











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
La necesidad de la aplicación de prácticas agrícolas sustentables ha llevado al desarrollo de bioinsumos en base a microorganismos capaces de promover el crecimiento de las plantas (Plant Growth Promoting Rhizobacteria) con el fin de aumentar el rendimiento en los cultivos20. En los últimos años el mercado mundial de bioinsumos ha crecido con una tasa sostenida cercana al 10% y con una expectativa de alcanzar los 4.092 millones de dólares en el 202518. Dentro de las Plant Growth Promoting Rhizobacteria uno de los géneros mejor caracterizados es Azospirillum, siendo Azospirillum brasilense una de las especies más estudiadas y ampliamente utilizada en la producción de formulados biológicos11,26. A lo largo de los anos, el género Azospirillum ha demostrado ser capaz de promover el crecimiento y desarrollo de numerosas especies vegetales de interés agrícola a través de numerosos mecanismos fisiológicos o bioquímicos4. Entre dichos mecanismos se incluyen, la fijación biológica de nitrógeno, que tuvo menos relevancia agronómica de la inicialmente esperada, o la producción de fitohormonas y otros reguladores del crecimiento de plantas, que ha tenido mayor importancia en los últimos anños4,5. En Sudamérica, existen alrededor de 106 productos comerciales formulados en base a A. brasilense y aproximadamente el 90% de estos productos son desarrollados en nuestro país, siendo la cepa Az39 de A. brasilense el principio activo más frecuentemente utilizado en América de Sur, con un 75% de prevalencia en el mercado nacional y brasilero11. La cepa Az39, fue aislada en el Instituto Nacional de Tecnología Agropecuaria-Instituto de Microbiología y Zoología Agrícola en la década del 80, mediante un programa de identificación y selección de cepas de Azospirillum spp. de la superficie de raíces esterilizadas de plantas de trigo en la localidad de Marcos Juárez en la provincia de Córdoba. Este programa, además, realizó la evaluación de la capacidad promotora del crecimiento de Az39 y otras cepas del mismo género en condiciones agronómicas, siendo la primera la más eficiente. Como consecuencia, el Servicio Nacional de Sanidad y Calidad Agroalimentaria, junto con las empresas productoras de inoculantes decidieron recomendarla inicialmente para la producción de bioinsumos destinados al tratamiento biológico de semillas de maíz, trigo, sorgo y otras especies de no leguminosas10,33 y a la fecha para más de 16 especies vegetales dentro de las que se incluyen las mencionadas y otras como algodón, tabaco, cebada, arroz y soja en coinoculación con Bradyrhizobium japonicum. Después de más de 30 años desde su aislamiento y de convertirse en una de las cepas más estudiadas in planta como a campo, el laboratorio de Fisiología Vegetal y de la Interacción Planta Microorganismo de la Universidad Nacional de Río Cuarto en el año 2012 inició un ambicioso proyecto para secuenciar el genoma de A. brasilense Az39, lo que culminó exitosamente con la obtención de la secuencia completa y cerrada del mismo en el año 201432. El genoma de Az39 consiste de 6.311 secuencias codificantes distribuidas en 6 replicones (un cromosoma, 3 crómidos y 2 plásmidos). La obtención de la secuencia del genoma fue considerada como la base para el desarrollo de numerosos estudios posgenómicos dentro de los que se incluyen la generación de herramientas moleculares para estudiar con mayor precisión y profundidad la capacidad de esta bacteria para promover el crecimiento vegetal.
Sin embargo, como se mencionó anteriormente, a pesar de que el uso de bioinsumos y dentro de ellos aquellos formulados con esta cepa, aumenta año a año, no hay normativas gubernamentales estrictas que regulen la calidad y trazabilidad de los productos en el sistema agroalimentario22. Más aun, en la República Argentina, no existen protocolos que permitan autenticar la identidad de la cepa Az39 de A. brasilense en formulaciones comercializadas para diferentes tipos de cultivo. Hasta el momento, la identificación de este microorganismo se realiza mediante metodologías de microbiología clásica, pero estas no son suficientes para asegurar con certeza la identidad del microorganismo a nivel de cepa10. Algunos autores han avanzado en este sentido y han desarrollado metodologías que permiten la identificación a nivel de cepas dentro de una misma especie12,13,28. De este modo, el desarrollo de herramientas moleculares basadas en la secuencia del ADN, permiten establecer la identidad de ciertos microorganismos de manera confiable y precisa. La utilización de marcadores moleculares del tipo Sequence Characterized Amplified Regions (SCAR) es una de las alternativas con la que se ha conseguido la identificación molecular a nivel de cepa. Los marcadores SCAR representan loci únicos, genéticamente definidos, que se identifican mediante amplificación por PCR usando fragmentos de ADN genómico con cebadores específicos. Teniendo en cuenta las consideraciones anteriores, el presente trabajo tuvo como objetivo emplear estas herramientas moleculares para desarrollar una metodología simple, rápida y certera que permita la identificación cepa-específica de A. brasilense Az39 en cultivos puros o mezclas de microorganismos.
