










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkMastozoología neotropical
versão impressa ISSN 0327-9383versão On-line ISSN 1666-0536
Mastozool. neotrop. v.11 n.2 Mendoza jul./dez. 2004
Micromamíferos (Didelphimorphia y Rodentia) del sudoeste de la provincia de Buenos Aires (Argentina): Taxonomía y distribución
Ulyses F. J.Pardiñas1, Agustín M. Abba2 y Mariano L. Merino2,3
1Centro Nacional Patagónico, Casilla de correo 128, 9120 Puerto Madryn, Chubut, Argentina. <ulyses@cenpat.edu.ar>. 2División Zoología Vertebrados, Facultad de Ciencias Naturales y Museo, Universidad Nacional La Plata, Paseo del Bosque s/n, 1900 La Plata, Argentina.3Comisión de Investigaciones Científicas de la Provincia de Buenos Aires.
ABSTRACT: Small mammals (Didelphimorphia and Rodentia) from southwestern Buenos Aires province, Argentina: Taxonomy and distribution. Species of small mammals from southwestern Buenos Aires province range within a wide variety of habitats, herbaceous (Pampean District), shrubs (Monte District), steppes, and xerophytic woodlands (Espinal District). Their natural history is still poorly known. In this paper we make a synthesis of the taxonomy and distribution of the marsupials (Marmosinae) and rodents (Sigmodontinae) of southwestern Buenos Aires province. Distributional records and taxonomical information were obtained from field survey, analyses of raptor pellets, revision of specimens housed in collections in Argentina, and literature. Between Sierra de la Ventana (38º S) and the río Negro (41ºS) occur two species of marmosines and 14 of sigmodontine rodents. Some of these species show a restricted distribution (e.g., Phyllotis xanthopygus, endemic for Sierra de la Ventana), and others are widespread (e.g. Akodon azarae or Calomys musculinus) across different phytogeographic units. A cluster analysis at local scale shows an ordination of small mammal assemblages with the topology (Sierra de la Ventana + Southern AtlanticCoast) (Espinal + Monte). The zoogeographic unicity of the Monte in the study area is evidenced by two sigmodontines, Akodon iniscatus and Oligoryzomys longicaudatus. The micromammal assemblage of Sierra de la Ventana may be characterized as pampasic. The wide distribution of A. azarae and C. musculinus may reflect the anthropic activities developed during the last century. These activities on shrub steppe have certainly diluted the original zoogeographic boundaries and allowed the dispersion of opportunistic sigmodontine species.
Key words. Antrophic impact. Argentina. Biogeography. Marmosinae. Sigmodontinae.
INTRODUCCIÓN
El conocimiento mastofaunístico de la provincia de Buenos Aires está asimétricamente distribuido tanto en el espacio como en el tiempo (Galliari et al., 1991). Una revisión de la información disponible indica que su desarrollo histórico puede ser dividido en dos grandes etapas:
a) Registros puntuales –con descripción de nuevas especies y subespecies– en localidades esparcidas en todo el territorio bonaerense, efectuados durante el siglo XIX y primeras décadas del XX;
b) Estudios de mayor detalle –que involucran análisis a nivel de población o comunidad, pero también descripción de nuevos taxones– en determinadas regiones, generalmente seleccionadas por su accesibilidad, importancia sanitaria o intereses particulares del investigador. Esta etapa se desarrolla entre 1960 y el presente y comprende, principalmente, Ciudad Autónoma de Buenos Aires y alrededores, Delta del Paraná (específicamente la región de Campana), partidos de Rojas y Pergamino (región endémica del virus Junín), agroecosistemas seleccionados del nordeste bonaerense, Mar del Plata, Balcarce y alrededores y laguna Chasicó (partido de Villarino).
Esta distribución asimétrica de los esfuerzos ha dejado importantes porciones del territorio (e. g., región al sudoeste de Bahía Blanca, área interserrana y sistemas serranos de Tandil y Ventana) prácticamente sin información básica. Paralelamente, el conocimiento de la distribución de algunas especies (e. g., Herpailurus yaguaroundi, Tolypeutes matacus, Zaedyus pichiy) se ha mantenido estático en los últimos 50 años (Galliari et al., 1991)
La importante modificación de la provincia por el impacto agropecuario en los últimos 150 años (Ghersa et al., 1998) ha repercutido profundamente en la estructura y composición de su biota. Asimismo, se han registrado cambios climáticos cíclicos de las temperaturas y precipitaciones medias (Hoffmann, 1988; Castañeda y Barros, 1994), que también deben haber afectado la distribución de ciertas especies estenoicas (e. g., Lyncodon patagonicus; Prevosti y Pardiñas, 2001). Sin embargo, la cuantificación e interpretación de los cambios recientes en los ensambles se ve limitada por la carencia de información mastofaunística previa a la etapa de mayor desarrollo antrópico en la región.
Particularmente, el sudoeste bonaerense ha recibido una escasa atención en cuanto a sus pequeños mamíferos. Pese a las tempranas contribuciones (Waterhouse, 1837, 1839; Thomas, 1919a) y a los esfuerzos posteriores de diferentes autores (e.g., Contreras, 1968, 1972, 1973a; Contreras y Maceiras, 1970; Massoia, 1973), la información que tenemos hoy es básicamente puntual y restringida mayoritariamente a su porción norte. Extensas regiones carecen de información básica, con el agravante de su rápida conversión en agroecosistemas o sectores de pastoreo por eliminación de la cubierta arbustiva.
Este trabajo tiene dos objetivos. En primer lugar, un aporte al conocimiento de los micromamíferos –marsupiales marmosinos y roedores sigmodontinos– del sudoeste de la provincia de Buenos Aires, mediante los resultados obtenidos de nuevos trampeos, la revisión de colecciones y el análisis de egagrópilas de aves rapaces. En segundo lugar, y sobre esta nueva base de información, un análisis zoogeográfico preliminar con relación a otras mastofaunas conocidas para latitudes medias de la Argentina.
MATERIALES Y MÉTODOS
Area de estudio
Corresponde a la porción de la provincia de Buenos Aires comprendida hacia el norte por los límites del partido de Saavedra y el arroyo Chasicó, la Sierra de la Ventana y el río Sauce Grande y hacia el sur y oeste, por el límite con las provincias de La Pampa y Río Negro (Fig. 1). Incluye los partidos de Bahía Blanca, Coronel de Marina L. Rosales, Monte Hermoso, Patagones, Saavedra, Tornquist y Villarino.
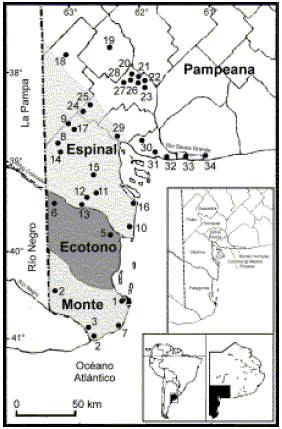
Fig. 1.Mapa del área de estudio, sudoeste de la provincia de Buenos Aires (Argentina), que incluye la ubicación geográfica de las localidades mencionadas en este trabajo sobreimpuestas al esquema fitogeográfico de Cabrera (1971). Para la referencia de los números, véase el Apéndice 1.
El clima para la región de Bahía Blanca y Sierra de la Ventana es subhúmedo seco, mesotérmico, con nulo o pequeño exceso de agua y concentración estival de la eficiencia térmica menor al 48%, que corresponde a un tipo transicional hacia los climas típicamente áridos. La temperatura media anual es de 15.7º C y la precipitación media anual ronda los 600 mm (Burgos y Vidal, 1951). Las áreas bonaerenses del Espinal y Monte poseen un clima semiárido con una temperatura media para el mes más cálido de 22.2 ºC y de 8.2 ºC para el mes más frío; la media anual de precipitaciones es de 466 mm (González Uriarte et al., 1987).
En el sudoeste bonaerense se destaca la Sierra de la Ventana, sistema serrano que genera ambientes heterogéneos –a nivel de relieve, flora y microclima– en el marco relativamente homogéneo de la pampasia (Cabrera, 1971; Kristensen, 1992). El área costera de la mencionada región está constituida por una barrera de médanos, con un ancho variable entre 4 y 10 km (Frenguelli, 1928).
Desde un punto de vista fitogeográfico, el área de estudio incluye porciones del Distrito Austral de la Provincia Fitogeográfica [PF] Pampeana, con una fisonomía de estepa o pseudoestepa de gramíneas, la PF del Espinal, representada por el Distrito del Caldén, y la PF del Monte, presente en la porción sur del partido de Patagones (Cabrera, 1971, 1976; Fig. 1). El sector correspondiente a la PF del Espinal se extiende hasta, aproximadamente, los 40º de latitud sur en el partido de Patagones (Lamberto, 1987). Es una variante empobrecida de esta unidad florística, con una fisonomía de arbustal y pastizales de gramíneas. Aparece aquí un estrato arbóreo compuesto por chañar (Geoffroea decorticans), piquillín (Condalia microphylla) y alpataco (Prosopis alpataco). La PF del Monte (Cabrera, 1976), que en el sudoeste bonaerense corresponde al Monte Oriental (sensu León et al., 1998), se caracteriza por la presencia de un estrato arbustivo de entre 1.5 m y 3 m de altura, integrado por jarilla (Larrea divaricata), piquillín y alpataco y un estrato herbáceo de escasa cobertura. El área comprendida entre los ríos Colorado y Negro constituye un extenso ecotono (Cabrera, 1971, 1976), aspecto que ha redundado en diversas interpretaciones en lo que se refiere al límite entre las PF del Monte y Espinal (Morello, 1958).
En la zona norte del área de estudio, la actividad productiva dominante es la explotación de tipo mixta agrícola-ganadera conducida en su variedad extensiva, con amplios cultivos de cereales. Las porciones bonaerenses de las PF del Espinal y Monte presentan dos grandes grupos según el uso de la tierra: áreas bajo riego –principalmente sobre el río Colorado– donde se cultivan hortalizas, frutales, cereales y forrajeras y áreas de secano, donde los cultivos mayoritarios son trigo, centeno, cebada forrajera y sorgo. En los sectores con regadío se encuentra muy desarrollada la cría de bovinos y ovinos; en las áreas de secano se practica la cría extensiva de ganado, principalmente bovino, ovino y caprino (González Uriarte et al., 1987).
Obtención de la información y taxonomía
La información utilizada en este trabajo surge de tres fuentes de evidencias:
a) Capturas de micromamíferos. Se llevaron a cabo viajes de colección, con el empleo de trampas de captura muerta (tipo Museum), captura viva (Sherman y jaulas) y cebos estándar, totalizando 1850 noches/trampa. En el Apéndice 1 se indican las localidades muestreadas;
b) Revisión de colecciones. Se obtuvo información correspondiente a los micromamíferos capturados en el área de estudio de las siguientes colecciones de Argentina: colección de mamíferos del Centro Nacional Patagónico, Puerto Madryn (CNP), colección Elio Massoia, Buenos Aires (CEM), Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Buenos Aires (MACN) y Museo de La Plata –División Zoología Vertebrados–, La Plata (MLP). En todos los casos, las determinaciones taxonómicas de catálogo o etiqueta fueron contrastadas con el examen directo del material;
c) Análisis de egagrópilas de aves rapaces. Se incluyen en este trabajo los ensambles de micromamíferos de 21 muestras de egagrópilas, mayoritariamente producidas por la lechuza de campanario, Tyto alba (Strigiformes: Tytonidae). De éstas, 9 fueron colectadas durante los trabajos de campo y las restantes corresponden a información publicada por diversos autores (Apéndice 2). Se tomaron también en cuenta los datos de un análisis de egagrópilas de laguna Guatraché, La Pampa (Massoia et al., 1997), ya que si bien es una localidad extra-bonaerense, su carácter limítrofe es informativo a los fines zoogeográficos. Es necesario aclarar que, pese a los sesgos que introducen lechuzas y búhos en la representación de especies de micromamíferos consumidas, los agregados generados mantienen, especialmente en el caso de T. alba, una estrecha relación con las ocurrencias reales a campo (Bellocq, 2000).
Las determinaciones taxonómicas se efectuaron mediante la comparación con colecciones de referencia y bibliografía. En el criterio sistemático se sigue a Galliari et al. (1996). Para la subdivisión ambiental del área de estudio y regionalización de las provincia de Buenos Aires se emplearon los esquemas y conceptos de Cabrera (1971), Lamberto (1987), González (1995, 1997) y León et al. (1998). Como base cartográfica se utilizaron las cartas topográficas del Instituto Geográfico Militar (Buenos Aires) a escalas 1:500.000 y 1:250.000. La ubicación geográfica de las localidades mencionadas en este trabajo se detalla en el Apéndice 1.
Se excluyen aquellos mamíferos exóticos silvestres (el lepórido Lepus europaeus y los múridos Musdomesticus y Rattus spp.) a los fines del tratamiento taxonómico ulterior (véase Resultados). Del mismo modo, tampoco se comentan las situaciones puntuales de los roedores caviomorfos Cavia aperea, Ctenomys spp., Galea musteloides y Microcavia australis. Si bien sus restos se registran en egagrópilas o algunos ejemplares han sido trampeados ocasionalmente, la información es escasa como para efectuar observaciones del mismo tenor que con respecto a los roedores sigmodontinos.
