










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkMastozoología neotropical
Print version ISSN 0327-9383On-line version ISSN 1666-0536
Mastozool. neotrop. vol.11 no.1 Mendoza Jan./June 2004
Sigmodontinos (Mammalia, Rodentia) del Holoceno Tardío del Nordeste de la provincia de Buenos Aires (Argentina)
Pablo Teta1, Daniel Loponte2 y Alejandro Acosta2
1Centro Nacional Patagónico, Boulevard Guillermo Brown s/n, 9120 Puerto Madryn, Chubut, Argentina. <antheca@yahoo.com.ar>.2Cátedra de Anatomía Comparada, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Argentina & Instituto Nacional de Antropología y Pensamiento Latinoamericano <danloponte@sinectis.com.ar>; <acosta@mail.retina.ar>
RESUMEN. Se estudiaron dos pequeñas muestras de roedores muroideos fósiles halladas en los sitios arqueológicos La Bellaca sitio 2 (34° 22' 79" S - 58° 39' 53” O) y Las Vizcacheras (34° 16' 81" S - 58° 48' 65” O). Dataciones radiocarbónicas permiten referir estos conjuntos al Holoceno tardío (1-0,6 ka). Los ensambles se hallan ampliamente dominados por el orizomino anfibio Holochilus brasiliensis, con frecuencias subordinadas de los akodontinos Bibimys cf. B.torresi, Necromys sp., Oxymycterus rufus y Scapteromys cf. S. aquaticus-S. tumidus. Las muestras de roedores para el Holoceno tardío del nordeste de Buenos Aires son escasas y de cronología limitada. Las especies registradas en estos agregados sugieren condiciones climáticas similares a las actuales en la región. Los conjuntos de sigmodontinos de La Bellaca sitio 2 y de Las Vizcacheras muestran elementos propios de los ambientes actualmente dominantes en el área: bañados, pequeñas lagunas, pajonales altos, selvas ribereñas y pastizales de gramíneas.
Palabras clave: Región Pampeana, humedales, Sigmodontinae, paleoambientes, tafonomía.
ABSTRACT. Late Holocene sigmodontines (Mammalia, Rodentia) from northeastern Buenos Aires province (Argentina).Remains of fossil muroid rodents found in La Bellaca 2 (34° 22' 79" S - 58° 39' 53” O) and Las Vizcacheras (34° 16' 81" S - 58° 48' 65” O) archeological sites were studied. These samples are Late Holocene in age (1-0,6 ka), according to several radiocarbon dates.The assemblages are largely dominated by the amphibious orizomyne Holochilus brasiliensis with low frequencies of the akodontines Bibimys cf. B.torresi, Necromys sp., Oxymycterus rufus and Scapteromys cf. S. aquaticus-S. tumidus. Samples of fossil muroids from the late Holocene in northeastern Buenos Aires wetlands are extremely isolated and chronologically limited. The scarce species registered in the archeological sites suggest climatic conditions similar to the present ones in the area. The sigmodontine fauna from la Bellaca II and Las Vizcacheras indicates an environment dominated by marshes, small fresh-water bodies, flooded grasslands and grass steppes.
Key words: Pampean Region, wetlands, Sigmodontinae, palaeoenvironments, taphonomy.
INTRODUCCIÓN
En el sector centro-oriental de la región pampeana y estrechamente asociado con el tramo inferior del río Paraná y sección superior del Río de La Plata, se desarrolla un extenso ambiente de humedal (Malvárez, 1997). Conformado en el territorio de la provincia de Buenos Aires por un sector insular en forma de delta y por una faja continental, de aproximadamente 80.000 has, sobre la margen derecha del río Paraná (González, 1995), esta unidad representa un espacio de engranaje entre las selvas marginales subclimáxicas de la Provincia Fitogeográfica Paranaense, los bosques xerófilos de talas de la Provincia del Espinal y las estepas de gramíneas de la Provincia Pampeana (Cabrera, 1968). Esta situación determina, conjuntamente con el régimen pulsátil del eje fluvial Paraná-de La Plata, una elevada complejidad ambiental que se expresa en un mosaico de humedales con características biogeográficas y ecológicas únicas dentro de la Argentina (Malvárez, 1997).
Para la región pampeana se han estudiado con cierto detalle las comunidades de roedores del Plioceno-Holoceno del sudeste de la provincia de Buenos Aires (e.g. Reig, 1978, 1987, 1994; Pardiñas, 1999a, b; Pardiñas, 2000a). En el nordeste de esta provincia, se ha destacado la existencia de muroideos del Pleistoceno temprano-Holoceno en casi una veintena de localidades (Ameghino, 1889, Rusconi, 1931, 1936, Pardiñas y Lezcano, 1995; Pardiñas, 1999a, b). En el área de bajíos ribereños bonaerenses y Delta del Paraná sólo dos yacimientos, en una extensión de terreno que supera las 1.730.000 has, cuentan con análisis de su registro de sigmodontinos (Lezcano, 1991; Pardiñas, 1999a). Únicamente dos taxones, Holochilus brasiliensis y Scapteromys sp., han sido documentados para estos sitios (López et al. 1991; Pardiñas, 1999a).
La presente contribución tiene como objetivo dar a conocer los resultados del estudio taxonómico de dos pequeñas muestras de roedores recuperadas de los sitios arqueológicos La Bellaca sitio 2 (LB II) y Las Vizcacheras (VZ), ambos ubicados sobre la franja de bajíos ribereños inundables del Paraná inferior. Además, se discuten algunos aspectos relativos a la significación paleoambiental y tafonomía de los hallazgos.
