










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkMastozoología neotropical
versão impressa ISSN 0327-9383versão On-line ISSN 1666-0536
Mastozool. neotrop. v.13 n.1 Mendoza jan./jun. 2006
Small mammal selection and functional response in the diet of the maned wolf, Chrysocyon brachyurus (Mammalia: Canidae), in southeast Brazil
Adriana A. Bueno and José Carlos Motta-Junior
Departamento de Ecologia, Instituto de Biociências, Universidade de São Paulo, 05508-900, São Paulo, SP, Brazil. < abueno@ib.usp.br>
ABSTRACT: Data reported here are intended to describe patterns of predation by a solitary hunter, as well as to bring new insights into the predator-prey relationship in the Neotropical region. The goal of this study was to verify if there was small mammal selection in the diet of the maned wolf. Also, we attempt to test for a functional response in relation to the consumption of small mammals. The study was carried out in the Ecological Station of Itirapina (ESI), São Paulo, Brazil. Faeces collection and small mammal abundance survey by pitfall trapping were conducted simultaneously during 2000-2002. The maned wolf was selective in the consumption of small mammals. Calomys tener and Oligoryzomys nigripes (Muridae) were consumed less than expected by chance, whereas Clyomys bishopi (Echimyidae) was preyed on more than expected. We did not detect a clear functional response by this canid in the ESI to overall shifts in abundance of small mammals. However, prey switching was detected by an increase in diet diversity in the wet season when small mammals become less available.
Key words. Chrysocyon brachyurus. Prey selection. Prey switching. Small mammals.
INTRODUCTION
Predators may influence prey population dynamics (Craighead and Craighead, 1956; Erlinge et al., 1983) either selecting prey species or consuming higher proportions of a particular size or age class (Brown and Twigg, 1971; Roberts and Wolfe, 1974; Lay, 1974; Fast and Ambrose, 1975; Fulk, 1976; Marti, 1976; Boonstra, 1977; Marti and Hogue, 1979; Jaksic and Yañez, 1979; Longland and Jenkins, 1987; Dickman et al., 1991; Bellocq and Kravetz, 1994; Trejo and Guthmann, 2003). On the other hand, predators may be affected by prey density (Jaksic and Simonetti, 1987). Predator's reaction to fluctuation in prey levels might involve changes in the foraging behaviour. For example, predators might increase the consumption of a particular prey when they are abundant or switch to a different prey type when their abundances decrease (Korpimäki and Norrdahl, 1989; Jaksic et al., 1992; White et al., 1996).
Jaksic (1989 a, b) defines an opportunistic predator as the one that consumes all prey types in similar frequencies as their availability in its hunting site and whose diet composition better fit with prey abundance than with prey size. Conversely, a selective predator consumes one or all prey in different proportions than those present in the field and its diet fits better with prey size than with prey abundance.
The functional response of predators to variation in prey abundance was reported in boreal forests of the Holoartic region (e.g. Korpimäki and Norrdahl, 1989; Dale et al., 1994), in a seasonal forest in Africa (e.g. Ray, 1998), and in subtropical areas (Jaksic et al., 1980, 1992; Silva et al., 1995; Branch et al., 1996) (see Marti, 1987, and Jaksic and Simonetti, 1987 for reviews). Although some authors have emphasized the need for additional studies on patterns of predation in Neotropical region (Jaksic and Yañez, 1979; Jaksic et al., 1981; Meserve et al., 1987; Jaksic et al., 1992), few studies on functional response, selection and prey vulnerability to mammalian carnivorous have been conducted in this region (e.g., Meserve et al., 1987; Iriarte et al., 1989; Corley et al., 1995; Motta-Junior 2000).
The maned wolf Chrysocyon brachyurus is a large canid (20 to 33 Kg for adults) that inhabits mostly savannah-like and grassland habitats. Its distribution comprises central South America, including all over Paraguay, north eastern Argentina, north western Uruguay, extreme south eastern Peru and large parts of central and eastern Bolivia, and central and southern Brazil (Langguth, 1975; Dietz, 1984, 1985; Nowak, 1999). Chrysocyon brachyurus is included in the IUCN Red List as a "lower risk - near threatened" species (IUCN, 2003), whereas in the Brazilian official list it is considered a "vulnerable" species (Machado et al., 2005).
