










Servicios Personalizados
Revista

Articulo
 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. v.12 n.2 Mendoza jul./dic. 2005
Biologia reprodutiva de Delomys dorsalis (Hensel, 1872) - Rodentia, Sigmodontinae - em área de floresta ombrófila mista, Rio Grande do Sul, Brasil
Cristina V. Cademartori 1 , Marta E. Fabián 2 e João O. Manegheti 2
1 Pós-Graduação e Pesquisa, UNILASALLE, Av. Victor Barreto 2288, Canoas, RS, Brasil, CEP 92010-000. titina@via-rs.net. 2 Depto. Zoologia, Instituto de Biociências, UFRGS, av. Bento Gonçalves 9500, Porto Alegre, RS, Brasil, CEP 90540-000. mfabian@vortex.ufrgs.br, meneghet@vortex.ufrgs.br.
ABSTRACT: Reproductive Biology of Delomys dorsalis (HENSEL, 1872) -Rodentia, Sigmodontinae- in an area of mixed forest with conifers, at Rio Grande do Sul, Brasil. Delomys dorsalis is restricted to the wet tropical and subtropical forests of southern and southeastern Brazil and northeastern Argentina . The reproductive cycle of this species was studied by the histological examination of the reproductive tracts of animals caught in an area of mixed forest with conifers (29º 23' S, 50º 23' W), in southern Brazil , and from observations on a laboratory group of individuals. Fourteen expeditions were carried out from February 1997 to April 1998, during approximately 3 nights, using 140 live traps set on the ground and on branches between 1.5 and 2.0 m high. Reproductively active individuals were observed year round, although winter was marked by a reduction in breeding activity resulting from recruitment. Females showed a post-partum oestrus and a gestation time between 21 and 22 days. The litter size in captivity ranged from two to four, though pregnant females collected from the field had even five embryos. External reproductive features, frequently used as indicatives of the reproductive status of small mammals in ecological studies, were not accurate and underestimated the number of active animals in the population. Some individuals showed a remarkable scansorial ability that should be considered in future studies about population dynamics.
Key words. Delomys dorsalis. Mixed forest with conifers. Reproduction. Sigmodontinae. Southern Brazil.
INTRODUÇÃO
Os padrões demográficos exibidos pelas populações de pequenos mamíferos freqüentemente refletem mudanças sazonais, com flutuações na abundância e alterações na taxa de sobrevivência e na estrutura etária (Streilein, 1982). A compreensão desses padrões depende, dentre outros fatores, da identificação das estratégias reprodutivas apresentadas pelas espécies de pequenos mamíferos como resposta aos diferentes desafios impostos pelo ambiente. Segundo Lacher (1992), os agentes determinantes dos padrões reprodutivos em roedores murídeos foram consideravelmente estudados em zonas temperadas. Na América do Sul, as exíguas informações acerca da biologia reprodutiva dos roedores sigmodontinos apontam para padrões altamente variáveis. Entretanto, conforme Lacher (1992), extensas regiões do continente e grande parte da fauna carecem de estudos científicos. No Brasil, a maior parte das pesquisas sobre o tema tem sido desenvolvida no Cerrado (Mello, 1980; Dietz, 1983; Alho e Pereira, 1985; Alho et al . , 1986; Nitikman e Mares, 1987; Mares e Ernest, 1995; Talamoni e Dias, 1999) e na Mata Atlântica (Fonseca e Kierulff, 1989; Olmos, 1991; Pereira et al., 1993; Bergallo, 1995; Bergallo e Magnusson, 1999). Em áreas de Mata Atlântica do extremo sul do Brasil, contudo, os esforços empreendidos na tentativa de compreender a dinâmica das populações de pequenos roedores têm sido insuficientes. Este estudo visa a fornecer informações a respeito da biologia reprodutiva de Delomys dorsalis , espécie restrita às florestas úmidas tropicais e subtropicais do sul e sudeste do Brasil, e do nordeste da Argentina, cujos hábitos permanecem praticamente desconhecidos (Voss, 1993). Especificamente, buscou-se identificar o período reprodutivo da espécie, através da análise histológica das gônadas de machos e fêmeas; observar características morfológicas externas de ambos os sexos que indicassem a condição reprodutiva, associando-as com o estado de maturação das células gaméticas; determinar o período de gestação e o número de filhotes por ninhada; definir o período de amamentação e registrar os principais eventos ontogenéticos dos filhotes.
