










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. v.11 n.2 Mendoza jul./dic. 2004
Características Químicas de salados y hábitos alimenticios de la Danta de montaña (Tapirus pinchaque Roulin, 1829) en los Andes Centrales de Colombia
Diego J. Lizcano y Jaime Cavelier
Laboratorio de Ecología Vegetal, Departamento de Ciencias Biológicas, Universidad de los Andes, Carrera 1 No. 18A-70, Bogotá, Colombia.
ABSTRACT: Chemical characteristics of salt licks and feeding habits of mountain tapir (Tapirus pinchaque) in the central Andes of Colombia. The chemical composition of salt licks and the diet of the mountain tapir (Tapirus pinchaque) were studied in the upper montane rain forests (2900-3700 m), in the Parque Regional Natural Ucumari and Parque Nacional Natural Los Nevados, Colombia. We collected water samples from the salt licks, 28 feces and voucher specimens of plants with tapir browsing sign. The chemical analysis of the water from salt licks showed higher concentrations of Na, N-NH4 y N-NO3when compared to the water of a nearby stream used as control. The nitrogen of the water at the salt lick is probably being fixed by Nostoc, a Cynobacteria associated with Gunneramanicata and G. magnifolia, two plants located in the upper part of the rocky cliff from where the water drains to the salt lick. It is likely that the tapirs are drinking these waters to complement their diet, especially in an environment like tropical montane forests where nitrogen is in low concentration. A total of 35 plant species showed browsing signs. The plant families that the tapir consumed the most were ferns (seven species), Melastomataceae (six species) and Rubiaceae (five species). In the feces there were mostly leaves and twigs and no fruits or seeds. Germination experiments should be carried out to determine if the mountain tapir is a seed dispersal as suggested in Ecuador.
Key words: Colombia. Diet. Salt licks. Tapir. Tapirus pinchaque.
INTRODUCCIÓN
La danta de montaña (Tapirus pinchaque Roulin, 1829) es la más pequeña de las cuatro especies de tapires vivientes, con una longitud total de 180 cm, alzada de 75 a 90 cm y un peso de hasta 250 kg. Se distingue de los otros tapires por sus labios blancos y pelaje de color negro relativamente largo (3-4 cm). Su hábitat comprende los bosques montanos (2000-3500 m) y páramos (>3500 m) (Shauenberg, 1969) y habita en los Andes de Colombia (Acosta et al., 1996), Ecuador (Downer, 1996) y norte del Perú (Lizcano y Sissa, 2003). La danta de montaña es una especie muy sensible a la intervención, fragmentación y destrucción del hábitat (Downer, 1997). Sus poblaciones se encuentran aisladas y sometidas a una alta presión de cacería, por lo que esta especie está incluida en el apéndice I de CITES y considerada como en peligro de extinción (UICN, 1996).
Los estudios realizados sobre la dieta de Tapirus terrestris (Salas y Fuller, 1996), T. bairdii (Naranjo, 1995) y T. pinchaque (Acosta et al., 1996; Downer, 1996; Schauenberg, 1969), así como comparaciones entre las especies neotropicales, muestran que las dantas son animales herbívoros que consumen un gran número de especies (Olmos, 1997). Existe controversia sobre su papel como dispersores de semillas. Algunos autores (Salas y Fuller, 1996) los consideran malos dispersores, debido a su hábito de defecar en el agua, o por destruir las semillas que consume (Janzen, 1981). Sin embargo, otros estudios indican que el tapir es un buen dispersor de semillas para varias especies (Fragoso, 1997; Fragoso y Huffman, 2000).
Los estudios realizados con la danta de montaña sugieren que esta especie se alimenta de una gran variedad de hojas frescas, plántulas, ramas de arbustos y árboles pequeños de los bosques andinos (p. ej. Chusquea sp. y Miconia sp.) y de algunos frutos, pastos, plantas acuáticas y de pantano (Schauenberg, 1969). En la actualidad se cuenta con un listado de plantas consumidas por T. pinchaque en los bosques montanos y páramos del Ecuador (Downer, 1996), como así también en los bosques montanos de Colombia, entre los 2400 y 2900 m de altitud, en la parte baja de su rango altitudinal (Acosta et al., 1996).
