










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista argentina de microbiología
versión impresa ISSN 0325-7541versión On-line ISSN 1851-7617
Rev. argent. microbiol. v.37 n.4 Ciudad Autónoma de Buenos Aires oct./dic. 2005
Caracterización fenotípica y genotípica de la resistencia enzimática a las cefalosporinas de tercera generación en Enterobacter spp.
E. Bertona1, M. Radice2*, C. H. Rodríguez3, C. Barberis3, C. Vay3, A. Famiglietti3, G. Gutkind2
1Carrera de Especialización en Bioquímica Clínica, área Bacteriología Clínica, Escuela de graduados, Junín 956 CP 1113.
2Cátedra de Microbiología, Facultad de Farmacia y Bioquímica, Universidad de Buenos Aires, Junín 956 CP 1113.
3Laboratorio de Bacteriología Clínica, Departamento de Bioquímica Clínica, Hospital de Clínicas José de San Martín, Facultad de Farmacia y Bioquímica, Universidad de Buenos Aires, Córdoba 2150 CP1113.
*Correspondencia. E-mail: mradice@ffyb.uba.ar
RESUMEN
Enterobacter spp. es un patógeno intrahospitalario que presenta múltiples mecanismos de resistencia a los antibióticos ß-lactámicos. Se caracterizaron fenotípica y genotípicamente las diferentes ß-lactamasas presentes en 27 aislamientos consecutivos e ininterrumpidos de Enterobacter spp. (25 Enterobacter cloacae y 2 Enterobacter aerogenes). También se evaluó la habilidad de diferentes métodos fenotípicos para detectar ß-lactamasas de espectro extendido (BLEE) en estos microorganismos. En 15/27 aislamientos (63%) se observó resistencia a las cefalosporinas de tercera generación. En 12 de los aislamientos resistentes se detectó un alto nivel de producción de cefalosporinasa cromosómica, siendo 6 de ellos también productores de PER-2. Dicha resistencia en los 3 aislamientos restantes se debió exclusivamente a la presencia de BLEE, PER-2 en 2 de ellos y CTX-M-2 en un caso. Sólo CTX-M-2 se detectó con todas las cefalosporinas probadas en los ensayos de sinergia, utilizando el método de difusión, mientras que cefepima mejoró la detección de PER-2 en 7/8 aislamientos productores de esta BLEE, 4/8 utilizando la prueba de doble disco y 7/8 comparando discos de cefepima con y sin el agregado de ácido clavulánico. El método de dilución empleado solo detectó 1/9 BLEE al comparar las cefalosporinas con y sin el agregado de inhibidor.
Palabras clave: Enterobacter spp., ß-lactamasas de espectro extendido, cefepima
SUMMARY
Phenotypic and genotypic characterization of resistance to third-generation cephalosporins in Enterobacter spp. Enterobacter spp. are becoming increasingly frequent nosocomial pathogens with multiple resistance mechanism to b-lactam antibiotics. We carried out the phenotypic and genotypic characterization of b-lactamases in 27 Enterobacter spp. (25 Enterobacter cloacae y 2 Enterobacter aerogenes), as well as the ability of different extended spectrum ß-lactamase (ESBL) screening methods. Resistance to third generation cephalosporins was observed in 15/27 (63%) isolates. Twelve resistant isolates produced high level chromosomal encoded AmpC ß-lactamase; 6 of them were also producers of PER-2. Resistance to third generation cephalosporins in the remaining 3 isolates was due to the presence of ESBLs, PER-2 in 2 cases, and CTX-M-2 in the other. Only CTX-M-2 production was detected with all tested cephalosporins using difusion synergy tests, while cefepime improved ESBLs detection in 7/8 PER-2 producers, 4/8 in the inhibitor aproximation test and 7/8 with double disk test using cefepime containing disk with and without clavulanic acid. Dilution method, including cephalosporins with and without the inhibitor detected 1/9 ESBLs producers.
