










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. v.11 n.1 Mendoza ene./jun. 2004
El registro de Ctenomys talarum durante el Pleistoceno Tardío - Holoceno de las Sierras de Tandilia Oriental
Carlos A. Quintana
Laboratorio de Arqueología, Universidad Nacional de Mar del Plata, <quintana@copetel.com.ar>
RESUMEN. Se discute la identidad de 1.004 restos craneanos y mandibulares de Ctenomys que provienen de la secuencia estratigráfica del sitio arqueológico Cueva Tixi. De acuerdo a un análisis de similitud basado en caracteres craneanos se concluye que los restos estudiados pertenecen a la especie Ctenomys talarum Thomas, 1898. Su abundancia, medida como porcentaje del número mínimo de individuos, disminuye paulatinamente desde el Pleistoceno tardío de Cueva Tixi, hasta desaparecer en tiempos históricos del área serrana.
Palabras clave: Ctenomys, Cueva Tixi, Tandilia, extinción, Argentina.
ABSTRACT. The Upper Pleistocene-Holocene record of Ctenomys talarum in EasternTandilia Range. The identity of 1,004 cranial and mandibular remains of Ctenomys from the stratigraphic sequence of the archaeological site Cueva Tixi, is discussed. According to a similarity analysis based on cranial characters the studied remains belong to the species Ctenomys talarum Thomas, 1898. Their abundance, measured as a percentage of the minimum number of specimens, diminishes gradually from the Late Pleistocene of Cueva Tixi, until disappearing in historical times from the mountain area.
Key words: Ctenomys, Tixi Cave, Tandilia, extinction, Argentina.
INTRODUCCIÓN
Los roedores Octodontidae de la subfamilia Ctenomyinae cuentan con un buen registro fósil, compuesto por géneros monoespecíficos o pauciespecíficos del Mioceno y Plioceno. Al menos desde el Plioceno se caracterizan por un modo de vida hipogeo y con el desarrollo de claras adaptaciones a la excavación (Reig et al., 1990; Quintana,1994; Verzi, 2002). Los tuco-tucos del género Ctenomys conforman el grupo de vertebrados con la mayor cantidad de especies de hábitos subterráneos (Reig et al., 1990). Su origen se remonta al Plioceno tardío, siendo Ctenomys chapalmalensis la especie más antigua y primitiva conocida. Esta antecede a la explosiva radiación adaptativa del género que se desarrolló durante el Pleistoceno, dando como resultado a alrededor de 60 especies actuales que se distribuyen en gran parte de América del Sur.
La vida subterránea propicia el desarrollo de adaptaciones morfológicas muy similares entre los mamíferos cavadores, ocasionando convergencias notables aun entre clados no emparentados de regiones distintas (Nevo, 1979). Así, la anatomía de las especies actuales de Ctenomys es muy homogénea a pesar de su diversidad, pero algunos caracteres muestran variabilidad ínter e intraespecífica lo que torna complejo el esclarecimiento del estado taxonómico de las entidades de tamaños similares o que se distribuyen en parapatría (Reig et al., 1965). Esta situación se magnifica en las formas extintas, las cuales suelen estar representadas por materiales de poca calidad de preservación anatómica. El registro fósil de Ctenomys es redundante, particularmente durante el Pleistoceno más tardío y el Holoceno, aunque sólo se cuenta con una contribución (Rusconi, 1931) que intentó elucidar su diversidad. El avance en el conocimiento de las especies vivientes llevó a que en el estudio de las fósiles se pasara de nominar especies nuevas a partir de restos fragmentarios y no diagnósticos, a una actitud más conservadora de citas sólo a nivel de género. Esta alternativa, si bien es más prudente, impide conocer las relaciones entre las entidades extintas entre sí y respecto de las vivientes. La solución a esta situación requiere de una revisión extensa de los restos fósiles, lo cual es una empresa de envergadura sobre todo si se considera que no está totalmente descripta la anatomía de las numerosas especies actuales. Estas carecen de estudios de su registro fósil, a pesar de la abundancia de hallazgos de este género durante el Holoceno. Un caso particular ocurre en el sitio arqueológico Cueva Tixi, el cual tiene cuantiosos ejemplares de una especie de Ctenomys distribuida durante más de 10.000 años (Quintana y Mazzanti, 1996; Quintana, 2001a). La misma fue referida a Ctenomystalarum Thomas, 1898 sin discutir los caracteres que sostienen su identificación (Quintana y Mazzanti, 1996) , por lo que esta contribución tiene el objetivo de discutir el estado taxonómico de esos restos y describir su registro fósil.