Al igual que otros ungulados, como Odocoileus virginianus (Weeks y Kirkpatrick, 1976), y primates (Izawa, 1993), las dantas frecuentan lugares con altas concentraciones de nutrientes donde las especies consumen agua y suelo, conocidos como salados (Downer, 1996; Montenegro, 1998). En el Parque Regional Natural Ucumari existe evidencia del uso de los salados por las dantas (Acosta et al., 1996), pero se desconoce la frecuencia de uso, así como la composición del agua de éstos.
Los objetivos de este trabajo fueron describir la composición de la dieta de la danta de montaña en los bosques montanos altos entre los 2900 y 3700 m y la composición química de los salados en el Parque Regional Natural Ucumari y parte del Parque Nacional Natural Los Nevados, Colombia.
ÁREA DE ESTUDIO
El estudio se llevó a cabo en la vertiente occidental de la Cordillera Central colombiana, entre los 2900 y los 3700 m, en el Parque Regional Natural Ucumari y el Parque Nacional Natural Los Nevados, Departamento de Risaralda (Fig. 1a). En esta fracción del gradiente altitudinal, se presentan las selvas andinas, o bosques montanos altos sensu Grubb (1977), las cuales tienen un dosel más bajo y una diversidad menor de especies leñosas que las selvas subandinas o bosques montanos bajos en la parte más baja del gradiente (Gentry, 1992). A mediados de la década de 1950, algunas áreas de bosque maduro fueron taladas para la extracción de maderas finas y producción de carbón vegetal, siendo convertidas finalmente en pastizales para la ganadería de altura (Londoño, 1994; N. Monsalve, com. pers. 1996). En la actualidad, estas áreas están cubiertas por bosques secundarios dominados por Weinmannia pubescens, Miconia spp. y Tibouchina grossa , junto con parches reducidos del pasto exótico Penisetum clandestinum.

Fig. 1.Localización del área de estudio en los AndesCentrales de Colombia. a) El recuadro corresponde al área donde se realizó el trabajo de campo. b) Área entre los ríos Barbo y Otún. Las líneas punteadas representan las curvas de nivel y las líneas continuas corresponden a los senderos de danta en los cuales se recolectaron las plantas consumidas. S = El "salado"; L = "lamedero". En el recuadro de este mapa se indica el área estudiada por Acosta et al. (1996).
La precipitación media anual varía entre 2500 mm a una altitud de 2120 m (Estación El Cedral) y 980 mm a 4000 m (Laguna del Otún). Las lluvias presentan un patrón bimodal con períodos menos lluviosos entre los meses de diciembre y enero y julio y agosto. Sin una época seca claramente definida. La temperatura media anual en El Cedral es de 15 ºC y en la Laguna del Otún, 5.5 ºC(Witte, 1993).
En la zona se pueden encontrar, además de la danta de montaña, otros mamíferos como pumas (Pumaconcolor), osos andinos (Tremarctos ornatus), venados (Mazama spp.), guaguas (Agoutitaczanowskii) y coaties (Nasuellaolivacea). La población local de dantas de montaña, estimada mediante censos de huellas, es de entre 11 y 15 individuos (Lizcano y Cavelier, 2000). Senderos con rastros y heces de danta son frecuentemente encontrados dispersos entre el bosque del área de estudio; algunas veces las heces pueden encontrarse agrupadas en letrinas (Acosta et al., 1996).