ESTRATIGRAFÍA Y CRONOLOGÍA
Los sitios arqueológicos La Bellaca sitio 2 (34° 22' 79" S - 58° 39' 53” O) y Las Vizcacheras (34° 16' 81" S - 58° 48' 65” O) se hallan ubicados en el nordeste de la provincia de Buenos Aires, en los partidos de Tigre y de Escobar, respectivamente (Fig. 1). Al igual que otros yacimientos emplazados en el área, la secuencia de estos depósitos se inicia con el suelo actual, compuesto por un horizonte “A” (nivel A), cuya potencia es de aproximadamente 45 a 50 cm. Seguidamente, se observa un nivel “C”, compuesto mayormente por arena con cantidades subordinas de arcilla. Este último es un sedimento plástico, que ocasionalmente presenta valvas de moluscos fluviales en posición de vida y que representa, dentro de la “Unidad Deltaica” (sensu Parker y Marcolini, 1992), un período de agregación fluvial. Entre ambas unidades se reconoce un nivel intermedio, denominado A/C, que varía entre 2 y 10 cm. Los hallazgos arqueológicos se registran estrictamente en el nivel estratigráfico “A”. Una muestra ósea recolectada en el sector medio del nivel A del depósito de La Bellaca sitio 2 arrojó una antigüedad corregida de 680 ± 80 años 14C AP (LATYR, LP-1263). Para el sitio Las Vizcacheras se cuenta con dos fechados. El primero corresponde a una muestra ósea recuperada en el sector medio del nivel A, que arrojó una edad de 1090 ± 40 años 14C AP (Beta 148.237). El segundo fue efectuado sobre restos carbonizados de semillas provenientes del sector más profundo del nivel A. Su antigüedad fue fijada en 1070 ± 60 años 14C AP (LATYR, LP-1401) (Loponte y Acosta, en prensa).

Fig. 1.Ubicación geográfica de los sitios arqueológicos Las Vizcacheras y La Bellaca sitio 2 (provincia de Buenos Aires, Holoceno Tardío).
Geographic location of Las Vizcacheras and La Bellaca 2 archaeological sites (Buenos Aires Province, Late Holocene).
MARCO AMBIENTAL
Los sitios arqueológicos La Bellaca sitio 2 y Las Vizcacheras se hallan ubicados sobre la franja inundable de bajíos ribereños que se extiende sobre la margen derecha del río Paraná. Esta unidad, que se prolonga entre la costa del río y las barrancas retrocedentes modeladas por la ingresión marina del Holoceno medio (Iriondo, 1988), posee, en promedio, cotas de 2 m de altura y se halla mayormente ocupada por llanuras bajas de drenaje impedido que dan lugar a extensos bañados y pequeñas lagunas (Bonfilds, 1962; Cavalloto et al., 1999). Los tipos de vegetación natural más frecuentes en el área son las praderas de herbáceas altas, tanto graminiformes (e.g. Scirpus giganteus) como equisetoides (e.g. S. californicus) (Malvárez, 1997). Los puntos más altos en esta unidad están representados por albardones de forma subcircular o longitudinal que quedan regularmente por encima de las inundaciones que periódicamente se presentan en el área. Ubicados a lo largo de ríos y en los bordes de bañados y lagunas internas, le corresponde a los mismos una fisonomía de selva ribereña empobrecida, que en la actualidad se halla mayormente desplazada por la forestación con frutales y salicáceas (Malvárez, 1997). Hacia el sur y el oeste se desarrolla una extensa estepa de gramíneas, conocida como Pampa ondulada, que posee cotas que superan los 20 m snm y que se continúa ampliamente en el sector central de la llanura Pampeana (Cabrera, 1968).
Las condiciones climáticas medias para el área son templadas, con temperaturas promedio de aproximadamente 23 ºC para enero y de 10 ºC para julio. Las precipitaciones totales anuales se hallan en el orden de los 1000 mm y se distribuyen en forma pareja a lo largo de todo el año (Burgos, 1968). Como característica propia de esta unidad, el sistema de humedales de la región genera un efecto de modificación sobre las principales variables climáticas que modera tanto las temperaturas extremas como la deficiencia hídrica temporaria. Esta situación origina condiciones más parecidas a las subtropicales húmedas que a las templado sub-húmedas de la zona circundante (Malvárez, 1997).
MATERIALES Y MÉTODOS
Se estudiaron 52 restos cráneo-mandibulares recuperados durante la excavación y el tamizado de los sedimentos de los sitios arqueológicos La Bellaca sitio 2 y Las Vizcacheras. La determinación taxonómica fue realizada a partir de la consulta de bibliografía (Pardiñas, 1996; Pardiñas y Galliari, 1998; Galliari y Pardiñas, 2000) y de la comparación con ejemplares actuales depositados en el Museo Argentino de Ciencias Naturales Bernardino Rivadavia. Los materiales estudiados se encuentran depositados en el Instituto Nacional de Antropología y Pensamiento Latinoamericano (INAPL).
El criterio taxonómico adoptado corresponde al discutido por Galliari et al (1996) con algunas modificaciones, a nivel de la ubicación tribal de algunos géneros, acordes con la literatura posterior (Smith y Patton, 1999; D´Elia, 2003.
En la descripción de las estructuras dentarias se siguió la terminología propuesta por Reig (1977) y, subsidiariamente, por Hershkovitz (1993). Las medidas, todas expresadas en milímetros, fueron tomadas con calibre manual (con precisión de 5 centésimas) y ocular micrométrico adosado a lupa binocular. Las longitudes para las series yugales son alveolares y los largos y anchos para los molares corresponden a los valores máximos.
Como parámetro actualístico de comparación para la realización de inferencias paleoambientales fueron utilizados los datos de varias muestras de egagrópilas referibles a Tyto alba (Aves; Tytonidae) de diversas localidades del nordeste bonaerense (Babarskas et al., 2003: Tabla 5).
Abreviaturas utilizadas en el texto y en las tablas: MD: maxilar derecho; MI: maxilar izquierdo; HD: hemimandíbula derecha; HI: hemimandíbula izquierda; M1, M2, M3, m1, m2 y m3: primero, segundo y tercer molar superior e inferior respectivamente; ka = 103años; MNI = número mínimo de individuos; NISP = número de especímenes identificados.
RESULTADOS
En líneas generales, ambas muestras se hallan signadas por una marcada dominancia de Holochilus brasiliensis y una moderada proporción de Necromys sp. En La Bellaca sitio 2 también fueron registrados Oxymycterus rufus y Scapteromys sp. El elenco de roedores sigmodontinos de Las Vizcacheras se completa, por otra parte, con las presencias de Scapteromys sp. y Bibimys cf. B. torresi (Tabla 1). Otros roedores detectados en estos yacimientos son los caviomorfos Cavia aperea, Ctenomys cf. C. talarum y Myocastor coypus y el múrido exótico Rattus cf. R. norvegicus. La presencia de restos de esta última especie en niveles >0,5 ka es claramente intrusiva y posiblemente se vincule con procesos de migración vertical descendente o con su introducción por causas eto-ecológicas (e.g. construcción de madrigueras).