Materiales y métodos Material biológico Los microorganismos utilizados en este trabajo se resumen en la tabla 1. Los cultivos bacterianos pertenecientes al género Azospirillum se obtuvieron a partir de colonias puras cultivadas a 30 °C o 37°C (según la cepa evaluada), durante 72 h en el medio de cultivo Luria Bertani (LB) modificado por la adición de solución de rojo Congo (LBRC)24. Las colonias se repicaron en tubos de ensayo conteniendo 5 ml de caldo LB8. Las cepas Pseudomonasprotegens CHAO y Bacillus sub-tilis BH8, cultivadas en caldo LB y aisladas en agar LB, se utilizaron como control negativo. El medio EMA40 se utilizó para el crecimiento de B. japonicum E109 y se determinó su pureza por ausencia de crecimiento en medio agar tripticasa soya1 2.
Análisis genotípico in silico y desarrollo de marcadores del tipo SCARPara el análisis in silico se utilizó el software http://insilico.ehu.eus/9. En primer lugar, se cargó la secuencia genómica de la cepa Az39 fraccionada en tres archivos, uno conteniendo el cromosoma, y otros dos con los plásmidosy crómidos, respectivamente. Luego, se simularon múltiples Amplified Fragment Length Polymorphism (AFLP-PCR) utilizando diferentes combinaciones enzimáticas y nucleótidos de selección, ordenados al azar. A continuación, se seleccionaron secuencias de ADN con longitudes de entre 200-800 pares de bases (pb). Estas secuencias fueron.
Extracción de ADN
La extracción del ADN genómico se realizó a partir de colonias aisladas o cultivos puros de Az39 empleando alternativamente dos metodologías: (1) el kit comercial Easy Pure Bacteria Genomic ADN de la firma Transgen Inc. (China), de acuerdo a las especificaciones del fabricante y (2) la resina de intercambio iónico Chelex®100, según la metodología propuesta por Alippi et al.2. En el caso de las colonias aisladas, la extracción solo se realizó por el método de resina y para ello las colonias fueron resuspendidas en 300 l de solución fisiológica estéril (SF) (8,5mg/ml de NaCl); mientras que en el caso de los cultivos se utilizaron ambos métodos y para ello se partió de un volumen de 1 ml de muestra que se centrifugó durante 10min a 10 000 rpm y cuyo pellet se resuspendió en 300 l de SF estéril. En el caso de la extracción con resina, los pellets fueron resuspendidos en 150 ^l de una solución de Chelex®100 al 6% (p/v) y las muestras se incubaron en un baño termostático a 56 °C durante 10min seguido de una incubación a 99°C durante 8min, sometiendo las muestras a agitación vigorosa entre cada período de incubación. Finalmente, se realizó un último paso de centrifugado durante 10min a 10 000 rpm. El sobrenadante se utilizó inmediatamente como molde para la realización de las reacciones de PCR o fue fraccionado y conservado a -20°C hasta su utilización.
Determinación de la temperatura óptima de hibridación de los cebadoresPara conocer la temperatura óptima de hibridación o annealing (Ta) para la reacción, se realizó una PCR a tiempo final con gradiente de temperatura, probando diferentes Ta en un rango de 5 °C. Como molde se utilizó el ADN genómico de A. brasilense Az39. La mezcla de reacción consistió en un volumen final de 20 ^l conteniendo 1 unidad de EasyTaq ADN polimerasa (TransGen Biotech, China); EasyTaq buffer 1X (TransGen Biotech, China); 0,4 mM de dNTPs (Promega, Madision, Wis.); 0,4 ^M de cada primer (IDT Technologies) y 1 ^l de ADN molde. Las condiciones de amplificación fueron las siguientes: una desnaturalización inicial de 2 min a 95 °C, seguida de 30 ciclos de desnaturalización por 0,5 min a 95 °C; hibridación de 0,5 min desde 55,2 °C hasta 61,4 °C y extensión de 0,5 min a 72 °C, con una extensión final de 5 min a 72 °C. La amplificación se realizó en un termociclador Ivema T21 (Ivema Desarrollos). Los productos de PCR fueron revelados mediante electroforesis en gel de agarosa 1% (p/v) teñnidos con Bromuro de Etidio (BrEt) y visualizados con un transiluminador UV (Maestrogen, MLB-16 124, Taiwan).
Determinación de la especificidadPara evaluar la especificidad de los cebadores diseñnados previamente, se utilizó como muestra el ADN genómico de diferentes cepas de A. brasilense, así como el ADN de otras especies pertenecientes al género y microorganismos distanciados filogenéticamente (tabla 1). Las condiciones de reacción de PCR y los ciclos de amplificación empleados se realizaron según lo descrito anteriormente, variando la Ta de acuerdo a los resultados óptimos obtenidos. Con los microorganismos pertenecientes al género Azospirillum descritos en la tabla 1, se realizó la amplificación por PCR con cebadores específicos para el gen 16S rRNA según Lin et al.21 a modo de control para confirmar la identidad de las cepas empleadas en el análisis. En el caso de los microorganismos distanciados filogenéticamente, se utilizaron los cebadores BOX-AR1 según Versalovic et al.39.