Key words: Enterobacter spp., expanded-spectrum b-lactamases, cefepime
INTRODUCCIÓN
El género Enterobacter incluye 14 especies o biogrupos clasificados de acuerdo a características bioquímicas y similitudes genómicas (1). Mientras que E. cloacae y E. aerogenes son aislados habitualmente como patógenos intrahospitalarios, otras especies como Pantoeae agglomerans, Enterobacter sakazakii, Enterobacter tayloreae (actualmente Enterobacter cancerogenus), Enterobacter gergoviae, Enterobacter absuriae son patógenos humanos poco frecuentes (19). La resistencia a los antibióticos ß-lactámicos en este género, y en muchos miembros de la familia Enterobacteriaceae, usualmente es mediada por la producción de enzimas con capacidad de hidrolizar dichos antibióticos y probablemente por alteraciones en la permeabilidad de la membrana externa; y en ciertos casos, la combinación de estos mecanismos (9).
Enterobacter spp., igual que otras enterobacterias como Citrobacter freundii, Serratia spp., Morganella morganii, Providencia stuartii y Providencia rettgeri producen ß-lactamasas cromosómicas inducibles denominadas Amp-C, no inhibidas por ácido clavulánico y otros inhibidores de ß-lactamasas (9). Los microorganismos productores de estas enzimas son resistentes a las aminopenicilinas con y sin los inhibidores de ß-lactamasas y a las cefalosporinas de primera generación. La resistencia a cefoxitina varía entre las distintas especies y a las cefalosporinas de tercera generación depende del nivel de expresión de la enzima cromosómica. En algunos casos la hiperproducción de estas enzimas puede otorgar resistencia a dichas cefalosporinas (6).
Por otro lado, es notable la diseminación a nivel mundial de ß-lactamasas de espectro extendido (BLEE) en enterobacterias productoras de enzimas cromosómicas inducibles, mecanismo que ya fue descripto en nuestro medio a finales de los años 90 (14, 16). Estas ß-lactamasas confieren resistencia a las penicilinas, cefalosporinas de primera, segunda, tercera y cuarta generación y monobactámicos. Escapan a su acción las cefamicinas y los carbapenemes, y son fácilmente inhibidas por ácido clavulánico y otros inhibidores relacionados (5).
Distintos estudios realizados en nuestro país, han demostrado que las BLEEs prevalentes en la mayoría de las enterobacterias son CTX-M-2 y PER-2. Un bajo porcentaje corresponde a las ß-lactamasas derivadas de SHV, que junto con las derivadas de TEM, son las más comúnmente halladas en enterobacterias en Europa y EEUU (3-5, 7, 15).
El objetivo de este trabajo fue investigar los mecanismos enzimáticos responsables de la resistencia a antibióticos ß-lactámicos en aislamientos clínicos de Enterobacter spp., así como caracterizar las ß-lactamasas presentes y comparar diferentes métodos fenotípicos de detección de la presencia de BLEEs en este género.
MATERIALES Y MÉTODOS
Microorganismos
Se estudiaron 27 aislamientos únicos, sucesivos e ininterrumpidos de Enterobacter spp. recuperados de pacientes atendidos en el Hospital de Clínicas "José de San Martín", en el período de Junio a Diciembre del año 1999. La identificación de género y especie se realizó según la metodología convencional. Los microorganismos fueron conservados a -20 °C, en agar cerebro-corazón con glicerol al 20%.