ÁREA DE ESTUDO
A área de estudo localiza-se no município de São Francisco de Paula, Rio Grande do Sul, em uma unidade de conservação do Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA), a Floresta Nacional de São Francisco de Paula (29º23'S, 50º23'W), incluída entre os ecossistemas que compõem a Reserva da Biosfera da Mata Atlântica no Estado desde 1993 (Marcuzzo et al., 1998). Essa unidade compreende uma área de 1606,7 ha, 901,9 dos quais são cobertos por Floresta Ombrófila Mista, cuja espécie dominante é Araucaria angustifolia . Uma descrição detalhada da área de estudo é encontrada em Cademartori et al. (2002).
O tipo climático predominante na região, segundo o sistema geral de Köppen-Geiger, é o subtropical úmido, classificado como Cfb, com temperaturas compreendidas entre -3 °C e 18 °C para o mês mais frio, entre 18,3 °C e 27 °C para o mês mais quente, e temperatura média anual igual a 18,5 °C (Galeti, 1972). A Floresta Nacional de São Francisco de Paula, situada a 930 m acima do nível do mar, junto à borda da escarpa do Planalto das Araucárias, localiza-se no centro da região de maior ocorrência do clima superúmido a úmido, o que resulta numa pluviometria com totais elevados (>2200 mm/ano) e distribuídos ao longo de todos os meses do ano (IBGE, 1986). É comum a ocorrência de geadas no outono e no inverno, bem como a precipitação de neve nos meses mais frios. Variações na precipitação e temperatura ao longo do período de estudo, em relação às médias obtidas para 30 anos (dados do Instituto Estadual de Meteorologia), podem ser observadas na Fig. 1 .

Fig. 1. Precipitação e temperatura média obtidas durante o período de estudo (janeiro/97 a abril/98) no município de São Francisco de Paula (29º20'S, 50º31'W), em relação às médias para 30 anos (1931 a 1961).
MATERIAIS E MÉTODOS
Atividades de Campo e Laboratório
Coletaram-se indivíduos para o exame histológico das gônadas, mensalmente, de fevereiro de 1997 a abril de 1998. Capturas no terreno e no sub-bosque (entre 1.5 e 2.0 m de altura) foram efetuadas com armadilhas de arame galvanizado do tipo gaiola, de tamanhos 9x9x22 cm e 14x14x30 cm. Utilizaram-se 140 armadilhas distribuídas em pontos eqüidistantes, sob a forma de grade, 70 ao nível do terreno e 70 no sub-bosque, os quais se constituíram em estações fixas de captura (a distância entre as estações foi de aproximadamente 10 m).
Como isca, foram empregadas fatias de milho-verde cobertas com pasta de amendoim. As armadilhas foram expostas, em média, durante 3 noites consecutivas e revisadas diariamente, objetivando-se substituir as iscas e solucionar eventuais imprevistos.
Os espécimes coletados, depositados na coleção científica do Laboratório de Mastozoologia do Museu de Ciências e Tecnologia da Pontifícia Universidade Católica do Rio Grande do Sul (PUCRS), foram medidos (de acordo com Moojen, 1943) e pesados. Também foram observadas características externas indicadoras do estado reprodutivo (posição dos testículos nos machos, abertura do orifício vaginal e tamanho dos mamilos nas fêmeas). A seguir, foram fixados em formol 10% e conservados em álcool 70%. A preparação e a análise histológica das gônadas transcorreram de acordo com a seqüência de procedimentos de rotina descritos sinteticamente a seguir.
a) Fixação : as peças foram fixadas em formol 10% por um período mínimo de 24 h;
b) Desidratação : sofreram um processo de desidratação, passando por uma bateria alcoólica ascendente;
c) Diafanização : foram mergulhadas em xilol;
d) Confecção dos blocos de parafin a: as peças foram incluídas em parafina líquida;
e) Seccionamento : os blocos de parafina foram seccionados, obtendo-se cortes semi-seriados com aproximadamente 5 ìm de espessura;
f) Coloração : após a montagem das lâminas, procedeu-se à coloração com Hematoxilina/Eosina.
A análise histológica, para a determinação do estado de maturação das células gaméticas, restringiu-se ao testículo e epidídimo esquerdos, para os machos, e abrangeu toda a amostra de indivíduos coletados (n=131). No caso das fêmeas, ambos os ovários foram seccionados; de um total de 110 espécimes, uma subamostra de 88 fêmeas foi observada quanto à presença de corpos lúteos, folículos pré-ovulatórios (de Graaf ou vesiculares) ou em estágios iniciais de desenvolvimento (primários e secundários). Em fêmeas grávidas, identificou-se o número de fetos ou embriões visíveis em cada corno uterino.