MATERIALES Y MÉTODOS
Los restos analizados provienen del sitio arqueológico Cueva Tixi, sierra La Vigilancia, Provincia de Buenos Aires (Mazzanti, 1997; 2001) (Fig. 1) . Para conocer la antigüedad de los mismos se obtuvieron dataciones radiocarbónicas de carbón vegetal de áreas de combustión (Mazzanti, 1997), procesadas con un acelerador de espectrometría de masas (AMS) en la Universidad de Arizona. El sitio fue excavado por medio de la técnica del decapado de los estratos naturales y depósitos antrópicos, contándose con un registro tridimensional de cada vestigio (Mazzanti, 2001). El material faunístico fue recuperado en planta o con cernidores bajo agua de malla de 2 mm de poro, y están depositados en la colección científica del Laboratorio de Arqueología de la Universidad Nacional de Mar del Plata. La estratigrafía fue descripta por Martínez y Osterrieth (2001).
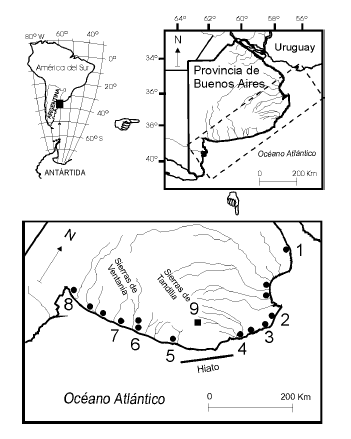
Fig. 1.Ubicación geográfica de las poblaciones actuales de Ctenomystalarum (Contreras y Reig, 1965; Massarini et al., 2002) y de Cueva Tixi. Poblaciones actuales de C. talarum: 1, Magdalena. 2, San Clemente del Tuyú. 3, Valeria del Mar. 4, Santa Clara del Mar. 5, Necochea. 6, Río Quequén Salado.7, Monte Hermoso. 8, Bahía Blanca. Cueva Tixi y Cueva El Abra: 9.
Geographic location of Ctenomys talarum (Contreras and Reig, 1965; Massarini et al., 2002) and Cueva Tixi. Living populations of C. talarum: 1 to 8. Cueva Tixi and Cueva El Abra: 9.
Para poner a prueba si los restos fósiles corresponden a Ctenomys talarum se siguieron los criterios desarrollados por Reig et al. (1965) (ver Apéndice), y también se evaluaron los caracteres allí descriptos con ejemplares actuales de esa especie procedentes de Mar de Cobo y Villa Gesell (provincia de Buenos Aires). El análisis de similitud global se realizó con el programa NTSYS-PC (Numerical Taxonomic System and Multivariate Analysis System, V 1.4). Se utilizó el coeficiente de asociación de Jaccard debido a que no ingresa los estados de caracteres ausentes compartidos (0, 0). Se calculó el coeficiente de correlación cofenético para evaluar la congruencia entre la matriz de asociación (producto de la aplicación del citado coeficiente) y el fenograma que deriva de ella (Crisci y López Armengol, 1983). Para conocer la abundancia de Ctenomys se calculó el número de especímenes óseos identificados (NISP) (Paine, 1975) y el número mínimo de individuos (MNI) (White, 1953). Para efectuar comparaciones entre distintos estratos se calculó el MNI como porcentaje del MNI del total de especies de cada estrato, de modo de evitar el efecto de los tamaños muestrales distintos. La proclividad de los incisivos se midió siguiendo los criterios de Lessa (1990) y las diferenciasentre los ejemplares fósiles y actuales se pusieron a prueba con el test no paramétrico de Mann-Withney.