Determinación de la sensibilidad del método para evaluar la sensibilidad del método, se utilizó ADN genómico como molde para la amplificación por PCR, el cual fue obtenido alternativamente por extracción con un kit comercial o por resina de intercambio iónico, como se describió previamente. A modo de resumen, 1 ml de un cultivo puro de A. brasilense Az39 fue diluido por duplicado en tubos conteniendo 9 ml de solución fisiológica estéril para obtener tubos con una dilución inicial de 10-1 a partir de los que se realizaron diluciones decimales hasta alcanzar un valor de 10-7. A continuación, se tomaron 20 ^l de cada dilución y se realizó un recuento por la técnica de microgota31 para confirmar el número de células viables. Del mismo tubo, se tomó 1 ml para la extracción con resina. El segundo tubo fue utilizado para la extracción de ADN mediante kit y la cuan-tificación de la concentración de ADN por la utilización de un NanoDrop One de la firma Thermo Fischer Inc. (EE. UU.). La determinación se realizó sobre el total de células (UFC/ml) obtenidas por centrifugación de cada dilución. Adicionalmente, se midió la concentración de ADN de diluciones en base decimal y centesimal del ADN obtenido a partir de una dilución 10-7 equivalente a un valor teórico de 1x102 UFC/ml. En todos los casos se utilizó como control negativo agua MilliQ esterilizada.
Determinación de la especificidad en cultivos mixtos con B. japonicum E109Cultivos mixtos de A. brasilense Az39 y B. japonicum E109, se obtuvieron independientemente en medios LB y EMA, hasta alcanzar una DO595 aprox. 1,9. Ambos cultivos se mezclaron en tubos plásticos de 1,5 ml de capacidad en diferentes proporciones de cada microorganismo (p. ej. 100% de E109; 70% E109 + 30% Az39; 50% E109 + 50% Az39; 30% E109 + 70% Az39 y 100% Az39). De cada una de las mezclas, se extrajo el ADN genómico mediante la metodología detallada anteriormente para cultivos en medio líquido. El ADN obtenido se utilizó como molde para reacciones de PCR específicas con los cebadores disenados en este trabajo (denominados TP5).
Determinación de la especificidad en cultivos mixtos con más de una cepa de AzospirillumSe realizaron tres mezclas diferentes de cultivos bacterianos pertenecientes al género Azospirillum, combinados en igual proporción. En el caso de la mezcla 1, se emplearon las cepas de la misma especie A. brasilense Az39, Sp7 y Sp245. Por otro lado, en la mezcla 2 se utilizaron las cepas A. brasilense Az39, Sp7 y Sp245, A. zeae N7, A. meli-nis TMYC 0552, A. halopraeferens Au4, A. canadense DS2 y A. rugosum IMMIB AFH-6 pertenecientes a distintas especies del género. Finalmente, en la mezcla 3 se utilizaron A. brasilense Az39, A. formosense CC-Nfb-7, A. fermentarium CC-LY743, A. oryzae COC8, A. palatum WW10, A. lipofe-rum Sp59b, A. agricola CC-HIH038 y A. soli CC-Ly788. Todas las cepas empleadas en estos experimentos se cultivaron en medio MMAB38 durante 24 h. Se tomó 1 ml de cada cultivo, se centrifugó a 10 000rpm durante 10 min y se ajustaron a igual DO595, con el objetivo de que todas las cepas tuvieran igual concentración (aprox. 1 x109 UFC/ml). Las mezclas se realizaron en microtubos de 1,5 ml a volumen final 1 ml, colocando iguales proporciones de los diferentes cultivos. Luego, a partir de cada mezcla bacteriana se extrajo el ADN con Chelex®100 según la metodología descrita anteriormente. Finalmente, el ADN genómico de las mezclas fue utilizado como molde para PCR con los cebadores específicos de Az39. Como control negativo se utilizaron mezclas sin Az39.
Control de calidad de formulaciones comercialesSe emplearon tres productos comerciales recomendados para el tratamiento de semillas de trigo y maíz, que en su membrete indicaban contener como principio activo A. brasilense Az39, a los que denominamos aleatoriamente I1, I2 e I3. En el caso del producto I1 declaraba una fecha de vencimiento tres meses posteriores a la fecha del análisis, en el caso de I2 se encontraba al límite de la fecha de vencimiento y en el caso de I3 había vencido 6 meses antes de realizar este análisis. A modo de aclaración, es necesario mencionar que, en Argentina, el valor de recuento límite a la fecha de vencimiento para productos comerciales conteniendo Azospirillum es de 1x107 UFC.ml-1. Para la toma de muestras, se homogenizaron las vejigas con el inoculante durante 10 min, se esterilizaron las superficies y se tomaron 2 ml de cada muestra para la posterior extracción de ADN por el método de Chelex®100. Adicionalmente, se tomó 1 ml para el recuento. Previo a la extracción de ADN, se realizó un lavado con solución fisiológica y las células se concentraron en un volumen final de 300 ^l. Luego se prosiguió con la metodología descrita previamente con la resina Chelex®100. El recuento se realizó por la técnica de microgota31 y para ello las muestras fueron colocadas en 9 ml de SF estéril para obtener la dilución 10-1 de la que se realizaron diluciones decimales para alcanzar una dilución de 10-7. A continuación, se tomaron 20 ^l de cada dilución, se sembraron en placas de LBRC y se incubaron a 37 °C durante 72 h. Se realizó el recuento de colonias y se calculó el número de UFC/ml. Adicionalmente, las muestras se sembraron en placas de LBRC en estrías por agotamiento y a partir de las colonias crecidas en este medio, se extrajo ADN con la resina Chelex®100 y se utilizó como molde para la amplificación por PCR con los cebadores TP5 a modo de confirmar la identidad de las colonias obtenidas.