Tabla 1
NISP para los roedores registrados en los agregados de La Bellaca sitio 2 y Las Vizcacheras (Holoceno tardío, nordeste de Buenos Aires)
NISP for registered rodents in Las Vizcacheras and La Bellaca 2 fossil assemblages (Buenos Aires Province, Late Holocene).
PALEONTOLOGÍA SISTEMÁTICA y DESCRIPTIVA
Orden Rodentia Bowdich, 1821
Familia Muridae Illiger, 1815
Subfamilia Sigmodontinae Wagner, 1843
Tribu Akodontini Vorontzov, 1959
Bibimys cf. B. torresi Massoia, 1979
Fig. 4a; Tabla 3
Descripción — La rama horizontal de la mandíbula es robusta y algo corta en sentido ántero-posterior. La base del incisivo no forma una proyección capsular notable, sino una repisa o plataforma extendida, suavemente arqueada, que termina en la parte posterior del proceso condilar. El proceso coronoides es corto y se inclina levemente hacia atrás. Los molares presentan un desgaste de tipo crestado y son moderadamente hipsodontes. Las cúspides labiales y linguales se muestran apenas alternantes. El m1 y m2 presentan un fléxido anteromediano bien marcado y el mesolófido fusionado con el entolofúlido. El m3 es subrectangular, con el hipofléxido largo y oblicuo.
Comentarios taxonómicos — La taxonomía del género Bibimys ha sido recientemente revisada por D´Elia et al. (en prensa). Aunque la validez específica de B. torresi ha sido preliminarmente aceptada por estos autores, como éstos mismos lo advierten, la evidencia morfológica y molecular sugiere una potencial conespecificidad con B. labiosus.
Distribución y microhábitat — Bibimys torresi se halla actualmente restringido al Delta del Paraná y áreas de influencia. El registro fósil de este múrido para los últimos 120 ka en la Región Pampeana muestra, por otra parte, una geonemia que se habría extendido por lo menos hasta el sudeste de la provincia de Buenos Aires (Pardiñas, 1995, 1996, 1999a, b, D´Elia et al., en prensa). Los ambientes de trampeo sugieren una relación estrecha con el agua (Massoia, 1979). Bianchini y Delupi (1993) citan la captura de esta especie en comunidades de seibales y en juncales de espadañas, ciperáceas y gramíneas.
Necromys sp.
Descripción — La rama mandibular es relativamente alta. La cresta masetérica está bien marcada y alcanza anteriormente el nivel del procíngulo del m1. La proyección capsular forma un abultamiento notable. Los molares son aterrazados y de cúspides opuestas. El cíngulo anterolabial está bien desarrollado. Los meta y protofléxido están orientados oblicuamente. El hipofléxido es transverso y de fondo amplio. El posterofléxido está bien marcado y define un posterolófido grande. En el m2 se evidencia claramente un protofléxido bien marcado.
Comentarios taxonómicos — Para la Región Pampeana han sido reconocidas dos especies del género Necromys: N. obscurus, con poblaciones en el litoral atlántico entre las localidades de Santa Clara del Mar y Necochea y en el extremo austral de las sierras de Tandilia y N. benefactus, que habita en el noroeste, centro-oeste y sudoeste del territorio bonaerense, alcanzando por el este el sistema serrano de Tandilia (Galliari y Pardiñas, 2000). Para el Holoceno tardío del nordeste de la provincia de Buenos Aires, en los sitios arqueológicos La Norma y La Higuera (partido de Berisso), se han registrado ejemplares con características morfométricas intermedias entre N. obscurus y N. benefactus (Pardiñas, 1999a). En función de esta situación, y considerando el rango de variabilidad morfológica reconocida para algunas especies del género, se opta por el uso de una taxonomía abierta hasta tanto los hallazgos aquí reportados puedan ser adecuadamente contextuados.
Distribución y microhábitat — Las especies del género Necromys ocupan con preferencia pastizales y bordes de campos cultivados (Mills et al., 1991). N. benefactus muestra, por otra parte, una relación estrecha con un anillo de menores precipitaciones por el oeste y sudoeste y con las formaciones florísticas xeromórficas del distrito del Tala por el norte y nordeste (Pardiñas, 1999a; Galliari y Pardiñas, 2000). La distribución actual del género en la provincia de Buenos Aires contrasta parcialmente con el registro fósil que evidencia una geonemia más extendida durante los últimos 10 ka (Pardiñas, 1999a; Galliari y Pardiñas, 2000).
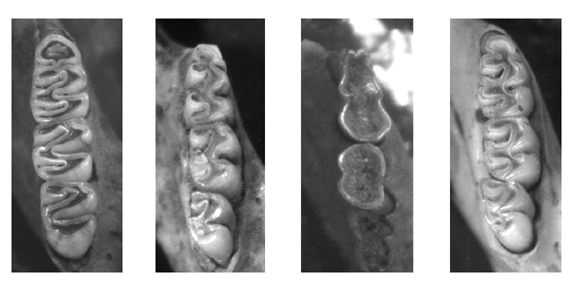
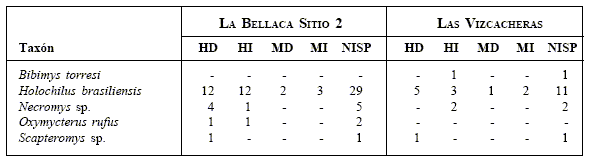
Fig. 2.Vistas oclusales de los molares derechos (entre paréntesis se indica la longitud de los elementos representados) de algunos sigmodontinos de los yacimientos Las Vizcacheras y La Bellaca sitio 2 (provincia de Buenos Aires, Holoceno Tardío). De izquierda a derecha: LB-1-60-65-A/Cb, Holochilus brasiliensis, m1-3 (8.25); LBII-1-35-40, Necromys sp., m1-3 (5.50); LBII-2-35-40: Oxymycterus rufus, m1-2 (4.3) y VZ-2-50-55, Scapteromys sp., m1-3 (6.6).
Oclusal views (the length of the teeth are given between parentheses) for some sigmodontines from Las Vizcacheras and La Bellaca 2 (Buenos Aires Province, Late Holocene). From left to right: Holochilus brasiliensis, Necromys sp., Oxymycterus rufus, and Scapteromys sp.