RESULTADOS
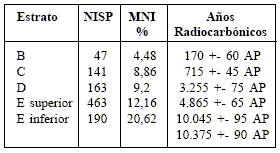
Se recolectaron 1.004 restos craneanos y mandibulares de Ctenomys, y numerosos huesos poscraneanos (Quintana, 2001a). Ctenomys está presente en los estratos B, C, D y E, de Cueva Tixi, su abundancia por estrato se describe en la Tabla 1.
Tabla 1
Abundancia y cronología de Ctenomys talarum en el registro estratigráfico de Cueva Tixi. Sólo se cuantificaron los restos craneanos y mandibulares. NISP: número total de restos identificados. MNI%: número mínimo de individuos en relación porcentual con el total del MNI de especies de mamíferos de cada estrato. AP: años antes del presente. Ver Fig. 4.
Abundance and chronology of Ctenomys talarum in the stratigraphic record of Cueva Tixi. Only the cranial and mandibular remains were quantified. NISP: total number of identified remains. MNI%: individuals’ minimum number in percentage relationship with the total of the MNI of species of mammals of each stratum. AP: years before present. See Fig. 4.
La comparación entre los ejemplares de Ctenomys de cada estrato no presentó diferencias que puedan ser atribuidas a taxones distintos. La muestra está compuesta por ejemplares juveniles, sólo unos pocos son adultos-jóvenes. El rostro presenta dos expresiones del contorno de los premaxilares: robusto, con extrusión de los alvéolos de los incisivos, y grácil sin esa extrusión. En los siguientes caracteres craneanos y mandibulares no se halló variación de nivel interespecífico: proclividad de los incisivos; forma de los nasales; expansión y forma del nacimiento del arco cigomático, presencia de una expansión del maxilar por encima del yugal; desarrollo de las apófisis del yugal; presencia de una depresión antero-dorsal en el yugal; forma y origen anterior de los agujeros incisivos; presencia y desarrollo del foramen interpremaxilar; forma de los frontales; desarrollo del proceso postcondiloideo de la rama mandibular.
El análisis de similitud entre Ctenomys de Cueva Tixi con otras siete especies del género, mostró una asociación con Ctenomystalarum al 100% de similitud (Fig. 2). La comparación detallada de la anatomía craneana de los restos fósiles con ejemplares de Ctenomystalarum actuales brindó una concordancia completa de estados de caracteres presentes compartidos (Fig. 3). Los siguientes caracteres de los ejemplares fósiles (Figs. 2 a 3) son coincidentes con los de Ctenomys talarum Thomas, 1898: tamaño pequeño (longitud de cráneo juvenil de la Fig. 3: 38 mm); incisivos: moderadamente proclives; nasales: cortos y estrechos abruptamente hacia atrás; su extremo anterior se proyecta hacia delante en forma de una V invertida; arco cigomático: moderadamente expandido, con su ancho máximo a nivel de la apófisis paraorbitaria del yugal; nace abrupto pero sin formar un ángulo recto con el eje sagital; maxilar: con una expansión por encima del yugal que participa en la delimitación ventral de la órbita; yugal: con una depresión ántero-dorsal poco profunda pero extendida, de la que participa la expansión orbitaria del maxilar, similar a la depresión cigomática de Osgood (ver Contreras y Berry, 1982); la apófisis paraorbitaria y la espina inferior del yugal son robustas; anchobimastoideo: menor que el ancho bicigomático; ampollas auditivas: pequeñas (apenas visibles en norma dorsal); agujeros incisivos: grandes, con un canal estrecho y de bordes nítidos; se originan a nivel de la mitad posterior de las apófisis palatinas; foramen interpremaxilar: presente y bien desarrollado; frontales: ligeramente estrechos por detrás de las apófisis postorbitarias; unidos a los escamosos mediante una sutura dorsal interna respecto de la cresta temporal; escamosos: forman con los frontales una apófisis posterior a la postorbitaria; osificaciones dorsales de los senos epitimpánicos: presentes y dispuestos entre el escamoso, parietales y la cresta occipital; interparietal: presente; foramen magnum: con el borde superior bajo, poco convexo y a la altura de los extremos dorsales de los cóndilos occipitales.
