRM. Checklist of Central and South American Agaricales (Basidiomycota). II: Strophariaceae. Mycosphere 2015; 6: 441-58. [ Links ]
47. Franco-Molano AE, Corrales A, Vasco-Palacios AM. Macrohongos de Colombia II. Listado de especies de los órdenes Agaricales, Boletales, Cantharellales y Russulales (Agaricomycetes, Basidiomycota). Actual Biología 2010; 32: 89-114. [ Links ]
48. Raithelhuber J. Nueva Flora Micológica Argentina. Mycosur, Stuttgart, 2004. [ Links ]
49. Gill M, Strauch RJ. Constituents of Agaricus xanthodermus Genevier: the first naturally endogenous azo compound and toxic phenolic metabolites. Z. Naturforsch. C 1984; 39: 1027-9. [ Links ]
50. Niveiro N, Albertó E. Checklist of the Argentine Agaricales 5. Agaricaceae. Mycotaxon 2013; 122: 491. [ Links ]
51. Grilli E. Studies on the genus Hebeloma, What is H. versipelle s. Bresadola? Riv Micol Romana, Boll Assoc Micol Ecol Romana 100-101, 2017; 33: 39-50. https://www.researchgate.net/publication/320466836_Studies_on_the_genus_Hebeloma_What_is_H_versipelle_s_Bresadola. Fecha de acceso: 5 de marzo de 2019. [ Links ]
52. Ho B-TQ, Pham N-DH, Shimizu K, Fukiharu T, Truong BN, Suzuki A. The first record of Hebeloma vinosophyllum (Strophariaceae) in Southeast Asia. Mycotaxon 2014; 128: 25-36. [ Links ]
53. Niveiro N, Albertó E. Checklist of the Argentine Agaricales 7. Cortinariaceae and Entolomataceae. Check List 2014; 10: 72-96. [ Links ]
54. Grilli E, Beker HJ, Eberhardt U, Schütz N, Leonardi M, Vizzini A. Unexpected species diversity and contrasting evolutionary hypotheses in Hebeloma (Agaricales) sections Sinapizantia and Velutipes in Europe. Mycol Prog 2016; 15: 1-46. [ Links ]
55. Fujimoto H, Suzuki K, Hagiwara H, Yamazaki M. New toxic metabolites from a mushroom, Hebeloma vinosophyllum. I. Structures of hebevinosides I, II, III, IV, and V. Chem Pharm Bull (Tokyo) 1986; 34: 88-99. [ Links ]
56. Fujimoto H, Hagiwara H, Suzuki K, Yamazaki M. New toxic metabolites from a mushroom, Hebeloma vinosophyllum. II. Isolation and structures of hebevinosides VI, VII, VIII, IX, X, and XI. Chem Pharm Bull (Tokyo) 1987; 35: 2254-60. [ Links ]
57. Fujimoto H, Maeda K, Yamazaki M. New toxic metabolites from a mushroom, Hebeloma vinosophyllum. III. Isolation and structures of three new glycosides, hebevinosides XII, XIII and XIV, and productivity of the hebevinosides at three growth stages of the mushroom. Chem Pharm Bull (Tokyo) 1991; 39: 1958-61. [ Links ]
58. Fujimoto H, Takano Y, Yamazaki M. Isolation, identification and pharmacological studies on three toxic metabolites from a mushroom, Hebeloma spoliatum. Chem Pharm Bull (Tokyo) 1992; 40: 869-72. [ Links ]
59. De Bernardi M, Fronza G, Gianotti MP, Mellerio G, Vidari G, Vita-Finzi P. Fungal metabolites XIII: New cytotoxic triterpene from Hebeloma species (Basidiomycetes). Tetrahedron Lett 1983; 24: 1635-8. [ Links ]