MÉTODOS
El trabajo de campo se efectuó desde enero de 1995 hasta febrero de 1996. Se realizaron 5 salidas de campo: la primera en abril y mayo (33 días), la segunda en agosto y septiembre (18 días), la tercera en diciembre de 1995 (15 días), la cuarta en enero (21 días) y la quinta en febrero de 1996 (15 días), totalizando 102 días de trabajo de campo. Durante estas salidas se invirtieron en promedio 7 horas diarias en el campo y se recorrieron los senderos de dantas entre los ríos Barbo y Otún y entre las cotas 2900 y 3700 m (Fig. 1b). Estos senderos totalizaron 42 km, los cuales se recorrieron cuidadosamente a una velocidad de uno a dos kilómetros por hora, en compañía de un experimentado ex cazador local. Los senderos se encontraron principalmente a lo largo de las crestas de las montañas y en las márgenes de quebradas. Se reconocían por ser de aproximadamente un metro de ancho y por estar desprovistos de vegetación debido al pisoteo del animal. La red de senderos de danta se ubicó en fotografías aéreas (1:20000) mediante mapas topográficos y coordenadas obtenidas en puntos de control con un posicionador satelital (GPS Panasonic 5500).
Durante los recorridos se buscaron salados y plantas ramoneadas por la danta, tanto a lo largo del camino como en "comederos" (claros en el bosque con signos de ramoneo de danta). Se colectaron las especies vegetales que presentaban señales de desgarramiento, principalmente en el cogollo, y huellas de danta a su alrededor. Adicionalmente, se colectaron 28 muestras fecales frescas, con menos de 48 horas de deposición, caracterizadas por su olor penetrante, color verde oscuro y brillante; 12 muestras se colectaron en abril-mayo y 16 en diciembre. Las muestras consistieron en dos o tres heces seleccionadas al azar de cada conjunto y fueron conservadas en una solución de alcohol etílico 90%, formol 5% y ácido acético5 %. Se analizaron bajo lupa binocular, separando tallos, hojas, semillas y pulpa que fueron pesados en seco con una balanza de precisión de 0.1 g (Bodmer, 1990).
Se colectaron muestras del agua estancada del salado de la región de La Suiza (El "Salado"), del agua que escurre por el "lamedero" de la Quebrada Paraíso (Fig. 1b) y de una quebrada cercana que se uso como control (Quebrada de las Mulas). Cada muestra de agua (1000 cc) fue colectada en dos botellas plásticas de 500 cc, evitando recoger residuos sólidos en las muestras. De cada una se determino el pH, conductividad y las concentraciones de Ca, Mg, K, Na, SO4, Cl, NO3, CO3y HCO3 mediante los métodos descritos en American Public Health Association (1980).
RESULTADOS
Se encontró un total de 35 especies de plantas consumidas por la danta de montaña, pertenecientes a 19 géneros y 15 familias (Tabla 1). Los grupos con más especies consumidas fueron los helechos (7 especies), Melastomataceae (6 especies) y Rubiaceae (5 especies) (Fig. 2). Las dantas se alimentaron principalmente en cuatro lugares: "comederos", a lo largo de los senderos, ubicados en las áreas abiertas en el ecotono entre el páramo y el bosque montano alto y, esporádicamente, en el páramo. Los "comederos" fueron en su mayoría claros de bosque donde se encontraron plantas como Chusqueafendlerii y Ericaceas (e.g., Cavendishia nitida). A lo largo de los caminaderos en los filos, la danta consumió arbustos de los géneros Blechnum, Miconia, y Oreopanax y, en los de fondos de quebradas, se alimentó de Gunnera manicata, Plantago australis y Myrridendron pennellii. En las áreas abiertas del limite superior del bosque de niebla consumió hojas de Lachemilla sp. En los análisis de heces se encontró un 72% (por peso) de hojas y un 28% de tallos y no se encontraron semillas > 2 mm de diámetro ni pulpa de frutas. Todas las heces se encontraron en tierra seca.
Tabla 1
Listado de plantas consumidas por la danta de montaña (Tapirus pinchaque), en el Parque Regional Natural Ucumari y el Parque Nacional Natural los Nevados comparada con la información disponible en Acosta et al. (1996).


Fig. 2.Número de especies vegetales por taxón que forman parte de la dieta de la danta de montaña entre las cotas 2400 y 3700 m s.n.m.