Tabla 2
Medidas de los molares y series dentarias inferiores en Oryzomyini y Akodontini de los agregados de La Bellaca sitio 2 y Las Vizcacheras (Holoceno tardío, nordeste de Buenos Aires). Se indica, cuando corresponde, media, desvío estándar y, entre paréntesis, número de ejemplares medidos.
Measurements of the molars and lower toothrows in Oryzomyini and Akodontini from Las Vizcacheras and La Bellaca 2 fossil assemblages (Buenos Aires Province, Late Holocene). Media, standard deviation, and sample size (between parentheses) are given.
Oxymycterus rufus Fischer (1814)
Descripción — La rama horizontal de la mandíbula es grácil y la cresta masetérica se presenta como un abultamiento de bordes suaves, cuya rama inferior alcanza anteriormente el nivel medio del m1. Los molares son hipsodontes y de corona crestada. En BII-2-35-40 el m1 muestra un fléxido anteromediano poco profundo, pero bien definido y un diseño oclusal totalmente simplificado por el desgaste. En LBII-1-35-40 el m3 es proporcionalmente largo y estrecho. El mesofléxido se opone, en dirección transversa, al hipofléxido, confiriéndole al diente una forma de 8 (ocho).
Comentarios taxonómicos — A pesar de los numerosos aportes realizados en los últimos años (Hershkovitz, 1994; Oliveira, 1998; Hoffman et al., 2002), la taxonomía del género Oxymycterus dista de ser clara. En una reciente revisión, Oliveira (1998) distingue como pertenecientes a O. rufus a las poblaciones de Entre Ríos, Buenos Aires, Santa Fe y Córdoba. Sin embargo, persiste cierta controversia en cuanto a la ubicación de la localidad tipo de este akodontino, restringida por algunos autores a la provincia de Entre Ríos (Musser y Carleton, 1993; Hershkovitz, 1994; Oliveira, 1998). Una re-evaluación de la obra literaria de Azara (1801; 1802), sugiere, por el contrario, que el ejemplar sobre el cual este autor basa la descripción de su “Rat Cinquiéme ou Rat Roux” y sobre el que se sustenta el Mus rufus de Fischer, habría sido coleccionado en el departamento de Misiones, Paraguay (Contreras y Teta, 2003). La validez de O. platensis (con localidad tipo en Río Santiago, Ensenada), promovida por Reig (1987) y por Hershkovitz (1994), y el estatus de las poblaciones serranas de Córdoba, San Luis y sur bonaerense, usualmente incluidas en la sinonimia de O. rufus, requiere de nuevas aproximaciones (Galliari et al., 1996).
Distribución y microhábitat — Oxymycterus rufus presenta poblaciones en el este de Paraguay y Brasil y en el nordeste, centro-este y centro de Argentina (Redford y Eisenberg, 1992). En el Delta del Paraná y áreas adyacentes es una especie frecuente en pastizales, cortaderales y microambientes arbustivos de Baccharis spicata (Cueto et al., 1995a).
Tabla 3
Medidas de los molares y series dentarias inferiores en Akodontini de los agregados de La Bellaca sitio 2 y Las Vizcacheras (Holoceno tardío, nordeste de Buenos Aires). Se indica, cuando corresponde, media, desvío estándar y, entre paréntesis, número de ejemplares medidos.
Measurements of the molars and lower toothrows in Akodontini from Las Vizcacheras and La Bellaca 2 fossil assemblages (Buenos Aires Province, Late Holocene). Media, standard deviation, and sample size (between parentheses) are given.

Scapteromys sp.
Descripción — La mandíbula es alargada y robusta. La cresta masetérica se halla moderadamente marcada. En el ejemplar VZ-2-50-55 los dientes yugales presentan el mesolófido largo y de posición oblicua. El m1 muestra un protofléxido rudimentario. El hipofléxido es largo y se orienta transversalmente. LBII-1-20-25 muestra un desgaste mayor de la superficie de oclusión molar, con pliegues apenas marcados, que lo acercan a la condición presentada por el individuo ilustrado por Massoia (1981) con el número CEM 987. En el m1 el mesolófilo y el metacónido se hallan fusionados. El m2 tiene forma de S, con el hipofléxido y el mesofléxido bien marcados.
Comentarios taxonómicos — Análisis de variación de secuencias de ADN mitocondrial sugieren que el género Scapteromys, largamente considerado monotípico, incluiría por lo menos dos clados bien diferenciados (D´Elia, 2002, D´Elia y Pardiñas, en prensa). Sobre la base de estos resultados y de una re-evaluación de la evidencia morfológica y cromosómica previa, D´Elia (2002) y D´Elia y Pardiñas (en prensa) han promovido el reconocimiento específico de las formas nominales S. aquaticus (con poblaciones en la Argentina y Paraguay) y S. tumidus (restringida al Uruguay y, posiblemente, al sur de Brasil). Las diferencias entre ambas entidades son difíciles de evaluar sobre la base de restos fragmentarios, razón que determina la asignación de los materiales al grupo Scapteromys cf. S. aquaticus - S. tumidus (Pardiñas, 1999a).
Distribución y microhábitat — El género Scapteromys se distribuye desde el norte de Paraguay y el sur de Brasil, hasta el Uruguay y el nordeste de la provincia de Buenos Aires (Massoia y Fornes, 1964; Hershkovitz, 1966; D´Elia, 2002). En la Argentina, Scapteromys aquaticus ha sido registrada en las provincias de Chaco, Formosa, Santa Fe, Corrientes, Entre Ríos y Buenos Aires (Massoia y Fornes, 1964). En Delta del Paraná Cueto et al. (1995b) han destacado su captura en cortaderales y pajonales altos de Scirpus giganteus y Panicum grumosum en terrenos bajos y anegadizos.