Observações em Cativeiro
Com o objetivo de se determinar o período de gestação, observar o cuidado parental, assim como o número de filhotes gerados, e aspectos do desenvolvimento ontogenético, foram mantidos em cativeiro 10 casais extraídos da área de estudo. Procurando-se minimizar as condições de estresse, os animais foram mantidos em caixas plásticas médias (comumente utilizadas na manutenção de cobaias), nas dependências do Biotério da PUCRS, de setembro de 1996, quando foram capturados, a agosto de 1998, quando houve perda total do grupo por intoxicação alimentar causada por ração contaminada. Mensalmente, os animais eram pareados e mantidos juntos durante 24 h, sendo então recolocados em caixas individuais. Durante os períodos em que foram obtidas ninhadas, os comportamentos envolvidos no cuidado parental foram observados qualitativamente e registrados, diariamente.
Análise Quantitativa
Utilizou-se o programa estatístico GraphPad Instat versão 3.01 na aplicação dos testes estatísticos relacionados a seguir.
1. Índice de Abundância
No intuito de se comparar a abundância de machos sexualmente ativos entre si, assim como a de inativos, ao longo dos meses, optou-se pelo uso de um índice de abundância, uma vez que os esforços de captura foram distintos no decorrer do estudo. Para a quantificação do esforço, levou-se em conta apenas o número de armadilhas efetivamente disponível para os machos. As fêmeas foram excluídas da avaliação quantitativa pelo fato da análise histológica das gônadas não ter abrangido a amostra completa. Logo, as armadilhas ocupadas pelas fêmeas, por outras espécies, ou indisponíveis por outros fatores, foram desconsideradas. O índice de abundância consistiu na razão entre o número de machos capturados (sexualmente ativos ou inativos) e o número de armadilhas disponível a cada expedição.
2. Estatística Não-Paramétrica
A prova de Kruskal-Wallis (KW) foi aplicada para testar a hipótese de ausência de diferenças entre mais de duas séries de índices de abundância, enquanto a prova de Mann-Whitney (U) permitiu a comparação entre duas séries. O teste complementar de Dunn possibilitou a identificação das séries que se diferenciaram significativamente.
A correlação entre a abundância de machos sexualmente ativos e a de inativos foi determinada pelo coeficiente de correlação de Spearman (r).
3. Estatística Paramétrica
Foram elaboradas curvas de distribuição de freqüência do comprimento cabeça-corpo de toda a amostra coletada durante o estudo, com o objetivo de se identificar períodos de recrutamento de jovens ao longo do ano. A normalidade das distribuições foi testada por meio da prova Kolmogorov-Smirnov (KS). Os graus de assimetria (g 1 ) e de curtose (g 2 ) também foram testados, considerando-se significativas as diferenças para valores de p<0,05 (Zar 1999). As médias de comprimento cabeça-corpo dos indivíduos capturados no outono-inverno e na primavera-verão seguinte foram comparadas por meio do teste t para amostras independentes, com correção de Welch. Empregou-se, ainda, ANOVA um fator com teste complementar Student-Newman-Keuls (SNK) no cotejo das médias mensais do período outono-inverno.
RESULTADOS E DISCUSSÃO
Verificaram-se flutuações na abundância de 46 machos sexualmente ativos, ao longo de 14 meses de estudo ( Fig. 2A ). Comparando-se os períodos fevereiro/97-março/97, maio/97-junho/97, agosto/97-setembro/97 e outubro/97-janeiro/98, encontrou-se uma diferença significativa (KW=7,064; p=0,0222). Houve um aumento gradativo na abundância de machos sexualmente ativos de fevereiro a setembro de 1997, interrompido pela ausência desta categoria no mês de julho/97. Após setembro/97, houve uma queda acentuada nos meses subseqüentes do mesmo ano. Os índices de abundância referentes ao período agosto/97-setembro/97, mais elevados, diferiram significativamente de outubro/97-janeiro/98. Entretanto, os períodos fevereiro/97-março/97, maio/97-junho/97 e agosto/97-setembro/97 não diferiram entre si, assim como os dois primeiros não diferiram de outubro/97-janeiro/98.

Fig. 2. Índice de abundância de machos de Delomys dorsalis capturados na Floresta Nacional de São Francisco de Paula (29º23'S, 50º23'W) de fevereiro/97 a abril/98: A - sexualmente ativos, B - sexualmente inativos.