Análisis zoogeográfico
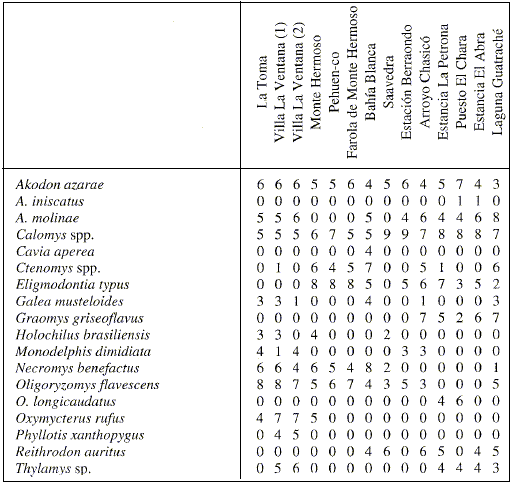
Se efectuaron análisis zoogeográficos a escala local y regional. El análisis local se desarrolló sobre una matriz básica de datos construida con la totalidad de los ensambles de egagrópilas con un número de individuos > 50 (14 sitios y 18 taxones). Esta matriz fue tratada tanto en su información nominal (presencia-ausencia) como cuantitativa (frecuencias estandarizadas por el método de octavas, Gauch, 1982; Tabla 1). Sobre ambas matrices se efectuó un análisis de agrupamiento mediante la aplicación de los coeficientes Jaccard (matriz nominal) y Euclidean distance (matriz cuantitativa estandarizada) y la técnica de ligamiento por la media aritmética no ponderada (UPGMA; Crisci y López Armengol, 1983).
Tabla 1
Matriz básica de datos (frecuencias estandarizadas) para el análisis zoogeográfico a escala local.
Para el análisis regional se volcó la información de las localidades en una grilla de 50 x 50 km. Las celdas correspondientes a otras áreas de la provincia de Buenos Aires fueron completadas con la información taxonómica y de distribución compilada por Pardiñas (1999). Con estos datos se construyó una matriz básica de datos de 11 unidades ambientales y 21 taxones, sólo a nivel nominal (Tabla 2). Las unidades ambientales consideradas fueron: Monte (MB), Espinal (ES), Sierra de la Ventana (VE), Litoral sur (LS), Litoral sudeste (SU), Tandilia (TA), Pampa deprimida (PD), Conurbano (CO), Pampa ondulada (PO), Selva marginal (SE) y Delta (DE). Esta matriz fue empleada en un análisis de agrupamiento siguiendo la misma metodología utilizada en el análisis de la matriz nominal a escala local. Además, sobre esta matriz se llevó a cabo un análisis de correspondencia (Ludwig y Reynolds, 1988), con la finalidad de explorar el comportamiento de las unidades y los taxones en forma conjunta.
Tabla 2
Matriz básica de datos (presencia-ausencia) para el análisis zoogeográfico a escala regional (para las referencias, véase el texto).

RESULTADOS
Taxonómicos y de distribución
Orden Didelphimorphia Gill, 1872
Familia Didelphidae Gray, 1821
Género Monodelphis Burnett, 1830
Monodelphisdimidiata (Wagner, 1875)
Especímenes examinados — MACN 14.954, MACN 14.961, MACN 15.718, MACN 15.719, MACN 18.658, MACN 18.659 (Abra de la Ventana); MLP 14.IX.99.65, MLP 14.IX.99.69, MLP 5.II.96.39 (Parque Provincial "Ernesto Tornquist").
Registros en egagrópilas — Arroyo de los Loros, cerro Destierro, cerro Dr. A. Serrano, La Toma, Villa La Ventana, Estación Berraondo, arroyo Chasicó y Estación Chasicó.
Registros adicionales — Abra de la Ventana (Crespo, 1964b).
Ambientes — Esta especie es dominante en cortaderales húmedos de baja y mediana altura (500-700 m s.n.m.) en Sierra de la Ventana, compartiendo este ambiente con Oxymycterus rufus. Sin embargo, también está presente en sectores completamente rocosos del mismo sistema serrano, sintópicamente con Phyllotis xanthopygus.
Comentarios — Desde un punto de vista taxonómico, aún queda pendiente clarificar el estatus de Monodelphis fosteri Thomas, 1923 (localidad tipo: Caleufu, 38º 52' S, 60º 12' O, La Pampa), si bien Reig (1964) planteó su carácter conespecífico respecto de M. dimidiata. Esta situación fue discutida por Pine et al. (1985) y, más recientemente, por Goin (2001), aunque sin una revisión del holotipo o de nuevos materiales topotípicos y, por consiguiente, de aportes a la cuestión. Cabe destacar que Pine et al. (1985) sugirieron mantener el status pleno de M. fosteri hasta tanto se documentara fehacientemente su probable sinonimia. M. dimidiata fue citada inicialmente (como Didelphysbrachyura) por Holmberg (1898) para el centro de la provincia, pero su notable abundancia es conocida tardíamente, a partir de los trabajos de Reig (1964, 1965) y Fornes y Massoia (1965). Coetáneamente, Crespo (1964b) la documentó por primera vez para el sudoeste bonaerense, a base del trampeo de un individuo en Sierra de la Ventana. Monodelphis presenta un peculiar patrón distribucional en la región pampeana –similar al del género Necromys, véase Galliari y Pardiñas (2000)– con poblaciones seguramente disyuntas en el sudeste y sudoeste. Su límite austral ha sido establecido en laguna Chasicó (Massoia, 1985; Massoia y Vetrano, 1988).
Género Thylamys Gray, 1821
Thylamys sp.
Especímenes examinados — MACN 16.263, MACN 18.656, MACN 18.657 (Sierra de la Ventana); MLP 14.IX.99.67 (Parque Provincial "Ernesto Tornquist"); MLP 21.V.74.1, MLP 24.V.74.5, MLP 27.X.95.10, MLP 27.X.95.11, MLP 27.X.95.12, MLP 30.X.95.14 (laguna Chasicó).
Registros en egagrópilas — Arroyo de los Loros, Villa La Ventana, Estancia La Petrona, Puesto El Chara, Estancia El Abra.
Registros adicionales —Sierra de la Ventana (Ringuelet, 1955; Crespo, 1964b), laguna Chasicó (Contreras, 1968, 1973a).
Ambientes —En el área de Sierra de la Ventana los especímenes trampeados fueron obtenidos en roquedales y pastizales a baja altura (500 m s.n.m.), dominados por Cardus sp. y Cortaderia sp.
Comentarios — No existe consenso sobre la taxonomía de las especies del género (Galliari et al., 1996); contribuciones recientes (e.g., Palma y Yates, 1998) sólo han llevado –en lo que hace a las formas patagónicas y centrales de Argentina– a una simplificación sin sustento explícito. Birney et al. (1996) destacan la presencia de dos morfotipos en el área patagónica, asimilados a las formas nominales T. pallidior (Matschie, 1916) (vinculada a la PF del Monte) y T. elegans (Waterhouse, 1839) (PF Patagónica). El primer registro de Thylamys en la provincia de Buenos Aires corresponde a la mención de Ringuelet (1955) para la Sierra de la Ventana (citada como Marmosa pallidior), posteriormente reafirmado por Crespo (1964b). Contreras (1968, 1973a) registró luego su presencia para el área de laguna Chasicó. En el sudoeste bonaerense se distribuye en forma más o menos continua desde Bahía Blanca hasta los límites provinciales con La Pampa y Río Negro y aun Chubut, provincias en las que no es infrecuente (Daciuk, 1974; Siegenthaler et al., 1993; Monjeau et al., 1997). Los ejemplares aquí estudiados corresponden a un único morfotipo de pequeño tamaño, congruente con lo destacado por Birney et al. (1996) para T. pallidior.
Orden Rodentia Bowdich, 1821
Familia Muridae Illiger, 1811
Tribu Akodontini Vorontzov, 1959
Género Akodon Meyen, 1833
Akodonazarae (Fischer, 1829)
Especímenes examinados — CEM 3504 (Estación Experimental del INTA Hilario Ascasubi); MACN 15.628, MACN 15.629, MACN 15.630, MACN 15.631, MACN 15.632, MACN 15.633, MACN 15.634, MACN 15.635, MACN 15.636, MACN 15.637 (laguna Chasicó); MLP 14.IX.99.4 (Faro Segunda Barranca); MLP 14.IX.99.12, MLP 14.IX.99.8 (Cabaña San José); MLP 14.IX.99.46, MLP 14.IX.99.28, MLP 14.IX.99.25, MLP 14.IX.99.21, MLP 14.IX.99.29, MLP 14.IX.99.27, MLP 14.IX.99.24, MLP 14.IX.99.23, MLP 14.IX.99.26, (Estancia La Elida); MLP 14.IX.99.63, MLP 14.IX.99.70, MLP 14.IX.99.66, MLP 14.IX.99.68, MLP 14.IX.99.72 (Parque Provincial "Ernesto Tornquist"); MLP 14.IX.99.48, MLP 14.IX.99.47, MLP 14.IX.99.54, MLP 14.IX.99.53, MLP 14.IX.99.58, MLP 14.IX.99.50, MLP 14.IX.99.49, MLP 14.IX.99.62, MLP 14.IX.99.56 (Balneario San Antonio); MLP 5.II.96.10, MLP 5.II.96.11, MLP 5.II.96.30, MLP 5.II.96.34, MLP 5.II.96.38, MLP 5.II.96.40, MLP 5.II.96.41, MLP 5.II.96.42, MLP 5.II.96.50, MLP 5.II.96.53, MLP 5.II.96.54, MLP 5.II.96.58, MLP 5.II.96.72, MLP 5.II.96.73, MLP 5.II.96.74, MLP 5.II.96.75 (Sierra de la Ventana); MLP 19.V.74.4, MLP 19.V.74.5, MLP 1.IX.70.8, MLP 1.XI.95.3, MLP 12.VI.70.33, MLP 12.VI.70.34, MLP 12.VI.70.35, MLP 12.VI.70.42, MLP 12.VI.70.43, MLP 12.VI.70.44, MLP 12.VI.70.46, MLP 12.VI.70.47, MLP 12.VI.70.48, MLP 12.VI.70.49, MLP 12.VI.70.50, MLP 18.VI.71.32, MLP 19.V.74.5, MLP 20.V.74.3, MLP 21.V.74.3, MLP 21.V.74.4, MLP 22.V.73.1, MLP 22.V.73.3, MLP 1.IX.70.8 (laguna Chasicó).
Registros en egagrópilas — Arroyo de los Loros, La Toma, Villa La Ventana, Monte Hermoso, Pehuén-co, Farola de Monte Hermoso, Bahía Blanca, Saavedra, Estación Berraondo, arroyo Chasicó, laguna Chasicó, Estancia La Petrona, Puesto El Chara, Estancia El Abra.
Registros adicionales —Laguna Chasicó (Contreras, 1968, 1972, 1973a, 1973b; Apfelbaum y Reig, 1989), Estación Experimental del INTA Hilario Ascasubi (Massoia, 1973).
Ambientes — Se han capturado ejemplares en diversos ambientes, incluyendo pastizales de Stipa spp. en Sierra de la Ventana, agroecosistemas, espartillares en las márgenes del río Negro, pastizales psamófilos y monte arbustivo.
Comentarios — Pese a ser uno de los sigmodontinos más abundantes de la región pampeana, A. azarae prácticamente no ha recibido atención en cuanto a la taxonomía de sus poblaciones (Reig, 1964), ni aun al complejo panorama generado por las sinonimias propuestas, muchas veces sin fundamentación explícita (Hershkovitz, 1990). Massoia (1990) indicó que las poblaciones del centro y sudoeste de Buenos Aires presentan mayores medidas corporales que las del norte y nordeste. Su límite austral fue originalmente establecido por Cabrera (1961) en la Sierra de la Ventana, posteriormente extendido por Contreras (1972, 1973a) hasta la región de laguna Chasicó. Los registros obtenidos por trampeos y análisis de egagrópilas indican que la especie alcanza la margen izquierda del río Negro, muy probablemente en simpatría con A. iniscatus. Si esta distribución parcialmente simpátrida es un efecto generado por el establecimiento de agroecosistemas al sur del río Colorado y su potencial como fuente de penetración de A. azarae, debe aún evaluarse en detalle.
Akodoniniscatus Thomas, 1919
Especímenes examinados — MLP 14.IX.99.61, MLP 14.IX.99.55 (Balneario San Antonio); MLP 5.X.99.4, MLP 5.X.99.5, CNP 779, CNP 780, CNP 781, CNP 782, CNP 783, CNP 784, CNP 785, CNP 786, CNP 787 (Bahía San Blas).
Registros en egagrópilas — Estación Experimental del INTA Hilario Ascasubi, Puesto El Chara, Estancia El Abra.