Resultados
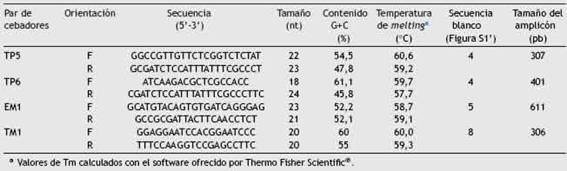
Análisis genotípico in silico y desarrollo de marcadores moleculares del tipo SCARMediante la simulación de 400 AFLP-PCR in silico, se seleccionaron 700 secuencias candidatas con un tamaño entre 200 y 800 pb. Cuando estas secuencias fueron comparadas mediante BLASTn, solo 12 de ellas no presentaron homología con secuencias depositadas en la base de datos o exhibieron fragmentos de al menos 200 pb únicos en el genoma de A. brasilense Az39 (fig. S1). Las secuencias únicas en el genoma de Az39 se localizaron en el cromosoma de dicha bacteria, algunas de ellas ubicadas en regiones no codificantes como en el caso de las secuencias 6, 7, 9 y 10 de la figura S2; mientras que otras se encontraron en regiones de secuencias que codifican para proteínas hipotéticas. Específicamente, en el caso de los productos de los cebadores evaluados in vitro en este trabajo TP5, TP6, TM 1 y EM1, todos se encontraron dentro de regiones que codifican para proteínas hipotéticas (fig. S2). Un total de 18 pares de cebadores fueron disenñados para la amplificación de las regiones no homólogas. Se observó que todos los cebadores fueron capaces de formar estructuras como homodímeros, heterodímeros y bucles dentro de la misma molécula de ADN, sin embargo, se escogieron 4 pares de cebadores cuyas características termodinámicas demostraron que las posibles estructuras secundarias eran inestables. Adicionalmente, cuando se simularon reacciones de PCR en los genomas de A. brasilense Sp45, A. lipoferum 4B, Azospirillum sp. B510 y Azospirillum spp. el resultado fue negativo. Más aún, no se observó amplificación en otras especies de Azospirillum, ni en otros géneros bacterianos presentes comúnmente en el suelo, entre ellos Bradyrhizo-bium, Pseudomonas, Bacillus, Rhodospirillum, Rhizobium, Sinorhizobium o Nitrosomonas. Las secuencias de los oligo-nucleótidos sintetizadas y probadas in vitro se muestran en la tabla 2 y estas amplifican las regiones número 4, 5 y 8 de la figura S1 y de la figura S3.
Análisis in vitro de los cebadores específicos para Az39Los 4 pares de cebadores disenñados fueron capaces de amplificar en todas las temperaturas evaluadas (fig. S4). En la figura S4A se observa que el par TP5 amplificó un fragmento de 307 pb; mientras que los pares de cebadores denominados TP6 y TM1 originaron amplicones de 401 pb y 306 pb respectivamente (fig. S4B). La amplificación con el par de cebadores denominado EM1 dio como resultado un amplicón esperado de 611 pb y una banda adicional que corresponde a un fragmento entre 100 pb y 200 pb (fig. S4C). Ambas bandas se observaron solo en la cepa Az39, inclusive cuando la Ta fue de 65 °C (datos no mostrados). En función de los resultados obtenidos en esta sección, se seleccionaron las siguientes Ta: 59,1 °C; 55,2 °C y 56,4°C para cada par de cebadores TP5, TP6 y TM1 respectivamente. Debido a que el par de cebadores EM1 era capaz de amplificar en regiones inespecíficas no fue utilizado para los ensayos posteriores de este trabajo.