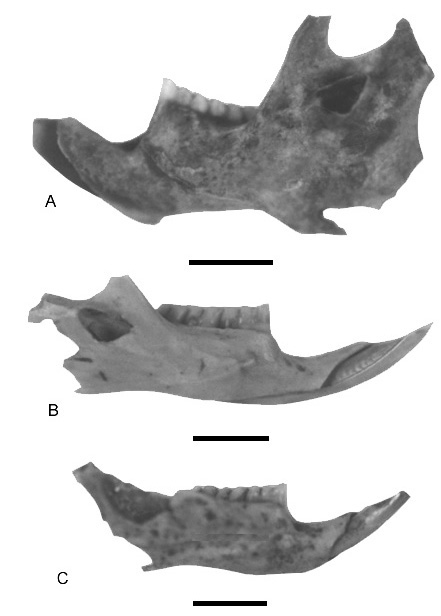
Fig. 3.Vistas labiales de mandíbulas de algunos sigmodontinos de los yacimientos Las Vizcacheras y La Bellaca sitio 2 (provincia de Buenos Aires, Holoceno Tardío). A, Holochilus brasiliensis, hemimandíbula izquierda (LB-1-60-65-A/Cb). B, Scapteromys sp., hemimandíbula derecha (VZ-2-50-55). C, Necromys sp., hemimandíbula derecha (LBII-1-35-40).
Labial views of mandibles of some sigmodontines from Las Vizcacheras and La Bellaca 2 (Buenos Aires Province, Late Holocene). A, Holochilus brasiliensis, left mandible. B, Scapteromys sp., right mandible. C, Necromys sp., right mandible.
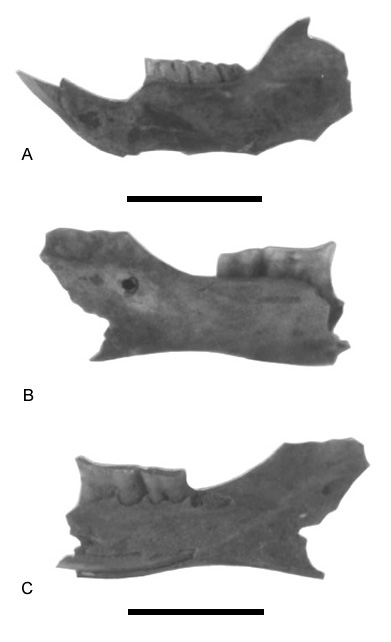
Fig. 4.Vistas labiales y linguales de mandíbulas de algunos sigmodontinos de los yacimientos Las Vizcacheras y La Bellaca sitio 2 (provincia de Buenos Aires, Holoceno Tardío). A, Bibimys torresi, hemimandíbula izquierda (VZ-2-30-35b). B, Oxymycterus rufus, hemimandíbula derecha (LBII-2-35-40), vista labial. C, vista lingual del ejemplar anterior.
Labial and lingual views of mandibles of some sigmodontines from Las Vizcacheras and La Bellaca 2 (Buenos Aires Province, Late Holocene). A, Bibimys torresi, left mandible. B, Oxymycterus rufus, right mandible, labial view. C, Oxymycterus rufus, right mandible lingual view.
Tribu Oryzomyini Vorontzov, 1959
Holochilus brasiliensis Desmarest (1819)
Descripción — El M1, M2 y M3 presentan el mesolofo moderadamente desarrollado. El M2 es de menor longitud que el M3. La rama horizontal de la mandíbula es robusta, con el proceso mentoniano bien desarrollado. Las crestas masetéricas superior e inferior están bien marcadas y suelen reunirse, formando un solo reborde que se extiende entre la raíz posterior del m1 y el foramen mental. El m1 muestra una fosétida antero-interna grande y alargada transversalmente. El metafléxido está bien desarrollado, alcanzando la línea media del diente. El proto e hipocónido presentan áreas subiguales de contorno casi triangular. El m2 es subcuadrangular en vista oclusal. El protofléxido forma una leve muesca sobre el borde labial. El hipofléxido se orienta transversalmente con el fondo opuesto al entolófido. El m3 es claramente sigmoide.
Comentarios taxonómicos — La taxonomía del género Holochilus presenta todavía numerosos interrogantes (Voss y Carleton, 1993; Pardiñas y Galliari, 1998). Al menos tres morfotipos, asimilables con las formas nominales H. brasiliensis, H. chacarius y H. sciureus, pueden ser reconocidos sobre la base de diferencias en la morfología cráneo-dentaria (Massoia, 1976; Voss y Carleton, 1993; Pardiñas y Galliari, 1998). En la propuesta de Massoia (1976), Holochilus brasiliensis incluye como subespecies a H. b. brasiliensis, H. b. vulpinus y H. b. darwini.
Distribución y microhábitat — Holochilus brasiliensis es una forma anfibia que ocupa, en áreas intertropicales a templadas, cuerpos de agua dulce, lénticos o lóticos, con abundante vegetación palustre. En Sudamérica, esta especie se distribuye desde el este de Paraguay y sur de Brasil hasta el Uruguay y la provincia de Buenos Aires (Hershkovitz, 1955, Massoia, 1976).
DISCUSIÓN Y CONCLUSIONES
Tafonomía
Los conjuntos arqueofaunísticos del nordeste bonaerense exhiben, en líneas generales, una alta integridad, que puede relacionarse, en buena medida, con un rápido enterramiento de los huesos en un ambiente de baja energía y con un desarrollo significativo de la cobertura vegetal (Acosta y Loponte, 1992; Loponte y Acosta, en prensa).
Los agregados de roedores sigmodontinos de los sitios La Bellaca sitio 2 y Las Vizcacheras se caracterizan por una baja riqueza y diversidad de especies. Esta situación sugiere, conjuntamente con una baja densidad de hallazgos, un proceso de incorporación fundamentalmente por causas eto-ecológicas (cf. Pardiñas, 2000b). Los restos estudiados muestran un muy buen estado de conservación y estadios muy bajos de meteorización. Las improntas de raíces, en forma de marcas dendríticas poco profundas, son abundantes en la mayoría de los especímenes (50-60%). No se han registrado evidencias de rodamiento o de corrosión estomacal. La abundancia de partes esqueletarias para las muestras de La Bellaca sitio 2 y Las Vizcacheras presenta máximos en los elementos mandíbula y fémur, con participación moderada de los huesos innominado, tibia y escápula (Tabla 4). La fragmentación de los restos es baja a moderada (Tabla 4).
Tabla 4
Categorías de fragmentación de los principales elementos óseos post-craneanos de los agregados de La Bellaca sitio 2 y Las Vizcacheras (Holoceno tardío, nordeste de Buenos Aires).