Os machos sexualmente inativos (n=85), representados na Fig. 2B , apresentaram uma diferença significativa quanto aos índices de abundância relativos aos períodos maio/97-agosto/97, mais elevados, e setembro/97-janeiro/98 (U=16,00; p=0,0286). Houve uma nítida predominância de machos inativos durante o outono e inverno.
O coeficiente de correlação de Spearman obtido (r=0,2233; p=0,4429) demonstrou a inexistência de associação entre os índices de abundância de machos sexualmente ativos e inativos, ou seja, um aumento no número de machos ativos não correspondeu a uma equivalente diminuição no número de inativos e vice-versa. Machos sexualmente ativos foram encontrados ao longo de todas as estações, embora em proporções diferenciadas, indicando uma estratégia reprodutiva do tipo panestacional.
O incremento de machos inativos durante o outono e inverno de 1997 deveu-se ao recrutamento de jovens, que, embora tenha ocorrido ao longo de todo o ano, se intensificou nessa fase. As curvas de distribuição de freqüência dos intervalos de comprimento cabeça-corpo apresentadas na Fig. 3 expressam a coexistência de jovens e adultos ao longo do ano inteiro, uma vez que se ajustam à função normal ( Fig. 3A : KS=0,07776, p>0,05; g 1 =0,0102, p>0,05; g 2 =0,1788, p>0,05 e Fig. 3B: KS=0,1135, p>0,05; g 1 =0,4145, p>0,05; g 2 =0,5374, p>0,05). Comparando-se os indivíduos capturados de maio/97 a agosto/97 ( Fig. 3A ) com os capturados de setembro/97 a março/98 ( Fig. 3B ), constatou-se a existência de intervalos de classe de menor comprimento no primeiro período, ausentes no segundo. Acrescentando-se a isso, o comprimento médio da amostra do outono-inverno de 1997 (média=109,77 mm; n=159) diferiu significativamente daquele obtido na primavera-verão subseqüente (média=115,59 mm; n=58), corroborando a hipótese do recrutamento prevalente de maio/97 a ago/97 (t=4,494, p<0,0001). É importante destacar que este período coincide com a fase de maturação das pinhas na Floresta Ombrófila Mista e, conseqüentemente, com a grande disponibilidade de um importante recurso alimentar exclusivo desse ambiente, os pinhões (sementes produzidas por Araucaria angustifolia ). Os estudos de Müller (1986) sobre a influência dos roedores e aves na regeneração da A. angustifolia em uma área de floresta nativa, próxima a Curitiba, no Estado do Paraná, apontam os pequenos roedores como os principais responsáveis pelo consumo de pinhões (em 20 transectos de 100 m 2 , 48% dos pinhões foram predados por pequenos roedores). Enfatiza-se, ainda, a redução da amplitude de comprimento cabeça-corpo na primavera-verão, resultante da ausência de indivíduos mais velhos, o que sugere um rápido rejuvenescimento da população.

Fig. 3. Curvas de distribuição de freqüência do comprimento cabeça-corpo (em mm) de Delomys dorsalis capturados na Floresta Nacional de São Francisco de Paula (29º23'S, 50º23'W): A - maio/97 a agosto/97, B - setembro/97 a março/98.
Da comparação dos comprimentos médios mensais das amostras coletadas de maio/97 a agosto/97 ( Tabela 1 ), mediante a aplicação de ANOVA um fator - SNK, resultou uma diferença significativa entre a média de junho/97, inferior, e as demais (p<0,0001). Dessa forma, é possível inferir que o pico do recrutamento ocorreu no referido mês. A possibilidade de uma breve redução da atividade reprodutiva no período de inverno, todavia, não pode ser descartada, posto que em julho/97, por exemplo, apareceram apenas machos inativos, apesar de serem encontradas fêmeas com folículos vesiculares ( Fig. 4 ). Krebs e Delong (1965) relatam a inibição da atividade sexual em subadultos de Microtus californicus , particularmente nos machos, em uma população com elevadas taxas de crescimento individual e populacional. Outra evidência pode ser verificada pela baixa freqüência de fêmeas grávidas em junho/97 e sua total ausência nos meses subseqüentes do período ( Fig. 4B ). Tal categoria só ressurgiu a partir de outubro/97, um mês após o reaparecimento de machos com espermatozóides no epidídimo ( Fig. 4A ). Há duas hipóteses que podem ser consideradas para explicar esse fenômeno: a primeira refere-se à maturação sexual tardia em machos de pequenos roedores, comparativamente às fêmeas (Fleming, 1971; Rickart, 1977; De Villafañe, 1981; Aguilera, 1985). Talvez, os machos recrutados no final do outono e meados do inverno tornem-se aptos à reprodução somente com a proximidade da primavera. Machos em início de atividade reprodutiva (princípio de espermiogênese nos túbulos seminíferos) foram observados em agosto/97, sendo que em setembro/97, em sua maioria, já apresentavam intensa espermiogênese nos túbulos seminíferos e moderada quantidade de espermatozóides no epidídimo ( Fig. 4A ). A segunda hipótese, complementar à primeira, está apoiada na ação de fatores climáticos, provavelmente das baixas temperaturas típicas do inverno, que reduziriam a probabilidade de sobrevivência da prole. Conforme Streilein (1982), a reprodução exige um investimento considerável de tempo e energia e o êxito reprodutivo depende da minimização do esforço empreendido em épocas inapropriadas. Fatores exógenos, tais como fotoperíodo, temperatura e precipitação, estão freqüentemente correlacionados com o sucesso ou o insucesso reprodutivo em pequenos mamíferos.
