










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista argentina de microbiología
versión impresa ISSN 0325-7541versión On-line ISSN 1851-7617
Rev. argent. microbiol. v.37 n.2 Ciudad Autónoma de Buenos Aires abr./jun. 2005
M.C. Nally1,2*, Y.P. Maturano1,2, F. Vázquez1,2, M.E. Toro1
1Instituto de Biotecnología, Facultad de Ingeniería, Universidad Nacional de San Juan. Avda. Libertador General San Martín 1109 (0este) (5400) San Juan. 2Departamento de Geofísica, Astronomía y Biología. Facultad de Ciencias Exactas, Físicas y Naturales - UNSJ - CUIM Av. I. De la Roza y Meglioli (5400) San Juan.
*Correspondencia. E-mail: cristinanally@yahoo.com.ar
RESUMEN
La composición química del vino constituye el fundamento de la posterior respuesta sensorial del producto y está determinada por varios factores, como las relaciones levadura-levadura. Se denomina fenómeno killer a la secreción por parte de ciertas cepas de levadura de una proteína tóxica que mata a células denominadas sensibles. El conocimiento del comportamiento en condición aeróbica de cultivos mixtos killer-sensible es útil para relacionarlo con la primera fase de la fermentación enológica, ya que en ella puede definirse la prevalencia o no de la cepa killer. Además, el empleo de mutantes con el plásmido curado permite comparaciones más precisas. El objetivo fue analizar el mecanismo de competencia por sustrato en levaduras killer de Saccharomyces cerevisiae y su mutante sensible con el plásmido curado, empleando distintas fuentes de nitrógeno. Si las muestras se incuban a temperatura de inactivación de la toxina, se evita la infraestimación de células sensibles. Los resultados del co-cultivo de las cepas en proporciones iguales muestran el rol desempeñado por la fuente de nitrógeno en la actividad killer. Cuando el inóculo es 10%K-90%S, el modelo de exclusión competitiva planteado para levaduras killer deja paso a otras variables de competencia.
Palabras clave: Saccharomyces cerevisiae, salvaje killer, isogénica sensible, cultivos mixtos, fuentes de nitrógeno
SUMMARY
Behaviour of a wild Saccharomyces cerevisiae killer yeast and its isogenic sensitive one with respect to different nitrogen sources in mixed cultures. Wine chemical composition is the outcome of complex chemosensory interactions that are difficult to predict because of the influences of many variables, like as yeast-yeast interactions. Killer phenomenon implicates the secretion of a toxic protein by some yeasts, that kill other yeasts called sensitive. The knowledge of the behaviour of killer-sensitive mixed cultures in aerobic conditions is useful to be related with the first stages of oenological fermentation. In these stages it can be defined the killer prevalence in the medium. Also, the use of cured plasmid mutants allows better comparisons. The objective was to analyse the mechanism of substrate competition in Saccharomyces cerevisiae killer strains and its sensitive cured plasmid mutant, using different nitrogen sources. When samples were incubated at the toxin inactivation temperature, the infraestimation of sensitive cells is avoided. Results obtained in co-cultures (50%K-50%S) show the role of the nitrogen source in killer activity. Results obtained with 10%K-90%S inoculum, show that there are another competence variables than the competitive exclusion model for killer yeasts.
Key words: Saccharomyces cerevisiae, wild killer yeast, isogenic sensitive yeast, mixed cultures, nitrogen sources
INTRODUCCIÓN
La composición química del vino constituye el fundamento de la posterior respuesta sensorial del producto y está determinada por varios factores. Estos incluyen el varietal de uva, las condiciones geográficas y viticulturales, la ecología microbiana de la uva y las prácticas relacionadas con el proceso fermentativo (5). Las interacciones entre las distintas especies ocurren de forma ininterrumpida y con características particulares a lo largo de las distintas etapas de la fermentación, por lo que el impacto en la eficiencia del proceso y la calidad del producto necesita ser identificado y evaluado. Por lo tanto, la particularidad en cuanto a la contribución de las levaduras a las propiedades sensoriales del vino depende de las especies que interactúan en su proceso y la ecología de la fermentación, con sus factores determinantes (5).