Registros adicionales —Bahía San Blas (Pardiñas y Galliari, 1999).
Ambientes — Los ejemplares trampeados fueron obtenidos en estepas arbustivas, parcialmente pastoreadas, dominadas por Atriplex sp. y Cyclolepis sp.
Comentarios — A esta especie se refieren las poblaciones de Akodon de pequeño tamaño desde el norte de Santa Cruz hasta el sudoeste de Buenos Aires, La Pampa y sur de Mendoza (e. g., Thomas, 1919a, 1919b, 1929; Tiranti, 1989; Barros et al., 1990). Una revisión integral de este taxón, incluyendo materiales topotípicos, es necesaria (véase la discusión en Pardiñas et al., 2003). Pardiñas y Galliari (1999) han documentado su presencia en Bahía San Blas y discutido en detalle las menciones previas para territorio bonaerense. Los registros de egagrópilas indican su ocurrencia hasta el río Colorado y sectores aledaños, como límite septentrional. Probablemente esta especie se extendiera, en un pasado reciente, más al norte, como lo sugiere su distribución colindante en la provincia de La Pampa (Estación Perú, 37º 38' S, 64º 09' O; Thomas, 1919a). Puede estimarse aquí un efecto simétricamente negativo al comentado para A. azarae: la eliminación del monte arbustivo habría producido la desaparición de las poblaciones de A. iniscatus, al menos hasta cercanías del río Colorado.
Akodonmolinae Contreras, 1968
Especímenes examinados — CEM 3503, CEM 3507, CEM 3508, CEM 3538, CEM 3547 (Estación Experimental del INTA Hilario Ascasubi); MACN 15.716, MACN 16.168, MACN 19.417, MACN 19.418, MACN 19.419, MLP 1.VIII.00.15 (Sierra de la Ventana); MACN 17.548 (Hilario Ascasubi); MACN 26.43 (Algarrobo); MACN 32.99 (Bahía San Blas); MLP 14.IX.99.13, MLP 14.IX.99.7, MLP 14.IX.99.5 (Cabaña San José); MLP 14.IX.99.16, MLP 14.IX.99.17 (Estancia El Abra); MLP 14.IX.99.3, MLP 14.IX.99.2, MLP 14.IX.99.1, MLP 14.IX.99.41 (Faro Segunda Barranca); MLP 14.IX.99.22, MLP 14.IX.99.42, MLP 14.IX.99.45, MLP 14.IX.99.43, MLP 14.IX.99.44, MLP 14.IX.99.40, MLP 14.IX.99.32, MLP 14.IX.99.37, MLP 14.IX.99.34, MLP 14.IX.99.33, MLP 14.IX.99.30, MLP 14.IX.99.39, MLP 14.IX.99.35, MLP 14.IX.99.36, MLP 14.IX.99.31 (Estancia La Elida); MLP 14.IX.99.51, MLP 14.IX.99.52 (Balneario San Antonio); MLP 30.VIII.99.8, MLP 18.V.74.4, MLP 1.IX.70.1, MLP 1.IX.70.2, MLP 1.IX.70.4, MLP 12.VI.70.55, MLP 12.VI.70.56, MLP 12.VI.70.57, MLP 12.VI.70.58, MLP 12.VI.70.59, MLP 21.V.74.2, MLP 3.X.70.38, MLP 3.X.70.39, MLP 3.X.70.40, MLP 3.X.70.41, MLP 3.X.70.42, MLP 3.X.70.43, MLP 3.X.70.44, MLP 3.X.70.45, MLP 3.X.70.46, MLP 3.X.70.47, MLP 18.V.74.5, MLP 18.V.74.3, MLP 18.V.74.8, MLP 10.VIII.76.1, MLP 19.V.74.2 (laguna Chasicó).
Registros en egagrópilas — Arroyo de los Loros, La Toma, Villa La Ventana, Bahía Blanca, Estación Berraondo, arroyo Chasicó, Estación La Petrona, Puesto El Chara, Estancia El Abra.
Registros adicionales — Sierra de la Ventana (Contreras, 1968), Laguna Chasicó (Contreras, 1968, 1973a; Apfelbaum y Reig, 1989), Estación Experimental del INTA Hilario Ascasubi (Massoia, 1973).
Ambientes — En Sierra de la Ventana, esta especie se registra en pastizales y roquedales desde los 500 m s.n.m. Hacia el sudoeste, es dominante en ambientes del monte arbustivo y subarbustivo.
Comentarios — A. molinae, originalmente descripto para el área de laguna Chasicó (localidad tipo: Vivero del Ministerio de Asuntos Agrarios, laguna Chasicó, partido de Villarino, provincia de Buenos Aires), presenta poblaciones abundantes en Sierra de la Ventana y en todo el sudoeste de la provincia de Buenos Aires, extendiéndose a La Pampa y Mendoza (Contreras, 1968, 1973a; Tiranti, 1988; Siegenthaler et al., 1990a y b, 1993). Su límite sudoriental puede establecerse en Península Valdés y alrededores de Puerto Madryn (42º 47' S, 65º 02' O), Chubut (Daciuk, 1974; Apfelbaum y Reig, 1989; Pardiñas et al., 2001). Se ha discutido ampliamente la posible sinonimia entre A. molinae y A. dolores Thomas, 1916 (localidad tipo: Yacanto, 32º 03' S, 65º 03' O, Córdoba), esta última tradicionalmente restringida a la provincia de Córdoba (e.g., Apfelbaum y Blanco, 1984; Bianchi y Merani, 1984; Wittouck et al., 1995). Adicionalmente, existe una continuidad geográfica y ambiental –aunque hoy parcialmente eliminada por la implantación de agroecosistemas en el este pampeano– entre sus geonemias, que sustentan su carácter conespecífico. Por otro lado, persiste una situación confusa en torno a la validez y extensión de A. neocenus Thomas, 1919 (localidad tipo: Neuquén, 38º 57' S, 68º 04' O). Para esta especie se ha referido un ejemplar con procedencia bonaerense (Myers, 1989; Pardiñas, 1993). A. molinae y A. neocenus presentan distribuciones superpuestas en el río Colorado (cf. Thomas, 1927; Contreras, 1968). En síntesis, la información disponible indica que estas especies del género Akodon (A. dolores, A. molinae y A. neocenus) conforman un grupo estrechamente relacionado, si no conespecífico, de amplia distribución en el monte y espinal del centro de Argentina (Pardiñas et al., 2003).
Género Necromys Ameghino, 1889
Necromysbenefactus (Thomas, 1919)
Especímenes examinados — MACN 15628 (laguna Chasicó); MLP 20.V.77.1, MLP 20.V.77.3, MLP 20.V.77.2, MLP 15.II.96.46, MLP 15.II.96.45, MLP 15.II.96.41, MLP 15.II.96.42, MLP 15.II.96.43, MLP 15.II.96.33, MLP 15.II.96.31, MLP 24.V.96.5, MLP 15.II.96.27, MLP 15.II.96.25, MLP 15.II.96.23, MLP 15.II.96.22, MLP 15.II.96.26, MLP 15.II.96.12, MLP 15.II.96.10, MLP 15.II.96.11, MLP 15.II.96.17, MLP 15.II.96.18, MLP 15.II.96.19, MLP 24.V.96.3, MLP 15.II.96.15, MLP 15.II.96.16, MLP 15.II.96.14, MLP 15.II.96.13, MLP 5.II.96.45, MLP 15.II.96.40, MLP 15.II.96.37, MLP 15.II.96.38, MLP 15.II.96.21, MLP 24.V.96.4, MLP 24.V.96.1, MLP 15.II.96.30, MLP 15.II.96.28, MLP 15.II.96.24, MLP 15.II.96.20, MLP 15.II.96.32, MLP 15.II.96.29, MLP 15.II.96.39, MLP 15.II.96.36, MLP 15.II.96.47, MLP 15.II.96.44, MLP 24.V.96.5, MLP 24.V.96.4, MLP 24.V.96.3, MLP 24.V.96.2, MLP 24.V.96.1 (Parque Provincial "Ernesto Tornquist").
Registros en egagrópilas — Arroyo de los Loros, cerro Destierro, cerro Dr. A. Serrano, La Toma, Villa La Ventana, Monte Hermoso, Pehuén-co, Farola de Monte Hermoso, Bahía Blanca, Saavedra.
Registros adicionales — 7 km al N Monte Hermoso (Apfelbaum y Reig, 1989), laguna Chasicó, Bahía Blanca, Monte Hermoso, Sierra del la Ventana, La Toma, Abra de la Ventana, Saavedra (Galliari y Pardiñas, 2000).
Ambientes — Frecuente en pastizales basales (500 a 800 m s.n.m.), vinculados a roquedales, en el área de Sierra de la Ventana.
Comentarios — La taxonomía de las especies bonaerenses de este género ha sido objeto de una reciente revisión (Galliari y Pardiñas, 2000). N.benefactus se considera una especie diferente de N. obscurus, esta última restringida al sudeste de la provincia. En el área de estudio, N.benefactus se registra con elevadas frecuencias en Sierra de la Ventana. Hacia el sudoeste, su número disminuye y su límite austral se encuentra en los alrededores de laguna Chasicó y Bahía Blanca. A este taxón deben referirse las menciones de Bolomysobscurus para las cercanías de Monte Hermoso (Apfelbaum y Reig, 1989). Cabe destacar que en las colecciones se han encontrado varios individuos referibles a esta especie, incluso con indicaciones de haber sido cariotipados, pero originalmente determinados como A. molinae. N.benefactus es la especie del género con poblaciones más australes.
Género Oxymycterus Waterhouse, 1837
Oxymycterusrufus (Fischer, 1814)
Especímenes examinados — MACN 14.939, MACN 14.940, MACN 14.941, MACN 14.944, MACN 14.945, MACN 14.946, MACN 14.948, MACN 14.949, MACN 14.950, MACN 14.957, MACN 14.958, MACN 14.959, MACN 14.970, MACN 14.973, MACN 14.974 (Sierra de la Ventana); MLP 14.IX.99.64, MLP 14.IX.99.71, MLP 5.II.96.4, MLP 5.II.96.6, MLP 5.II.96.22, MLP 5.II.96.17, MLP 5.II.96.20, MLP 5.II.96.1, MLP 5.II.96.57, MLP 5.II.96.71, MLP 5.II.96.52, MLP 5.II.96.23, MLP 5.II.96.51, MLP 5.II.96.76, MLP 5.II.96.56, MLP 5.II.96.60, MLP 5.II.96.70, MLP 5.II.96.65, MLP 5.II.96.59, MLP 5.II.96.63, MLP 5.II.96.62, MLP 5.II.96.68, MLP 5.II.96.66, MLP 5.II.96.35, MLP 5.II.96.32, MLP 5.II.96.37, MLP 5.II.96.67, MLP 5.II.96.29, MLP 5.II.96.28, MLP 5.II.96.25, MLP 5.II.96.27, MLP 5.II.96.31, MLP 5.II.96.36 (Parque Provincial "Ernesto Tornquist").
Registros en egagrópilas — Arroyo de los Loros, cerro Destierro, cerro Dr. A. Serrano, La Toma, Villa La Ventana, Monte Hermoso.
Registros adicionales —Abra de la Ventana (Crespo, 1964b).
Ambientes — Este sigmodontino es dominante en los pastizales y formaciones densas de Cortaderia de los pedemontes de Sierra de la Ventana, al menos hasta los 800 m s.n.m. También es frecuente en las márgenes de los arroyos de la vertiente sur de Sierra de la Ventana, incluyendo las porciones que atraviesan la cadena de médanos litorales.
Comentarios — Esta especie presenta poblaciones abundantes en los pastizales de Sierra de la Ventana, habiendo sido registrada –al igual que M. dimidiata– en fecha relativamente reciente para el centro y sur de Buenos Aires (Crespo, 1962, 1964b; Reig, 1964, 1965; Fornes y Massoia, 1965). Su frecuencia disminuye hacia el sudoeste y su límite austral se fija en cercanías de Bahía Blanca. Recientes análisis morfológicos y moleculares, que incluyen muestras del sudoeste bonaerense (Hoffmann et al., 2002; Gonçalves y de Oliveira, 2004), señalan una marcada consistencia de la especie en todo su rango de distribución.
Tribu Oryzomyini Vorontzov, 1959
Género Holochilus Brandt, 1835
Holochilusbrasiliensis (Desmarest, 1819)
Especímenes examinados — Ninguno.
Registros en egagrópilas — La Toma, Villa La Ventana, Monte Hermoso, Saavedra.
Registros adicionales — Bahía Blanca (Waterhouse, 1839), laguna Chasicó (Contreras, 1973a), Sauce Grande (Massoia, 1976).
Ambientes — En pajonales de cuerpos de agua lénticos y lóticos.