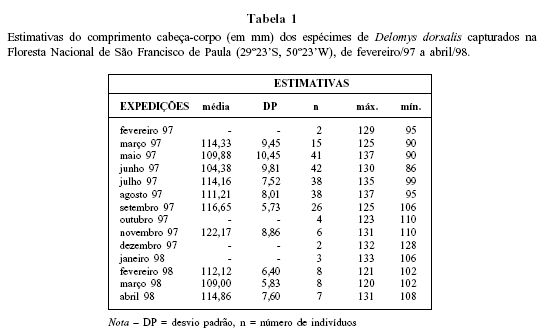

Fig. 4. Condição reprodutiva dos espécimes de Delomys dorsalis capturados na Floresta Nacional de São Francisco de Paula (29º23'S, 50º23'W) de fevereiro/97 a abril/98: A - machos, B - fêmeas.
Olmos (1991) trabalhou em área de Floresta Atlântica, no Estado de São Paulo, a uma altitude de 850 m e condições climáticas semelhantes, registrando D. dorsalis como a espécie mais abundante. Sugere que a reprodução se interrompa durante o inverno (maio-agosto), com a redução na disponibilidade de alimento, indicando que os indivíduos de mais baixo peso foram capturados nesse período. As informações obtidas nesta investigação, contudo, tornam tal afirmação incompleta, pois o autor, ao atribuir a inatividade reprodutiva e o mais baixo peso, exclusivamente, à redução na disponibilidade de alimento, exclui a possibilidade desse resultado ser fortemente influenciado pelo recrutamento. Chama-se a atenção para o fato de ambas as áreas terem revelado o mesmo padrão, apesar da oferta diferencial de alimento. Como já foi mencionado, o outono-inverno é caracterizado pela disponibilidade de um importante recurso alimentar na Floresta Ombrófila Mista, os pinhões, ausentes na área estudada por Olmos. Conclui-se, então, que a inatividade reprodutiva descrita pelo referido autor, provavelmente, não seja explicada, unicamente, pela falta de alimento, mas possa ser atribuída, também, à incorporação de jovens ainda imaturos à população.
A partir de outubro/97, a acentuada redução na abundância de machos sexualmente ativos e inativos ( Fig. 2 ), bem como a diminuição no número de fêmeas registradas no local de estudo ( Fig. 4B ), podem estar associadas a uma ampliação da área de domínio vital e à ocupação do estrato arbóreo. Essa hipótese fundamenta-se na captura de espécimes no sub-bosque (20% do total amostrado), na observação de indivíduos escalando rapidamente árvores de mais de 20 m de altura, e na análise de Voss (1993), que estabelece correspondência entre determinadas características morfológicas da espécie e a habilidade escansorial, não obstante Carvalho (1965), Olmos (1991) e Fonseca et al. (1996) apontem D. dorsalis como exclusivamente ou essencialmente terrestre. Davis (1945) refere-se à possibilidade de ocupação do estrato arbóreo por pequenos roedores em função da flutuação sazonal na disponibilidade de recursos. Fonseca e Kierulff (1989), Bergallo e Magnusson (1999) relatam que as flutuações populacionais intra-anuais parecem ser reguladas pela disponibilidade de alimento (frutos ou artrópodos) em ecossistemas tropicais, influenciada pela dinâmica das chuvas. A hipótese apresentada, contudo, deve ser testada por meio da amostragem do estrato arbóreo propriamente dito e do estudo simultâneo da flutuação dos recursos disponíveis na área, principalmente durante a primavera-verão. Outrossim, não se pode descartar a interferência de outros fatores na dinâmica populacional, tais como a predação e a emigração.