El "lamedero" de la Quebrada Paraíso es un escarpe rocoso por donde escurre agua lentamente en forma permanentemente. El agua en este lugar es incolora y no presenta ningún olor. Este lugar está cubierto en la parte alta por plantas de Gunnera manicata (hoja de pantano) y en la parte baja por musgos y hepáticas (Marchantia sp.). El salado de la región de La Suiza es una gruta de 3 m de alto con un pequeño pozo de agua estancada de 10 m por 4 m sobre el que hay pequeñas estalactitas. El agua en este lugar presenta un color lechoso y un leve olor a óxido. Además de los salados naturales se encontró que las dantas también visitan los salados artificiales dispuestos para el ganado. No se encontraron indicios de geofagia. Los análisis químicos de las aguas que consume la danta en el "lamedero" y en el salado son similares en cuanto a las concentraciones de sulfatos, nitrógeno y zinc (Tabla 2) y tienen concentraciones de iones más altas que el agua de la quebrada que se usó como control. Las concentraciones de iones fueron más altas en el salado que en el "lamedero" (Tabla 2).
Tabla 2
Análisis químicos de las aguas de la Quebrada de las Mulas (3600 m) usada como control, el "lamedero" (3400 m) y El "salado" (2800 m).

DISCUSIÓN
En este estudio se encontró un total de 35 especies de plantas consumidas por la danta de montaña. Esta lista y la reportada por Acosta et al., (1996) en la parte baja del gradiente altitudinal sugieren que la danta de montaña es un herbívoro generalista, como se ha sugerido tanto para T. terrestris (Bodmer, 1990) como para T. bairdii (Naranjo, 1995). Las familias de plantas más consumidas (Helechos, Melastomataceae y Rubiaceae) son típicas del sotobosque y de estadios sucesionales de los bosques montanos altos. Dentro de la dieta de la danta de montaña se registran especies como Myrridendron pennellii (Apaiceae) y Chusquea fendleri (Poaceae) que según datos de ex-cazadores son el alimento preferido por estos animales (N. Monsalve, com. pers.). De las 35 especies vegetales que se encontraron con signos de ramoneo, solamente 7 habían sido reportadas previamente (Acosta et al., 1996; Tabla 1). Estas diferencias pueden atribuirse a que las colecciones en estos dos estudios se realizaron a diferentes altitudes y al estrecho rango altitudinal de muchas especies de los bosques montanos andinos (Gentry, 1992).
El análisis de las heces de T. pinchaque mostró que el total del material está compuesto de tallos y hojas. A pesar de que las colecciones de heces se realizaron tanto en la época lluviosa (abril y mayo) como en la época menos lluviosa del año (diciembre y enero), no se encontraron diferencias visibles en la composición de las heces. En estas muestras no se encontraron semillas, aunque no es posible descartar la presencia de aquellas de muy pequeño tamaño, como las de los géneros Miconia (Melastomataceae) y Palicourea (Rubiaceae), que se hacen visibles únicamente cuando germinan. Tampoco se observaron semillas germinando sobre las heces, quizás debido a que la mayoría de las heces se encontraron en lugares sombreados y relativamente fríos y no en claros propicios (Bazzas, 1979).
En contraste, en los análisis de heces de las especies de tierras bajas, T. terrestris y T. bairdii (Tabla 3) se encontraron algunas frutas, situación atribuible al menor tamaño y abundancia de frutos en los bosques montanos. T. terrestris es considerada como dispersora de semillas en los bosques de tierras bajas del Brasil (Fragoso, 1997; Fragoso y Huffman, 2000), Perú (Bodmer, 1990), Argentina y Brasil (Rodríguez et al., 1993) y T. bairdii, en Costa Rica (Janzen, 1982; Naranjo, 1995).
Tabla 3
Composición (%) de la dieta según análisis de heces de las tres especies de tapires neotropicales. El asterisco (*) indica la suma de hojas y tallos.