Determinación de la especificidad y sensibilidad del métodoEn la figura 1 se muestran los resultados obtenidos en la evaluación de la especificidad del par de cebadores denominados TP5 en cultivos puros. El ciclo de reacción se llevó a cabo con una temperatura de alineamiento de 59,1 °C. Cuando se utilizó como molde el ADN genómico de cepas de A. brasilense filogenéticamente relacionadas con Az39, los resultados fueron negativos (fig. 1A), lo que demuestra el alto grado de especificidad de los cebadores diseñados. Las cepas evaluadas fueron A. brasilense Sp245, FP2, Abv5 y Abv6 (utilizadas en la producción de inoculantes en Brasil); Az 19, Az 36, Az 45, Az 48, Az 63 y Az 65 (pertenecientes a la colección del Instituto Nacional de Tecnología Agropecuaria-Instituto de Microbiología y Zoología Agrícola). Tampoco se observaron productos de PCR en otras cepas pertenecientes a diferentes especies del género como A. formosense CC-Nfb-7, A. fermentarium CC-LY743, A. halopraeferens Au4, A. oryzae COC8, A. palatum WW10, A. melinis TMYC 0552, A. lipoferum Sp59b, A. canadense DS2, A. rugosum IMMIB AFH-6, A. agrícola CC-HIH038, A. soli CC-Ly788 y A. zeae N7 (fig. 1B). Otras cepas no relacionadas filogenéticamente con este género, pero frecuentemente utilizadas para la formulación comercial de inoculantes combinados con A. brasilense Az39 fueron evaluadas. Estas cepas fueron P pro-tegens (ex-P! fluorescens) CHAO, B. japonicum E109 y B. subtilis BH8 y ninguna presentó resultados positivos para la amplificación con los cebadores TP5. En las figura 1C y D se observa la amplificación del gen 16S rRNA con los cebadores específicos para el género Azospirillum utilizados como control21. Por otro lado, se comprobó la amplificación por PCR con cebadores del tipo BOX-AR1 en las cepas pertenecientes a otros géneros microbianos (datos no mostrados). La evaluación con los cebadores TP6 y TM1 presentó resultados inesperados ya que hubo amplificación inespecífica en algunas cepas del género, por lo que los mismos fueron descartados para ensayos posteriores de este trabajo. Si bien, se analizó el total de las cepas presentadas en la tabla 1 con los cebadores TP6 y TM1 (datos no mostrados), en la figura S5 se presentan a modo de ejemplo algunos de los productos inespecíficos amplificados por dichos cebadores. Algunas cepas amplificaron bandas únicas, definidas y de peso molecular diferente al esperado para el caso de Az39; mientras que en otras se observó más de un producto de PCR. A pesar de que los cebadores TP6 y TM1 amplificaron de manera inespecífica para otros microorganismos, fueron eficientes para diferenciar la cepa Az39 del resto de las cepas evaluadas ya que, aun existiendo amplificación inespecífica, fue posible distinguir A. brasilense Az39 de otras cepas.
En cuanto a la sensibilidad, la metodología fue probada con el par de cebadores denominado TP5 ya que resultó ser el más específico para la reacción de PCR. En la figura 2A se observan los productos de amplificación de PCR empleando el par de cebadores TP5 para los que se utilizó como molde ADN extraído por la resina Chelex® 100, a partir de diluciones decimales de un cultivo puro de Az39. Nuestros resultados demuestran que el punto de corte con esta metodología se obtiene en la dilución 10-4 equivalente a un título bacteriano de 2,5x105 UFC/ml o una concentración de ADN equivalente a 4,5 ng/^l ADN. En la figura 2B se observan los productos de amplificación de PCR, utilizando como molde ADN extraído por un kit comercial. Nuestros resultados muestran que el punto de corte de esta metodología, teniendo en cuenta el número de células, se obtiene en dilución 10-7, equivalente a un título de 4,5 x 102 UFC/ml y con un contenido equivalente de ADN de 0,88 ng/^l; sin embargo, el límite de detección en base a la concentración de ADN fue establecido donde la amplificación se alcanzó con 0,098 ng/^l de ADN, a pesar de que la amplificación fue tenuemente observada en una concentración inferior de ADN (0,0092 ng/^l). El recuento inicial del cultivo del cual se realizaron las diluciones fue 2,5x109 UFC/ml. Es importante aclarar que cuando el ADN fue extraído con la metodología de Alippi et al.2 se obtuvieron resultados similares para las amplificaciones realizadas con los cebadores TP6 (datos no mostrados). En función de los resultados obtenidos a nivel de la especificidad y sensibilidad del método en cultivos puros, se pudo determinar la especificidad en cultivos mixtos. En la figura 3A se observa la amplificación por PCR empleando los cebadores TP5, a partir de mezclas de cultivos puros de A. brasilense Az39 y B. japonicum E109 en distintas proporciones. Los resultados demostraron que cuando la cepa Az39 está presente, la amplificación por PCR es positiva independientemente de las proporciones de los microorganismos presentes en la mezcla. La especificidad en mezclas de Azospirillum se evaluó mezclando por un lado cepas de la misma especie (mezcla 1) y cepas pertenecientes a distintas especies (mezcla 2 y 3). En la figura 3B se observa el resultado para las tres mezclas de cultivos evaluadas, donde el producto de 307 pb se obtuvo en todos los casos en los que estuvo presente Az39; mientras que los resultados negativos fueron pertenecientes a las mezclas control sin el agregado de esta cepa.

Figura 1:A y B: Electroforesis en gel de agarosa 1% de los productos de amplificación con los cebadores TP5, específicos para la cepa Az39. C y D) Electroforesis en gel de agarosa 1% de los productos de amplificación con los cebadores 16S rRNA específicos de género (Lin et al., 2011) utilizados como control positivo para las cepas Azospirillum spp. Referencias: (MM) marcador de peso molecular 100 bp Trans Plus ADN Ladder; (1) A. brasilense Az39; (2) A. brasilense Sp245; (3) A. brasilense FP2; (4) A. brasilense Abv5; (5) A. brasilense Abv6; (6) A. brasilense Az 19; (7) A. brasilense Az 36; (8) A. brasilense Az 45; (9) A. brasilense Az 48; (10) A. brasilense Az 63; (11) A. brasilense Az 65; (12) Pseudomonas protegens CHA0; (13) Bradyrhizobium japonicum E109; (14) A. formosense CC-Nfb-7; (15) A. fermentarium CC-LY743; (16) A. halopraeferens Au4; (17) A. oryzae COC8; (18) A. palatum WW10; (19) A. melinis TMYC 0552; (20) A. lipoferum Sp59b; (21) A. canadense DS2; (22) A. rugosum IMMIB AFH-6; (23) A. agrícola CC-HIH038; (24) A. soli CC-Ly788; (25) A. zeae N7; (C-) control negativo (agua).