Determinación de la susceptibilidad a antibióticos
La determinación de la concentración inhibitoria mínima (CIM) se realizó por el método de dilución en agar siguiendo las recomendaciones del NCCLS (actualmente CLSI) (11). Se incluyeron los siguientes antibióticos: amoxicilina (AMX), amoxicilina/ ác. clavulánico, (2:1) (AMC), piperacilina (PIP), piperacilina/ tazobactam (PTZ) (tazobactam en concentración constante de 4 µg/ml), cefalotina (CTN), cefoxitina (FOX), cefotaxima (CTX), ceftazidima (CAZ), cefoperazona (CPZ), cefepima (FEP), aztreonam (AZT), imipenem (IMI), meropenem (MER), ciprofloxacina (CIP) y gentamicina (GEN). Para la detección de microorganismos productores de BLEE se incluyeron CTX, CAZ y FEP combinados con concentraciones constantes de ácido clavulánico (2 µg/ml). Se confirmó fenotípicamente su presencia cuando la CIM de alguna de las cefalosporinas utilizadas con el inhibidor, presentó un descenso de por lo menos tres diluciones, con respecto a la cefalosporina sola.Detección fenotípica de la presencia de ß-lactamasas inducibles
Se realizó un ensayo de doble disco, empleando discos conteniendo FOX (30 µg) (Laboratorio Britania), ácido clavulánico (10 µg) y ácido 6-amino-penicilánico (6APA) (60 µg) colocados a 2 cm de distancia borde a borde de discos conteniendo CTX (100 µg), sobre placas de agar Mueller-Hinton inoculadas con 108 microorganismos/ml. Los discos conteniendo ácido clavulánico y 6APA fueron preparados en el laboratorio. La inducción fue interpretada como un achatamiento en el halo de inhibición del disco de CTX en la zona adyacente a los discos conteniendo los distintos inductores. Los discos conteniendo 100 µg de CTX fueron preparados en el laboratorio debido a que la mayoría de las cepas no presentaban halo de inhibición para este antibiótico empleando los discos comerciales de CTX (30 µg).
Detección de la presencia de BLEEs por difusión en agar
Se realizó el ensayo de detección de la presencia de BLEE en 15 aislamientos que resultaron resistentes a alguna de las cefalosporinas ensayadas. Se emplearon discos conteniendo CTX (30 µg) y CAZ (30 µg) solos, y con el agregado de 10 µg de ácido clavulánico, CTXC y CAZC, de acuerdo a las recomendaciones del NCCLS para la detección de BLEE en E. coli y Klebsiella spp. (12). Se incluyó en el ensayo los discos de FEP (30 µg) y FEP-ácido clavulánico (FEPC) (30 µg/10 µg), para evaluar su utilidad en la detección de BLEE en microorganismos productores de enzimas AmpC inducibles.
Se realizó también otro ensayo de sinergia, colocando discos de CTX (30 µg) y FEP (30 µg) a 20 mm de un disco de AMC (20/ 10 µg) (8).
La presencia de BLEEs fue interpretada como un aumento en el halo de inhibición de la cefalosporina asociada al inhibidor ≥ 5 mm con respecto a la cefalosporina sola, y/o observación de sinergia en el ensayo por aproximación de disco.
Determinación de los puntos isoeléctricos (pI) de las ß-lactamasas
Los extractos enzimáticos se obtuvieron a partir de cultivos de los microorganismos en 50 ml de caldo cerebro-corazón con 100 µg/ml de ampicilina, mediante ruptura ultrasónica en baño de hielo para evitar la desnaturalización proteica, utilizando un sonicador Vibra Cell (Sonics & Materials Inc, USA). El pI de las enzimas fue determinado por isoelectroenfoque analítico empleando geles de poliacrilamida de amplio rango (pH: 3-10) (10). Se empleó un equipo LKB 2117 Multiphor II (LKB Produkter AB, Suecia). Como patrones se incluyeron enzimas de pI conocido. Las ß-lactamasas fueron reveladas por el método iodométrico en agar utilizando ampicilina (500 µg/ml) (18). La inhibición de las enzimas se evaluó sumergiendo el gel en una solución 1mM de clavulanato de potasio y revelando posteriormente por el método iodométrico, empleando ampicilina 500 µg/ml como sustrato (14).