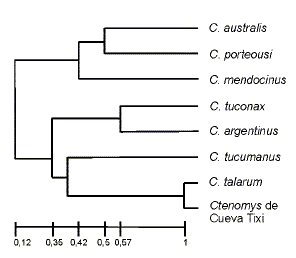
Fig. 2.Relaciones de similitud entre 7 especies vivientes de Ctenomys y los restos de Cueva Tixi. Fenograma de 13 caracteres craneanos, utilizando ligamiento promedio, UPGMA y el coeficiente de Jaccard. Coeficiente de Correlación Cofenético= 0,954.
Phenetic relationships among the remains of Cueva Tixi and 7 living Ctenomys species. Phenogram of 13 cranial characters, using UPGMA and Jaccard coefficient. Cophenetic correlation coefficient= 0,954.
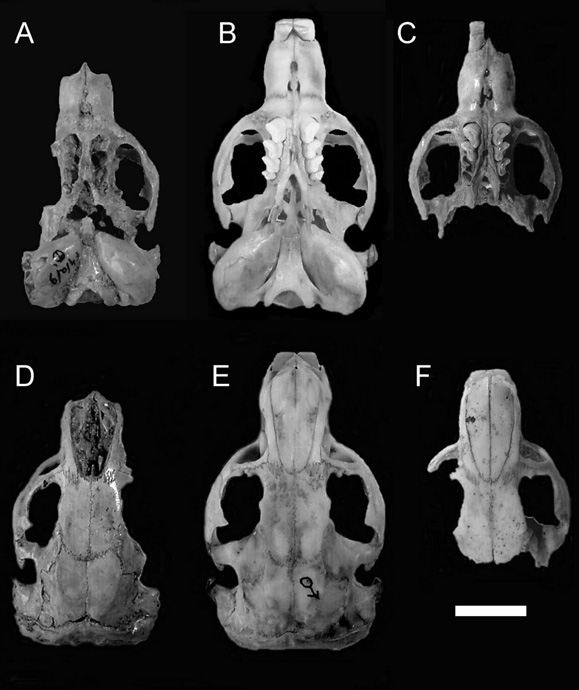
Fig. 3.Cráneos de Ctenomystalarum. A y D, Cueva Tixi, Estrato E superior. B y E, Mar de Cobo, macho adulto. C, Cueva Tixi, Estrato D. F, Cueva Tixi, Estrato D. Escala = 10 mm.
Skulls of Ctenomys talarum. A and D, Cueva Tixi , upper level E). B and E, Mar de Cobo, adult male . C, Cueva Tixi, level D. F, Cueva Tixi, level D. Scale = 10 mm.

Fig. 4.Variación de la abundancia (MNI%) de Ctenomys talarum en los estratos de Cueva Tixi, comparado con el resto de los roedores caviomorfos registrados en ese sitio. Ver Tabla 1.
Variation of the abundance (MNI%) of Ctenomys talarum in Cueva Tixi , respect to the remainder caviomorph species recorded in the site. See Table 1.
La proclividad de los incisivos de la muestra de Cueva Tixi aparenta ser mayor que la de las poblaciones actuales de Ctenomystalarum, pero su evaluación no brindó diferencias significativas ( P> 0,11). Los incisivos de los fósiles, por causas tafonómicas, suelen preservarse desplazados dentro del alvéolo, lo que en algunos casos incrementa, en apariencia, la proclividad. Los casos extremos impidieron la medición.
En el sitio arqueológico Cueva El Abra, que se encuentra a unos 10 kilómetros de Cueva Tixi, también se hallaron restos de Ctenomys (Quintana et al., 2003) pero no son identificables al nivel de especie porque se trata de fragmentos mandibulares escasos y mal preservados.