Control de calidad de formulados comercialesDesde el punto de vista del análisis microbiológico de los productos comerciales empleados en este trabajo, podemos mencionar que los resultados obtenidos a partir del empleo de la técnica de recuento por microgota arrojaron los siguientes títulos de recuento: 5,8 x 108 UFC/ml para el formulado I1; 3,38 x 107 UFC/ml y para el formulado I2 y < 1 x 105 UFC/ml para el formulado I3. En el caso de I1 e I2 se observaron colonias color rojo escarlata, con una superficie plana-craquelada, con bordes redondeados y textura seca; morfología típica para este grupo bacteriano. En el caso de I3 solo se observaron estas colonias después de sembrar de manera directa una alícuota del producto en una placa conteniendo medio LBRC agarizado. Cuando el ADN molde utilizado fue obtenido a partir de colonias puras cultivadas de los recuentos (I1 e I2) o de la siembra en directo del producto I3, la electroforesis en gel de agarosa, presentó resultados positivos para los tres productos comerciales (datos no mostrados). Mientras que cuando se utilizó como molde el ADN obtenido directamente de los formulados, resultados positivos se observaron para los inoculantes I1 e I2; sin embargo, en el caso del inoculante I3, no se observó amplificación (fig. 4).

Figura 2: Electroforesis en gel de agarosa 1% de los productos de amplificación con el par de cebadores TP5 para la determinación de la sensibilidad del método a partir de ADN molde obtenido por A) resina Chelex® 100 y B) kit comercial. Referencias A): (MM) marcador molecular 100 bp Trans Plus ADN Ladder; (1). A. brasilense Az39 109 UFC/ml); (2) 108 UFC/ml; (3) 107 UFC/ml; (4) 106 UFC/ml; (5) 105 UFC/ml; (6) 104 UFC/ml; (7) 103 UFC/ml; (8) 102 UFC/ml. Referencias B): (MM) marcador molecular 100bp Trans Plus ADN Ladder; (1) A. brasilense Az39 8,5x105 UFC/ml); (2) 5,3x104 UFC/ml; (3) 6,7x103 UFC/ml; (4) 2,5x102 UFC/ml (equivalente a 0,88 ng/pl ADN); (5)* 0,098 ng/pl ADN y (6)** 0,0092 ng/pl de ADN. Dilución *decimal y ** centesimal de la muestra obtenida de ADN en (4).

Figura 3: Electroforesis en gel de agarosa al 1% de los productos de amplificación con los cebadores TP5 en la evaluación de la especificidad en cultivos mixtos. Referencias A): (MM) marcador de peso molecular 1 kb GeneRuler™; (1) 100% B. japonicum E109; (2) 70% B. japonicum E109 + 30% A. brasilense Az39; (3) 50% B. japonicum E109 + 50% A. brasilense Az39; (4) 30% B. japonicum E109 + 70% A. brasilense Az39; (5) 100% A. brasilense Az39. Referencias B): (1) mezcla 1; (2) mezcla 2; (3) mezcla 3; (MM) marcador de peso molecular 100 bp Trans Plus ADN Ladder; (4) control negativo mezcla 1; (5) control negativo mezcla 2; (6) control negativo mezcla 3.

Discusión
Nuestro trabajo consistió en realizar una identificación certera de A. brasilense Az39, a través de una metodología rápida y accesible, que pueda ser utilizada como una estrategia de control de calidad de inoculantes comerciales. Esto se logró mediante el desarrollo de marcadores moleculares del tipo SCAR. Hasta el momento, la identificación cepa-específica se podía realizar mediante el marcado de la misma con genes reporteros, pero esto involucra modificaciones a nivel genético, y solo puede aplicarse con fines de investigación14. La amplificación por PCR de secuencias específicas de cada cepa es una metodología simple y permite detectar particularidades de la cepa sin necesidad de su modificación genética. El uso de herramientas bioinfor-máticas y el acceso a la secuencia completa del genoma de Az39 nos permitió hallar fragmentos únicos en la secuencia de ADN de esta cepa. La búsqueda se realizó mediante la simulación de AFLP-PCR in silico. La mayoría de los trabajos donde se aborda la problemática de la identificación específica de cepas, se realizan de manera experimental a través de la búsqueda de secuencias únicas utilizando metodologías del tipo fingerprinting15,16. Esta alternativa resulta más costosa, laboriosa e implica una mayor inversión de tiempo, pero es de gran utilidad cuando se desconoce la secuencia genómica de la bacteria de interés. Entre los trabajos publicados donde obtienen marcadores SCAR específicos para otros microorganismos del género Azospirillum de manera experimental, se encuentran los realizados por Couillerot et al.12,13 y de Priya et al.28. Estos autores lograron desarrollar protocolos de identificación y cuantificación por qPCR con cebadores disenados para amplificar secuencias específicas de las cepas A. lipoferum CRT113, A. brasilense UAP-154 y CFN-53512 y Azospirillum sp. Az204 y A. brasilense Sp728, respectivamente. Cabe destacar que no se han encontrado trabajos donde se lleve a cabo de manera in vitro la búsqueda de secuencias SCAR a partir de AFLP-PCR; sin embargo, la disponibilidad de secuencias genómicas en diferentes bases de datos, permite el desarrollo de estas novedosas herramientas como alternativa. Tal es así que se ha logrado encontrar secuencias específicas de manera in silico, mediante el corte en pequeños fragmentos de 500 pb (simulación de digestión enzimática) del ADN genómico de una cepa de interés. Con el posterior análisis mediante BLAST sobre el genoma de una cepa cercanamente relacionada35 (hibridación y eliminación de los fragmentos compartidos). De este modo, se detectaron fragmentos únicos para la identificación de A. brasilense FP2 mediante la amplificación por PCR de secuencias específicas35. Este abordaje permitió la identificación certera de la cepa de una manera diferente a la planteada en este trabajo, pero igualmente efectiva. La herramienta molecular utilizada en nuestro trabajo ha permitido desarrollar una metodología simple, rápida y económica, comparada con los reportes previos; que permite de igual modo conseguir una gran cantidad de secuencias específicas para la identificación de la cepa.