60. Garlaschelli L, Vidari G, Virtuani M, Vita-Finzi P, Mellerio G. The structures of new lanostane triterpenes from the fruiting bodies of Hebeloma senescens. J Nat Prod 1995; 58: 992-1002. [ Links ]
61. Shao HJ, Qing C, Wang F, Zhang YL, Luo DQ, Liu JK. A new cytotoxic lanostane triterpenoid from the basidiomycete Hebeloma versipelle. J Antibiot 2005; 58: 828-31. [ Links ]
62. Oliveira AG, Desjardin DE, Perry BA, Stevani CV. Evidence that a single bioluminescent system is shared by all known bioluminescent fungal lineages. Photochem Photobiol Sci 2012; 11: 848-52. [ Links ]
63. Kirchmair M, Morandell S, Stolz D, Pöder R, Sturmbauer C. Phylogeny of the genus Omphalotus based on nuclear ribosomal DNA-sequences. Mycologia 2004; 96: 1253-60. [ Links ]
64. Bresinsky A, Besl H. A Colour Atlas of Poisonous Fungi: a Handbook for pharmacists, doctors, and biologists. 1a ed, London, Wolfe Publishing Ltd., 1990. [ Links ]
65. Miller OK Jr. Observations on the genus Omphalotus in Australia. Mycol Helv 1994; 6: 91-100. [ Links ]
66. Shirmila Jose G, Radhamany PM. Identification and determination of antioxidant constituents of bioluminescent mushroom. Asian Pac J Trop Biomed 2012; 2 (1 Suppl): S386-91. [ Links ]
67. Moreno G, Esteve-Raventós F, Pöder R, Ayala N. Omphalotus olivascens var. indigo, var. nov. from Baja California (Mexico). Mycotaxon 1993; 48: 217-22. [ Links ]
68. Omphalotus japonicus (Kawam.) Kirchm. & O.K. Mill. MycoBank. International Mycological Association. Disponible en: http://www.mycobank.org/BioloMICS.aspx?Table=Mycobank&Rec=411196&Fields=All. Fecha de acceso: 5 de marzo de 2019. [ Links ]
69. Redhead SA, Neda H. (1741) Proposal to conserve the name Pleurotus japonicus against Agaricus guepiniformis and Pleurotus harmandii (Basidiomycota). Taxon 2006; 55: 1032-3. [ Links ]
70. Norvell LL. Report of the Nomenclature Committee for Fungi: 14. Taxon 2008; 57: 637-9. [ Links ]
71. Fukasawa Y, Osono T, Takeda H. Beech log decomposition by wood-inhabiting fungi in a cool temperate forest floor: a quantitative analysis focused on the decay activity of a dominant basidiomycete Omphalotus guepiniformis. Ecol Res 2010; 25: 959-66. [ Links ]
72. Ka KH, Park H, Hur TC, Bak WC. Formation of fruiting body of Omphalotus japonicus by sawdust cultivation. Korean J Mycol 2010; 38: 80-2. [ Links ]
73. Yang ZL, Feng B. The genus Omphalotus (Omphalotaceae) in China. Mycosystema 2013; 32: 545- 56. [ Links ]
74. Tsurida S, Akai K, Hiwaki H, Suzuki A, Akiyama H. Multiplex real-time PCR assay for simultaneous detection of Omphalotus guepiniformis and Lentinula edodes. Biosci Biotechnol Biochem 2012; 76: 1343-9. [ Links ]
75. Diaz JH. Syndromic diagnosis and management of confirmed mushroom poisonings. Crit Care Med 2005; 33: 427-36. [ Links ]