Comentarios — Los registros del sudoeste bonaerense pueden referirse a la subespecie H. b. darwini Thomas, 1897, cuyo límite austral estaría en la laguna Chasicó (Contreras, 1973a; Massoia, 1976). Esta especie es rara en el área de estudio, a juzgar por su escaso registro, tanto en trampeos como en egagrópilas de aves rapaces. Sin embargo, no puede desestimarse un sesgo atribuible a su gran tamaño corporal en relación con el de las trampas usualmente empleadas. Pese a las citas existentes (Hershkovitz, 1955), no hay registros documentados para La Pampa, si bien durante el Holoceno tardío alcanzó la cuenca del río Limay (Pardiñas, 1999), situación que sugiere variaciones recientes en su corología.
Género Oligoryzomys Bangs, 1900
Oligoryzomys flavescens (Waterhouse, 1837)
Especímenes examinados — MACN 14.468, MACN 14.475, MACN 14.476, MACN 14.477, MACN 19.224 (Sierra de la Ventana); MLP 5.II.96.49, MLP 5.II.96.69, MLP 5.II.96.55, MLP 5.II.96.33, MLP 5.II.96.26, MLP 5.II.96.43, MLP 5.VIII.98.11 (Parque Provincial "Ernesto Tornquist"), MLP 1.XII.76.3, MLP 1.IX.70.7 (laguna Chasicó).
Registros en egagrópilas — Arroyo de los Loros, cerro Dr. A. Serrano, La Toma, Villa La Ventana, Monte Hermoso, Pehuén-co, Farola de Monte Hermoso, Bahía Blanca, Saavedra, Estación Berraondo, arroyo Chasicó, Estación Chasicó.
Registros adicionales — Laguna Chasicó (Contreras, 1968, 1973a; Contreras y Alvarado, 1968).
Ambientes — Pastizales en relación con cuerpos de agua.
Comentarios — Este roedor, típico componente de los agroecosistemas bonaerenses, tiene su límite sudoriental en los alrededores de Bahía Blanca y laguna Chasicó (Contreras y Alvarado, 1968).
Oligoryzomys longicaudatus (Bennett, 1832)
Especímenes examinados — CEM 3512, CEM 3513, CEM 3520, CEM 3522 (Estación Experimental del INTA Hilario Ascasubi); CNP 788, CNP 789 (Bahía San Blas).
Registros en egagrópilas — Estación Experimental del INTA Hilario Ascasubi, Estancia La Petrona, Puesto El Chara.
Registros adicionales — Estación Experimental del INTA Hilario Ascasubi (Massoia, 1973).
Ambientes — Los ejemplares obtenidos en Bahía San Blas fueron trampeados en una estepa arbustiva de Atriplex lampa con estrato herbáceo de pasturas introducidas.
Comentarios — Massoia (1973) citó originalmente esta especie para Buenos Aires, considerándola una nueva subespecie, O. longicaudatus pampanus (localidad tipo: Estación Experimental del INTA Hilario Ascasubi). Se distingue de O. flavescens por su mayor tamaño, molares más robustos, bordes de los frontales formando suaves plataformas y escotadura lunar muy poco excavada (Massoia, 1973). Los registros de egagrópilas y trampeos permiten definir una geonemia relativamente extendida en el sudoeste bonaerense, en ningún caso simpátrica con O. flavescens. Para La Pampa, la especie fue mencionada originalmente por Tiranti (1988, 1989) y, recientemente, por De Santis et al. (2001). En estos casos, aparentemente ocurre en simpatría con O. flavescens. Recientes estudios moleculares sobre ejemplares de Bahía San Blas indican una escasa divergencia, pese a su grado de aislamiento, con respecto a poblaciones cordilleranas argentinas y chilenas de O. longicaudatus (Palma et al., en prensa).
Tribu Phyllotini Vorontzov, 1959
Género Calomys Waterhouse, 1837
Calomys spp.
Especímenes examinados — CEM 3509, CEM 3546 (Estación Experimental del INTA Hilario Ascasubi); MACN 32.234, MACN 32.235, MACN 32.236, MACN 32.237, MACN 32.238, MACN 32.239, MACN 32.240, MACN 26.49, MACN 26.50, MACN 26.51, MACN 26.52, MACN 26.53, MACN 26.54 (Algarrobo); MACN 15.715 (Sierra de la Ventana); MLP 14.IX.99.11, MLP 14.IX.99.15 (Cabaña San José); MLP 14.IX.99.38, MLP 14.IX.99.20 (Estancia La Elida); MLP 14.IX.99.59, MLP 14.IX.99.60, MLP 14.IX.99.57 (Balneario San Antonio); MLP 2.I.73.1 (laguna Chasicó).
Registros en egagrópilas — Arroyo de los Loros, cerro Dr. A. Serrano, La Toma, Villa La Ventana, Monte Hermoso, Pehuén-co, Farola de Monte Hermoso, Bahía Blanca, Saavedra, Estación Berraondo, arroyo Chasicó, Estación Chasicó, Estancia La Anita, Estación Experimental del INTA Hilario Ascasubi, Estancia La Petrona, Puesto El Chara, Estancia El Tigre, Estancia El Abra.
Registros adicionales — Laguna Blanca y Algarrobo (Massoia et al., 1968); Estación Experimental del INTA Hilario Ascasubi (Massoia, 1973); Monte Hermoso, Bahía Blanca (Waterhouse, 1839; Contreras y Justo, 1974).
Ambientes — Al igual que A. azarae, los representantes del género Calomys se registran en múltiples ambientes, desde agroecosistemas hasta monte arbustivo de Larrea divaricata y Prosopis sp.
Comentarios — C. laucha (Fischer, 1814) y C. musculinus (Thomas, 1913) son dudosamente diferenciables con restos cráneo-mandibulares fragmentarios como los obtenidos en egagrópilas y, por esta razón, se tratan aquí en forma conjunta (Pardiñas y Lezcano, 1995). C. musculinus presenta una extensa distribución que alcanza hacia el sur la provincia de Santa Cruz (Massoia y Pardiñas, 1994). Es la única especie hasta ahora citada en el ámbito patagónico (Pardiñas et al., 2003) y los estudios moleculares parecen confirmarlo (Salazar-Bravo et al., 2001). El límite sur en la distribución de C. laucha fijado en cercanías del río Colorado (Hershkovitz, 1962) debe ser estudiado en detalle. La dominancia de Calomys en varias regiones de Patagonia ha sido vinculada con el deterioro ambiental por efecto antrópico (Pardiñas, 1999; Pardiñas et al., 2000).
Género Eligmodontia Cuvier, 1837
Eligmodontia typus F. Cuvier, 1837
Especímenes examinados — MLP 11.II.36.47, MLP 11.II.36.25, MLP 11.II.36.47, MLP 11.II.36.25, MLP 11.II.36.25, MLP 22.V.58.2 (Monte Hermoso); MLP 8.XI.95.3, MLP 8.XI.95.4, MLP 8.XI.95.7, MLP 8.XI.95.6, MLP 8.XI.95.8, MLP XI.95.9, MLP 18.V.71.12, MLP 18.V.71.13, MLP 18.V.71.14, MLP 18.V.71.15, MLP 18.V.71.17, MLP 18.V.71.18, MLP 18.V.71.19, MLP 18.V.71.20, MLP 18.V.71.21, MLP 18.V.71.23, MLP 18.V.71.24, MLP 18.V.71.25, MLP 18.V.71.26, MLP 18.V.71.27, MLP 18.V.71.28, MLP 18.V.71.29, MLP 18.V.74.1, MLP 24.V.73.3, MLP 20.V.74.6, MLP 5.V.71.4, MLP 21.VI.96.1, MLP 21.VI.96.2, MLP 1.IX.70.14, MLP 1.IX.70.16, MLP 1.IX.70.21, MLP 28.IX.95.3, MLP 5.V.71.3, MLP 14.XII.72.1, MLP 12.VI.70.52, MLP 12.VI.70.53 (laguna Chasicó); MLP 28.IX.95.4 (Pehuén-co).
Registros en egagrópilas — Monte Hermoso, Pehuén-co, Farola de Monte Hermoso, Bahía Blanca, Estación Berraondo, arroyo Chasicó, Estación Chasicó, Estancia La Petrona, Puesto El Chara, Estancia El Abra.
Registros adicionales — Pehuén-co (Massoia y Fornes, 1964); laguna Chasicó (Contreras, 1968, 1972, 1973a), Salinas Chicas, Cañada de los Leones, Mayor Buratovich (Contreras, 1972), Bahía Blanca (Waterhouse, 1839).
Ambientes — Frecuente en pastizales de ambientes de médanos, en el sudoeste del litoral atlántico bonaerense y en estepas arbustivas con suelo desnudo.
Comentarios — Massoia y Fornes (1964) restringieron su localidad tipo al sudoeste de la provincia y descartaron las menciones sobre la presencia de este roedor en Corrientes y Delta del Paraná (Cabrera, 1961; Redford y Eisenberg, 1992; Musser y Carleton, 1993). Originalmente documentada para Bahía Blanca (terra typica de Mus elegans, sinonimizada con E. typus; Hershkovitz, 1962), Massoia y Fornes (1964) la citaron por primera vez para Pehuén-co, límite nordeste posteriormente ampliado por Contreras (1972). Los registros obtenidos en egagrópilas permiten ubicar este límite en los alrededores de Monte Hermoso. La penetración oriental de este roedor se realiza a través de la faja medanosa del litoral sur, lo cual explicaría su ausencia en Sierra de la Ventana. Cabe destacar que los hallazgos paleontológicos indican la presencia de Eligmodontia en el sudeste de la provincia, en cercanías de Mar del Plata, hace 300 años (Pardiñas, 1999).
Género Graomys Thomas, 1916
Graomys griseoflavus (Waterhouse, 1837)
Especímenes examinados — CEM 3521, CEM 3526, CEM 3527, CEM 3528, CEM 3529, CEM 3530, CEM 3531, CEM 3532, CEM 3533, CEM 3534, CEM 3535, CEM 3541, CEM 3542, CEM 3543, CEM 3544, CEM 3545 (Estación Experimental del INTA Hilario Ascasubi); MACN 26.33, MACN 26.34 (Algarrobo); MLP 14.IX.99.6, MLP 14.IX.99.11 (Cabaña San José); MLP 14.IX.99.18 (Estancia La Elida); MLP 18.V.71.34, MLP 2.I.79.1, MLP 2.I.79.19, MLP 2.I.79.2, MLP 12.I.79.15, MLP 24.V.73.13, MLP 22.V.73.5, MLP 14.XII.72.3, MLP 14.XII.72.6, MLP 14.XII.72.12, MLP 14.XII.72.20, MLP 2.I.73.3, MLP 7.III.73.1, MLP 14.VIII.73.3, MLP 22.V.73.7, MLP 14.XII.72.19, MLP 14.XII.72.16, MLP 14.XII.72.11, MLP 14.XII.72.10, MLP 14.XII.72.2, MLP 14.XII.72.18, MLP 14.VIII.73.1, MLP 22.V.73.8, MLP 22.V.73.4, MLP 2.I.73.2, MLP 24.V.73.1, MLP 31.X.80.5, MLP 24.V.73.7, MLP 2.I.79.22, MLP 1.X.70.21, MLP 30.X.79.16, MLP 2.I.79.25, MLP 30.X.79.15, MLP 2.I.79.11, MLP 2.I.79.5, MLP 2.I.79.16, MLP 2.I.79.10, MLP 2.I.79.24, MLP 2.I.79.13, MLP 2.I.79.14, MLP 2.I.79.17, MLP 30.X.79.17, MLP 18.V.71.36, MLP 30.X.95.6, MLP 30.X.95.7, MLP 9.VIII.71.10, MLP 30.X.95.4, MLP 9.VIII.71.9, MLP 31.X.73.7, MLP 31.X.73.9, MLP 31.X.73.3, MLP 31.X.73.5, MLP 31.X.73.1, MLP 7.VI.96.2, MLP 7.VI.96.4, MLP 7.VI.96.5, MLP 7.VI.96.6, MLP 7.VI.96.7, MLP 7.VI.6.9, MLP 24.V.73.4, MLP 29.IV.99.10, MLP 31.X.73.10, MLP 24.V.73.6, MLP 24.V.73.2, MLP 2.I.73.8, MLP 14.XII.72.9, MLP 14.XII.72.14, MLP 24.V.73.5, MLP 2.I.74.9, MLP 26.V.73.3, MLP 2.I.73.10, MLP 14.XII.72.8, MLP 14.XII.72.15, MLP 5.V.71.2, MLP 20.V.74.1, MLP 21.V.74.5, MLP 18.V.71.5, MLP 18.V.71.43, MLP 18.V.71.6, MLP 18.V.71.10, MLP 18.V.71.2, MLP 18.V.71.1, MLP 18.V.71.38, MLP 18.V.71.42, MLP 18.V.71.4, MLP 18.V.71.16, MLP 18.V.71.7, MLP 14.XII.72.4, MLP 14.XII.72.7, MLP 14.XII.72.5, MLP 2.I.79.26, MLP 2.I.79.3, MLP 2.I.79.9, MLP 31.X.73.4, MLP 22.V.73.6, MLP 20.V.74.6 (laguna Chasicó).
Registros en egagrópilas — Arroyo Chasicó, Estancia La Petrona, Puesto El Chara, Estancia El Abra.