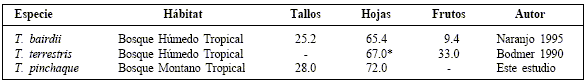
En este estudio se encontró que las dantas defecan regularmente en tierra. De igual forma, en algunos lugares el tapir de tierras bajas, T. terrestris, defeca con mas frecuencia en tierra firme que en el agua (Fragoso, 2000), mientras que en otros lugares ocurre a la inversa (Salas y Fuller, 1996). Por otro lado, T. bairdii defeca más frecuentemente en el agua que en tierra (Naranjo, 1995). Estas diferencias pueden deberse a diferencias innatas en el comportamiento de las especies (Naranjo, 1995), aunque seguramente existe una gran variabilidad de acuerdo a las características de cada región.
Durante este estudio se comprobó el uso de salados por la danta de montaña. El sodio, que esta presente en altas concentraciones en El "salado" (Tabla 2), ha sido identificado como uno de los cationes mas importantes en los salados usados por T. terrestris (Montenegro, 1998), loros y guacamayas en el Perú (Emmons y Stark, 1979; Munn, 1988), venados en Nepal (Moe, 1993) y es el ion preferido experimentalmente en mamíferos como Odocoileus virginianus, Sciurus niger y Marmota monax en Norteamérica (Weeks y Kirkpatrick, 1976; 1978). El nitrógeno encontrado en las aguas del "lamedero" de la Quebrada Paraíso está siendo fijado probablemente por Nostoc, una Cyanobacteria asociada con Gunnera spp. (Silvester y Smith, 1969; Meeks, 1998). Plantas de este género fueron observadas en la parte superior del "lamedero" y forman parte de la dieta de la danta de montaña tanto en Ucumarí y el Parque los Nevados (Tabla 1) como en los bosques montanos del Ecuador (Downer, 1996). El alto contenido de nitrógeno (NH4 y NO3) de las aguas estancadas del salado de la región de La Suiza puede provenir del proceso de nitrificación de la orina de los tapires, ya que se ha encontrado que éstos orinan frecuentemente en estos lugares (Montenegro, 1998). Este suplemento de nitrógeno puede ser de particular importancia en la dieta de la danta de montaña ya que se ha encontrado que tanto el nitrógeno como el fósforo son elementos que se encuentran en bajas concentraciones en estos bosques (Grubb, 1977; Tanner, 1977; Tanner et al., 1998). Es posible que la danta de montaña ingiera agua con alto contenido de sodio para complementar sus necesidades minerales y que el papel del nitrógeno sea "fertilizar" los microorganismos de su cámara fermentadora, ya que se ha encontrado que el nitrógeno del amonio puede ser incorporado a nucleótidos por los microorganismos del rumen (Huntington, 1986; Koening et al., 2000). Al parecer, el uso del agua de los salados tiene como función complementar las necesidades minerales de la dieta del animal (Hebert y Cowan, 1971; Emmons y Stark, 1979) o neutralizar las toxinas que contienen algunas de las plantas que consumen (Johns, 1986; Munn, 1988), en especial los helechos, que constituyen una parte destacable de la dieta de la danta de montaña (25%). Establecer el origen del nitrógeno en los salados es de particular importancia para futuros estudios.
La creciente población humana y la tendencia de transformación de los Andes en Colombia (Etter y Van Wyngaarden, 2000) sugieren que el hábitat de los tapires esta disminuyendo rápidamente. Se requieren estudios adicionales para determinar las plantas preferidas por esta especie, teniendo en cuenta la disponibilidad y el consumo y cómo éstas se relacionan con la abundancia de dantas. De esta forma podríamos enfocar nuestros esfuerzos de conservación a los hábitats más propicios para las dantas.
AGRADECIMIENTOS
Agradecemos a la Corporación Autónoma Regional del Risaralda (CARDER) y a Wildlife Conservation Society (WCS) por financiar este proyecto, así como a las Empresas Públicas de Pereira (hoy, Aguas y Aguas de Pereira) por el apoyo logístico durante la realización de este trabajo. Este agradecimiento se hace extensivo a todos los habitantes del páramo de Cortaderal por su valiosa colaboración y, en forma especial, a Javier Monsalve (guardabosque de las Empresas Públicas de Pereira) y al Sr. Noel Monsalve, por ser nuestros guías en el páramo. Finalmente, queremos expresar nuestro reconocimiento a Craig C. Downer por transferirnos sus conocimientos y experiencia de campo sobre tapires en Ucumari, a Eduardo Londoño y la gente de La Pastora, por la colaboración y el apoyo logístico, a William Vargas, por la determinación del material botánico y a Kirsten Silvius y 2 revisores anónimos por sus valiosos comentarios sobre versiones preliminares de este manuscrito.