Except in the breeding season, the maned wolf is a solitary animal, being active mainly during the crepuscular-nocturnal period. Its home range extends from 21.7 to 115.0 km2 (Dietz, 1984, 1985; Carvalho and Vasconcellos, 1995). It is omnivorous, relying on fruits (mostly wolf's fruit Solanum lycocarpum) and small vertebrates (mainly small mammals) for most of its nutrition (Dietz, 1984; Motta-Junior et al., 1996; Motta-Junior, 2000; Aragona and Setz, 2001; Bueno et al., 2002; Juarez and Marinho-Filho, 2002; Santos et al., 2003; Bueno and Motta-Junior, 2004; Jácomo et al., 2004). Although several studies about its diet were recently published, only one reported on small mammal selection and functional response (Motta-Junior, 2000).
The goal of this study was to verify if the maned wolf selects small-mammal prey in southeast Brazil. Also, we attempted to test for a functional response and prey switching in relation to the consumption of small mammals. Data reported here were intended to describe patterns of predation by this solitary hunter as well as to bring new insights into the predator-prey relationship in Neotropical region.
MATERIAL AND METHODS
Study area
The study was carried out in the Ecological Station of Itirapina - ESI (2 300 ha, 22 o 15' S; 47 o 49' W), São Paulo State, southeast Brazil. This Ecological Station comprises Cerrado (savannah-like) vegetation and it is one of the last remains of natural savannahs and grasslands in São Paulo State (Parker and Willis, 1997; SEMA, 2000). The physiognomies include grassland (locally named "campo limpo"), grassland savannah ("campo sujo", and "campo cerrado"), savannah ("cerrado sensu stricto") (see Coutinho, 1978) and small patches of marshes and gallery forests. The ESI is surrounded by plantations of Pinus sp., Eucalyptus sp., Citrus sp., and by pastures. According to Köppen (1948), the climate is Cwa with well distinct wet (October to March) and dry (April to September) seasons. A general description of the Cerrado biome can be found in Eiten (1994). The Cerrado is the second most important Brazilian hotspot, largely threatened by the expansion of pastures and agriculture (Ratter et al., 1997).
Faeces collection
Roads and trails were walked and driven on each month from February 2000 to July 2002 in order to search for faecal samples. Faeces were taken to the laboratory to be washed in water through a fine (1 mm) mesh screen (Emmons, 1987; Motta-Junior et al., 1996) or in a washing machine (e.g., Springer and Smith, 1981; Uresk and Sharps, 1986). Afterwards, they were oven dried (50ºC) during 24-48 hours, stored and individually examined.
Prey identification
Prey remains of teeth and bones were identified to the most detailed taxonomic level possible by comparison with a reference collection of the study site. Pairs of mandibles were used to count the minimum number of individual animals consumed (Emmons, 1987). The prey biomass consumption was estimated by counting the number of individuals in faeces and then multiplying this number by the mean body mass of each prey species, obtained at the study site by pitfall trap surveys (Emmons, 1987; Motta-Junior et al., 1996).
Prey availability
Three sets of pitfall traps were installed in three different physiognomies of Cerrado to survey small-mammal abundance ("campo sujo", "campo cerrado" and an ecotone "campo sujo"/forest gallery). Each set consisted of two lines separated by 100 m with four buckets (100 l) each, 15 m apart, connected by a plastic drift-fence (45 cm length and 0.5 cm high). A total of nine sets or 18 lines with 72 buckets (four buckets/line) were checked three days per month, from February 2000 to July 2002, resulting in 6 480 pitfall (bucket)-nights. All captured rodents were identified, sexed, weighed, ear-tagged with numbered monel tags, and then released. Only first captures were considered to obtain the small mammal frequencies in the field.
Boonstra and Krebs (1978), Iriarte et al. (1989), Jaksic (1989 a, b) and Dickman et al. (1991) highlighted the influence of sampling methods on prey capturability and on studies of prey selection. Traps should be as efficient as the predators in capturing different prey species, besides revealing the real prey composition and abundance in the environment. Some authors have suggested that the use of pitfall traps could avoid such potential methodological biases (Boonstra and Krebs, 1978; Korpimäki and Norrdahl 1989; Dickman et al., 1991). These traps are more efficient than cages in capturing smaller and younger individuals, and may invoke a more faithful picture of prey community.