Fragmentation categories of main skeletal elements of Las Vizcacheras and La Bellaca 2 fossil assemblages (Buenos Aires Province, Late Holocene).

Paleoambientes
Con excepción de Necromys sp., todas las especies registradas en La Bellaca sitio 2 y Las Vizcacheras habitan actualmente en el nordeste bonaerense. Se trata mayormente de taxones de clara afinidad subtropical, cuyo rango de distribución abarca desde el sur de Brasil y Paraguay hasta el Uruguay y la provincia de Buenos Aires.
Holochilus brasiliensis y Scapteromysaquaticus son dos elementos frecuentes en los ambientes palustres del litoral parano-platense. La elevada frecuencia del primero es consistente con un ambiente dominado por amplios espacios de agua emergida bajo condiciones climáticas templado-cálidas (Hershkovitz, 1955; Carleton y Olson, 1999). La segunda es una especie abundante en el Delta del Paraná y las formaciones ribereñas higrófilas y selva marginal (Massoia y Fornes, 1964; Pardiñas, 1999a).
Bibimystorresi se halla actualmente restringido al Delta del Paraná, donde ocupa pastizales periselváticos y comunidades higrófilas (Bianchini y Delupi, 1993; Pardiñas, 1996). El registro actual más austral para esta especie corresponde a la Reserva Natural Estricta Otamendi, ubicada a 10 km al NO de Las Vizcacheras. Oxymycterus rufus es una especie abundante en el nordeste bonaerense. Cueto et al. (1995a) destacan su captura en cortaderales y en microambientes arbustivos de Baccharis spicata. La significación paleoambiental de Necromys resulta, en virtud de la indefinición taxonómica, algo más difícil de precisar. En la Región Pampeana, las especies de este género son frecuentes en pastizales naturales y bordes de campos cultivados (Mills et al., 1991; Galliari y Pardiñas, 2000).
En síntesis, los conjuntos de sigmodontinos de La Bellaca sitio 2 y de Las Vizcacheras son coincidentes en señalar un ambiente dominado por bañados, cortaderales, pajonales altos y pastizales de gramíneas. La presencia de Bibimys cf. B. torresi y Scapteromys sp. podría, en este contexto, ser indicativa de una mayor extensión de la selva marginal subclimáxica y comunidades asociadas hacia 1-0.6 ka. Actualmente, la expresión de esta unidad en el área de bajíos ribereños se halla fuertemente restringida por la deforestación y el reemplazo con frutales y salicáceas (Malvárez, 1997). En efecto, sobre tres muestras actuales de egagrópilas coleccionadas en las proximidades del Río Luján, 5 km al NO de estos sitios, no se han registrado ejemplares de estas dos especies (Babarskas et al., 2003). Alternativamente, la extensión austral de la geonemia de Bibimys podría vincularse con un aumento de la temperatura hacia el Holoceno más tardío (Pardiñas, 1999a, b, 2000a). Otros mamíferos registrados en estos yacimientos (e.g. Lutreolina crassicaudata, Myocastor coypus, Blastocerus dichotomus, Ozotoceros bezoarticus, véase Loponte y De Santis, 1995; Loponte y Acosta, en prensa) son consistentes con esta interpretación y coinciden en indicar condiciones ambientales similares a las actuales, previas a la etapa más profunda del impacto antrópico (Salemme, 1987; Lezcano, 1991; Loponte y De Santis, 1995; Pardiñas, 1999a).
AGRADECIMIENTOS
Ulyses Pardiñas leyó críticamente el manuscrito aportando sugerencias e información inédita. Luciano J. De Santis realizó comentarios a una primera versión de este trabajo. María Guiomar Vucetich y dos evaluadores anónimos contribuyeron, con sus observaciones, a mejorar la claridad de este trabajo. Luciano Pafundi participó del análisis de las muestras.
LITERATURA CITADA
ACOSTA, A. y D. LOPONTE. 1992. Una aproximación a los procesos tafonómicos en el área norte de la Provincia de Buenos Aires. Palimpsesto, Revista de Arqueología, 2:21-28. [ Links ]
AMEGHINO, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias en Córdoba, 6:1-1027 y Atlas: 98 pls. [ Links ]
AZARA, F. de. 1801. Essais sur l´Historie Naturelle des Quadrupédes de la Province du Paraguay. Tome Second. Imprière C. Pougens, París, pp. 1-449. [ Links ]
AZARA, F. de. 1802. Apuntamientos para la Historia Natural de los Quadrúpedos del Paraguay y Río de La Plata. Tomo segundo. Imprenta de la Viuda de Ibarra, Madrid, pp. 1-328. [ Links ]
BABARSKAS, M., E. HAENE y J. PEREIRA. 2003. Aves de la Reserva Natural Otamendi. Pp. 47-113. En: Fauna de Otamendi (Haene, E. y J. Pereira, eds.). Temas de Conservación y Naturaleza, Monografía de Aves Argentinas, 3. [ Links ]
BIANCHINI, J.J. y H. DELUPI. 1993. Mammalia. Pp. 1-79. En: Fauna de agua dulce de la República Argentina (Zulma A. de Castellanos, dir.). 44, Fascículo 2 (Actualización), La Plata. [ Links ]
BONFILDS, 1962. Los suelos del Delta del Río del Paraná. Factores generadores, clasificación y uso. Revista de Investigaciones Agrícolas, INTA, 16(3):1-257. [ Links ]
BURGOS, J.J. 1968. El clima de la provincia de Buenos Aires en relación con la vegetación natural y el suelo. Pp. 33-99. En: Flora de la provincia de Buenos Aires (Cabrera, A.L., ed.), INTA, Colección Científica, 4(1). [ Links ]
CABRERA, A.L. 1968. Vegetación de la Provincia de Buenos Aires. Pp. 101-120. En: Flora de la provincia de Buenos Aires (Cabrera, A.L., ed.), INTA, Colección Científica, 4(1). [ Links ]