Registros adicionales —Cercanías de Carmen de Patagones, boca del río Negro (Waterhouse, 1839), laguna Chasicó (Contreras, 1973a; Bianchini y Delupi, 1975); Algarrobo (Hershkovitz, 1962), Estación Experimental del INTA Hilario Ascasubi (Massoia, 1973), Bahía San Blas (Pardiñas, datos no publicados).
Ambientes — Frecuente en estepas arbustivas dominadas por Larrea sp. y Prosopis sp.
Comentarios — Tiranti (1998) distinguió dos citotipos asignando los binomios G. centralis (Thomas, 1902) (2n = 42) y G. griseoflavus (2n = 36-38), e indicó que este último es el registrado en Buenos Aires (Theiler y Gardenal, 1996). Cabe destacar la inexistencia de estudios citogenéticos sobre ejemplares topotípicos (i.e., aquellos provenientes de la desembocadura del río Negro). Graomys griseoflavus presenta poblaciones importantes en toda el área de estudio al sur de Bahía Blanca.
Género Phyllotis Waterhouse, 1837
Phyllotis xanthopygus (Waterhouse, 1837)
Especímenes examinados — MACN 14.914, MACN 14.915, MACN 14.916, MACN 14.917, MACN 14.918, MACN 14.919, MACN 14.920 (Abra de la Ventana); MLP 27.XII.01.11, MLP 27.XII.01.12, MLP 27.XII.01.13, MLP 27.XII.01.14 (Parque Provincial "Ernesto Tornquist").
Registros en egagrópilas — Cerro Destierro, cerro Dr. A. Serrano, Villa La Ventana.
Registros adicionales —Abra de la Ventana (Crespo, 1964a). La mención de Phyllotisdarwini efectuada por Contreras (1968) para laguna Chasicó, debe atribuirse a un lapsus en favor de G. griseoflavus (véase también Ringuelet, 1955).
Ambientes — Roquedales en Sierra de la Ventana desde los 500 m s.n.m.
Comentarios — P. bonariensis (localidad tipo: Abra de la Ventana; Crespo, 1964a), descripta como una subespecie de P. darwini (= P. xanthopygus), fue elevada a rango específico por diversos autores (Reig, 1978; Musser y Carleton, 1993; Galliari et al., 1996). Aquí se mantiene su status original ante la inexistencia de estudios comparados. Este roedor se presenta como un endemite de Sierra de la Ventana en territorio bonaerense. Los registros más cercanos para la especie corresponden a Pampa de Achala y Embalse de Río Tercero, Córdoba (Polop et al., 1985; Polop, 1989) y al Cerro Negro, La Pampa (Tiranti, 1989). Phyllotis acusa variaciones corológicas durante el Pleistoceno-Holoceno, como documentan los hallazgos de restos fósiles en cercanías de Bahía Blanca y Mar del Plata (Pardiñas y Deschamps, 1996; Pardiñas, 1999).
Género Reithrodon Waterhouse, 1837
Reithrodonauritus (Fischer, 1814)
Especímenes examinados — Ninguno.
Registros en egagrópilas — Bahía Blanca, Saavedra, arroyo Chasicó, Estación Chasicó, Estancia La Petrona, Estancia El Abra.
Registros adicionales — Epu Leufú (Contreras, 1972), laguna Chasicó (Contreras, 1973a).
Ambientes — Estepas graminosas y pasturas introducidas.
Comentarios — Sobre la base de los registros en egagrópilas, R. auritus presenta un particular patrón distribucional en el área de estudio (Pardiñas y Galliari, 2001)y está ausente en Sierra de la Ventana (al menos en la vertiente sudoriental) y en el litoral atlántico entre las localidades de Monte Hermoso y Pehuén-co. Cabe destacar que registros paleontológicos documentan su presencia en el litoral atlántico, en cercanías de Monte Hermoso, durante el Holoceno temprano (Pardiñas, 2001).
Cuantitativos y zoogeográficos
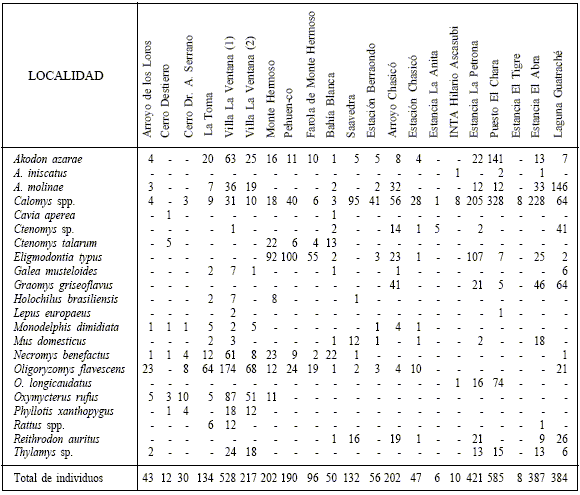
La Tabla 3 muestra las frecuencias de individuos en las localidades con análisis de egagrópilas del área de estudio. El sector de Sierra de la Ventana se caracteriza por elevadas frecuencias de Akodon azarae, Oligoryzomys flavescens y Oxymycterus rufus, acompañadas subsidiariamente por Akodon molinae y Necromys benefactus. Aquí se destacan las ausencias de Eligmodontia typus, Graomys griseoflavus y Reithrodon auritus. En la barrera sur de médanos litorales es dominante E. typus, acompañado por N. benefactus, O. flavescens y, en menor medida, Calomys spp. El Espinal en la provincia de Buenos Aires aparece dominado por Calomys spp.; una situación similar se observa en el Monte, pero participando también A. azarae.
Tabla 3
Número de individuos por especie en los análisis de egagrópilas (Apéndice 2) incluidos en este trabajo.
El análisis de agrupamientos de la matriz de presencia/ausencia de las localidades estudiadas (Fig. 2a) muestra una topología bien consistente a nivel geográfico. Se diferencian dos grandes grupos, uno que involucra las localidades de Sierra de la Ventana, más aquellas de la barrera sur de médanos litorales (a las que se agrega Saavedra), y otro que contiene a las restantes muestras. En el primer grupo, las localidades de Ventana forman un núcleo de alta similitud; lo propio hacen aquellas del litoral sur, a las que se une Saavedra a un bajo nivel de similitud. El segundo grupo muestra dos subconjuntos, uno conformado por localidades del Espinal empobrecido y otro por aquellas del Monte Oriental, quedando la localidad de Estación Berraondo como la más disímil de las estudiadas. En síntesis, el análisis de agrupamiento muestra la topología ((Ventana + Litoral Sur) (Espinal + Monte)), evidenciando el gradiente ambiental nordeste-sudoeste que rige la región pampeana y una congruencia entre los ensambles de micromamíferos y las unidades fitogeográficas. El análisis de agrupamientos sobre la base de las frecuencias, muestra resultados parcialmente distintos (Fig. 2b). Se observan dos grupos principales, uno conformado por las localidades del sistema de Ventana y otro, de topología compleja, donde pueden reconocerse con estructura geográfica, las localidades del Litoral sur, el Espinal y el Monte.
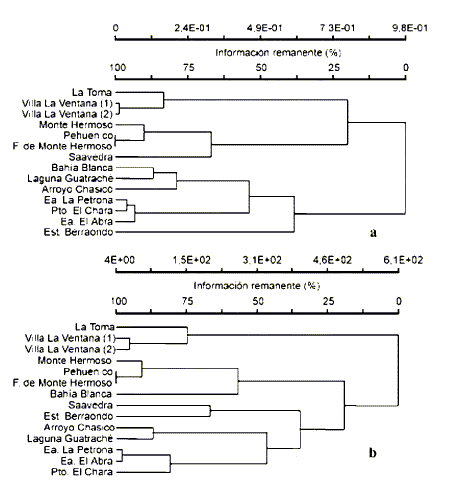
Fig. 2.Análisis zoogeográfico a escala local: relaciones fenéticas entre las muestras estudiadas del sudoeste de la provincia de Buenos Aires (Argentina): (a) sobre la matriz de presencia-ausencia, (b) sobre la matriz de frecuencias estandarizadas. CCC = 0,797.
A una escala regional, el análisis de agrupamientos por presencia-ausencia (Fig. 3a) destaca a la porción bonaerense del Monte como la unidad más disímil en el conjunto de las analizadas. Esta distinción es consistente con la presencia en la misma de dos especies, A. iniscatus y O. longicaudatus. Los restantes sectores conforman un amplio grupo con dos subconjuntos mayores, que separan al Delta y Selva marginal por un lado y a las restantes unidades por el otro. De estas últimas, Sierra de la Ventana aparece como la más diferenciada, mientras que el Litoral sur y el Espinal se unen en un núcleo de alta similitud, vinculado a las restantes áreas pampásicas. Estos resultados son congruentes con los obtenidos mediante el análisis de correspondencia (Fig. 3b), donde con evidente efecto arco se disponen las unidades ambientales y especies de micromamíferos en un gradiente nordeste-sudoeste que copia la tendencia de las precipitaciones y temperaturas medias.
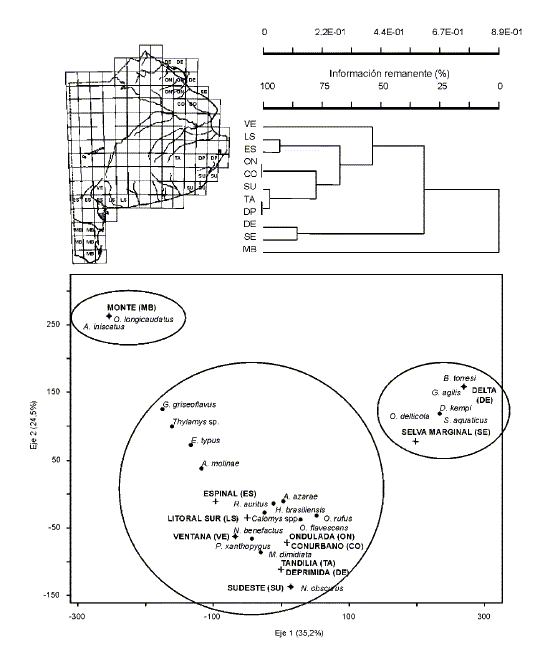
Fig. 3.Análisis zoogeográfico a escala regional: (a) relaciones fenéticas entre unidades ambientales de la provincia de Buenos Aires (matriz de presencia-ausencia), (b) análisis de correspondencia incluyendo unidades ambientales y especies de micromamíferos registradas (ejes 1 y 2).
DISCUSION Y CONCLUSIONES
Aspectos taxonómicos y zoogeográficos
Los resultados obtenidos permiten brindar un panorama actualizado sobre la distribución de varias especies de pequeños mamíferos en el sudoeste bonaerense. Entre los representantes del género Akodon, es novedosa la extensión de A. azarae hasta, al menos, la margen norte del río Negro; asimismo, su simpatría (no necesariamente sintopía) con A. iniscatus. Esto último era esperable, teniendo en cuenta los registros de principios de siglo para la localidad pampeana de Perú (Thomas, 1919a). Cabe destacar que es necesaria una revisión integral, incluyendo materiales topotípicos, de las poblaciones asignadas a este taxón en todo su rango de distribución (Pardiñas et al., 2003).
Una contribución significativa es la nueva geonemia de Oligoryzomys longicaudatus. Restringida a la localidad tipo de O. l. pampanus, los trampeos y análisis de egagrópilas indican su presencia en todo el sudoeste bonaerense entre los ríos Colorado y Negro. Los registros puntuales para las regiones patagónicas y pampeanas muestran la existencia de poblaciones aparentemente aisladas (García-Esponda et al., 1998; De Santis et al., 2001; Pardiñas et al., 2003). Habida cuenta del riesgo sanitario que esta especie implica (Mills y Childs, 1998), es necesario no sólo un adecuado estudio de su estatus taxonómico (Palma et al., en prensa), sino también una exploración integral de la misma en la Patagonia extra-andina.
Una síntesis de los patrones globales de distribución se brinda en la Tabla 4. Sobre esta base se pueden efectuar los siguientes comentarios, siempre para el área de estudio:
1.Únicamente Akodonazarae y Calomysmusculinus presentan una distribución extendida que involucra a todas las unidades fitogeográficas que ocurren en el sudoeste bonaerense;
2.Akodoniniscatus y Oligoryzomyslongicaudatus presentan un patrón distribucional común, ambas restringidas a la faja entre el río Colorado y sus inmediaciones y el río Negro;
3.Calomys laucha, Holochilus brasiliensis, Monodelphisdimidiata, Necromys benefactus y Oligoryzomys flavescens son especies que tienen como límite sudoriental de sus distribuciones el eje Chasicó-Bahía Blanca. Todas ocupan Sierra de la Ventana;
4.Akodon molinae y Thylamys sp. presentan un patrón distribucional comúne ingresan en Sierra de la Ventana pero están ausentes en la faja medanosa entre Bahía Blanca y el río Sauce Grande y regiones colindantes;
5.Eligmodontia typus y Graomys griseoflavus también presentan un patrón común, ambas ausentes en Sierra de la Ventana, pero con la particularidad de que E. typus ocupa ampliamente la franja medanosa costera entre Bahía Blanca y el río Sauce Grande;
6.Reithrodon auritus presenta una geonemia particular, ausente en Sierra de la Ventana y en la franja medanosa costera entre Bahía Blanca-río Sauce Grande;
7.Oxymycterus rufus y Phyllotis xanthopygus son aquellas especies de distribución más restringida, limitándose el primero a la franja medanosa costera y Sierra de la Ventana y el segundo exclusivamente a esta última.