Una de las interacciones levadura-levadura del tipo antagónico, es el denominado fenómeno killer que implica la secreción por parte de ciertas cepas de levadura de una proteína tóxica de baja masa molecular (llamada toxina killer, a la cual ellas son inmunes) que mata a células denominadas sensibles, las cuales pueden ser del mismo o diferentes géneros.
Estudios genéticos en Saccharomyces cerevisiae han demostrado que la capacidad de producir micotoxinas es heredada citoplasmáticamente y está relacionada con la presencia de plásmidos de doble cadena lineal de ARN (dsRNA), encapsulados, formando partículas tipo virus (VLP) no infecciosas dentro del citoplasma de las células (20).
Aunque existe más de un tipo de variable que afecta la expresión de los fenotipos killer y sensibles, abunda evidencia de que este tipo de interacciones pueden determinar la evolución de las distintas poblaciones de levaduras durante la fermentación. En algunas ocasiones una cepa killer de Saccharomyces cerevisiae predomina al final del proceso fermentativo, sugiriendo que la expresión de la toxina le permitió conducir parte de la vinificación (5).
Durante la fermentación enológica, el crecimiento de levaduras indígenas puede conferir propiedades sensoriales inadecuadas al vino. La metodología fundamental para reducir el riesgo de este tipo de efecto es el de conducir la fermentación con un starter de Saccharomyces cerevisiae en presencia de SO2. El adicionar SO2 a la fermentación enológica suprime el crecimiento de buena parte de las levaduras indígenas potenciando así la prevalencia y acción fermentativa del starter. Las levaduras killer pueden proveer un método alternativo para el control de levaduras no deseadas (21).
De acuerdo con estudios previos llevados a cabo en bodegas (17), el efecto killer se ve significativamente influenciado por la proporción inicial de células killer y sensibles en el inicio de la fermentación. No se han detectado efectos notables en proporciones 1:7, pero sí se observó una actividad killer pronunciada con una proporción inicial de células killer y sensibles de 1:1. Sin embargo, es necesario destacar que en condiciones controladas y volúmenes pequeños, los resultados pueden diferir.
El mosto de uva contiene una amplia variedad de compuestos nitrogenados (ión amonio, aminoácidos, péptidos y polipéptidos de cadena corta) que las levaduras usan como fuente de nitrógeno. Este tipo de compuestos, además de ser esenciales para el proceso de vinificación en cuanto a su decisiva influencia en el crecimiento de las levaduras, también afecta la formación de alcoholes superiores que contribuyen al aroma del vino exaltando su calidad (10). Cuando una cepa de levadura posee el carácter killer, esto puede alterar el metabolismo del nitrógeno en cuanto a que la toxina killer decrece el gradiente de iones a través de la membrana plasmática de la célula sensible, interrumpiendo el transporte acoplado de protones y aminoácidos. Además, la toxina causa la pérdida por parte de la célula de metabolitos pequeños como ATP, glucosa y aminoácidos (15).
Desde el inicio de las investigaciones sobre esta propiedad hasta la actualidad, se han producido y empleado mutantes obtenidos a partir de cepas de levadura killer para uso tanto en investigación básica como aplicada (2, 3, 6, 8,13, 18, 19). Resulta entonces, particularmente útil su empleo para elucidar aspectos poco investigados de este fenómeno, como es el comportamiento de cepas killer en cultivos mixtos killer-sensible.
El objetivo de este trabajo fue analizar el mecanismo de competencia por el sustrato, con énfasis en la relación killer - sensible, en levaduras killer de la especie S. cerevisiae y su mutante con el plásmido curado, empleando distintas fuentes de nitrógeno en el medio de cultivo.
Este trabajo fue presentado en el XVII Congreso Latinoamericano de Microbiología y X Congreso Argentino de Microbiología, realizado en la Ciudad Autónoma de Buenos Aires del 17 al 21 de octubre de 2004.
MATERIALES Y MÉTODOS
Microorganismos
Cepa BSc411. Mutante sensible 41S obtenido mediante el curado con Naranja de Acridina de la cepa BSc411 (1). Todas las cepas pertenecen al Cepario del Instituto de Biotecnología de la U.N.S.J.