Data analysis
The diet of the maned wolf was evaluated in relation to changes in prey availability along the years, using the Spearman correlation coefficient (r s) (Zar, 1984; Silva et al., 1995). To provide adequate sample sizes for analyses, diet data were pooled in two or three-month periods, however avoiding the combination of different seasons. At least eight faeces per period were collected in the study area (mean ± standard deviation = 24.5 ± 10.5 faeces/period). The food-niche breadth was calculated for each group of months defined above using the standardized Levins measure (B st) (Krebs 1999). Small mammal abundance in the diet was compared to their availability in the environment using G test (Jaksic, 1979, 1989a; Zar, 1984), and employing Bonferroni confidence intervals according to Byers and Steinhorst (1984). In all tests, a statistical significance of 0.05 was considered.
RESULTS
Eleven potential small mammal prey species and 1 223 individuals were captured in ESI during the study period. Calomys tener (83.0%) was the most abundant, followed by Oligoryzomys nigripes (12.7%) and Clyomys bishopi (2.2%). The remaining species comprised only 2.1% of the total individuals captured (Table 1).

The study was based on a total sample of 252 faeces. Clyomys bishopi and C. tener were the most abundant rodents found in the faecal samples (Table 2). Although some other rodent species, such as Oxymycterus delator and Nectomys squamipes, were present in the study area, they were not found in the collected faeces during the study period.

Although the maned wolf consumed more small mammals in the dry season, when they were more abundant in the study area (Tables 1 and 2), we did not detect a functional response by this canid in ESI to shifts in the abundance of the small mammals. The whole assemblage data showed a negative non-significant correlation. (rs = -0.018, p=0.957, Fig. 1). The association between abundance in the field and consumption by the wolves for each prey species also was not significant. The mean Bst was higher in the wet season (0.57 ± 0.14; n=5 periods) than in the dry season (0.37 ± 0.09; n = 6) (Mann-Whitney test, U=3, p=0.0285). Also, there was no significant correlation, neither between Bst and small mammal abundance in the field (rs =-0.020, p=0.959, n= 11) nor between Bst and small mammal consumption by the wolves (rs =0. 209, p=0.537, n= 11) (Fig. 1).
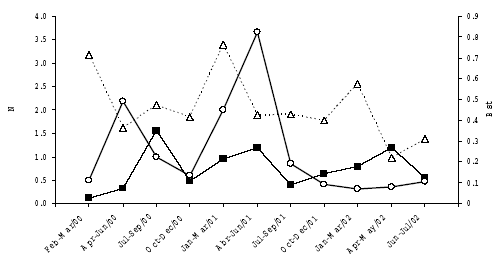
Fig. 1. Number of small mammals in the field (individuals per pitfall line), number of small mammals per faecal sample and Levins standardized food niche breadth (B st), in relation to groups of months
According to the overall analysis, the maned wolf was selective in relation to the consumption of small mammals (G = 245.76; d.f. = 2; p << 0.001). Calomys tener and O. nigripes were consumed less than expected by chance, whereas C. bishopi was preyed on more than expected (Table 3).
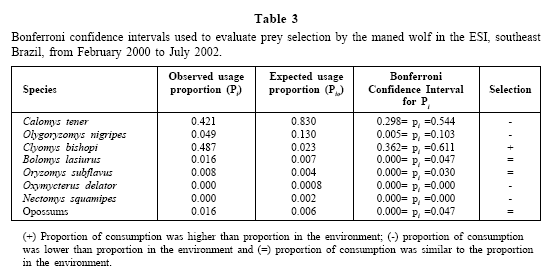
Both in the wet season (G=75.87; d.f.= 1; p <0.001) and in the dry season (G=57,0; d.f.=1; p <0.001), the maned wolf selected small mammal species (Tables 4 and 5). Calomys tener was always consumed in lower proportions, while C. bishopi was invariably consumed more than expected by chance.