LITERATURA CITADA
ACOSTA H, J CAVELIER y S LONDOÑO. 1996. Aportes al conocimiento de la biología de la danta de montaña, Tapirus pinchaque, en los Andes Centrales de Colombia. Biotropica 28:258-265. [ Links ]
AMERICAN PUBLIC HEALTH ASSOCIATION. 1980. Standard methods for examination of water and waste water. 15 edition, AMS Press, Washington. D.C. 465 pp. [ Links ]
BAZZAS FA. 1979. The physiological ecology of plant succession. Annual Review Ecology and Systematics 10:351-371. [ Links ]
BODMER RE. 1990. Fruit patch size and frugivory in the lowland tapir (Tapirus terrestris). Journal of Zoology 222:121-128. [ Links ]
DOWNER CC. 1996. The mountain tapir, endangered "flagship" species of the high Andes. Oryx 30:45-58. [ Links ]
DOWNER CC. 1997. Status and action plan of the mountain Tapir (Tapirus pinchaque). Pp. 10-22, en: Tapirs, status survey and conservation action plan (DM Brooks, RE Bodmer y S Matola, eds.). IUCN/SSC Tapir specialist group. IUCN, Gland, Switzerland and Cambridge, UK 164 pp. [ Links ]
EMMONS LH. y NM STARK. 1979. Elemental composition of a natural mineral lick in Amazonia. Biotropica 11:311-313. [ Links ]
ETTER A y W VAN WYNGAARDEN. 2000. Patterns of landscape transformation in Colombia, with emphasis in the Andean region. Ambio 29:432-439. [ Links ]
FRAGOSO JMV. 1997. Tapir-generated seed shadows: scale-dependent patchiness in the Amazon rain forest. Journal of Ecology 85:519-529. [ Links ]
FRAGOSO JMV y JM HUFFMAN. 2000. Seed-dispersal and seedling recruitment patterns by the last Neotropical megafaunal element in Amazonia, the tapir. Journal of Tropical Ecology 16:369-385. [ Links ]
GENTRY AH. 1992. Diversity and floristic composition on Andean forests of Peru and adjacent countries: Implications for their conservation. Memorias del Museo de Historia Natural U. N. M. S. M. (Lima) 21:11-29. [ Links ]
GRUBB PJ. 1977. Control of forest growth and distribution on wet tropical mountains: with special reference to mineral nutrition. Annual Review of Ecology and Systematics 8:83-107. [ Links ]
HEBERT D y IMCT COWAN. 1971. Natural salt licks as a part of the ecology of the mountain goat. Canadian Journal of Zoology 49:605-610. [ Links ]
HUNTINGTON GB. 1986. Uptake and transport of no protein nitrogen by the ruminant gut. Federation Proceedings 45:2272-2276. [ Links ]
IUCN. 1996. IUCN Red list of threatened animals. IUCN, Gland, Switzerland and Cambridge, UK 286 pp. [ Links ]
IZAWA K. 1993. Soil eating by Alouata and Ateles. International Journal of Primatology 14:229-242. [ Links ]
JANZEN DH. 1981. Digestive seed predation by a Costa Rican Baird´s tapir. Biotropica 13:59-63. [ Links ]
JANZEN DH. 1982. Seeds in tapir dung in Santa Rosa National Park, Costa Rica. Brenesia 19/20:129-135. [ Links ]
JOHNS T. 1986. Detoxification function of geophagy and domestication of the potato. Journal of Chemical Ecology 12:635-646. [ Links ]
KOENING KM, CJ NEWBOLD, FM MCINTOSH y LM RODE. 2000. Effects of protozoa on bacterial nitrogen recycling in the rumen. Journal of Animal Science 78:2431-2445. [ Links ]