Possivelmente, variações interanuais na disponibilidade de recursos e em outros fatores ecológicos sejam também responsáveis pelos padrões observados e, embora existam indivíduos sexualmente ativos ao longo do ano inteiro, uma intensificação na atividade reprodutiva, em períodos favoráveis, pode maximizar a sobrevivência da prole. A elevada abundância de machos sexualmente ativos em fevereiro/98 ( Fig. 2A ), assim como a captura exclusiva de fêmeas grávidas em fevereiro/97- março/97 ( Fig. 4B ), parecem sugerir um pico de atividade reprodutiva no final do verão.
As análises histológicas detalhadas efetuadas nas gônadas das fêmeas e as observações de animais em cativeiro permitem afirmar que as mesmas apresentam estro pós-parto. Tal hipótese é demonstrada pela existência de fêmeas grávidas, exibindo, simultaneamente, corpos lúteos e folículos vesiculares no ovário. Acrescente-se a isso a observação, em cativeiro, de duas fêmeas, que durante o final do período de amamentação, obtiveram uma segunda ninhada. O estro pós-parto já foi relatado para várias espécies de pequenos roedores (Jameson, 1953; Richmond e Conaway, 1969; Lee et al., 1970; Rickart, 1977; Aguilera, 1985; Nelson, 1985) e, conforme Conaway (1971), constitui-se num mecanismo que permite aumentar a produtividade em pequenos mamíferos, cuja expectativa de vida é curta.
A partir da análise de 16 fêmeas grávidas com implantações visíveis, foram evidenciados 27 embriões no corno uterino direito e 29 no esquerdo, considerando-se uma variação de dois a cinco embriões por fêmea. Davis (1947) registrou uma variação de dois a quatro embriões para cinco fêmeas grávidas coletadas. Do total de fêmeas analisadas, sete apresentaram um maior número de implantações no corno esquerdo, seis, no direito, e em três delas o número de implantações foi idêntico nos dois cornos uterinos. Embora Pereira et al. (1993) tenham relatado uma clara tendência de implantação no corno direito para três espécies de cricetídeos da Floresta Atlântica, sugerindo uma maior atividade ou sucesso de fertilização do ovário correspondente, isso parece inválido para D. dorsalis.
Das características comumente referidas na literatura como indicadoras de atividade reprodutiva em fêmeas, a que se mostrou mais confiável foi a presença de mamas conspícuas, posto que das 60 fêmeas sexualmente ativas, em cujos ovários foram encontrados corpos lúteos ou folículos vesiculares, 55% apresentaram tal característica. A abertura do orifício vaginal, todavia, não foi fidedigna, tendo sido observada em apenas 10% da amostra analisada. A posição dos testículos, nos machos, também não permitiu uma avaliação precisa. Em apenas 20 dos 46 machos definidos como sexualmente ativos mediante a análise histológica, os testículos encontraram-se em posição escrotal. Os estudos de McCravy e Rose (1992) apontam a posição dos testículos como um indicativo relativamente confiável (87-94%), contrariamente aos resultados aqui obtidos. As características externas das fêmeas também se mostraram pouco consistentes, sendo o tamanho dos mamilos a mais fiel.
Desenvolvimento Ontogenético
em Cativeiro
Em cativeiro, foi obtido um total de oito ninhadas ao longo dos meses de abril e novembro de 1997, janeiro, fevereiro, março, junho (n=2) e agosto de 1998. O período de gestação, para quatro fêmeas acompanhadas desde o dia do acasalamento, ficou compreendido entre 21 e 22 dias, não discrepando de dados citados para outras espécies de pequenos roedores (Aguilera, 1985). Das oito gestações, cinco resultaram em apenas dois filhotes, duas resultaram em três e uma, em quatro. Os filhotes nasceram com os olhos fechados, desprovidos de pêlos, mas com o dorso parcialmente pigmentado. O peso médio ao nascimento, para as fêmeas, foi de 3.8 g ± 0.31 (n=9) e, para os machos, foi de 4.1 g ± 0.81 (n=8). O dorso e as orelhas tornaram-se intensamente pigmentados até o quinto dia. No sétimo dia, o dorso apresentou-se densamente coberto por pêlos e a listra dorsal, característica da espécie, nítida. Neste mesmo dia, os incisivos inferiores erupcionaram. A erupção dos superiores, todavia, deu-se entre o 10º e o 11º dias, período em que o ventre se tornou densamente coberto por uma pelagem branco-acinzentada. Os filhotes abriram os olhos entre o 12º e o 13º dias, e entre o 16º e o 17º dias saíram do ninho, na companhia da mãe, alimentando-se de frutos e sementes. O término do período de amamentação foi registrado somente entre o 22º e o 25º dias.