Detección molecular de los genes codificantes de las ß-lactamasas plasmídicas
De acuerdo a los resultados observados en los ensayos anteriores, se amplificaron los genes codificantes de las ß-lactamasas empleando distintos pares de cebadores (5´-3´) (GIBCO BRL, USA) blaTEM-1a: ATAAAATTCTTGAAGACGAAA, blaTEM1b: GACAGTTACCAAYGCTTAATCA, blaCTX-M-2 I:TTAAT-GATGACTCAGAGCATTC, blaCTX-M-2 II: GATACCTCGCTC-CATTTATTG, blaPER2-a: TGTGTTTTCACCGCTTCTGCTCTG, blaPER2-b: AGCTC-AAACTGATAAGCCGCTTG. Como molde se utilizó ADN plasmídico obtenido por lisis alcalina de las cepas de Enterobacter spp. resistentes a cefalosporinas de tercera generación. La reacción de amplificación consistió en desnaturalización a 95 °C durante 5 minutos, "annealing" a 55 °C por 15 minutos (a los 6 minutos se agregó la Taq polimerasa) y un período de extensión de 6 minutos a 72 °C; seguido de 25 ciclos de 1 minuto a 95 °C, 90 segundos a 55 °C y 90 segundos a 72 °C; y un período de extensión final de 20 minutos a 72 °C. Se empleó un termociclador Matercycler 5330 (Eppendorf, Alemania). Los fragmentos amplificados fueron secuenciados automáticamente en ambas cadenas empleando un secuenciador ABI PRISM 3700 DNA.RESULTADOS
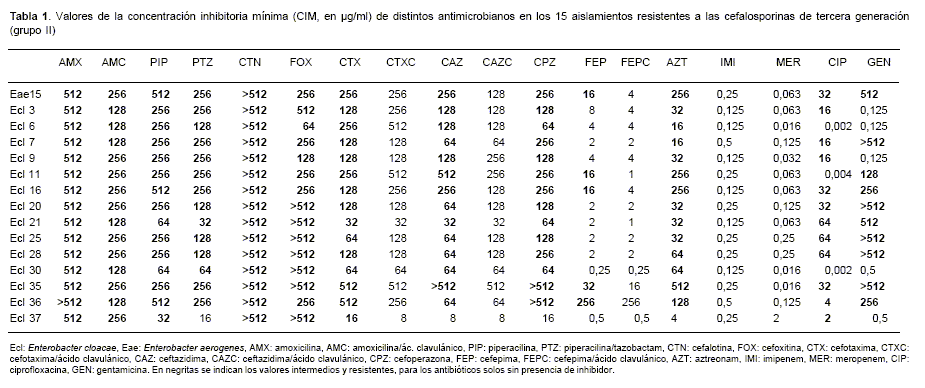
En el período analizado se recuperaron 27 aislamientos de Enterobacter spp., 25 correspondieron a E. cloacae y 2 a E. aerogenes. Todos ellos presentaron las resistencias naturales a AMX, AMC, CTN y FOX, y sensibilidad a los carbapenemes. Doce de los 27 aislamientos fueron sensibles a CTX (grupo I), mientras que 15 presentaron sensibilidad intermedia o fueron resistentes (grupo II). Los aislamientos del grupo I fueron sensibles a gentamicina y ciprofloxacina, excepto uno de ellos resistente a ciprofloxacina (datos no mostrados). Todos los microorganismos del grupo II, excepto uno, fueron también resistentes a CAZ, CPZ y AZT. Solo 5/15 presentaron sensibilidad disminuida o resistencia a FEP. Las resistencias asociadas en este grupo fueron: ciprofloxacina en 12/15 y gentamicina en 10/15. En la tabla 1 se observan los valores de CIM de los antibióticos ensayados para los aislamientos del grupo II.
Tabla 1. Valores de la concentración inhibitoria mínima (CIM, en µg/ml) de distintos antimicrobianos en los 15 aislamientos resistentes a las cefalosporinas de tercera generación (grupo II)
Cuando se analizaron los valores de CIM de CTX y CAZ en presencia de ácido clavulánico no se observó disminución de tres diluciones respecto de los valores obtenidos de la droga sola, no detectándose la presencia de BLEE por este método. La combinación FEP/ácido clavulánico permitió la detección de la presencia de BLEE en un solo caso (cepa Ecl 11, Tabla 1).