DISCUSIÓN
Debido a la redundancia de los estados de caracteres de los restos de Ctenomys analizados en la secuencia estratigráfica de Cueva Tixi, se supone que se trata de una única especie. Sobre la base de los análisis y comparaciones con especies actuales la misma se refiere a C. talarum. La diversidad descripta en la forma del rostro coincide con la hallada en la población actual de Mar de Cobo: las hembras inmaduras presentan un rostro grácil, mientras que los machos adultos o adultos jóvenes tienen un rostro más robusto y con las raíces de los incisivos más divergentes. Es de destacar que C. talarum es una de las especies más variables del género y con un marcado dimorfismo sexual (Reig et al., 1992; Massarini et al., 2002; Tolosa, 2003; Malizia y Bush, 1991; Zenuto et al., 1999).
A pesar de que los ejemplares estudiados provienen de un sitio arqueológico y se distribuyen junto a objetos culturales, su presencia en la secuencia sedimentaria se debe a un mismo proceso natural. Los restos de C.talarum de Cueva Tixi fueron ingresados a la cueva a través de egagrópilas de aves rapaces (Quintana, 2001b). Este origen concuerda con la mayor abundancia de individuos juveniles en la muestra. Las poblaciones actuales de C.talarum son predadas por aves que seleccionan los tamaños menores de sus presas, conformando una muestra sesgada hacia individuos juveniles (Vassallo et al., 1994) lo que ocasiona un registro similar al de Cueva Tixi.
La distribución actual de las poblaciones de C.talarum comprende una extensa franja costera del litoral Atlántico, situada entre la ciudad de Magdalena y Bahía Blanca. Pero presenta un hiato de unos 150 km entre las ciudades de Santa Clara y Necochea que separa a las poblaciones hacia el norte y hacia el sur (Reig et al., 1965; Contreras y Reig, 1965) (Fig. 1). Estas poblaciones fueron referidas a sendas subespecies, pero recientemente se demostró que las mismas carecen de fundamento fáctico (Tolosa, 2003). También se conocen dos poblaciones de latitudes más continentales en las localidades de Saladillo y Cazón, oeste de la provincia de Buenos Aires (Piantanida et al., 2000). En todos los casos C.talarum excava sus madrigueras en suelos no totalmente arenosos, ya que se trata de suelos más compactos que, por ejemplo, el de las dunas inmediatamente lindantes con el mar y con una buena biomasa vegetal por encima y debajo de la superficie (Malizia et al., 1991). La presencia de C.talarum durante 10.500 años en los valles de Sierra La Vigilancia plantea una paleodistribución algo diferente a la distribución actual. El aspecto más resaltante es su registro en el ámbito serrano, donde actualmente no se desarrollan suelos arenosos. Sin embargo, hacia el Pleistoceno tardío-Holoceno temprano los suelos estaban compuestos por menos arcillas que en la actualidad, debido a la gran proporción de loess no edafizado (médano invasor) (Frengüelli, 1950). Esta constitución sedimentaria, posiblemente, favoreció el asentamiento de C. talarum en el ambiente serrano. Durante el Holoceno se desarrollaron suelos más compactos, por lo que la tolerancia a los tipos de suelo de esta especie fue todavía mayor que el conocido. Actualmente la línea de costa se encuentra a unos 50 km hacia el este de Cueva Tixi (Fig. 1), pero esa cota fue mayor en el pasado, durante el Pleistoceno tardío-Holoceno temprano se estima una distancia de 100 km entre esta cueva y el océano (Fray y Ewing, 1963), lo cual diferencia todavía más la geografía de las poblaciones de La Vigilancia respecto de las actuales que se ubican en la franja costera. Las poblaciones actuales de C. talarum serían un relicto reciente de una distribución histórica de mayor amplitud, o la consecuencia de procesos migratorios (Contreras y Bidau, 1999).