LIZCANO DJ y J CAVELIER. 2000. Densidad Poblacional y Disponibilidad de Hábitat de la Danta de Montaña (Tapirus pinchaque) en los Andes Centrales de Colombia. Biotropica 32:165-173 [ Links ]
LIZCANO DJ y A SISSA. 2003. Notes on the Distribution, and Conservation Status of Mountain Tapir (Tapirus pinchaque) in Northern Peru. Tapir Conservation, newsletter of IUCN/SSC Tapir Specialist Group 12: 21-24. [ Links ]
LONDOÑO EM. 1994. Parque regional natural Ucumari, un vistazo histórico. Pp. 25-38, en: Ucumari, un caso típico de la biodiversidad biótica Andina (OJ Rangel, ed.). CARDER. Pereira, Colombia. 451 pp. [ Links ]
MEEKS JC. 1998. Symbiosis between nitrogen-fixing Cyanobacteria and plants. BioScience 48:266-276. [ Links ]
MONTENEGRO OL. 1998. The behavior of lowland tapir (Tapirus terrestris) at a natural mineral lick in the Peruvian Amazon. Master thesis, Wildlife ecology and conservation, University of Florida, Gainesville 78 pp. [ Links ]
MUNN CA. 1988. Macaw biology in Manu National Park, Peru. Parrot letter 1:18-21. [ Links ]
MOE SR. 1993. Mineral content and wildlife use of soil licks in southwestern Nepal. Canadian Journal of Zoology 71:933-936. [ Links ]
NARANJO EJ. 1995. Hábitos de alimentación del tapir (Tapirus bairdii) en un bosque tropical húmedo de Costa Rica. Vida Silvestre Neotropical 4:32-37. [ Links ]
OLMOS F. 1997. Tapirs as seed dispersers and predators. Pp. 3-9, en: Tapirs, status survey and conservation action plan (DM Brooks, RE Bodmer y S. Matola, eds.). IUCN/SSC Tapir Specialist Group. IUCN, Gland, Switzerland and Cambridge, UK. 164 pp. [ Links ]
RODRIGUEZ M, F OLMOS y M GALETTI. 1993. Seed dispersal by tapir in southeastern Brazil. Mammalia 57:460-461. [ Links ]
SALAS LA y TK FULLER. 1996. Diet of the lowland tapir (Tapirus terrestris L.) in the TabaroRiver valley, southern Venezuela. Canadian Journal of Zoology 74:1444-1451. [ Links ]
SCHAUENBERG P. 1969. Contribution à l'étude du Tapir pinchaque, Tapirus pinchaque Roulin 1829. Revue Suisse de zoologie 76:211-256. [ Links ]
SILVESTER WH y DR SMITH. 1969. Nitrogen fixation by Gunnera-Nostoc symbiosis. Nature 224:1231. [ Links ]
TANNER EVJ. 1977. Four montane rain forest of Jamaica: a quantitative characterization of the floristics, the soils and the foliar mineral levels, and a discussion of the interrelations. Journal of Ecology 65:883-918. [ Links ]
TANNER EVJ, PM VITOUSEK y E CUEVAS. 1998. Experimental investigation of nutrient limitations of forest growth on wet tropical mountains. Ecology 79:10-22. [ Links ]
WEEKS HP Jr y CM KIRKPATRICK. 1976. Adaptations of white-tailed deer to natural occurring sodium deficiencies. Journal of Wildlife Management 40:610-625. [ Links ]
WEEKS HP Jr y CM KIRKPATRICK. 1978. Salt preferences and sodium drive phenology in fox squirrels and woodchucks. Journal of Mammalogy 59:531-542. [ Links ]
WITTE HJ. 1993. Distribución estacional y altitudinal de la precipitación, la temperatura y la humedad en el transecto Parque los Nevados (Cordillera Central, Colombia). Instituto Geográfico Agustín Codazzi, Bogotá, Bogotá, Colombia 78 pp [ Links ]
Recibido 17 mayo 2004.
Aceptación final 2 agosto 2004.