Medios de cultivo
Conservación y multiplicación: YEPD (g l-1): extracto de levadura 10; peptona 20; glucosa 20 (agar 20 cuando es necesario) pH 4,5.
YCB – SO4 (NH4)2 (g l-1): Yeast- Carbon Base (Difco) 11,7, SO4(NH4)2 1.
YEPD – MB (g l-1): YEPD, Azul de Metileno 0,03.
Cultivo mixto en biorreactor
Las cepas de levadura se activaron en frascos Erlenmeyer de 250 ml de capacidad, conteniendo 50 ml de medio YEPD pH 4,5, hasta la fase exponencial de crecimiento, a una temperatura de 25 ºC, con agitación constante (200 rpm).
Las cepas se inocularon a una concentración final de 2x106 células ml-1 en un biorreactor de 1000 ml con 600 ml de medio YEPD, pH 4,6, con agitación continua (300 rpm), suministro de aire estéril (0,6 l min-1) y temperatura controlada a 25 ºC.
Se tomaron muestras cada 90 minutos, durante 720 min. La concentración de biomasa se determinó por recuento celular en cámara de Neubauer.
Los cultivos mixtos se llevaron a cabo con dos proporciones de inóculo: condición A: 50% Killer- 50% Sensible, condición B: 10% Killer -90% Sensible
Detección de actividad killer y sensible
Se empleó, con modificaciones, el sistema descripto por da Silva (4). Aproximadamente 105 células/ml de la cepa sensible (41S) se suspendieron en 20 ml de YEPD - agar MB tamponado a pH 4,5 con buffer citrato-fosfato. La cepa a determinar se inoculó sobre la superficie del medio agarizado, incubándoselo a 23 ºC durante 72 hs. La formación de halo de inhibición sobre la cepa sensible, implicó que la cepa problema poseía el fenotipo killer.
Determinación de las proporciones killer-sensible en los ensayos en biorreactor
Las muestras extraídas del biorreactor se incubaron en YEPD-agar en una dilución apropiada, durante 72 hs a dos temperaturas: 25 ºC y 37 ºC, esta última en función de inactivar la toxina y de esa forma evitar la infraestimación de células sensibles. Una vez desarrolladas se eligieron al azar 250 colonias que se transfirieron a YEPD-MB para determinar su fenotipo.
Análisis estadístico de los resultados
Test de Tukey (12) p<0,05. InfoStat/Estudiantil, versión 2,0.
RESULTADOS Y DISCUSIÓN
Si bien existen algunos antecedentes de investigaciones sobre estudios de cultivos mixtos K-S en condiciones de fermentación enológica (4, 7, 9, 11), son escasos los trabajos previos considerando cultivos mixtos K-S (14) en condición aeróbica, aunque su utilidad se hace evidente como modelo a aplicar en la primera fase de la fermentación enológica, donde la disponibilidad de oxígeno disuelto es mayor que en el resto del proceso. Es en esta etapa donde se define, en muchos casos, la prevalencia o no de la cepa killer sobre la sensible (14); entonces, resulta de vital importancia definir las características de la exclusión competitiva bajo dicha condición de cultivo. El empleo de mutantes con el plásmido curado permite comparaciones más precisas sobre la influencia de los distintos factores de cultivo en la relación entre las cepas killer y sensibles (8, 13).
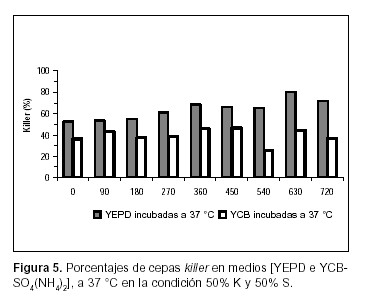
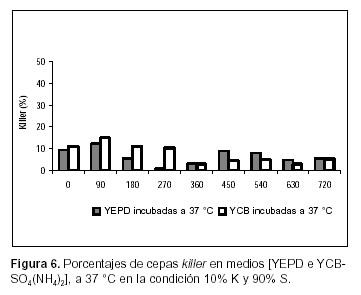
La evaluación del comportamiento de las poblaciones killer y sensibles de S. cerevisiae se ha visto restringida debido a la dificultad existente en la cuantificación de las diferentes cepas cuando están creciendo en cultivos mixtos. El repique de un número representativo de colonias para ser evaluadas, el uso de medios diferenciales y la resistencia a antibióticos (4, 13, 14), han sido algunas de las metodologías empleadas para tal fin. En todos los casos citados, la temperatura de incubación de la muestra fue de 18º a 28 ºC, la cual posibilita la expresión de la toxina killer en S. cerevisiae. En el presente estudio se sometió la muestra a una temperatura de inhibición de la acción de la toxina (37 ºC) en función de comparar los resultados con aquellos obtenidos a 25 ºC. Los datos obtenidos muestran diferencias significativas (Test de Tukey, p<0,05) (12) en cuanto a las células killer en todos los casos ensayados (Figuras 1, 2, 3, 4, 5 y 6).