CARLETON, M.D. y S.L. OLSON. 1999. Amerigo Vespucci and the Rat of Fernando de Noronha: a new genus and species of Rodentia (Muridae: Sigmodontinae) from a volcanic Island off Brazil´s continental shelf. American Museum Novitates, 3256:1-59. [ Links ]
CAVALLOTO, J.L., R. VIOLANTE y G. PARKER. 1999. Historia evolutiva del Río de la Plata durante el Holoceno. XIV Congreso Geológico Argentino, Actas, (1):508-515. [ Links ]
CONTRERAS, J.R. y P. TETA. 2003. Acerca del estatus taxonómico y de la localidad típica de Oxymycterus rufus (Fisher, 1814) (Rodentia: Muridae: Sigmodontinae). Nótulas Faunísticas, 14:1-5 [ Links ]
CUETO, V.R., M.J. PIANTANIDA y M. CAGNONI. 1995a. Population demography of Oxymycterus rufus (Rodentia: Cricetidae) inhabiting a patchy environment of the Paraná river, Argentina. Acta Theriologica, 40(2):123-130. [ Links ]
CUETO, V.R., M. CAGNONI y M.J. PIANTANIDA. 1995b. Habitat use of Scapteromys tumidus (Rodentia: Cricetidae) in the delta of the Paraná river, Argentina. Mammalia, 59(1):25-34. [ Links ]
D´ELÍA, G. 2002. Testing Patterns and Processes of Diversification of a South American Group of Land Mammals (Rodentia, Cricetidae, Sigmodontinae). Ph.D. Dissertation, The University of Michigan, Ann Arbor, pp. i-xi + 1-174. [ Links ]
D´ELÍA, G. 2003. Phylogenetics of Sigmodontinae (Rodentia, Muroidea, Cricetidae), with special reference to the akodont group, and with additional comments on historical biogeography. Cladistics, 19:307-323. [ Links ]
D´ELÍA, G. y U.F.J. PARDIÑAS. En prensa. Systematics of Argentinean, Paraguayan, and Uruguayan swamp rats of the genus Scapteromys (Rodentia, Cricetidae, Sigmodontinae). Journal of Mammalogy. [ Links ]
D´ELÍA, G., U.F.J. PARDIÑAS y P. MYERS. En prensa. An introduction to the genus Bibimys (Rodentia: Sigmodontinae): Phylogenetic position and alpha taxonomy. En: Mammalian diversification: from population genetics to Biogeography (Lacey, E. y P. Myers, eds.). Berkeley, University of California Press. [ Links ]
GALLIARI, C.A. y U.F.J. PARDIÑAS. 2000. Taxonomy and distribution of the sigmodontine rodents of genus Necromys in central Argentina and Uruguay. Acta Theriologica, 45(2):211-232. [ Links ]
GALLIARI, C.A., U.F.J. PARDIÑAS y F.J. GOIN. 1996. Lista comentada de los mamíferos argentinos. Mastozoología Neotropical, 3(1):39-61. [ Links ]
GONZÁLEZ, M.A. 1995. Diagnóstico ambiental de la provincia de Buenos Aires. Tomo I: aspectos básicos. Ediciones Banco Provincia, Buenos Aires. [ Links ]
HERSHKOVITZ, P. 1955. South American marsh rats genus Holochilus, with a summary of sigmodont rodents. Fieldiana, Zoology, 37:639-673. [ Links ]
HERSHKOVITZ, P. 1966. South American swamp and fossorial rats of the Scapteromyine group (Cricetinae, Muridae) with comments on the glans penis in Murid taxonomy. Sond. Zeitschrift für Säugetierkunde Bd., 2:81-149. [ Links ]
HERSHKOVITZ, P. 1993. A new central brazilian genus and species of sigmodontine rodent (Sigmodontinae) transitional between akodonts and oryzomyines with a discussion of muroid molar morphology and evolution. Fieldiana, Zoology, new series, 75:1-18. [ Links ]
HERSHKOVITZ, P. 1994. The Description of a New Species of South American Hocicudo, or Long Nosed Mouse, Genus Oxymycterus (Sigmodontinae, Muroidea), with a Critical Review of the Generic Context. Fieldiana, Zoology, new series, 79:1-43. [ Links ]
HOFFMAN, F.G., E.P. LESSA y M.F. SMITH. 2002. Systematics of Oxymycterus with description of a new species from Uruguay. Journal of Mammalogy, 83(2):408-420. [ Links ]
IRIONDO, M. 1988. Llanura aluvial del Río Paraná. Simposio Internacional sobre el Holoceno en América del Sur. INQUA-CADINQUA. Guía de campo Nº 4, pp. 1-20. [ Links ]
LEZCANO, M.J. 1991. Análisis preliminar de los restos faunísticos hallados en el sitio Laguna Grande, partido de Campana, provincia de Buenos Aires. Boletín del Centro, 2:95-104. [ Links ]
LÓPEZ, J.A., E. MASSOIA y A. MORICI. 1991. Fauna asociada a restos arqueológicos en el sitio "Las Vizcacheras", costa del río Luján, partido de Escobar, provincia de Buenos Aires. Boletín Científico, Asociación para la Protección de la Naturaleza, 14:2-5. [ Links ]
LOPONTE, D. y A. ACOSTA. En prensa. Arqueología de cazadores-recolectores del sector centro-oriental de la Región Pampeana. Runa, Facultad de filosofía y Letras. [ Links ]
LOPONTE D. y L. DE SANTIS. 1995. Variabilidad ambiental y problemas metodológicos: el caso del registro arqueofaunístico en el norte de la provincia de Buenos Aires. I Jornadas Chivilcoyanas en Ciencias Sociales y Naturales, Actas: 137-143. [ Links ]
MÁLVAREZ, A.I. 1997. El Delta del Río Paraná como mosaico de humedales. Pp. 1-49. En: Tópicos sobre humedales Sudamericanos (Malvárez, A.I. y P. Kandus, eds.). Facultad de Ciencias Exactas y Naturales, Departamento de Ciencias Biológicas, UBA. [ Links ]
MASSOIA, E. 1976. Mammalia. En: Ringuelet, R. (dir.), Fauna de Agua Dulce de la República Argentina, Fundación editorial Ciencia y Cultura, Buenos Aires, 44:1-128. [ Links ]
MASSOIA, E. 1979. Descripción de un género y especie nuevos: Bibimys torresi (Mammalia-Rodentia-Cricetidae-Sigmodontinae-Scapteromyini). Physis, C, 38(95):1-7. [ Links ]