Los fragmentos de ADN encontrados, distinguidos por estar presentes únicamente en el genoma de Az39, fueron utilizados como molde para el disenño de los cebadores. Cuatro de estos pares de cebadores fueron evaluados in vitro. Solo el par denominado TP5 demostró ser específico desde una primera instancia, el mismo fue seleccionado para las evaluaciones posteriores. En el caso de los otros cebadores, el grado de especificidad fue bajo, sin embargo, es posible aumentar la misma variando condiciones como lo son la Ta, el tiempo de elongación o la concentración de cebadores. A pesar de esta diferencia observada entre los ensayos in vitro e in silico, para los cebadores TP6 y TM1, con respecto a otros microorganismos, siempre se observó el producto esperado en AZ39. Adicionalmente, ensayos preliminares realizados en nuestro laboratorio, demostraron que el aumento de la Ta mejora la especificidad con respecto a la amplificación sobre otras cepas (datos no mostrados); sin embargo, en el caso de los cebadores EM1, no se observó el producto esperado y este tipo de diferencias entre los resultados obtenidos in vitro e in silico también han sido observados en otros trabajos17,25. De manera general, los pares de cebadores TP6, TM1 y EM1 fueron descartados a lo largo de los experimentos debido a que el par TP5 presentó el mejor comportamiento.
En cuanto a la prueba de especificidad en una mezcla bacteriana, en el trabajo de Shime-Hattori et al.34 utilizaron una cepa de Bacillus en diferentes proporciones. Ensayos similares fueron realizados en nuestro laboratorio, empleando una mezcla de A. brasilense Az39 con B. japonicum E109.
Existen reportes que demuestran que la coinoculación con ambas cepas en plantas de soja, genera un efecto positivo en la estimulación del crecimiento de este cultivo7,29,30. En base a esta evidencia, y teniendo en cuenta que en la actualidad se comercializan inoculantes formulados con ambas bacterias, probamos que los cebadores disenñados para A. brasilense Az39 fueron eficientes. Así como también fueron capaces de distinguir la cepa de interés en mezclas con microorganismos del mismo género y especie.
En los ensayos con mezclas bacterianas, el par TP5 demostró su aptitud de diferenciar Az39 de otras cepas, pero siempre respetando una concentración final de células de Az39 por encima del valor de 105 UFC/ml cuando se emplea el método de extracción con Chelex® 100. A pesar de las desventajas observadas en la extracción de ADN con esta resina fue propuesta como un método alternativo debido a que a los fines de control de calidad de inoculantes es más accesible y económico que la utilización de kit comerciales. Otra metodología alternativa para la detección directa desde formulados comerciales consistiría en la utilización de Sephadex G100 al 10%28.