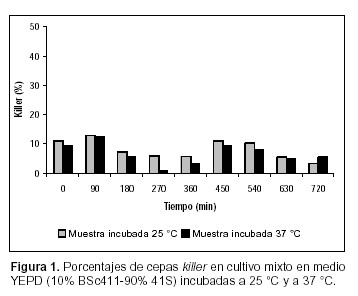
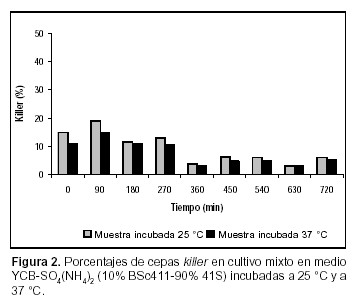

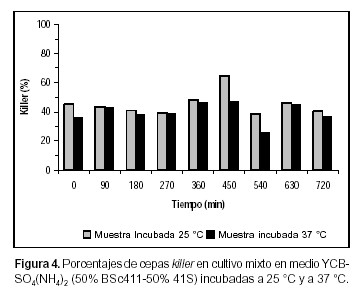
Esto se hace evidente incluso en la estimación de proporciones del inóculo. En las figuras 3 y 4 (condición A) se observa que la proporción de células killer detectadas cuando la muestra se incubó a 37 ºC fue entre 3,9% (YEPD) y 9,5% [YCB-SO4(NH4)2] menor que cuando se las cultivó a 25 ºC. En la condición B, las diferencias en el inóculo son de 1,5% en el ensayo realizado en YEPD (Figura 1) y de 4% cuando el cultivo se llevó a cabo en YCB-SO4(NH4)2. En el caso de los inóculos, dada la posibilidad de contrastar los datos porcentuales con el conteo celular inicial, es posible ver claramente la infraestimación de las células sensibles. Una posible explicación a estos resultados puede residir en que durante el desarrollo de las colonias, previo a la evaluación de su fenotipo, algunas células sensibles presentes en el biorreactor pueden ser eliminadas del medio de desarrollo de la muestra debido a la cercanía de células killer, lo cual puede conducir a una infraestimación del tamaño de la población sensible presente en el biorreactor. Este tipo de dato adquiere importancia cuando está referido a la eliminación completa de la cepa sensible.
En las Figuras 1 y 2 se muestra que en la condición de cultivo B, tanto en medio YEPD como en YCB-SO4(NH4)2, la cepa killer no incrementó en forma significativa su presencia en el medio, por lo tanto no se verificó la exclusión de la cepa sensible, ya sea cuando las muestras se incubaron a 25 ºC como a 37 ºC (Test de Tukey, p< 0.05) (12). Al co-cultivar ambas cepas en iguales proporciones en medio de cultivo YEPD, la cepa BSc411 desplazó a la mutante alcanzando el 79% del total de la biomasa producida (540 min) cuando las muestras fueron incubadas a 25 ºC. A 37 ºC, la cepa K alcanzó un valor similar a los 630 min de comenzado el experimento. El dominio del medio de cultivo ocurrió en el momento en que la cepa BSc411 se hallaba en fase logarítmica de multiplicación. Sin embargo, en YCB no se observa dicha prevalencia, manteniéndose en proporciones similares a las sembradas (Figuras 3 y 4).