Mediante el ensayo de inducción realizado en los 15 aislamientos pertenecientes al grupo II, ésta fue detectada en 7 de ellos, mientras que en el resto la metodología fue insuficiente para poder diferenciar entre un mayor nivel de expresión de las enzimas cromosómicas y la derrepresión total de las mismas.
Utilizando el método de difusión propuesto por el NCCLS para Escherichia coli, Klebsiella spp. y Proteus mirabilis para la detección de BLEE, se observó aumento del halo de inhibición en 1/15 aislamientos del grupo II empleando CTX o CAZ con el agregado de ácido clavulánico. En cambio, con FEPC se incrementó la detección en 8/15 aislamientos. Del mismo modo, cuando se empleó la prueba de doble disco se observó sinergia sólo en 1 de 15 aislamientos del grupo II enfrentando CTX a AMC y en 5 de 15 enfrentando FEP a AMC. En la tabla 2 se observan los resultados de los distintos ensayos empleados para la detección de BLEE en los microorganismos del grupo II.
Tabla 2. Caracterización fenotípica y genotípica de las ß-lactamasas presentes en los 15 aislamientos resistentes a las cefalosporinas de tercera generación (grupo II)
Todos los extractos enzimáticos excepto uno, presentaron bandas detectables en el isoelectroenfoque. Probablemente en dicho caso la actividad enzimática fuera menor al límite de detección del método iodométrico empleado en el revelado. El resto presentó enzimas de pI 5,4 y/o 8,2 activas frente a ampicilina 500 µg/ml. Todas las enzimas de pI 5,4 y sólo una de las de pI 8,2 fueron inhibidas por clavulanato. En el resto de los extractos la enzima de pI 8,2 no fue inhibida lo que sugiere la presencia de una enzima de tipo AmpC.
En la Tabla 2 se observan, también, los resultados de la caracterización fenotípica y genotípica de las ß-lactamasas presentes en los microorganismos del grupo II.
DISCUSIÓN
La resistencia a las cefalosporinas de tercera generación en los aislamientos de Enterobacter incluidos en este trabajo (15/27) fue similar a los niveles de resistencia informados por el Sistema Informático de Resistencia (SIR) donde se observó aproximadamente un 60% de resistencia a CTX y CAZ en el año 2000 (2). Igual a lo observado en otras enterobacterias, la resistencia asociada a antibióticos no ß-lactámicos es más frecuente en aquellas cepas resistentes a cefalosporinas de tercera generación que en las sensibles (17). La frecuencia de aislamiento de las distintas especies del género Enterobacter se vio representada en la muestra analizada (Informe SIR 2001, www.aam.org.ar).
Los 15 aislamientos resistentes a cefalosporinas de tercera generación correspondieron al grupo II. En 6 aislamientos de E. cloacae pertenecientes a dicho grupo se observó un alto nivel de expresión de la ß-lactamasa cromosómica de pI 8,2 no inhibible por ácido clavulánico, capaz de conferir resistencia por sí misma a las cefalosporinas de tercera generación. En este estudio no se evaluó la participación de posibles alteraciones en la permeabilidad que contribuyan a la resistencia a dichos antibióticos. Algunos de ellos fueron a su vez, productores de la enzima TEM-1. El ensayo de inducción sólo fue positivo en dos de estos aislamientos no detectándose en el resto si correspondieron a altos niveles de expresión de la enzima o a mutantes totalmente derreprimidas. En este subgrupo la CIM de FEP osciló entre 0,25-16 µg/ ml. En otros 6 aislamientos (5 E. cloacae y 1 E. aerogenes) en los que se observó alto nivel de producción de AmpC, también se detectó la presencia de PER-2. Esto coincide con comunicaciones nacionales previas en cuanto al tipo de BLEE predominante en Enterobacter spp. (13). En estos aislamientos, la CIM de FEP mostró un rango de 2-16 µg/ml, coincidiendo con los valores esperados para cepas productoras de esta enzima. En los tres aislamientos restantes del grupo II el mecanismo de resistencia a las cefalosporinas de tercera generación fue la producción de BLEE, que correspondió a PER-2 en dos casos y CTX-M-2 en el restante. Esta última enzima fue detectada con todos los métodos de difusión confirmatorios de BLEE, mientras que PER-2 fue detectada en 7/8 aislamientos con el ensayo de difusión empleando FEP/FEPC.