MASSOIA, E. 1981. Variabilidad morfológica de los molares de Scapteromys del Delta del Paraná (Mammalia, Rodentia, Cricetidae, Sigmodontinae). Historia Natural, 2(9):57-63. [ Links ]
MASSOIA, E. y A. FORNES. 1964. Notas sobre el género Scapteromys (Rodentia-Cricetidae). I. Sistemática, distribución geográfica y rasgos etoecológicos de Scapteromys tumidus (Waterhouse). Physis, 24(68):279-297. [ Links ]
MILLS, J.N., B.A. ELLIS, K.T. MCKEE, J.I. MAISTEGUI y J.E. CHILDS. 1991. Habitat associations and relative densities of rodent populations in cultivated areas of central Argentina. Journal of Mammalogy, 72:470-479. [ Links ]
MUSSER, G.G. y M.D. CARLETON. 1993. Family Muridae. Pp. 501-755. En: Mammal Species of the World (Wilson D.E. y A.M. Reeder, eds). Second Edition. Smithsonian Inst. Press, Washington & Londres. [ Links ]
OLIVEIRA, J. 1998. Morphometric assesment of species group in the South American rodent genus Oxymycterus Sigmodontinae, with taxonomic notes based on the analysis of the type material. Ph.D. Dissertation, TexasTechUniversity, Lubbock. [ Links ]
PARDIÑAS, U.F.J. 1995. Novedosos cricétidos (Mammalia, Rodentia) en el Holoceno de la Región Pampeana, Argentina. Ameghiniana, 32(2):197-203. [ Links ]
PARDIÑAS, U.F.J. 1996. El registro fósil de Bibimys Massoia, 1979 (Mammalia, Rodentia). Consideraciones preliminares sobre la distribución de los Scapteromyini (Cricetidae, Sigmodontinae) en la región pampeana durante el Pleistoceno-Holoceno. Mastozoología Neotropical, 3(1):15-38. [ Links ]
PARDIÑAS, U.F.J. 1999a. Los roedores muroideos del Pleistoceno tardío-Holoceno en la región pampeana (sector este) y Patagonia (República Argentina): aspectos taxonómicos, importancia bioestratigráfica y significación paleoambiental. Tesis doctoral, Facultad de Ciencias Naturales y Museo, Universidad Nacional La Plata, pp. i-ix +1-283. [ Links ]
PARDIÑAS, U.F.J. 1999b. Fossil murids: taxonomy, paleoecology, and paleoenvironments. En: Quaternary of South America and Antarctic Peninsula (Tonni, E.P. y A.L. Cione, eds.). 12:225-254. [ Links ]
PARDIÑAS, U.F.J. 2000a. Micromamíferos y paleoambientes del Holoceno en el sudeste de la provincia de Buenos Aires (Argentina): el caso de Cueva Tixi. Cuaternario y Ciencias Ambientales, Publicación Especial, 4:31-36. Buenos Aires. [ Links ]
PARDIÑAS, U.F.J. 2000b. Tafonomía de microvertebrados en yacimientos arqueológicos de Patagonia (Argentina). Arqueología, 9:265-340. [ Links ]
PARDIÑAS, U.F.J. y C. GALLIARI. 1998.Sigmodontinos (Rodentia, Muridae) del Holoceno inferior de Bolivia. Revista Española de Paleontología, 13(1):17-25. [ Links ]
PARDIÑAS, U.F.J. y M. LEZCANO. 1995. Cricétidos (Mammalia, Rodentia) del Pleistoceno tardío del nordeste de la provincia de Buenos Aires (Argentina). Aspectos sistemáticos y paleoambientales. Ameghiniana, 32(3):249-265. [ Links ]
PARKER, G. y S. MARCOLINI. 1992. Geomorfología del delta del Paraná y su extensión al Río de la Plata. Revista de la Asociación Geológica Argentina, 47(2):243-249. [ Links ]
REDFORD, K.H y J.F. EISENBERG. 1992. Mammals of the Neotropics, Vol. 2. The Southern cone: Chile, Argentina, Paraguay and Uruguay.University of Chicago Press. Chicago, pp. 1-338. [ Links ]
REIG, O.A. 1977. A proposed unified nomenclature for the enamelled components of the molar teeth of the Cricetidae (Rodentia). Journal of Zoology, 181:227-241. [ Links ]
REIG, O.A. 1978. Roedores cricétidos del Plioceno superior de la Provincia de Buenos Aires (Argentina). Publicaciones del Museo Municipal de Ciencias Naturales de Mar del Plata "Lorenzo Scaglia", 2(8):164-190. [ Links ]
REIG, O.A. 1987. An assessment of the systematics and evolution of the Akodontini, with the description of new fossil species of Akodon (Cricetidae, Sigmodontinae). Pp. 347-399. En: Studies in Neotropical Mammalogy (Patterson, B.D. y R.M. Timm, eds.). Fieldiana, Zoology, New Series, 39. [ Links ]
REIG, O.A. 1994. New species of akodontine and scapteromyine rodents (Cricetidae) and new records of Bolomys (Akodontini) from the upper Pliocene and Middle Pleistocene of Buenos Aires Province, Argentina. Ameghiniana, 31(2):99-113. [ Links ]
RUSCONI, C. 1931. Dos nuevas especies de roedores del piso ensenadense de Buenos Aires. Notas Preliminares del Museo de La Plata, 1(3):333-338. [ Links ]
RUSCONI, C. 1936. Distribución de los vertebrados fósiles del Ensenadense. Boletín de la Academia Nacional de Ciencias de Córdoba, 33(3):183-214. [ Links ]
SALEMME, M. 1987. Paleoetnozoología del sector bonaerense de la región Pampeana. Tesis doctoral, Facultad de Ciencias Naturales y Museo, Universidad Nacional La Plata, pp. 1-283. [ Links ]
SMITH, M.F. y J.L. PATTON 1999. Phylogenetic relationships and the radiation of sigmodontine rodents in South America: evidence from cytochrome b. Journal of Mammalian Evolution, 6:89-128 [ Links ]
VOSS, R.S. y M.D. CARLETON. 1993. A new genus for Hesperomys molitor Winge and Holochilus magnus Hershkovitz (Mammalia, Muridae) with an analysis of its phylogenetic relationships. American Museum Novitates, 3085:1-39 [ Links ]
Recibido 28 agosto 2003.
Aceptación final 27 febrero 2004.