CONCLUSÕES
A reprodução, em D. dorsalis , ocorreu ao longo de todo o ano, observando-se, entretanto, uma redução no período de inverno, em decorrência do recrutamento de jovens. Diferentemente da maioria dos ecossistemas, o outono-inverno, na Floresta Ombrófila Mista, é caracterizado pela abundância de um importante recurso alimentar, os pinhões. O curto período de gestação e o estro pós-parto constatados constituem estratégias vantajosas em ambientes sob condições instáveis. As características externas (posição dos testículos, abertura do orifício vaginal e tamanho dos mamilos), comumente utilizadas como indicadoras do estado de maturação sexual, revelaram-se pouco consistentes, pois subestimaram a magnitude da atividade reprodutiva na população.
AGRADECIMENTOS
À Capes, pelo suporte financeiro concedido na forma de bolsa; ao Unilasalle, pela concessão de auxílio destinado ao aprimoramento acadêmico; ao IBAMA, especialmente ao Eng. Florestal Artur José Soligo, administrador da FLONA de São Francisco de Paula e demais funcionários, pelo auxílio e cooperação; às colegas Rosane Vera Marques e Susi Missel Pacheco, pela colaboração sistemática nas atividades de campo; ao Eng. Eletricista Fernando Ramos, pelo inestimável apoio técnico; ao Prof. Júlio César González, coordenador do Laboratório de Mastozoologia do MCT/PUCRS, pelo apoio e estímulo constantes; ao biólogo Marcos Machado, pelas contribuições e revisão do manuscrito.
LITERATURA CITADA
AGUILERA MM. 1985. Growth and reproduction in Zygodontomys microtinus (Rodentia, Cricetidae) from Venezuela in a laboratory colony. Mammalia 49:75-83. [ Links ]
ALHO CJR e LA PEREIRA. 1985. Population ecology of a cerrado rodent community in central Brazil. Revista Brasileira de Biologia 45:597-607. [ Links ]
ALHO CJR, LA PEREIRA e AC PAULA. 1986. Patterns of habitat utilization by small mammal populations in Cerrado biome of central Brazil . Mammalia 50:447-460. [ Links ]
BERGALLO HG. 1995. Comparative life-history characteristics of two species of rats, Proechimys iheringi and Oryzomys intermedius , in an Atlantic Forest of Brazil. Mammalia 59:51-64. [ Links ]
BERGALLO HG e WE MAGNUSSON. 1999. Effects of climate and food availability on four rodent species in southeastern Brazil . Journal of Mammalogy 80:472-486. [ Links ]
CADEMARTORI CV, RV MARQUES, SM PACHECO, LRM BAPTISTA e M GARCIA. 2002. Roedores ocorrentes em Floresta Ombrófila Mista (São Francisco de Paula, Rio Grande do Sul) e a caracterização do seu habitat. Comunicações do Museu de Ciências e Tecnologia - PUCRS, Serie Zoologia 15:61-86. [ Links ]
CARVALHO CT. 1965. Bionomia de pequenos mamíferos em Boracéia. Revista de Biologia Tropical 13:239-257. [ Links ]
CONAWAY CH. 1971. Ecological adaptation and mammalian reproduction. Biology of Reproduction 4:239-247. [ Links ]
DE VILLAFAÑE G. 1981. Reproduccion y crecimiento de Akodon azarae azarae (Fischer, 1829). Historia Natural 1:193-204. [ Links ]
DAVIS DA. 1945. The annual cycle of plants, mosquitoes, birds and mammals in two Brazilian forests. Ecological Monographs 15:243-295. [ Links ]
DAVIS DA. 1947. Notes on the life histories of some Brazilian mammals. Boletim do Museu Nacional (76):1-8. [ Links ]
DIETZ JM. 1983. Notes on the natural history of some small mammals in central Brazil . Journal of Mammalogy 64:521-523. [ Links ]
FLEMING TH. 1971. Population ecology of three species of Neotropical rodents. Miscellaneous Publications Museum of Zoology (143):1-77. [ Links ]
FONSECA GAB e MCM KIERULFF. 1989. Biology and natural history of Brazilian Atlantic Forest small mammals. Bulletin of the Florida State Museum 34:99-152. [ Links ]
FONSECA GAB, G HERRMANN, YLR LEITE, RA MITTERMEIER , AB RYLANDS e JL PATTON. 1996. Lista anotada dos mamíferos do Brasil. Occasional Papers in Conservation Biology (4):1-38. [ Links ]