Tabla 4
Patrones de distribución de los sigmodontinos y marmosinos registrados en el sudoeste de la provincia de Buenos Aires, Argentina. Los sectores geográficos se han ordenado de izquierda a derecha por latitud creciente.

La mayor parte de los pocos intentos zoogeográficos en cuanto a los mamíferos argentinos han estado regidos por las divisiones fitogeográficas de base (pero véase Real et al., 2003 para una aproximación diferente). De hecho, como tempranamente señalara Ringuelet (1961), el ajuste entre unidades zoo y fitogeográficas es algo esperable y hasta deseable (Pardiñas et al., 2003). Justamente, en el sudoeste de Buenos Aires se observa el engranaje de tres dominios zoogeográficos que, de norte a sur, son el Pampásico, el Central o Subandino y el Patagónico (Ringuelet, 1955, 1961). Estas unidades calcan aproximadamente el esquema florístico, en el mismo orden, PF Pampeana, PF del Espinal y PF del Monte (Cabrera, 1971). Sin embargo, es importante destacar que mientras los dominios Central y Patagónico pertenecen a la Subregión Andino-Patagónica, el dominio Pampásico corresponde a la Subregión Guayano-Brasílica (Ringuelet, 1961).
El análisis zoogeográfico regional, desarrollado en este trabajo sobre la base de roedores sigmodontinos y pequeños marsupiales, reconoce una división de mayor jerarquía entre el Monte y el resto de las unidades consideradas. El ecotono para estos dos bloques faunísticos se da en un sector poco conocido y muestreado, entre el eje Laguna Chasicó-Bahía Blanca y el río Colorado. Probablemente este ecotono ha fluctuado, desplazando su posición hacia el sudoeste debido al avance de la frontera agropecuaria (véase más abajo). En este esquema, la unicidad de Sierra de la Ventana como entidad zoogeográfica es discutible. Si bien presenta un taxón exclusivo, P. xanthopygus, están ausentes claros elementos característicos de los dominios Central y Patagónico (Eligmodontia typus, Graomys griseoflavus). Su ensamble de micromamíferos puede definirse como pampásico con algunos elementos incurrentes propios de hábitats más xéricos (e.g., Necromysbenefactus, Thylamys sp.). Lo mismo puede decirse de la franja medanosa litoral entre Bahía Blanca y el río Sauce Grande, penetrada por Eligmodontia typus. Esta franja estaría actuando de vía de ingreso para algunas especies que, aprovechando las características edáficas y microclimáticas, alcanzan posiciones orientales en Buenos Aires (Contreras y Reig, 1965).
En síntesis, los resultados obtenidos contrastan parcialmente con el esquema zoogeográfico de Ringuelet (1955, 1961). Los ensambles de pequeños mamíferos permiten reconocer una división de mayor jerarquía y determinan dos grandes bloques faunísticos en el ámbito bonaerense. Por otro lado, vinculan la Sierra de la Ventana con el Dominio Pampásico. Sin embargo, existe en ambas propuestas una fuerte coincidencia en señalar al área del río Colorado y aledaños como una "bisagra" zoogeográfica. Asimismo, el análisis de correspondencia señala que, dentro de las "unidades pampásicas", aquellas correspondientes al nordeste de la provincia (Delta-Selva marginal) se diferencian por un conjunto exclusivo de micromamíferos (e.g., Bibimys torresi, Oligoryzomys delticola); el reconocimiento de este grupo con la penetración del Dominio Brasílico de Ringuelet (1961) es clara. El análisis zoogeográfico pormenorizado a escala regional escapa a los fines de este trabajo. Aun así es justo reconocer que los resultados sobre la base de pequeños mamíferos marcan una incongruencia con el límite austral sugerido para las provincias Monte y Pampa (ambas pertenecientes a la Subregión Chaqueña) del esquema biogeográfico propuesto por Morrone (2001: figura 1).
Micromamíferos y cambios
en el uso de la tierra
El área de estudio reconoce un prolongado proceso de modificación antrópica. El ejemplo del partido de Patagones es elocuente. La reducción de la cobertura arbustiva/arbórea alcanzó al 49% de la superficie durante el período 1975-1999 (Radar, 2000), con una pérdida de 357.000 hectáreas, de las 911.171 que poseía el mencionado partido. La eliminación de esta cobertura vegetal natural redundó, en el mejor de los casos, en una sustitución por cultivos de trigo, alfalfa o girasol o en una explotación ganadera más intensa. Estos cambios a nivel macroambiental tienen que haber repercutido en la composición de los ensambles de micromamíferos, independientemente de nuestra posibilidad de percibirlos. La eliminación de la cubierta arbórea/arbustiva ha sido considerada como una de las causas principales de la desaparición de los componentes de la fauna (Roig, 1989).
Pardiñas (1999) y Pardiñas et al. (2000) plantearon, como hipótesis de trabajo, que la distribución y abundancia de ciertas especies de roedores sigmodontinos muestran una estrecha relación con las modificaciones antrópicas. El análisis de diversas secuencias de micromamíferos fósiles en las regiones pampeana y patagónica revela cambios dramáticos en algunas especies de micromamíferos, traducidos tanto en extinciones locales como en variaciones de frecuencias. Así, para el sudeste bonaerense, se verifica un incremento continuo de Calomys spp. durante los últimos 300 años. Situaciones similares se detectan en varias regiones de Patagonia, sometidas durante el último siglo a explotación agrícola y ganadera (Pardiñas, 1999; Pardiñas et al., 2000). Procesos similares, con pauperización de ensambles de micromamíferos por impacto antrópico, han sido reconocidos en diversos sectores del Hemisferio Sur (e.g., Avery, 1988, 1997; Simonetti 1994; Simonetti y Saavedra, 1997).
Calomys musculinus y Akodon azarae se distribuyen ampliamente en el área de estudio. Esta distribución puede ser un reflejo de un avance de estas especies oportunistas, gracias a las modificaciones antrópicas. Bilenca et al. (1994) han documentado para A. azarae una asociación negativa en cuanto a la duración del período reproductivo y del tamaño de camada con respecto al aumento latitudinal. Estos autores sugieren que el límite sur en la distribución de la especie estaría dado por restricciones ambientales con relación a la cantidad de alimento y a las condiciones ambientales óptimas para la reproducción. La progresiva transformación agrícola del sudoeste de la provincia de Buenos Aires, favorecida por un ciclo de mayores precipitaciones medias durante los últimos 30 años (Castañeda y Barros, 1994), habría permitido la paulatina expansión de estos sigmodontinos oportunistas al punto de convertirse en formas dominantes (e.g., Puesto El Chara). Por otra parte, la eliminación de hábitats naturales, especialmente aquellos arbustivos de la franja Laguna Chasicó y el Río Colorado, habría contribuido a retraer las poblaciones de ciertos micromamíferos, como Akodon iniscatus y Oligoryzomys longicaudatus. Estas modificaciones provocarían una dilución del límite entre los grandes bloques faunísticos previamente reseñados, generando el corrimiento de esta frontera zoogeográfica hacia el sur.
APÉNDICE 1
Ubicación geográfica de las localidades mencionadas en el texto, correspondientes al sudoeste de la provincia de Buenos Aires, Argentina (véase la Fig. 1). El símbolo t señala aquellas donde fueron efectuados trampeos de micromamíferos (este trabajo).
Partido de Patagones:
1.Bahía San Blas t (40º 33' S, 62º 13´ O)
2.Boca del Río Negro (41º 2' S, 62º 48´ O)
3.Cabaña San José t(40º 57' 51" S, 62º 47' 58" O)
4.Estancia El Abra t(40º 30' 09" S, 63º 22' 46"O)
5.Estancia El Tigre t(39º 50' S, 62º 23' O)
6.Estancia La Elida t(39º 21' 32" S, 63º 19' 21" O)
7.Faro Segunda Barranca t(40º 53' 50" S, 62º 40' 45" O)
Partido de Villarino:
8.Algarrobo (38º 54' S, 63º 8' O)
9.Arroyo y laguna Chasicó (38º 38' S, 63º 06'O)
10.Balneario San Antonio t(39º 39' 06" S, 62º 7' 09" O)
11.Estación Experimental del INTA Hilario Ascasubi (39º 22' S, 62º39' O)
12.Estancia La Anita (39º 16' S, 62º 37' O)
13.Estancia La Petrona (39º 26' S, 62º 46' O)
14.Laguna Blanca (38º 50' S, 63º 10' O)
15.Mayor Buratovich (39º 16' S, 62º 37' O)
16.Puesto El Chara (39º 27' S, 62º 03' O)
17.Salinas Chicas (38º 47' S, 63º 00' O)
Partido de Puán:
18.Epu Leufú (no localizada)
Partido de Saavedra:
19.Saavedra (37º 46' S, 62º 21' O)
Partido de Tornquist:
20.Abra de la Ventana (38º 04' S, 62º 02' O)
21.Arroyo de los Loros t(38º 03' S, 61º 55' O)
22.Parque Provincial "Ernesto Tornquist" - Cerro Destierro t(38º 01' 25" S, 62º 01' 04" O)
23.Cerro Dr. A. Serrano (38º 04' S, 61º 57' O)
24.Estación Berraondo (38º 25' S, 62º 50' O)
25.Estación Chasicó (38º 20' S, 62º 38' O)
26.La Toma t(38º 06' S, 61º 58' O)
27.Parque Provincial "Ernesto Tornquist" - Refugio t(38º 02' 23" S, 61º 59' 26"O)
28.Villa La Ventana (38º 08' S, 61º 47' O)
Partido de Bahía Blanca:
29.Bahía Blanca (38º 44' S, 62º 16' O)
Partido de Coronel de Marina L. Rosales:
30.Cañada de los Leones (39º 04' S, 61º 30' O)
31.Pehuén-co (39º 00' S, 61º 33' O)
32.Farola de Monte Hermoso (39º 00'S, 61º 56' O)
Partido de Monte Hermoso:
33.Monte Hermoso (38º 59' S, 61º 18' O)
34.Sauce Grande (38º 23' S, 61º 48' O)
APÉNDICE 2
Muestras de egagrópilas empleadas en este trabajo. Se indica localidad, especie de ave que generó la muestra y referencias bibliográficas cuando corresponde.
Arroyo de los Loros, Tyto alba, Pardiñas (1999); cerro Destierro, T. alba, Pardiñas (1999); Villa La Ventana (1), T. alba, Massoia et al. (1993); Villa La Ventana (2), T. alba, Pardiñas (1999); cerro Dr. A. Serrano, T. alba, Pardiñas (1999); La Toma, T. alba, Pardiñas (1999); Monte Hermoso, T. alba, Pardiñas (1999); Farola de Monte Hermoso, ave indeterminada, Pardiñas (1999); Bahía Blanca, T. alba, Pardiñas (1999); arroyo Chasicó, Asio flammeus, Massoia (1985); Pehuén-co, T. alba; Estación Chasicó, T. alba; Estación Berraondo, T. alba; Puesto El Chara, T. alba; Estancia El Abra, T. alba; Estancia El Tigre, T. alba; Estancia La Anita, Athene cunicularia; Estación Experimental INTA Hilario Ascasubi, T. alba; Estancia La Petrona, T. alba; Saavedra, T. alba, Pardiñas (1999); Laguna Guatraché, T. alba, Massoia et al. (1997).
AGRADECIMIENTOS
D. Birochio, A. Carlini, F. Cremonte, V. Di Martino, E. Etcheverry, C. Galliari, F. Goin, R. González Ittig, F. Moschione y A. Scorolli efectuaron o participaron en diversos trabajos de campo (recolección de egagrópilas, trampeos, etc.) y permitieron gentilmente el uso de la información obtenida. F. Goin corroboró las determinaciones taxonómicas de marmosinos. R. Bogdaschewsky y L. M. Pérez participaron en el procesamiento de las muestras. E. Massoia (†), M. Piantanida (†), O. Vaccaro y D. Verzi facilitaron el acceso a las colecciones consultadas. G. Navone y M. Lareschi brindaron equipo empleado en los trabajos de campo. C. Deschamps contribuyó en el resumen en inglés. Este trabajo ha sido solventado con fondos del Consejo Nacional de Investigaciones Científicas y Técnicas, la Fundación Antorchas, el Ministerio de Agricultura, Ganadería y Alimentación de la provincia de Buenos Aires y la Comisión de Investigaciones Científicas de la Provincia de Buenos Aires. A todas las personas e instituciones mencionadas, el especial reconocimiento de los autores, únicos responsables de posibles errores u omisiones.