76. Saviuc P, Danel V. New syndromes in mushroom poisoning. Toxicol Rev 2006; 25: 199-209. [ Links ]
77. McMorris TC, Kelner MJ, Wang W, Estes LA, Montoya MA, Taetle R. Structure-activity relationship of illudins: analogs with improved therapeutic index. J Org Chem 1992; 57: 6876-83. [ Links ]
78. McMorris TC, Anchel M. The structures of the basidiomycete metabolites illudin S and illudin M. J Am Chem Soc 1963; 85: 831-2. [ Links ]
79. Arnone A, Cardillo R, Nasini G, de Pava OV. Secondary mould metabolites. Part 31. Isolation and structure elucidation of illudins A and B, and illudalenol, new sesquiterpenoids from Clitocybe illudens. J Chem Soc, Perkin Trans 1 1991; 733-7. [ Links ]
80. Liu J-K. N-Containing compounds of macromycetes. Chem Rev 2005; 105: 2723-44. [ Links ]
81. Tanaka K, Inoue T, Tezuka Y, Kikuchi T. Metabolism by rat liver cytosol of illudin S, a toxic substance of Lampteromyces japonicus. II. Characterization of illudin S-metabolizing enzyme. Xenobiotica 1992; 22: 33-9. [ Links ]
82. Tanaka K, Inoue T, Tezuka Y, Kikuchi T. Metabolism of illudin S, a toxic substance of Lampteromyces japonicus: urinary excretion of mercapturic acids in rat. Xenobiotica 1996; 26: 347-54. [ Links ]
83. Tanaka K, Inoue T, Tezuka Y, Kikuchi T. Michael-type addition of illudin S, a toxic substance from Lampteromyces japonicus, with cysteine and cysteine-containing peptides in vitro. Chem Pharm Bull (Tokyo) 1996; 44: 273-9. [ Links ]
84. Kirchmair M. Identification of illudins in Omphalotus nidiformis and Omphalotus olivascens var. indigo by column liquid chromatography–atmospheric pressure chemical ionization tandem mass spectrometry. J Chromatogr A 1999; 832: 247-52.
85. Burgess ML, Zhang YL, Barrow KD. Characterization of new illudanes, illudins F, G, and H from the basidiomycete Omphalotus nidiformis. J Nat Prod 1999; 62: 1542-4. [ Links ]
86. Beattie KD, Ulrich R, Grice ID, Uddin SJ, Blake TB, Wood KA, et al. Ethanolic and aqueous extracts derived from Australian fungi inhibit cancer cell growth in vitro. Mycologia 2011; 103: 458-65. [ Links ]
87. Sterner O, Etzel W, Mayer A, Anke H. Omphalotin, a new cyclic peptide with potent nematicidal activity from Omphalotus olearius II. Isolation and structure determination. Nat Prod Lett 1997; 10: 33-8. [ Links ]
88. Mayer A, Anke H, Sterner O. Omphalotin, A new cyclic peptide with potent nematicidal activity from Omphalotus olearius I. Fermentation and biological activity. Nat Prod Lett 1997; 10: 25-32. [ Links ]
89. Mayer A, Kilian M, Hoster B, Sterner O, Anke H. In vitro and in vivo nematicidal activities of the cyclic dodecapeptide omphalotin A. Pestic Sci 1999; 55: 27-30. [ Links ]
90. Liermann JC, Opatz T, Kolshorn H, Antelo L, Hof C, Anke H. Omphalotins E–I, five oxidatively modified nematicidal cyclopeptides from Omphalotus olearius. Eur J Org Chem 2009; 2009: 1256-62.