GALETI PA. 1972. Conservação do solo, reflorestamento, clima. Impres, Campinas, SP. [ Links ]
IBGE. 1986. Folha SH.22 Porto Alegre e parte das folhas SH.21 Uruguaiana e SI.22 Lagoa Mirim: Geologia, Geomorfologia, Pedologia, Vegetação, Uso Potencial da Terra. IBGE, Rio de Janeiro . [ Links ]
JAMESON EW. 1953. Reproduction of Deer Mice ( Peromyscus maniculatus and P. boylei ) in the Sierra Nevada, California . Journal of Mammalogy 34:44-58. [ Links ]
KREBS CJ e KT DELONG. 1965. A Microtus population with supplemental food. Journal of Mammalogy 46:566-573. [ Links ]
LACHER TE. 1992. Ecological aspects of reproductive patterns in South American small rodents. Pp. 283-294, in : Reproductive biology of South American vertebrates (WC Hamlett, ed.). Springer-Verlag , New York . [ Links ]
LEE C, DJ HORVATH, RW METCALFE e EK INSKEEP. 1970. Ovulation in Microtus pennsylvanicus in a laboratory environment. Laboratory Animal Care 20:1098-1102. [ Links ]
MARCUZZO S, SM PAGEL e MIS CHIAPPETTI. 1998. A Reserva da Biosfera da Mata Atlântica no Rio Grande do Sul. Situação atual, ações e perspectivas. Conselho Nacional da Reserva da Biosfera da Mata Atlântica, São Paulo. [ Links ]
MARES MA e KA ERNEST. 1995. Population and community ecology of small mammals in a gallery forest of central Brazil . Journal of Mammalogy 76:750-768. [ Links ]
McCRAVY KW e RK ROSE. 1992. An analysis of external features as predictors of reproductive status in small mammals. Journal of Mammalogy 73:151-159. [ Links ]
MELLO DA. 1980. Estudo populacional de algumas espécies de roedores do Cerrado (norte do município de Formosa, Goiás). Revista Brasileira de Biologia 40:843-860. [ Links ]
MOOJEN J. 1943. Captura e preparação de pequenos mamíferos para coleções de estudo. Imprensa Nacional, Rio de Janeiro. [ Links ]
MÜLLER JA. 1986. A influência dos roedores e aves na regeneração da Araucaria angustifolia (Bert.) O. Ktze. Dissertação de Mestrado, Setor de Ciências Agrárias, Universidade Federal do Paraná, Curitiba, PR, Brasil. [ Links ]
NELSON RJ. 1985. Photoperiod influences reproduction in the Prairie Vole ( Microtus ochrogaster ). Biology of Reproduction 33:596-602. [ Links ]
NITIKMAN LZ e MA MARES. 1987. Ecology of small mammals in a gallery forest of central Brazil . Annals of Carnegie Museum 56:75-95. [ Links ]
OLMOS F. 1991. Observations on the behavior and population dynamics of some Brazilian Atlantic Forest rodents. Mammalia 55:555-565. [ Links ]
PEREIRA LA, WA CHAGAS e JE da COSTA. 1993. Ecologia de pequenos mamíferos silvestres da Mata Atlântica, Brasil. I. Ciclos reprodutivos de Akodon cursor , Nectomys squamipes e Oryzomys nigripes (Rodentia, Cricetinae). Revista Brasileira de Zoologia 10:389-398. [ Links ]
RICHMOND M e CH CONAWAY. 1969. Management, breeding and reproductive performance of the vole, Microtus ochrogaster , in a laboratory colony. Laboratory Animal Care 19:80-87. [ Links ]
RICKART EA. 1977. Reproduction, growth and development in two species of cloud forest Peromyscus from southern Mexico . Occasional Papers Museum of Natural History (67):1-22. [ Links ]
STREILEIN KE. 1982. The ecology of small mammals in the semiarid Brazilian Caatinga. III. Reproductive biology and population ecology. Annals of Carnegie Museum 51:251-269. [ Links ]
TALAMONI SA e MM DIAS. 1999. Population and community ecology of small mammals in southeastern Brazil . Mammalia 63:167-181. [ Links ]
VOSS RS. 1993. A revision of the brazilian muroid rodent genus Delomys with remarks on "Thomasomyine" characters. American Museum Novitates (3073):1-44. [ Links ]
ZAR JH. 1999. Biostatistical Analysis. Prentice Hall, New Jersey . [ Links ]
Recibido 16 marzo 2005.
Aceptación final 1º julio 2005.