La baja sensibilidad de las cefalosporinas de 3ª generación se debería al balance entre el efecto inductor del ácido clavulánico sobre la expresión de la enzima cromosómica inducible, y la inhibición de la BLEE. El efecto final dependerá del balance entre ambas actividades. Como fue observado por otros autores (13, 14) y recomendado por la Sociedad Francesa de Microbiología (20), la utilización de FEP mejora notablemente la detección de las BLEE en Enterobacter spp. Los métodos de difusión ensayados para detectar la presencia de BLEE, fueron superiores al método de dilución. Esta diferencia probablemente se deba a la menor concentración de ácido clavulánico utilizada en la CIM. La concentración constante de 2 µg/ml versus 4 µg/ml recomendada por NCCLS (11) se eligió en función de trabajos anteriores (21) y considerando que una menor concentración provocaría menor efecto inductor en la ß-lactamasa cromosómica.
En el género Enterobacter la hiperproducción de AmpC resultó el mecanismo principal de resistencia a las cefalosporinas de tercera generación, asociada en muchos casos con la producción de PER-2. Si bien la presencia de BLEE fue detectada en 1 solo caso por los métodos propuestos por el NCCLS para E. coli, Klebsiella spp. y P. mirabilis, la incorporación de FEP/FEPC incrementó sensiblemente su detección. Discos de FEP y FEPC podrían utilizarse para detectar BLEE en aislamientos de Enterobacter spp. y probablemente en otras enterobacterias con AmpC inducibles. La importancia de detectar la presencia de BLEE en aislamientos sensibles a FEP, radica en que esta cefalosporina no debería indicarse cuando se detecta la producción de BLEE, aunque un ensayo negativo no descartaría su presencia.
Agradecimientos: Este trabajo fue financiado en parte por los subsidios ANPCyT, BID 1201/ OC-AR PICT 6931 a Gabriel Gutkind y Fundación Roemmers a Marcela Radice.
BIBLIOGRAFÍA1. Abbot S (1999) Klebsiella, Enterobacter, Citrobacter, and Berratia. En: Murray P, Baron E, Pfaller M, Tenover F, Yolken (Ed), Manual of Clinical Microbiology. 7th edition. ASM Press, Washington, Estados Unidos de América, p. 475-482. [ Links ]
2. Bantar C, Famiglietti A, Goldberg M, Committee TA, Group. TNSPSP (2000) Three-year surveillance study of nosocomial bacterial resistance in Argentina. Int. J. Infect. Dis. 4: 85-90. [ Links ]
3. Bauernfeind A, Stemplinger I, Jungwirth R, Mangold P, Amann S, Akalin E, et al. (1996) Characterization of ß-lactamase gene blaPER-2 which encodes an extended-spectrum class A b-lactamase. Antimicrob Agents Chemother. 40: 616-620. [ Links ]
4. Bauernfeind A, Stemplinger Y, Jungwirth R, Ernst S, Casellas JM (1996) Sequences of b-lactamase genes encoding CTX-M-1 (MEN-1) and CTX-M-2 and relationship of their amino acid sequences with those of other b-lactamases. Antimicrob. Agents Chemother. 40: 509-513. [ Links ]
5. Bradford PA (2001) Extended-spectrum b-lactamases in the 21stcentury: Characterization, epidemiology and detection of this important resistance threat. Clin. Microbiol. Rev. 14: 933-951. [ Links ]
6. Famiglietti A, Quinteros M, Vázquez M, Marín M, Nicola F, Radice M, et al. (2005) Consenso sobre las pruebas de sensibilidad a los antimicrobianos en Enterobacteriaceae. Rev. Argent. Microbiol. 37: 57-66. [ Links ]