91. Schobert R, Knauer S, Seibt S, Biersack B. Anticancer active illudins: recent developments of a potent alkylating compound class. Curr Med Chem 2011; 18: 790-807. [ Links ]
92. Baekelandt M. Irofulven (MGI Pharma). Curr Opin Investig Drugs 2002; 3: 1517-26. [ Links ]
93. Staake MD, Kashinatham A, McMorris TC, Estes LA, Kelner MJ. Hydroxyurea derivatives of irofulven with improved antitumor efficacy. Bioorg Med Chem Lett 2016; 26: 1836-8. [ Links ]
94. Yue Q, Gao G, Zou G, Yu H, Zheng X. Natural products as adjunctive treatment for pancreatic cancer: recent trends and advancements. Biomed Res Int 2017; 2017: 8412508. [ Links ]
95. Wawrzyn GT, Quin MB, Choudhary S, López-Gallego F, Schmidt-Dannert C. Draft genome of Omphalotus olearius provides a predictive framework for sesquiterpenoid natural product biosynthesis in Basidiomycota. Chem Biol 2012; 19: 772-83. [ Links ]
96. Ortega A, Esteve-Raventós F, Gómez J, Reyes JD. Contributo allo studio della micoflora dell' Andalusia (Spagna), XIV, Agaricales VII. Boll Gruppo Micol G. Bresadola, Nuova Seria BGMB 2000; 43: 45-7. [ Links ]
97. Klemm G. Beobachtungen über den Verlauf einer Massenvergiftung mit dem Bruchkreizker Lactarius helvus Fries. Mykol Mitteilungsbl 1961; 5: 1-4. [ Links ]
98. Arora D. All that the rain promises and more: a hip pocket guide to Western mushrooms. Berkeley, California: Ten Speed Press 1991; p. 19. [ Links ]
99. Reichl F-X, Ritter L. Illustrated Handbook of Toxicology. Stuttgart, Alemania: Thieme. 2011; p. 296. [ Links ]
100. Barceloux DG. Medical toxicology of natural substances: foods, fungi, medicinal herbs, plants, and venomous animals. John Wiley & Sons, Inc. 2008. [ Links ]
101. Sterner O, Bergman R, Kihlberg J, Wickberg B. The sesquiterpenes of Lactarius vellereus and their role in a proposed chemical defense system. J Nat Prod 1985; 48: 279-88. [ Links ]
102. Anke H, Sterner O. Comparison of the antimicrobial and cytotoxic activities of twenty unsaturated sesquiterpene dialdehydes from plants and mushrooms. Planta Med 1991; 57: 344-6. [ Links ]
103. Gry J, Andersson C. Mushrooms traded as food. Vol. II, sec. 2. Nordic Council of Ministers, Copenhague, Dinamarca, 2014. [ Links ]
104. Gilardoni G, Malagòn O, Tosi S, Clericuzio M, Vidari G. Lactarane sesquiterpenes from the European mushrooms Lactarius aurantiacus, L. subdulcis, and Russula sanguinaria. Nat Prod Commun 2014; 9: 319-22. [ Links ]
105. Vidari G, Vita-Finzi P. Sesquiterpenes and other secondary metabolites of genus Lactarius (Basidiomycetes): Chemistry and biological activity. En: Atta-ur-Rahman (ed), Studies in Natural Products Chemistry. Vol. 17, Part D, Elsevier, 1995; pp. 153-206. [ Links ]
106. Kobata K, Kano S, Shibata H. New lactarane sesquiterpenoid from the fungus Russula emetica. Biosci Biotechnol Biochem 1995; 59: 316-8. [ Links ]
107. Rapior S, Fons F, Bessière J-M. The fenugreek odor of Lactarius helvus. Mycologia 2000; 92: 305-8. [ Links ]
108. Dickschat JS. Fungal volatiles - a survey from edible mushrooms to moulds. Nat Prod Rep 2017; 34: 310-28. [ Links ]
109. Suortti T. Improved analytical and preparative methods for necatorin from Lactarius necator (Fr.) Karst. mushroom. J Chromatogr 1984; 301: 303-7. [ Links ]
110. Suortti T. Stability of necatorin, a highly mutagenic compound from Lactarius necator mushroom. Food Chem Toxicol 1984; 22: 579-81. [ Links ]
111. Klamann J-D, Fugmann B, Steglich W. Alkaloidal pigments from Lactarius necator and L. atroviridis. Phytochemistry 1989; 28: 3519-22. [ Links ]
112. Tanaka Y, Kawahara S, Eng AH, Takei A, Ohya N. Structure of cis-polyisoprene from Lactarius mushrooms. Acta Biochim Pol 1994; 41: 303-9. [ Links ]