Los resultados obtenidos en este estudio respecto de la expresión de la toxina killer en medios con distinta calidad y cantidad de fuente de nitrógeno muestran diferencias significativas al comparar las dos condiciones. En las Figuras 5 y 6 se observa que, cuando se incuba a 37 ºC y la fuente de nitrógeno, es suministrada por la peptona y el extracto de levadura (medio de cultivo YEPD), el porcentaje de BSc411 (K) en la condición A, durante todo el experimento (es evidente, incluso en la cuantificación del inóculo), y en la B durante los 450 min finales del ensayo, es significativamente mayor (Test de Tukey, p< 0.05) (12) que el encontrado cuando el experimento se realizó en YCB-SO4(NH4)2. Estos resultados son consistentes con los obtenidos por Medina K. et al. (11) comparando en condiciones de fermentación enológica, la abundancia de células killer en mosto fresco respecto de mosto adicionado con sales de amonio, donde se concluye que la interacción killer-sensible depende, además de la proporción de las distintas células en el inóculo, del tipo y concentración de fuente de nitrógeno en el inicio de la fermentación y probablemente de la demanda del mismo cuando las cepas están en competencia (11).
Los datos obtenidos en condición aeróbica en el presente estudio pueden relacionarse con los de la primera fase del proceso de vinificación, y muestran la influencia de la fuente de nitrógeno en la expresión del carácter killer.
En condiciones experimentales comparables, Ramón–Portugal F. et al. (14), encuentran que una proporción del 10% de cepa killer desplaza a la levadura sensible, en cultivos mixtos por lote. En la condición B (considerando los valores obtenidos con la muestra cultivada a 37 ºC) de este trabajo, cuando se utilizó YCB-SO4(NH4)2 como medio de cultivo, la proporción de BSc411 fluctuó entre el 11% (0 min del experimento) y 5,3% (720 min). De la misma forma, cuando el cultivo mixto se llevó a cabo en YEPD, la proporción de cepa killer varió desde el 9,5% al comienzo del experimento hasta el 5,6% al finalizar la experiencia. El comportamiento fluctuante respecto de la proporción de cepas killer en co-cultivos con células sensibles, registra antecedentes tanto en condiciones de cultivo aeróbicas como en procesos de vinificación (11, 14). En ambos casos, esta situación se produce cuando el inóculo es insuficiente para permitir el desarrollo de la cepa killer. En los experimentos llevados a cabo en condiciones enológicas (11), la imposibilidad de la cepa killer de ejercer su predominio se atribuye a que las células sensibles habrían contado con el tiempo suficiente para acumular el nitrógeno asimilable del medio de cultivo antes de que la máxima actividad killer pudiera expresarse. Este tipo de explicación podría también ser adecuada para los cultivos en condición aeróbica. Cabe destacar que en ambos medios de cultivo la proporción de BSc411 se redujo prácticamente a la mitad en el final del experimento, lo cual puede atribuirse a que la estabilidad de la toxina puede alterarse debido a cambios producidos en el pH durante el transcurso del ensayo.
Los resultados del experimento del co-cultivo de las cepas en proporciones iguales muestran el rol desempeñado por la fuente de nitrógeno en la actividad killer. Por otro lado, cuando el inóculo K es minoritario respecto del mutante sensible, el modelo de exclusión competitiva planteado para levaduras con carácter killer deja paso a otras variables de competencia.
BIBLIOGRAFÍA
1. Cabrera NE, Toro ME, Vázquez F (2001) Plásmido killer en levaduras de interés enológico. Curado mediante diferentes agentes mutagénicos. X Simposio Latinoamericano y VII Simposio Argentino de Farmacobotánica, Resumen 111, p. 86, Comodoro Rivadavia, Chubut, Argentina. [ Links ]
2. Cansado J, Longo E, Agrelo D, Villa TG (1989) Curing of the killer character of Saccharomyces cerevisiae with Acridine orange. FEMS Microbiol. Letters 65: 233-238. [ Links ]
3. Chen WB, Han YF, Jong SC, and Chang SC (2000). Isolation, purification, and characterization of a killer protein from Schwanniomyces occidentalis. Appl. Environm. Microbiol. 66: 5348-5352. [ Links ]
4. da Silva GA (1996) The occurrence of killer, sensitive, and neutral yeasts in Brasilian Riesling Italico grape must and the effect of neutral strains on killing behaviour. Appl. Microbiol. Biotechnol. 46: 112-121. [ Links ]
5. Fleet GH (2003) Yeast interactions and wine flavour. Int. J. Food Microbiol. 86: 11-22. [ Links ]
6. Flegelová H, Novotná D, Vojtíšková K, and Jandrová B (2002) Isolation and characterization of Saccharomyces cerevisiae mutants with a different degree of resistance to killer toxins K1 and K2. FEMS Yeast Research. 2: 73-79.