DISCUSSION
The diet of the maned wolf in the ESI was described in detail in Bueno et al. (2002). Small mammals represented 22.5% in terms of frequency of occurrence (total of 1 054 occurrences), 45.8% of the number of ingested animals (total of 507 individuals) and 16.2% of the ingested biomass estimated (total of 185,323 g).
The high consumption of small mammals by the maned wolf was reported mainly in the dry season when they are more abundant (Dietz, 1984; Motta-Junior, 2000; Bueno and Motta-Junior, 2004; Jácomo et al., 2004). However, we did not find any functional response in this study. Motta-Junior (2000) also reported this lack of functional response in relation to small mammals by the maned wolf elsewhere. The lack of significant results occurred probably because the maned wolf included many individuals of C. bishopi in its diet during the wet season, despite having reduced its overall consumption of small mammals in this period (Tables 1 and 2). This species comprised almost 50% of all small mammal individuals consumed in these months (Table 2).
However, prey switching -a concept closely associated with functional response- was observed. The maned wolf increased the consumption of fruits in the wet season (Bueno et al., 2002), when small mammals became less available (Table 1). According to White et al. (1996), prey switching enables the predator to survive and reproduce when their primary prey is scarce. When analysing the diet as a whole, and not only the consumption of small mammals, the higher values of Bst for the wet season may be understood by the fact that during these months, the maned wolf more often consumes food items other than small mammals, such as wild fruits (Solanum lycocarpum and Campomanesia glabra, among others) and other small vertebrates (Bueno et al., 2002). The maned wolf tends to show a more specialist diet during dry months, when small mammal abundance is higher and the predation is restricted to less rodent species, mostly C. bishopi. It also included higher proportions of S. lycocarpum fruits in its diet during drier months. Similar results were reported in Jácomo et al. (2004) for the maned wolf in central Brazil. White et al. (1996) and Jaksic et al. (1992) argued that if predators react functionally to fluctuations in densities of small mammals, then diet diversity might increase as mammals decline in abundance. This was reported in our study.
Among the rodents, the results showed strong preference for larger species. Such pattern was observed along the two-year study in the ESI. As a large mammalian predator, the high energetic needs of the maned wolf are fulfilled with the consumption of more profitable prey. The mean weights of the rodents in the environment are in Table 1. Differences in the selection between the two seasons may reflect prey availability.
The largest and most consumed rodent in ESI, C. bishopi, is a semi-fossorial and herbivorous species (Vieira, 1997, Bueno et al., 2004), that inhabits patches of savannah-like vegetation. A possible explanation to its high consumption in ESI may be related to its higher availability in this area. Enviromental conditions such as open vegetation also might contribute to the high rate of predation of this rodent species. It seems to be a vulnerable species above ground, with no display of erratic movements or bipedal leaps during escape (Bueno, 2003; see also discussions on locomotor adaptations to predation escape in Bartholomew and Caswell, 1951; Bartholomew and Cary, 1954; Kotler, 1984, 1985).
The genus Calomys was reported in the diet of the maned wolf by several authors (Motta-Junior et al., 1996; Motta-Junior, 2000; Bueno and Motta-Junior, 2004). Probably, these small sized rodents bring low benefits to such a large predator. On the other hand, they were very abundant, their bad hearing (relative small tympanic bulla, 12.38 ± 2.43 ml) and locomotion capabilities (Bueno, 2003; see Webster and Webster, 1971; Kotler, 1984, 1985 for the positive association between hearing and tympanic bulla size) might make them highly vulnerable to predation by maned wolves.
Morphological and behavioural components of both prey and predators may explain differential predation (Lay, 1974; Dickman et al., 1991; Castro and Jaksic, 1995). Selectivity may involve predator morphological limits in handling prey (Longland and Jenkins, 1987), as well as behavioural aspects of prey species (Jaksic and Marti, 1981; Kotler, 1985; Nashimura and Abe, 1988; Taylor, 1994). Handling time, rates of energy and nutrient intake, risk of injury as well as sensorial capabilities of prey and predator are involved in the process of selection (Endler, 1986). Differential predation may be a result of real selection by predators reflecting prey vulnerability. Future studies dealing with prey vulnerability could bring new insights to the understanding of predator-prey relationship (see Corley et al., 1995).