El mayor cambio en la composición específica de la secuencia de Cueva Tixi se presenta hacia tiempos históricos (Quintana, 2001a) lo cual está manifestado por la extinción de Galea tixiensis, una especie de roedor Caviinae endémica de Tandilia (Quintana, 2001c), la extinción de las poblaciones locales de dos especies de roedores cricétidos (Pseudorizomys simplex y Bibimystorresi), de dos ungulados (Lama guanicoe y Ozotoceros bezoarticus) y de un ave (Rhea americana). En este contexto también desaparecieron las poblaciones de C. talarum. Pero, a diferencia de los restantes taxones citados, su abundancia sufrió una disminución constante desde el Pleistoceno tardío. Los valores absolutos (NMI) de restos de Ctenomys muestran la tendencia de esta disminución (Tabla 1), pero su cuantificación homogeneizando los números muestrales distintos de cada estrato (MNI%) muestran más claramente la retracción paulatina de las poblaciones serranas de C. talarum (Fig. 4) hasta su extinción en tiempos históricos cercanos al establecimiento de agroecosistemas.
APÉNDICE
Descripción de los caracteres y matriz usada para la construcción del fenograma de la figura 2. Description of the characters and matrix used for the construction of the cluster of the figure 2.
1 Nasal: Se angosta de modo abrupto hacia atrás 1. Se angosta de modo moderado hacia atrás 0.
2 Frontal: Aproximadamente plano en su mitad posterior y convexo anteriormente 1. Convexo posteriormente y casi plano anteriormente 0.
3 Frontal: Conserva aproximadamente su ancho hacia atrás 1. Se angosta hacia atrás 0.
4 Constricción interorbitaria: Bien marcada y amplia, con apófisis post-orbitarias del frontal conspicuas 1. Poco marcada y con apófisis post-orbitarias del frontal apenas evidentes 0.
5 Origen del arco cigomático respecto del maxilar: Abrupto, forma un ángulo aproximadamente recto 1. Gradual, forma un ángulo agudo 0.
6 Diámetro transverso máximo del arco cigomático: Aproximadamente a la misma altura de las apófisis paraorbitarias del yugal 1. Por detrás de esas apófisis 0.
7 Proceso frontal del yugal: Robusto, con las apófisis muy desarrolladas 1. Grácil, con esas apófisis poco desarrolladas 0.
8 Ampollas auditivas: Pequeñas y no visibles desde la norma superior del cráneo 1. Grandes y visibles desde la norma superior del cráneo 0.
9 Bordes del canal de los agujeros incisivos: Aproximadamente paralelos 1. No paralelos 0.
10 Nacimiento del canal de los agujeros incisivos: Hacia la mitad anterior del vómer, el cual es bien visible 1. Hacia la mitad posterior del vómer, el cual es poco visible 0
11 Foramen Interpremaxilar o de Hill: Bien desarrollado, ovalado y separado por el vómer de los agujeros incisivos 1. Poco desarrollado o ausente y forma un conjunto con los agujeros incisivos delimitado por el canal de éstos 0.
12 Borde superior del foramen magnum: Bajo, poco convexo y a la altura de los extremos dorsales de los cóndilos occipitales 1. Alto, convexo y por encima de los cóndilos 0
13 Forma de la cresta masetérica del yugal: Aproximadamente recta y se eleva suavemente hacia atrás 1. se eleva abruptamente hacia atrás 0.

AGRADECIMIENTOS
A los doctores Diego Verzi y Aldo Vasallo cuyas sugerencias permitieron mejorar el manuscrito. Esta investigación fue financiada en el marco del proyecto 04-09594 de la Agencia Nacional de Promoción Científica y Tecnológica y de la Universidad Nacional de Mar del Plata.