Hasta el momento, el control de calidad de inoculantes a base de Azospirillum se realiza según lo expresa el manual de Procedimientos de la REDCAI10. El mismo consiste en el recuento de células viables mediante la metodología de siembra en placas por extensión en superficie en medio rojo Congo. Para la evaluación de microorganismos contaminantes, el manual de procedimientos sugiere la siembra directa en medio agar tripticasa soya para los microorganismos en general y en agar Sabouraud para hongos saprofíticos. Adicionalmente se sugiere una coloración mediante la técnica de Gram y la observación al microscopio de un preparado fresco de la muestra. Por otra parte, debido al dimorfismo de colonias de Azospirillum sp., en placas de medio rojo Congo, se realiza una prueba de la capacidad diazotrófica en condiciones de microaerofilia en medio NFb, de las colonias consideradas presuntivas. En nuestro trabajo proponemos complementar la metodología propuesta por la REDCAI con la confirmación molecular de la identidad de la cepa. Los pares de cebadores disenñados en nuestro trabajo cumplieron con los requisitos esperados respecto a la especificidad de la cepa Az39 en muestras comerciales. Nuestros resultados demostraron que la combinación de las metodologías de microbiología clásica y molecular permiten la evaluación eficiente de la calidad de inoculantes con diferente aptitud comercial, ya que se pudo distinguir entre productos dentro y fuera de la fecha de vencimiento y con recuentos por encima y debajo de la garantía de tales productos. En cuanto al límite de detección de la reacción de PCR, el mismo fue dependiente de la metodología de extracción de ADN empleada, siendo en este caso para la resina Chelex® 100 del orden de 105 UFC/ml y equivalente a 4,5 ng/^l ADN. Si bien, la sensibilidad del método es baja empleando la resina iónica, es necesario destacar que los productos comerciales formulados con Azospirillum deben contener no menos de 1x109 UFC/ml a la fecha de elaboración y no menos de 1x107 UFC/ml a la fecha de vencimiento, por lo que el método de extracción con resina, a pesar de su baja eficiencia, todavía sería útil si consideramos la definición de producto apto para uso agrícola con más de 1x107 UFC/ml. Por otro lado, un aumento significativo de la sensibilidad del método fue obtenido cuando el ADN bacteriano se extrajo con un kit comercial, para el que un título bacteriano de 102 UFC/ml o una concentración equivalente de ADN de 0,88 ng/^l fue suficiente para su amplificación por PCR. De la misma manera, concentraciones de ADN diez y cien veces inferior a la equivalente en 102 UFC/ml (0,098 o 0,0092 ng/^l) resultaron positivas para la amplificación, aunque en el último caso, tenuemente. Esto indica claramente que el límite de la metodología de amplificación por PCR está por debajo del límite de detección del método de recuento de bacterias viables disponible en este momento. Finalmente y de manera complementaria, podemos decir que otros investigadores han desarrollado metodologías similares para la identificación de microorganismos a nivel de género, que pueden ser utilizadas de manera complementaria para la evaluación del control de calidad de inoculantes6,21,23,34.
Conclusión
La caracterización fenotípica no es suficiente para la identificación inequívoca de un microorganismo, pero puede bridar conocimientos sobre la pureza de un cultivo y el número de microorganismos presentes en el mismo. Esto reafirma que, en muchos casos, la combinación de técnicas microbiológicas y moleculares es una estrategia clave para establecer un procedimiento efectivo de control de calidad. Muchas técnicas moleculares han sido descritas para la identificación de Azospirillum pero ninguna destinada al reconocimiento específico de A. brasilense Az39, la cepa más utilizada para la formulación de inoculantes en la República Argentina. Hasta el momento, la identificación de esta cepa empleando herramientas moleculares se puede realizar por genotipado (16S rRNA y rpoD) de una manera relativamente rápida; sin embargo, estas metodologías requieren de amplificación y secuenciación, por lo que las hace imprácticas para el análisis cotidiano. En este trabajo, la disponibilidad de herramientas bioinformáticas accesibles de manera gratuita y la ventaja de contar con la secuencia del genoma de A. brasilense Az39, nos permitió realizar el análisis del polimorfismo de ADN en nuestra cepa de interés. Esta estrategia condujo al hallazgo de secuencias únicas en el genoma de Az39. El diseño de cebadores, dirigidos a dichas secuencias, demostró no solo ser específico a nivel in silico, sino que también permite determinar la presencia de A. brasilense Az39 en un cultivo puro tan solo con amplificación y electroforesis en gel de agarosa. Estas metodologías resultan ser prácticas y accesibles, además hacen posible una identificación de rutina certera. A pesar de la desventaja que presenta la técnica desarrollada en cuanto al bajo poder de detección obtenido, donde la extracción del ADN genómico de las muestras se obtiene por técnicas robustas, sencillas, rápidas y de bajo costo, es exitosa tanto en cultivos puros como en mezclas bacterianas. Así como también, los resultados obtenidos para formulados comerciales permitieron demostrar que los productos evaluados estaban formulados con la cepa Az39 de A. brasilense recomendada por el Servicio Nacional de Sanidad Animal (Servicio Nacional de Sanidad y Calidad Agroalimentaria) en la República Argentina para la producción de inoculantes para numerosas especies vegetales de interés agrícola.
Financiación
Este trabajo fue financiado por el Consejo Nacional de Investigación Científica Tecnológica de Argentina (CONICET) y el Fondo Nacional de Ciencia y Tecnología (FONCyT), a través de sus proyectos PICT-2012-1051 y PICT 2015-1599.
Conflicto de intereses
Los autores declaran no tener ningún conflicto de intereses.
AgradecimientosLos autores del trabajo quieren mostrar su agradecimiento a la Universidad Nacional de Río Cuarto (UNRC); al Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET) y al Instituto Nacional de Tecnología Agropecuaria (INTA). En particular, agradecemos al Dr. Emanuel de Souza de la Universidad Federal de Paraná (Curitiba, Brasil); al Prof. C.C. Young de la Universidad Nacional Chung Hsing University (Chung Hsing, Taiwán) y a la Prof.a Florence Wisnieski-Dyé del Laboratorio de Ecología de la Rizosfera de la Universidad de Lyon (Lyon, Francia), por facilitarnos diferentes cepas de Azospirillum utilizadas en este trabajo.
Anexo. Material adicional Se puede consultar material adicional a este artículo en su versión electrónica disponible en doi:10.1016/ j.ram.2019.02.004.