ACKNOWLEDGEMENTS
We thank to FAPESP and WWF/Ford Brazil for the financial support. R. M. Shimizu made invaluable suggestions on an earlier version of the manuscript. C. A. Grelle, F. M. Jaksic and E. M. Vieira made valuable suggestions for the final version. Darius Tubelis reviewed the English. Cinthia Brasileiro and Sonia Belentani aided in the fieldwork.
LITERATURE CITED
ARAGONA M and EZF SETZ. 2001. Diet of the maned wolf, Chrysocyon brachyurus (Mammalia: Canidae), during wet and dry seasons at Ibitipoca State Park, Brazil. Journal of Zoology, (London) 254:131-136. [ Links ]
BARTHOLOMEW-JR GA and HH CASWELL-JR. 1951. Locomotion in Kangaroo rats and its adaptative significance. Journal of Mammalogy 32:155-159. [ Links ]
BARTHOLOMEW-JR GA and GR CARY. 1954. Locomotion in Pocket mice. Journal of Mammalogy 35(3):386-392. [ Links ]
BELLOCQ MI and FO KRAVETZ. 1994. Feeding strategy and predation of the barn owl (Tyto alba) and the burrowing owl (Speotyto cunicularia) on rodent species, sex, and size, in agrosystems of central Argentina. Ecología Austral 4:29-34. [ Links ]
BOONSTRA R. 1977. Predation on Microtus townsendii populations: impact and vulnerability. Canadian Journal of Zoology 55:1631-1643. [ Links ]
BOONSTRA R and CJ KREBS. 1978. Pitfall trapping of Microtus townsendii. Journal of Mammalogy 59:136-148. [ Links ]
BRANCH LC, M PESSINO, and D VILLARREAL. 1996. Response of pumas to a population decline of the plains vizcacha. Journal of Mammalogy 77(4):1132-1140. [ Links ]
BROWN JC and GI TWIGG. 1971. Mammalian prey of the barn owl (Tyto alba) on Skomer Island, Pembrokeshire. Journal of Zoology, (London) 165:527-530. [ Links ]
BUENO AA, SCS BELENTANI, and JC MOTTA-JUNIOR. 2002. Ecologia alimentar do lobo-guará, Chrysocyon brachyurus (Illiger, 1815) (Mammalia: Canidae), na Estação Ecológica de Itirapina, Estado de São Paulo. Revista Biota Neotropica 2(2):1-9. http://www.biotaneotropica.cria.org.br/v2n2/en/abstract?article. [ Links ]
BUENO AA. 2003. Vulnerabilidade de pequenos mamíferos de áreas abertas a vertebrados predadores na Estação Ecológica de Itirapina, SP. Dissertação, Instituto de Biociências, Universidade de São Paulo, São Paulo, Brasil. [ Links ]
BUENO AA and JC MOTTA JUNIOR. 2004. Feeding ecology of two syntopic canids, the maned wolf (Chrysocyon brachyurus) and the crab-eating fox (Cerdocyon thous), in Southeastern Brazil. Revista Chilena de Historia Natural 77:5-14. [ Links ]
BUENO AA, MJ LAPENTA, MF OLIVERA, and JC MOTTA-JUNIOR. 2004. Association of Clyomys bishopi (Rodentia: Echimyidae) with palm trees and armadillo burrows in southeastern Brazil. Revista de Biología Tropical 52(4):1009-1011. [ Links ]
BYERS CR and RK STEINHORST. 1984. Clarification of a technique for analysis of utilization-availability data. Journal of Wildlife Management 48(3):1050-1053. [ Links ]
CASTRO SA and FM JAKSIC. 1995. Great Horned Owl and Barn Owl prey differentially according to the age/size of a rodent in Northcentral Chile. Journal of Raptor Research 29(4):245-249. [ Links ]
CARVALHO CT and LEM VASCONCELLOS. 1995. Disease, food and reproduction of the maned wolf Chrysocyon brachyurus (Illiger) (Carnivora, Canidae) in southeast Brazil. Revista Brasileira de Zoologia 12:627-640. [ Links ]