113. Tanaka Y, Kawahara S, Aikhwee E, Shiba K, Ohya N. Initiation of biosynthesis in cis polyisoprenes. Phytochemistry 1995; 39: 779-84. [ Links ]
114. Panchak LV, Antoniuk VO. [Purification of lectin from fruiting bodies of Lactarius rufus (Scop.: Fr.) Fr. and its carbohydrate specificity]. Artículo en idioma ucraniano, resumen en inglés. Ukr Biokhim Zh 2007; 79: 123-8. [ Links ]
115. Kobata K, Wada T, Hayashi Y, Shibata H. Volemolide, a novel norsterol from the fungus Lactarius volemus. Biosci Biotechnol Biochem 1994; 58: 1542-4. [ Links ]
116. Sivakumar M, Bhat SV, Nagasampagi BA. Volemitol en L. volemus. En: Chemistry of Natural Products. Berlin, Germany: Springer. 2005; p. 495. [ Links ]
117. Yue J-M, Chen S-N, Lin Z-W, Sun H-D. Sterols from the fungus Lactarius volemus. Phytochemistry 2001; 56: 801-6. [ Links ]
118. Muszyńska B, Sułkowska-Ziaja K. Analysis of indole compounds in edible Basidiomycota species after thermal processing. Food Chem 2012; 132: 455-9. [ Links ]
119. Muszyńska B, Maślanka A, Sułkowska-Ziaja K, Krzek J. TLC-UV analysis of indole compounds and other nitrogen-containing bases in the fruiting bodies of Lactarius deterrimus. J Planar Chromatogr 2007; 20: 57-60. [ Links ]
120. Stone TW, Mackay GM, Forrest CM, Clark CJ, Darlington LG. Tryptophan metabolites and brain disorders. Clin Chem Lab Med 2003; 41: 852-9. [ Links ]
121. Guillot J, Giollant M, Damez M, Dusser M. Isolation and characterization of a lectin from the mushroom Lactarius deliciosus. J Biochem 1991; 109: 840-5. [ Links ]
122. Giollant M, Guillot J, Damez M, Dusser M, Didier P, Didier E. Characterization of a lectin from Lactarius deterrimus. Plant Physiol 1993; 101: 513-22. [ Links ]
123. Bergendorff O, Sterner O. The sesquiterpenes of Lactarius deliciosus and Lactarius deterrimus. Phytochemistry 1988; 27: 97-100. [ Links ]
124. Koul SK, Taneja Sc, Ibraham SP, Dhar KL, Atal CK. A C-formylated azulene from Lactarius deterrimus. Phytochemistry 1985; 24: 181-2. [ Links ]
125. Niveiro N, Popoff OF, Albertó EO. Hongos comestibles silvestres: especies exóticas de Suillus (Boletales, Basidiomycota) y Lactarius (Russulales, Basidiomycota) asociadas a cultivos de Pinus elliottii del Nordeste Argentino. Bonplandia 2009; 18: 65-71. [ Links ]
126. Barroetaveña C, Cázares E, Rajchenberg M. Ectomycorrhizal fungi associated with ponderosa pine and Douglas-fir: a comparison of species richness in native western North American forests and Patagonian plantations from Argentina. Mycorrhiza 2007; 17: 355-73. [ Links ]
127. Suzuki K, Une T, Fujimoto H, Yamazaki M. [Studies on the toxic components of Rhodophyllus rhodopolius. I. The biological activities and screening of the toxic principles]. Artículo en idioma japonés. Yakugaku Zasshi 1987; 107: 971-7. [ Links ]
128. Suzuki K, Une T, Yamazaki M. [Studies on the toxic components of Rhodophyllus rhodopolius. II. Partial purification and properties of the hemolysin from Rhodophyllus rhodopolius: examination on the condition of the hemolysis]. Artículo en idioma japonés. Yakugaku Zasshi 1988; 108: 221-5. [ Links ]
129. Suzuki K, Une T, Yamazaki M, Takeda T. Purification and some properties of a hemolysin from the poisonous mushroom Rhodophyllus rhodopolius. Toxicon 1990; 28: 1019-28. [ Links ]
130. Chapuis J-R. Jahresbericht des Verbandstoxikologen für das Jahr 1983. Schweiz Z Pilzk 1984; 62: 196-7. [ Links ]
131. Bocchi A, Bracchi PG, Delbono G, Cadonici O. Segnalazioni di intossicazioni da funghi nel Parmense. Ann Fac Medic Vet di Parma 1995; 15: 251-6. [ Links ]