7. Franken DB, Ariatti M, Pretorius IS, Gupthar AS (1998) Genetic and fermentation properties of the K2 killer yeast, Saccharomyces cerevisiae T206. Antonie van Leeuwenhoek. 73: 263-269. [ Links ]
8. Longo E, Cansado J, Sieiro C, Calo P, Velázquez JB, Villa TG (1992) Influence of the curing of the killer phenotype in Saccharomyces cerevisiae wine strains on their fermentative behaviour. World J. Microbiol. Biotechnol. 8: 147-150. [ Links ]
9. Longo E, Velázquez JB, Cansado J, Calo P, Villa TG (1990) Role of killer effect in fermentations conducted by mixed cultures of Saccharomyces cerevisiae. FEMS Microbiol. Letters. 71: 331-336. [ Links ]
10. Mauricio JC, Valero E, Millán C, Ortega JM (2001) Changes in nitrogen compounds in must and wine during fermentation and biological aging by flor Yeasts. J. Agric. Food Chem. 49: 3310-3315. [ Links ]
11. Medina K, Carrau FM, Gioia O, Bracesco N (1997) Nitrogen availability of grape juice limits killer yeast growth and fermentation activity during mixed-culture fermentation with sensitive commercial yeast strains. Appl. Environm. Microbiol. 63: 2821-2825. [ Links ]
12. Miller JC, Miller JN (1988) Significance tests. En: Chalmers RA, Masson M. (Eds), Statistics for analytical chemistry, Ellis Horwood Limited, England, p. 53-80 [ Links ]
13. Pérez F, Ramírez M, Regodón JA (2001) Influence of killer strains of Saccharomyces cerevisiae on wine fermentation. Antonie van Leeuwenhoek. 79: 393-399. [ Links ]
14. Ramon-Portugal F, Delia M, Strehaiano P, Riba J (1998) Mixed culture of Killer and sensitive Saccharomyces cerevisiae strains in batch and continuous fementations. World J. Microbiol. Biotechnol. 14: 83-87. [ Links ]
15. Torrea-Goñi D, Ancín-Azpilicueta C (2002) Nitrogen metabolism in chardonnay musts inoculated with killer strains of Saccharomyces cerevisiae. J. Biosc. Bioengin. 94: 15-22. [ Links ]
16. Van Vuuren HJJ, Wingfield BD (1986) Killer yeasts, Cause of stuck fermentations in a wine cellar. S. Afric. J. Enol. Vitic. 7: 113-118. [ Links ]
17. Versavaud A, Courcoux P, Roulland C, Dulau L, Hallet JN (1995) Genetic diversity and geographical distribution of wild Saccharomyces cerevisiae strains from the wine-producing area of Charentes, France. Appl. Environm. Microbiol. 10: 3521-3529. [ Links ]
18. Vodkin M (1977) Induction of yeast killer factor mutations. J. Bacteriol. 132: 346-348. [ Links ]
19. Wickner RB (1974) "Killer character" of Saccharomyces cerevisiae: curing by growth at elevated temperature. J. Bacteriol. 117: 1356-1357. [ Links ]
20. Wickner RB (1996) Double-stranded RNA viruses of Saccharomyces cerevisiae. Microbiol. Rev. 60: 250-265. [ Links ]
21. Yap NA, de Barros Lopes M, Langridge P, Henschke PA (2000) The incidence of killer activity of non-Saccharomyces yeasts towards indigenous yeast species of grape must: potential application in wine fermentation. J. Appl. Microbiol. 89: 381-389. [ Links ]
Recibido: 9/11/04
Aceptado: 27/6/05