CORLEY JC, GJ FERNANDEZ, AF CAPURRO, AJ NOVARO, MC FUNES, and A TRAVAINI. 1995. Selection of cricetine prey by the culpeo fox in Patagonia: a differential prey vulnerability hypothesis. Mammalia 59(3):315-325. [ Links ]
COUTINHO LM. 1978. O conceito de Cerrado. Revista Brasileira de Botânica 1:17-23. [ Links ]
CRAIGHEAD JJ and FC CRAIGHEAD Jr. 1956. Hawks, owls and wildlife. Harrisburg, Stackpole Books. [ Links ]
DALE BW, LG ADAMS and RT BOWYER. 1994. Functional response of wolves preying on barren-ground caribou in a multiple-prey ecosystem. Journal of Animal Ecology 63:644-652. [ Links ]
DICKMAN CR, M PREDAVEC, and AJ LYNAM. 1991. Differential predation of size and classes of mice by the barn owl, Tyto alba. Oikos 62:67-76. [ Links ]
DIETZ JM. 1984. Ecology and social organization of the maned wolf (Chrysocyon brachyurus). Smithsonian Contributions to Zoology 392:1-51. [ Links ]
DIETZ JM. 1985. Chrysocyon brachyurus. Mammalian Species 234:1-4. [ Links ]
EITEN G. 1994. Vegetação do cerrado. Pp. 17-73, in: Cerrado: caracterização, ocupação e perspectives (MN Pinto, ed.). Editora Universidade de Brasília, Brasília, Brasil. [ Links ]
EMMONS LH. 1987. Comparative feeding ecology of felids in a neotropical rainforest. Behavioral Ecology and Sociobiology 20:271-283. [ Links ]
ENDLER JA. 1986. Defense against predators. Pp. 109-134, in: Predator-prey relationships (ME Feder and GV Lauder, eds.). University of Chicago Press, Chicago. [ Links ]
ERLINGE S, G GÖRANSSON, L HANSSON, G HÖGSTEDT, O LIBERG, IN NILSSON, T NILSSON, T VON SCHANTZ, and M SYLVÉN. 1983. Predation as a regulating factor on small rodent populations in southern Sweden. Oikos 40:36-52. [ Links ]
FAST SJ and HW AMBROSE. 1975. Prey preference and hunting habitat selection in the barn owl. American Midland Naturalist 96:503-507. [ Links ]
FULK GW. 1976. Owl predation and rodent mortality: a case study. Mammalia 40:423-427. [ Links ]
IUCN. 2003. Red List of Threatened Species. Gland, Switzerland and Cambridge, UK. [ Links ]
IRIARTE JA, JE JIMENEZ, LC CONTRERAS, and FM JAKSIC. 1989. Small-mammal availability and consumption by the fox, Dusicyon culpaeus, in Central Chilean scrublands. Journal of Mammalogy 70(3):641-645. [ Links ]
JÁCOMO ATA, L SILVEIRA, and JAF DINIZ-FILHO. 2004. Niche separation between the maned wolf (Chrysocyon brachyurus), the crab-eating fox (Dusicyon thous) and the hoary fox (Dusicyon vetulus) in central Brazil. Journal of Zoology (London) 262:99-106. [ Links ]
JAKSIC FM. 1979. Técnicas estadisticas simples para evaluar selectividad dietária en Strigiformes. Medio Ambiente 4:114-118. [ Links ]
JAKSIC FM. 1989 a. What do carnivorous predators cue in on size or abundance of mammalian prey? A crucial test in California, Chile, and Spain. Revista Chilena de Historia Natural 62:237-249. [ Links ]
JAKSIC FM. 1989 b. Opportunism vs. selectivity among carnivorous predators that eat mammalian prey: a statistical test of hypotheses. Oikos 56(3):427-430. [ Links ]
JAKSIC FM and JL YAÑEZ. 1979. The diet of the barn owl in central Chile and its relation to the availability of prey. Auk 96:619-621. [ Links ]
JAKSIC FM, RP SCHLATTER, and JL YAÑEZ. 1980. Feeding ecology of central Chilean foxes Dusicyon culpaeus and Dusicyon griseus. Journal of Mammalogy 61:254-260. [ Links ]