132. Trueb L, Carron P-N, Saviuc P. Intoxication par les champignons. Rev Med Suisse 2013; 9: 1465-72. [ Links ]
133. Kondo K, Nakamura K, Ishigaki T, Sakata K, Obitsu S, Noguchi A, et al. Molecular phylogenetic analysis of new Entoloma rhodopolium-related species in Japan and its identification method using PCR-RFLP. Sci Rep 2017; 7: Article number: 14942. [ Links ]
134. Alder AE. Die Pilzvergiftungen in der Schweiz, während 40 Jahren. Bull Suisse Mycol 1960; 38: 65-73. [ Links ]
135. Shidei Y. A case report of poisoning by a Rhodophyllus sp. probably Rhodophyllus rhodopolius. Trans Mycol Soc Jpn 1976; 17: 95-6. [ Links ]
136. Alder AE. Erkennung und Behandlung der Pilzvergiftung. Dtsch Med Wochenschr 1961; 86: 1121-7. [ Links ]
137. Maeder A. Rapport toxicologique 1974. Bull Suisse Mycol 1975; 53: 150-4. [ Links ]
138. Ishihara Y, Yamaura Y. Descriptive epidemiology of mushroorn poisoning in Japan. (Nippon Eiseigaku Zasshi) Japan J Hyg 1992; 46: 1071-8. [ Links ]
139. Azéma RC. Mycotoxicologie. Bull Soc Mycol Fr 1982; 98: 23-9. [ Links ]
140. Bastida J, Viladomat F, Llabrés JM, Codina C. Screening of higher fungi from Catalonia for alkaloids-II. Int J Crude Drug Res 1987; 25: 129-32. [ Links ]
141. Tuzen M, Sesli E, Soylak M. Trace element levels of mushroom species from East Black Sea region of Turkey. Food Control 2007; 18: 806-10. [ Links ]
142. Falandysz J. Mercury in mushrooms and soil of the Tarnobrzeska Plain, south-eastern Poland. J Environ Sci Health, Part A 2002; 37: 343-52. [ Links ]
143. Byrne AR, Slejkovec Z, Stijve T, Fay L, Gössler W, Gailer J, et al. Arsenobetaine and other arsenic species in mushrooms. Appl Organometallic Chem 1995; 9: 305-13. [ Links ]
144. Maki T, Takahaski K, Shibata S. Isolation of the vomiting principles from the mushroom Rhodophyllus rhodopolius. J Agric Food Chem 1985; 33: 1204-5. [ Links ]
145. Agerer R. Colour Atlas of Ectomycorrhizae. Schwäbisch Gmünd: Einhorn-Verlag. 2002; p. 116-7. [ Links ]
146. Knudsen H, Vesterholt J. (eds) Funga Nordica: agaricoid, boletoid and cyphelloid genera. Nordsvamp, 2008. [ Links ]
147. Odenthal KP, Seeger R, Braatz R, Petzinger E, Moshaf H, Schmitz-Dräger C. Damage in vitro to various organs and tissues by rubescenslysin from the edible mushroom Amanita rubescens. Toxicon 1982; 20: 765-81. [ Links ]
148. Albertó E. Síndrome de intoxicación producido por el hongo Chlorophyllum molybdites. Síntomas y diagnóstico botánico. Bol Farmacoterap Toxicol 2001; 8: 13-20. [ Links ]
149. Romano GM, Iannone L, Novas MV, Carmarán C, Romero AI, Lopez SE, et al. Hongos tóxicos en la ciudad de Buenos Aires y alrededores. Medicina (Buenos Aires) 2013; 73: 406-10. [ Links ]
150. Blayney D, Rosenkranz E, Zettner A. Mushroom poisoning from Chlorophyllum molybdites. West J Med 1980; 132: 74-7. [ Links ]
151. Ge Z-W, Jacobs A, Vellinga EC, Sysouphanthong P, van der Walt R, et al. A multi-gene phylogeny of Chlorophyllum (Agaricaceae, Basidiomycota): new species, new combination and infrageneric classification. MycoKeys 2018; 65-90. [ Links ]
152. Chlorophyllum molybdites. En: Urban mushrooms. Accesible en: http://urbanmushrooms.com/index.php?id=4. Fecha de acceso: 5 de marzo de 2019. [ Links ]
153. Stenklyft PH, Augenstein WL. Chlorophyllum molybdites--severe mushroom poisoning in a child. J Toxicol Clin Toxicol 1990; 28: 159-68. [ Links ]
154. Lehmann PF, Khazan U. Mushroom poisoning by Chlorophyllum molybdites in the Midwest United States. Cases and a review of the syndrome. Mycopathologia 1992; 118: 3-13. Comentario en: Rippon JW. Mycetismus ‘mushroom poisoning'. Mycopathologia 1992; 118: 1-2.