LITERATURA CITADA
APFELBAUM LI y A BLANCO. 1984. Genetic similarity between species of Akodon (Rodentia-Cricetidae). Journal of Experimental Zoology 229:1-5. [ Links ]
APFELBAUM LI y OA REIG. 1989. Allozyme genetic distances and evolutionary relationships in species of akodontine rodents (Cricetidae: Sigmodontinae). Biological Journal of the Linnean Society 38:257-280. [ Links ]
AVERY DM. 1988. The Holocene environment of central South Africa: micromammalian evidence. Palaeoecology of Africa 19:335-345. [ Links ]
AVERY DM. 1997. Micromammals and the Holocene environment of RoseCottageCave. South African Journal of Science 93:445-448. [ Links ]
BARROS MA, RC LIASCOVICH, L GONZÁLEZ, MS LIZARRALDE y OA REIG. 1990. Banding pattern comparison between Akodon iniscatus, and Akodonpuer (Rodentia, Cricetidae). Zeitschrift für Säugetierkunde 55:115-127. [ Links ]
BELLOCQ MI. 2000. A review of the trophic ecology of the Barn Owl in Argentina. Journal of Raptor Research 34:108-119. [ Links ]
BIANCHI N y S MERANI. 1984. Cytogenetics of South American Akodont rodents (Cricetidae). X. Karyological distances at generic and intergeneric levels. Journal of Mammalogy 65:206-219. [ Links ]
BIANCHINI JJ y H DELUPI. 1975. Taxonomía experimental en cricétidos argentinos. I. Sobre el género Phyllotis (Waterhouse, 1837) (Rodentia, Cricetidae). Neotrópica 21(64):42-45. [ Links ]
BILENCA D, G ZULETA y F KRÁVETZ. 1994. Latitudinal variation in litter size and length of the breeding season in populations of pampas mice, Akodonazarae. Mammalia 58:563-568. [ Links ]
BIRNEY EC, RS SIKES, JA MONJEAU, N GUTHMANN y CJ PHILLIPS. 1996. Comments on Patagonian marsupials of Argentina. Pp. 149-154, en: Contribution in mammalogy, a memorial volume honoring Dr. J. Knox Jones Jr. (HH Genoways y RJ Baker, eds.). Museum of TexasTechUniversity Press. [ Links ]
BURGOS JJ y AL VIDAL. 1951. Los climas de la República Argentina, según la clasificación de Thornthwaite. Meteoros 1:3-32. [ Links ]
CABRERA A. 1961. Catálogo de los mamíferos de América del Sur. Parte II. Revista del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia," Ciencias Zoológicas 4:309‑732. [ Links ]
CABRERA AL. 1971. Fitogeografía de la República Argentina. Boletín de la Sociedad Argentina de Botánica 16:1-42. [ Links ]
CABRERA AL. 1976. Regiones Fitogeograficas Argentinas. Enciclopedia Argentina de Agricultura y Jardinería 2(1):1-85 . [ Links ]
CASTAÑEDA E y V BARROS. 1994. Las tendencias de la precipitación en el Cono Sur de América al Este de los Andes. Meteorológica 19:23-32. [ Links ]
CONTRERAS JR. 1968. Akodon molinae una nueva especie de ratón de campo del sur de la provincia de Buenos Aires. Zoología Platense, Investigaciones zoológicas y paleontológicas 1(2):9-12. [ Links ]
CONTRERAS JR. 1972. Nuevos datos acerca de la distribución de algunos roedores en las provincias de Buenos Aires, La Pampa, Entre Ríos, Santa Fe y Chaco. Neotrópica 18(55):27-30. [ Links ]
CONTRERAS JR. 1973a. La mastofauna de la zona de la Laguna Chasicó, provincia de Buenos Aires. Physis, Sección C 32(84):215‑219. [ Links ]
CONTRERAS JR. 1973b. Ecología del ratón de campo Akodon azarae en la región semiárida del sudoeste de Buenos Aires, Laguna Chasicó, Pdo. de Villarino. Deserta (Mendoza) 4:15-24. [ Links ]
CONTRERAS JR y LJ ALVARADO. 1968. Notas sobre los roedores cricétidos bonaerenses I Oryzomys nigripesflavescens en laguna Chasicó. Zoología Platense, Investigaciones zoológicas y paleontológicas 1(4):17-19. [ Links ]
CONTRERAS JR y E JUSTO. 1974. Aportes a la mastozoología pampeana. I. Nuevas localidades para roedores Cricetidae (Mammalia, Rodentia). Neotrópica 20(62):91-96. [ Links ]
CONTRERAS JR y A MACEIRAS. 1970. Relaciones entre tucu-tucos y los procesos del suelo en la región semiárida del sudoeste bonaerense. Agro, Publicación Técnica 12(17):3-17. [ Links ]
CONTRERAS JR y OA REIG. 1965. Datos sobre la distribución del género Ctenomys en la zona costera de la provincia de Buenos Aires comprendida entre Necochea y Bahía Blanca. Physis 25(69):169-186. [ Links ]
CRESPO JA. 1962. El ratón hocicudo rojizo, Oxymycterus rufus platensis. Neotrópica 8(27):116. [ Links ]
CRESPO JA. 1964a. Descripción de una nueva subespecie de roedor filotino (Mammalia). Neotrópica 10(33):99-101. [ Links ]
CRESPO JA. 1964b. Cita de mamíferos para el sudoeste de la provincia de Buenos Aires. Neotrópica 10(33):102. [ Links ]
CRISCI JV y MF LOPEZ ARMENGOL. 1983. Introducción a la teoría y práctica de la taxonomía numérica. Monografías de la OEA, Serie Biología, 26:1-129. [ Links ]
DACIUK J. 1974. Notas faunísticas y bioecológicas de Península Valdés y Patagonia. XII. Mamíferos colectados y observados en la Península Valdés y zona litoral de los golfos San José y Nuevo (provincia de Chubut, República Argentina). Physis, Sección C 33:23-39. [ Links ]
DE SANTIS LJM, E JUSTO y M KIN. 2001. Nuevos datos acerca de la distribución geográfica de Oligoryzomys longicaudatus (Rodentia: Sigmodontinae) en regiones extra-andinas de la República Argentina. Neotrópica 47:109-111. [ Links ]
FORNES A y E MASSOIA. 1965. Micromamíferos (Marsupialia y Rodentia) recolectados en la localidad bonaerense de Miramar. Physis 25(69):99‑108. [ Links ]
FRENGUELLI J. 1928. Observaciones geológicas en la región costanera sur de la Provincia de Buenos Aires. Anales de la Facultad de Ciencias de la Educación 2:1-145. [ Links ]
GALLIARI CA y UFJ PARDIÑAS. 2000. Taxonomy and distribution of the sigmodontine rodents of genus Necromys in central Argentina and Uruguay. Acta Theriologica 45:211-232. [ Links ]
GALLIARI CA, WD BERMAN y FJ GOIN.1991. Mamíferos. Pp. 1-35, en: Situación ambiental de la provincia de Buenos Aires. A: Recursos y rasgos naturales en la evaluación ambiental (HL López y EP Tonni, eds.). Comisión de Investigaciones Científicas Provincia de Buenos Aires, La Plata. [ Links ]
GALLIARI CA, UFJ PARDIÑAS y FJ GOIN. 1996. Lista comentada de los mamíferos argentinos. Mastozoología Neotropical 3:39-61 [ Links ]
GARCÍA ESPONDA CM, LJM DE SANTIS, JI NORIEGA, GO PAGNONI, GJ MOREIRA y M BERTELLOTTI. 1998. The diet of Tyto alba (Strigiformes: Tytonidae) in the lower Chubut valley (Argentina). Neotrópica 44:57-63. [ Links ]
GAUCH HG. 1982. Multivariate Analysis in Community Ecology. CambridgeUniversity Press, New York. [ Links ]
GHERSA CM, MA MARTINEZ-GHERZA y RJC LEON. 1998. Cambios en el paisaje pampeano y sus efectos sobre los sistemas de soporte de la vida. Pp. 38-71, en: Hacia una agricultura productiva y sostenible en la pampa (OT Solbrig y L Vainesman, comp.). Harvard University Press y Consejo Profesional de Ingeniería Agronómica, Buenos Aires. [ Links ]
GOIN FJ. 2001. Marsupiales (Didelphidae: Marmosinae y Didelphinae). Pp. 75-113, en: Cueva Tixi: cazadores y recolectores de las sierras de Tandilia Oriental. 1. Geología, Paleontología y Zooarqueología (D Mazzanti y C Quintana, eds.). Publicación de los editores, sin pie de imprenta. [ Links ]
GONÇALVES PR y JA DE OLIVEIRA. 2004. Morphological and genetic variation between two sympatric forms of Oxymycterus (Rodentia: Sigmodontinae): an evaluation of hypotheses of differentiation within the genus. Journal of Mammalogy 85:148-161. [ Links ]
GONZALEZ URIARTE M, F GONZALEZ MARTIN, H KRUGER, S LAMBERTO, G ARBANESI y VG de VERCESI. 1987. Evaluación expeditiva del recurso suelo y uso y cobertura de la tierra en el sur de la Provincia de Buenos aires. Estación Experimental Agropecuaria Hilario Ascasubi, Informe Técnico 28:1-75. [ Links ]
GONZÁLEZ MA. 1995. Diagnóstico ambiental de la provincia de Buenos Aires. Tomo I: aspectos básicos. Ediciones Banco Provincia, Buenos Aires. [ Links ]
GONZÁLEZ MA. 1997. Diagnóstico ambiental de la provincia de Buenos Aires. Tomo II: descripción de los sistemas ambientales (de Tandilia, interserranos, de Ventania, y de la vertiente austral de Ventania). Ediciones Banco Provincia, Buenos Aires. [ Links ]
HERSHKOVITZ P. 1955. South American marsh rats genus Holochilus, with a summary of sigmodont rodents. Fieldiana, Zoology 37:639‑673. [ Links ]
HERSHKOVITZ P. 1962. Evolution of Neotropical cricetine rodents (Muridae), with special reference to the Phyllotine Group. Fieldiana, Zoology 46:1‑524. [ Links ]
HERSHKOVITZ P. 1990. Mice of the Akodon boliviensis size class (Sigmodontinae, Cricetidae), with the description of two species from Brazil. Fieldiana, Zoology, new series 57:1‑35. [ Links ]
HOFFMANN JA. 1988. Las variaciones climáticas ocurridas en la Argentina desde fines del siglo pasado hasta el presente. Pp. 275-290, en: El deterioro del ambiente en la Argentina (suelo, agua, vegetación, fauna). Fundación para la educación de la ciencia y la cultura, Buenos Aires. [ Links ]
HOFFMANN FG, EP LESSA y MF SMITH. 2002. Systematics of Oxymycterus with description of a new species from Uruguay. Journal of Mammalogy 83:408-420. [ Links ]
HOLMBERG ES. 1898. La fauna de la República Argentina. Pp. 383-602, en: Segundo Censo de la República Argentina, 10-V-1895, Volumen I, Territorios, I, parte 6ª [ Links ].