7. Galas M, Rapoport M, Pasterán F, Melano R, Petroni A, Ceriana P, et al. (1999) High distribution of CTX-M-2 beta-lactamases among Klesiella spp. isolates in argentinean extended-spectrum beta-lactamase (esbla) surveillance program. 39 th. Interscience Conference on Antimicrobial Agents and Chemotherapy (ICCAC) Abstract 1474, pag. 165, San Francisco, California, USA. [ Links ]
8. Jarlier V, Nicolas MH, Fournier G, Philippon A (1988) Extended broad-spectrum b-lactamases conferring transferable resistance to newer ß-lactam agents in Enterobacteriaceae: Hospital prevalence and susceptibility patterns. Rev. Infect. Dis. 10: 867-878. [ Links ]
9. Livermore DM (1995) b-lactamases in laboratory and clinical resistance. Clin. Microbiol. Rev. 8: 557-584. [ Links ]
10. Matthew M, Harris AM, Marshall MJ, Ross GW (1975) The use of analytical isoelectric focusing for detection and identification of b-lactamases. J. General Microbiol. 88: 169-178. [ Links ]
11. National Committee for Clinical Laboratory Standards (2004) Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically. Sixth edition, Approved standard, Wayne, Pa., Estados Unidos de América . [ Links ]
12. National Committee for Clinical Laboratory Standards (2004) Performance standards for antimicrobial disk susceptibility tests. Eight edition, Approved standard, Wayne, Pa., Estados Unidos de América. [ Links ]
13. Pasteran F, Melano R, Galas M (1999) High proportion of extended spectrum beta-lactamases (esbla) among Amp-C producers enterobacteria in Argentina. 39 th. Interscience Conference on Antimicrobial Agents and Chemotherapy (ICCAC) Abstract 1475, p. 166, San Francisco, California, USA. [ Links ]
14. Power P, Radice M, Barberis C, de Mier C, Mollerach M, Maltagliatti M, et al. (1999) Cefotaxime-hydrolysing b-lactamases in Morganella morganii. Eur. J. Clin. Microbiol. Infect. Dis. 18: 743-747. [ Links ]
15. Quinteros M, Radice M, Gardella N, Rodríguez MM, Costa N, Korbenfeld D, et al. (2003) Extended-spectrum b-lactamases in Enterobacteriaceae in Buenos Aires, Argentina, public hospitals. Antimicrob. Agents Chemother. 47: 2864-2869. [ Links ]
16. Quinteros M, Radice M, Power P, Matteo M, Mollerach M, Conza JD, et al. (1999) Detection of extended-spectrum b-lactamases in microorganisms harboring an AmpC ß-lactamase. International Congress on b-lactamases, Abstract A27, L'Aquila, Italia. [ Links ]
17. Rodriguez C, Juárez J, de Mier C, Pugliese L, Blanco G, Vay C, et al. (2003) Resistencia a antibióticos en gram negativos aislados en unidades de cuidados intensivos. Análisis comparativo de dos períodos (1998-2001) Medicina. 63: 21-27. [ Links ]
18. Rossi A, Lopardo H, Woloj M, Picandet AM, Mariño M, Galas M, et al. (1995) Non-typhoid Salmonella spp. resistant to cefotaxime. J. Antimicrob. Chemother. 36: 697-702. [ Links ]
19. Sanders E, Sanders C (1997) Enterobacter: a pathogen poised to flourish at the turn of the century. Clin. Microbiol. Rev. 10: 220-241 [ Links ]
20. Soussy C, Carret G, Cavallo J-D, Chardon H, Chidiac C (2000) Antibiogram committee of the french microbiologysociety. Report 2000-2001. Pathol. Biol. 48: 832-871. [ Links ]
21. Tzelepi E, Giakkoupi P, Sofianou D, Loukova V, Kemeroglou A, Tsakris A (2000) Detection of extended-spectrum b-lactamases in clinical isolates of Enterobacter cloacae and Enterobacter aerogenes. J. Clin. Microbiol. 38: 542-546. [ Links ]
Recibido: 08/08/05
Aceptado: 31/10/05


{kind=link}