155. Schmitt C, Pineau-Delabi F, Olivier J, Simon N, de Haro L. Intoxication par ingestion de Chlorophyllum molybdites sur l'Île de la Réunion: à propos d'un cas. [Chlorophyllum molybdites poisoning in Reunion Island: A case study]. Toxicol Anal Clin 2016; 28: 143-5. [ Links ]
156. Bijeesh C, Vrinda KB, Pradeep CK. Mushroom poisoning by Chlorophyllum molybdites in Kerala. J Mycopathol Res 2017; 54: 477-83. [ Links ]
157. Eilers FI, Nelson LR. Characterization and partial purification of the toxin of Lepiota morganii. Toxicon 1974; 12: 557-63. [ Links ]
158. Yamada M, Tokumitsu N, Saikawa Y, Nakata M, Asano J, Miyairi K, et al. Molybdophyllysin, a toxic metalloendopeptidase from the tropical toadstool, Chlorophyllum molybdites. Bioorg Med Chem 2012; 20: 6583- 8. [ Links ]
159. Ohta T, Inoue H, Kusanao G, Oshima Y. Lepiotins A and B, new alkaloids from the mushrooms, Macrolepiota neomastoidea and Chlorophyllum molybdites. Heterocycles 1998; 47: 883-91. [ Links ]
160. Yoshikawa K, Ikuta M, Arihara S, Matsumura E, Katayama S. Two new steroidal derivatives from the fruit body of Chlorophyllum molybdites. Chem Pharm Bull (Tokyo) 2001; 49: 1030-2. [ Links ]
161. Kobayashi Y, Kobayashi K, Umehara K, Dohra H, Murata T, Usui T, et al. Purification, characterization, and sugar binding specificity of an N-glycolylneuraminic acid-specific lectin from the mushroom Chlorophyllum molybdites. J Biol Chem 2004; 279: 53048-55. [ Links ]
162. Su Z, Wang P, Yuan W, Li S. Chemical constituents from the fruit body of Chlorophyllum molybdites. Nat Prod Commun 2013; 8: 1227-8. [ Links ]
163. Al-Fatimi M, Schröder G, Kreisel H, Lindequist U. Biological activities of selected basidiomycetes from Yemen. Pharmazie 2013; 68: 221-6. [ Links ]
164. Niveiro N, Albertó E. Checklist of the Argentine Agaricales 5. Agaricaceae. Mycotaxon 2013; 122: 491. [ Links ]
165. Kuo M. Leucocoprinus birnbaumii. MushroomExpert. Com. Disponible en: http://www.mushroomexpert.com/leucocoprinus_birnbaumii.html. Fecha de acceso: 5 de marzo de 2019. [ Links ]
166. Bartsch A, Bross M, Spiteller P, Spiteller M, Steglich W. Birnbaumin A and B: two unusual 1-hydroxyindole pigments from the “flower pot parasol” Leucocoprinus birnbaumii. Angew Chem Int Ed Engl 2005; 44: 2957-9.
Recibido: 18 de septiembre de 2018
Aceptado: 7 de marzo de 2019