KRISTENSEN J. 1992. Características microclimáticas de la Sierra de la Ventana y su relación con la vegetación. Tesis de doctorado inédita, Facultad de Ciencias Naturales y Museo, Universidad Nacional La Plata, La Plata, Argentina. [ Links ]
LAMBERTO S. 1987. Vegetación Natural. Pp. 51-53, en: Evaluación expeditiva del recurso suelo y uso y cobertura de la tierra en el sur de la Provincia de Buenos aires (M Gonzalez Uriarte, M González, H Kruger, S Lamberto, G Arbanesi y VG de Vercesi, comp.). Estación Experimental Agropecuaria Hilario Ascasubi, Informe Técnico. [ Links ]
LEÓN RJC, D BRAN, M COLLANTES, JM PARUELO y A SORIANO. 1998. Grandes unidades de vegetación de la Patagonia extra andina. Pp. 125-144. en: Ecosistemas patagónicos (M Oesterheld, MR Aguiar y JM Paruelo, eds.). Ecología Austral 8:75-308. [ Links ]
LUDWIG JA y JF REYNOLDS. 1988. Statistical Ecology. John Wiley and Sons, New York. [ Links ]
MASSOIA E. 1973. Presencia y rasgos bioecológicos de Oryzomys longicaudatus pampanus, nueva subespecie en la provincia de Buenos Aires (Mammalia - Rodentia - Cricetidae). Revista de Investigaciones Agropecuarias, INTA, Serie 1, Biología y Producción Animal 10(1):43-49. [ Links ]
MASSOIA E. 1976. Mammalia. En: Fauna de Agua Dulce de la República Argentina (R Ringuelet, dir.). Fundación Editorial Ciencia y Cultura, Buenos Aires 44:1-128. [ Links ]
MASSOIA E. 1985. Análisis de regurgitados de Asio flammeus del arroyo Chasicó. ACINTACNIA, Año 2(15):7‑9. [ Links ]
MASSOIA E. 1990. Roedores depredados por Tyto albatuidara en Mar del Tuyú, Partido de General Lavalle, Provincia de Buenos Aires. Boletín Científico, Asociación para la Protección de la Naturaleza 17:17-20. [ Links ]
MASSOIA E y A FORNES. 1964. Comentarios sobre Eligmodontia typus typus (Rodentia-Cricetidae). Physis 24(68):298. [ Links ]
MASSOIA E y UFJ PARDIÑAS. 1994. La depredación de mamíferos por Bubo virginianus y Tyto alba en Cerro Casa de Piedra, Lago Burmeister, Parque Nacional Perito Moreno, provincia de Santa Cruz. Boletín Científico, Asociación para la Protección de la Naturaleza 26:6-12. [ Links ]
MASSOIA E y AS VETRANO. 1988. Análisis de regurgitados de Tyto alba de Villa Regina, General Roca, provincia de Río Negro. Boletín Científico, Asociación para la Protección de la Naturaleza 3:10‑20. [ Links ]
MASSOIA E, A FORNES, R WAINBERG y TG de FRONZA. 1968. Nuevos aportes al conocimiento de las especies bonaerenses del género Calomys (Rodentia-Cricetidae). Revista de Investigaciones Agropecuarias, INTA, Serie 1, Biología y Producción Animal 5(4):63-92. [ Links ]
MASSOIA, E, S TIRANTI y AJ DIEGUEZ. 1997. Pequeños mamíferos depredados por Tytoalba en la provincia de La Pampa, según sucesivas recolecciones. Boletín Científico, Asociación para la Protección de la Naturaleza 32:19‑21. [ Links ]
MILLS J y J CHILDS. 1998. Ecologic studies of rodent reservoirs: Their relevance for human health. Emerging Infectious Diseases 4:529-537. [ Links ]
MONJEAU JA, RS SIKES, EC BIRNEY, N GUTHMANN y CJ PHILLIPS. 1997. Small mammal community composition within the major landscape divisions of Patagonia, southern Argentina. Mastozoología Neotropical 4:113-127. [ Links ]
MORELLO J. 1958. La provincia Fitogeográfica del Monte. Opera Lilloana 2:1-155 [ Links ]
MORRONE JJ. 2001. Biogeografía de América Latina y el Caribe. M&T-Manuales & Tesis SEA, Madrid. [ Links ]
MUSSER G y M CARLETON. 1993. Family Muridae. Pp. 501-756, en: Mammal species of the World. A taxonomic and geographic reference (DE Wilson y DAM Reeder, eds.). Smithsonian Institution Press, Washington. [ Links ]
MYERS P. 1989. A preliminary revision of the varius group of Akodon (A. dayi, dolores, molinae, neocenus, simulator, toba and varius). Pp. 5-54, en: Advances in Neotropical Mammalogy (KH Redford y JF Eisenberg , eds.). Sandhill Crane Press, Gainesville. [ Links ]
PALMA E y TL YATES. 1998. Phylogeny of southern South America mouse opossums (Thylamys, Didelphidae) based on alloenzyme and chromosomal data. Zeitschrift für Säugetierkunde 63:1-15. [ Links ]
PALMA RE, E RIVERA-MILLA, J SALAZAR-BRAVO, F TORRES-PÉREZ, UFJ PARDIÑAS, PA MARQUET, AE SPOTORNO, AP MEYNARD y TL YATES. En prensa. Phylogeography of Oligoryzomys longicaudatus (Rodentia: Sigmodontinae) in temperate South America. Journal of Mammalogy. [ Links ]
PARDIÑAS UFJ. 1993. El registro más antiguo (Pleistoceno temprano a medio) de Akodon azarae (Fischer, 1829) (Mammalia, Rodentia, Cricetidae) en la provincia de Buenos Aires, Argentina. Ameghiniana 30:149-153. [ Links ]
PARDIÑAS UFJ. 1999. Los roedores muroideos del Pleistoceno tardío-Holoceno en la región pampeana (sector este) y Patagonia (República Argentina): aspectos taxonómicos, importancia bioestratigráfica y significación paleoambiental. Tesis de doctorado inédita, Facultad de Ciencias Naturales y Museo, Universidad Nacional La Plata, La Plata, Argentina. [ Links ]
PARDIÑAS UFJ. 2001. Condiciones áridas durante el Holoceno temprano en el sudoeste de la provincia de Buenos Aires (Argentina): vertebrados y tafonomía. Ameghiniana 38:227-236. [ Links ]
PARDIÑAS UFJ y C DESCHAMPS. 1996. Sigmodontinos (Mammalia, Rodentia) pleistocénicos del sudoeste de la provincia de Buenos Aires (Argentina): aspectos sistemáticos, paleozoogeográficos y paleoambientales. Estudios Geológicos 52:367-379. [ Links ]
PARDIÑAS UFJ y CA GALLIARI. 1999. La presencia de Akodon iniscatus (Mammalia:Rodentia) en la provincia de Buenos Aires (Argentina). Neotropica 45:115-117. [ Links ]
PARDIÑAS UFJ y CA GALLIARI. 2001. Reithrodon auritus (Fischer, 1814). Mammalian Species, American Society of Mammalogists 664:1-8. [ Links ]
PARDIÑAS UFJ y MJ LEZCANO. 1995. Cricétidos (Mammalia, Rodentia) del Pleistoceno tardío del nordeste de la provincia de Buenos Aires (Argentina). Aspectos sistemáticos y paleoambientales. Ameghiniana 32:249-265. [ Links ]
PARDIÑAS UFJ, S CIRIGNOLI y DH PODESTA. 2001. Nuevos micromamíferos registrados en la Península de Valdés (provincia de Chubut, Argentina). Neotrópica 47:101-102. [ Links ]
PARDIÑAS UFJ, G MOREIRA, C GARCÍA-ESPONDA y LJM DE SANTIS. 2000. Deterioro ambiental y micromamíferos durante el Holoceno en el nordeste de la estepa patagónica (Argentina). Revista Chilena de Historia Natural 72:541-556. [ Links ]
PARDIÑAS UFJ, P TETA, S CIRIGNOLI y DH PODESTA. 2003. Micromamíferos (Didelphimorphia y Rodentia) de norpatagonia extra andina, Argentina: taxonomía alfa y biogeografía. Mastozoología Neotropical 10:69-113. [ Links ]
PINE R, P DALBY y J MATSON. 1985. Ecology, postnatal development, morphometrics, and taxonomic status of the short-tailed opossum, Monodelphis dimidiata, an apparently semelparous annual marsupial. Annals of CarnegieMuseum 54:195-231. [ Links ]
POLOP J. 1989. Distribution and ecological observations of wild rodents in Pampa de Achala, Córdoba, Argentina. Studies on Neotropical Fauna & Environment 24:53-59. [ Links ]
POLOP J, R MARTÍNEZ y M TORRES. 1985. Distribución y abundancia de poblaciones de pequeños roedores en la zona de embalse de Río Tercero, Córdoba. Historia Natural 5(5):33-44. [ Links ]
PREVOSTI FJ y UFJ PARDIÑAS. 2001. Variaciones corológicas de Lyncodonpatagonicus (Carnivora, Mustelidae) durante el Cuaternario. Mastozoología Neotropical 8:21-39 [ Links ]
RADAR. 2000. Cuestión de Imagen. Boletín Informativo Red Agroeconómica de Administración de Recursos, 2:7. [ Links ]
REAL R, M BARBOSA, D PORRAS, M KIN, A MARQUEZ, J GUERRERO, LJ PALOMO, E JUSTO y JM VARGAS. 2003. Relative importance of environment human activity and spatial situation in determining the distribution of terrestrial mamal diversity in Argentina. Journal of Biogeography 30:939-947. [ Links ]
REDFORD KH y JF EISENBERG. 1992. Mammals of the Neotropics. Volumen 2. The Southern Cone. Chile, Argentina, Uruguay, Paraguay. University of Chicago Press, Chicago. [ Links ]
REIG OA. 1964. Roedores y marsupiales del partido de General Pueyrredón y regiones adyacentes (provincia de Buenos Aires, Argentina). Publicaciones del Museo Municipal de Ciencias Naturales de Mar del Plata 1(6):203-224. [ Links ]
REIG OA. 1965. Datos sobre la comunidad de pequeños mamíferos de la región costera del Partido de General Pueyrredón y los partidos limítrofes. Physis 25(69): 205‑211. [ Links ]
REIG OA. 1978. Roedores cricétidos del Plioceno superior de la Provincia de Buenos Aires (Argentina). Publicaciones del Museo Municipal de Ciencias Naturales de Mar del Plata "Lorenzo Scaglia" 2(8):164‑190. [ Links ]
RINGUELET R. 1955. Panorama zoogeográfico de la provincia de Buenos Aires. Notas del Museo de La Plata 18(156):1-45. [ Links ]
RINGUELET R. 1961. Rasgos fundamentales de la zoogeografía de la Argentina. Physis 22(63):151‑170. [ Links ]
ROIG VG. 1989. Desertificación y distribución geográfica de mamíferos en la República Argentina. En: Desertificación, detección y control. Conferencias, trabajos y resultados del Curso Latinoamericano (F Roig, ed.). UNEP-IADIZA, Mendoza. [ Links ]
SALAZAR-BRAVO J, JW DRAGOO, DS TINNIN y TL YATES. 2001. Phylogeny and evolution of the neotropical rodent genus Calomys: Inferences from mitochondrial DNA sequence data. Molecular Phylogenetics and Evolution 20:173-184. [ Links ]
SIEGENTHALER GB, E FIORUCCI, S TIRANTI, P BORRAZ, M URIOSTE y A GARCÍA. 1990a. Informe de avance del plan de relevamiento de los vertebrados de la provincia de La Pampa. Agro Pampeano 18:38-48. [ Links ]
SIEGENTHALER GB, E FIORUCCI, S TIRANTI, P BORRAZ, M URIOSTE, J GOBBI y A GARCÍA. 1990b. Plan de relevamiento de los vertebrados de la provincia de La Pampa. Agro Pampeano 19:4-11. [ Links ]
SIEGENTHALER GB, S TIRANTI y CM DUCO. 1993. Relevamiento de los vertebrados de la provincia de La Pampa. Tercer informe. 5 Jornadas Pampeanas de Ciencias Naturales 1:139-147. [ Links ]
SIMONETTI JA. 1994. Impoverishment and netedness in caviomorph assemblages. Journal of Mammalogy 75:979-984. [ Links ]
SIMONETTI JA y B SAAVEDRA. 1997. Holocene variation in the small mammal fauna of central Chile. Zeitschrift für Säugetierkunde 63:58-62. [ Links ]
THEILER GR y NC GARDENAL. 1996. Patterns of evolution in Graomys griseoflavus (Rodentia, Cricetidae). II. Reproductive isolation between cytotypes. Journal of Mammalogy 77:776-784. [ Links ]
THOMAS O. 1919a. On small mammals collected by Sr. E. Budin in North-western Patagonia. Annals and Magazine of Natural History (London), 9, 3:199-212. [ Links ]
THOMAS O. 1919b. Two new Argentine species of Akodon. Annals and Magazine of Natural History (London), 9, 3:213-215. [ Links ]
THOMAS O. 1927. On further Patagonian mammals from Neuquen and the Rio Colorado collected by Señor E. Budin. Annals and Magazine of Natural History (London), 20, 17:199-205. [ Links ]
THOMAS O. 1929. The mammals of Señor Budin's Patagonian expedition, 1927-28. Annals and Magazine of Natural History (London), 10, 4:35-45. [ Links ]
TIRANTI SI. 1988. Análisis de regurgitados de Tytoalba de la provincia de La Pampa. Boletín Científico, Asociación para la Protección de la Naturaleza 11:8-12. [ Links ]
TIRANTI SI. 1989. Tres roedores cricétidos nuevos para la fauna de la provincia de La Pampa. 1 Jornada Nacional de Fauna Silvestre, Actas:489-494. [ Links ]
TIRANTI SI. 1998. Cytogenetics of Graomysgriseoflavus (Rodentia: Sigmodontinae) in central Argentina. Zeitschrift für Säugetierkunde 63:32-36. [ Links ]
WATERHOUSE GR. 1837. Characters of new species of the genus Mus, from the collection of Mr. Darwin. Proceedings of the Zoological Society of London 5(50-51):15-21. [ Links ]
WATERHOUSE GR. 1839. Mammalia, pt. 2, Rodents. Pp. 1-97 + 35 láms., en: The zoology of the voyage of H.M.S. Beagle, under the command of Captain Fitzroy, R.N., during the years 1832 to 1836 (C Darwin, ed.). London. [ Links ]
WITTOUCK P, E PINNA SENN, C SOÑEZ, M PROVENSAL, J POLOP y J LISANTI. 1995. Chromosomal and synaptonemal complex analysis of robertsonian polymorphisms in Akodon dolores and Akodon molinae (Rodentia, Cricetidae) and their hybrids. Cytologia 60:93-102. [ Links ]
Recibido octubre 2002.
Aceptación final setiembre 2004.