LITERATURA CITADA
CONTRERAS, J. y O. REIG. 1965. Datos sobre la distribución del género Ctenomys (R. O.) en la zona costera de la provincia de Bs. As. comprendida entre Necochea y Bahía Blanca. Physis, 25(69):169-186. [ Links ]
CONTRERAS, J. y C. BIDAU. 1999. Líneas generales del panorama evolutivo de los roedores excavadores sudamericanos del género Ctenomys (Mammalia, Rodentia, Caviomorpha: Ctenomyidae). Ciencia Siglo XXI, 1:1-22. [ Links ]
CONTRERAS, J. y L. BERRY. 1982. Ctenomys argentinus, una nueva especie de Tuco-tuco procedente de la provincia del Chaco Rep. Arg. (Rod. Oct.). Historia Natural 2, 20:165-173 [ Links ]
CRISCI, J. y M. LOPEZ ARMENGOL. 1983. Introducción a la teoría y práctica de la taxonomía numérica. OEA, Programa Regional de desarrollo científico y tecnológico, Monografía 26, Serie Biología, 128 pp. [ Links ]
FRAY, C. y M. EWING. 1963. Pleistocene sedimentation and fauna of the Argentine shelf. I. Wisconsin sea level as indicated in Argentine Continental shelf sediments. Proceedings, Academy Natural Science Philadelphia, 115:113-126. [ Links ]
FRENGÜELLI, J. 1950. Rasgos generales de la morfología y la geología de la Prov. de Bs. As. LEMIT 2 (33). 72 pp. [ Links ]
LESSA, E. 1990. Morphological evolution of subterranean mammals. Integrating structural, functional and ecological perspectives. Pp: 211-230. En: Evolution of subterranean mammals at the organismal and molecular levels (Nevo, E. y O. Reig, eds.). A.R. Liss-Wiley, New York. [ Links ]
MALIZIA, A. y C. BUSCH. 1991. Reproductive parameters and growth in the fossorial rodent Ctenomys talarum (Rodentia Octodontidae). Mammalia, 55:293-305 [ Links ]
MALIZIA, A., A. VASSALLO y C. BUSCH. 1991. Population and habitat characteristics of two sympatric species of Ctenomys (Rodentia, Octodontidae). Acta Theriologica, 36(1-2):87-94. [ Links ]
MARTÍNEZ, G. y M. OSTERRIETH. 2001. Estratigrafía, procesos formadores y paleoambientes. Pp.19-34. En: Cueva Tixi: cazadores y recolectores de las sierras de Tandilia oriental. I. Geología, Paleontología y Zooarqueología (Mazzanti, D y C. Quintana, eds.). Publicación Especial 1. Laboratorio de Arqueología, Universidad Nacional de Mar del Plata. [ Links ]
MASSARINI, A., D. MIZRAHI, S. TIRANTI, A. TOLOZA, F. LUNA y C. SCHLEICH. 2002. Extensive chromosomal variation in Ctenomystalarumtalarum from the Atlantic coast of Buenos Aires Province, Argentina (Rodentia: Octodontidae). Mastozoología Neotropical, 9(2):199-207. [ Links ]
MAZZANTI, D. 1997. Excavaciones arqueológicas en el sitio Cueva Tixi, Buenos Aires, Argentina. Latin American Antiquity, 8(1):55-62. [ Links ]
MAZZANTI, D. 2001. Las Investigaciones en Cueva Tixi. Pp. 3-7. En: Cueva Tixi: Cazadores y recolectores de las sierras de Tandilia Oriental. 1. Geología, Paleontología y Zooarqueología (Mazzanti D. y C. Quintana, eds.). Publicación Especial 1, Laboratorio de Arqueología, Universidad Nacional de Mar del Plata. [ Links ]
NEVO, E. 1979. Adaptative convergence and divergence of subterranean mammals. Annual Review of Ecology and Systematics, 10:269-308. [ Links ]
PAINE, S. 1975. Partial recovery and sample bias. Pp:7-17. En: Archaeozoological Studies (Clason, A., ed.). Amsterdam. [ Links ]
PIANTANIDA, M., J. CONTRERAS, A. DI GIACOMO y Y. DAVIES. 2000. Una interesante población bonaerense de Ctenomys talarum Thomas 1898 y sus relaciones con las demás poblaciones conocidas de la especie (Rodentia, Ctenomyidae). IX Congreso Iberoamericano de Biodiversidad y Zoología de Vertebrados, pp:176-178. Buenos Aires. [ Links ]
QUINTANA, C. 1994. Sistemática y anatomía funcional del roedor Ctenomyinae Praectenomys (Caviomorpha: Octodontidae) del Plioceno de Bolivia. Revista Técnica de YPFB, 15(1-2):175-185. Bolivia. [ Links ]
QUINTANA, C. 2001a. Composición y cambios en la secuencia faunística. Pp. 37-64. En: Cueva Tixi: cazadores y recolectores de las sierras de Tandilia oriental. I. Geología, Paleontología y Zooarqueología. (Mazzanti y Quintana, eds.). Publicación Especial 1. ARBO-UNMDP. [ Links ]
QUINTANA, C. 2001b. Formación del depósito faunístico. Pp. 123-133. En: Cueva Tixi: cazadores y recolectores de las sierras de Tandilia oriental. I. Geología, Paleontología y Zooarqueología. (Mazzanti y Quintana, eds.). Publicación Especial 1. ARBO-UNMDP. [ Links ]
QUINTANA, C. 2001c. Galea (Rodentia, Caviidae) del Pleistoceno Superior y Holoceno de las sierras de Tandilia Oriental, Provincia de Buenos Aires, Argentina. Ameghiniana, 38(4):399-408. [ Links ]
QUINTANA, C. y D. MAZZANTI. 1996 Secuencia faunística del sitio arqueológico Cueva Tixi (Pleistoceno tardío- Holoceno), provincia de Buenos Aires. Actas VI Jornadas Pampeanas de Ciencias Naturales. Actas pp:187-194. Santa Rosa. Argentina. [ Links ]
QUINTANA, C., F. VALVERDE y A. ALBINO, 2003. Registro de fauna del sitio Cueva El Abra, Tandilia Oriental, Provincia de Buenos Aires. Actas XIII Congreso Nacional de Arqueología Argentina 3:317-324. [ Links ]
REIG, O., J. CONTRERAS y M. PIANTANIDA. 1965. Contribución a la elucidación de la sistemática de las entidades del género Ctenomys (Rodentia, Octodontidae). U.B.A., F.C.E. y N., Contribución Científica, Serie Zoológica, 1:301-352. [ Links ]
REIG, O., C. BUSCH, M. ORTELLS y J. CONTRERAS. 1990. An overview of evolution, systematics, population biology, cytogenetics, molecular biology and speciation in Ctenomys. Pp.71-96. En: Evolution of subterranean mammals at the organismal and molecular levels(Nevo E. y Reig O., eds.) A.R. Liss-Wiley, New York. [ Links ]
REIG, O., A. MASSARINI, M. ORTELLS, M. BARROS, S. TIRANTI S. y F. DYZNCHAUS. 1992. New karyotypes and C-banding patterns of the subterranean rodents of the genus Ctenomys (Caviomorpha, Octodontidae) from Argentina. Mammalia, 56(4):603-623. [ Links ]
RUSCONI, C. 1931. Las especies fósiles del género Ctenomys. Con descripción de nuevas especies. Anales de la Sociedad Científica Argentina, 113:217-236. [ Links ]
TOLOSA, A. 2003. Análisis Citogenético y Morfométrico de poblaciones de roedores subterráneos del género Ctenomys (Rodentia: Octodontidae) de las Provincias de Buenos Aires y de Salta. Tesis, Universidad de Buenos Aires, Facultad de Ciencias Exactas y Naturales, 91 pp. [ Links ]
VASSALLO, A., M. KITTLEIN y C. BUSCH. 1994. Owl predation on two sympatric species of tuco-tucos (Rodentia: Octodontidae). Journal of Mammalogy, 75(3):725-732. [ Links ]
VERZI, D. 2002. Patrones de evolución morfológica en Ctenomyinae (Rodentia: Octodontidae). Mastozoología Neotropical, 9(2):309-328. [ Links ]
WHITE, T. 1953. A method of calculating the dietary percentage of various food animals utilized by aboriginal peoples. American Antiquity, 19:396-398. [ Links ]
ZENUTO, R., A. MALIZIA y C. BUSCH. 1999. Sexual size dimorphism, testes size and mating system in two populations of Ctenomys talarum (Rodentia: Octodontidae). Journal of Natural History, 33(2):305-314. [ Links ]
Recibido 16 marzo 2004.
Aceptación final 8 julio 2004.