JAKSIC FM, HW GREENE, and JL YAÑEZ. 1981. The guild structure of a community of predatory vertebrates in central Chile. Oecologia (Berlin) 49:21-28. [ Links ]
JAKSIC FM and CD MARTI. 1981. Trophic ecology of Athene owls in Mediterranean-type ecosystems: a comparative analysis. Canadian Journal of Zoology 59:2331-2340. [ Links ]
JAKSIC FM and JA SIMONETTI. 1987. Predator/prey relationships among terrestrial vertebrates: an exhaustive review of studies conducted in Southern South America. Revista Chilena de Historia Natural 60:221-244. [ Links ]
JAKSIC FM, JE JIMÉNEZ, SA CASTRO and P FEINSINGER. 1992. Numerical and functional response of predators to a long-term decline in mammalian prey at a semi-arid Neotropical site. Oecologia 89:90-101. [ Links ]
JUAREZ KM and J MARINHO-FILHO. 2002. Diet, habitat use and home ranges of sympatric canids in central Brazil. Journal of Mammalogy 83(4):925-933. [ Links ]
KOTLER BP. 1984. Risk of predation and the structure of desert rodent communities. Ecology 65(3):689-701. [ Links ]
KOTLER BP. 1985. Owl predation on desert rodents which differ in morphology and behaviour. Journal of Mammalogy 66(4):824-828. [ Links ]
KÖPPEN W. 1948. Climatologia. Ed. Fondo Cultura Economica, Mexico. [ Links ]
KORPIMÄKI E and K NORRDAHL. 1989. Predation of Tengmalm´s owls: numerical responses, functional responses and dampening impact on population fluctuations of microtines. Oikos 54:154-164. [ Links ]
KREBS CJ. 1999. Ecological methodology. 2 nd edition. Benjamin-Cumming, Menlo Park, California. [ Links ]
LANGGUTH A. 1975. Ecology and evolution in the South American canids. Pp. 192-206, in: The wild canids: their systematics, behavioral ecology and evolution (MW Fox, ed.). Van Nostrand Reinhold Co., New York. [ Links ]
LAY DM. 1974. Differential predation on gerbils (Meriones) by the little owl, Athene brahma. Journal of Mammalogy 55:608-614. [ Links ]
LONGLAND WS and SH JENKINS. 1987. Sex and age affect vulnerability of desert rodents to owl predation. Journal of Mammalogy 68(4):746-754. [ Links ]
Machado AB, CS Martins, and GM Drummond. 2005. Lista da Fauna Brasileira Ameaçada de Extinção. Fundação Biodiversitas, Belo Horizonte. [ Links ]
MARTI CD. 1976. A review of prey selection by the long-eared owl. Condor 78:331-336. [ Links ]
MARTI CD and JG HOGUE. 1979. Selection of prey size in screech owls. Auk 96:319-327. [ Links ]
MARTI CD. 1987. Predator-Prey interactions: a selective review of North American research results. Revista Chilena de Historia Natural 60:203-219. [ Links ]
MESERVE PL, EJ SHADRICK, and DA KELT. 1987. Diets and selectivity of two Chilean predators in the northern semi-arid zone. Revista Chilena de Historia Natural 60(1):93-99. [ Links ]
MOTTA-JUNIOR JC. 2000. Variação temporal e seleção de presas na dieta do lobo-guará, Chrysocyon brachyurus (Mammalia: Canidae), na Estação Ecológica de Jataí, Luiz Antônio, SP. Pp. 331-346, in: Estudos integrados em ecossistemas. Estação ecológica de Jataí (JE Santos. and JSR Pires, eds.). Vol. I. Rima Editora, São Carlos. [ Links ]
MOTTA-JUNIOR JC, SA TALAMONI, JA LOMBARDI, and K SIMOKOMAKI. 1996. Diet of the maned wolf, Chrysocyon brachyurus, in central Brazil. Journal of Zoology, (London) 240:277-284. [ Links ]
NASHIMURA K and MT ABE. 1988. Prey susceptibilities, prey utilization and variable attack efficiencies of Ural owls. Oecologia 77:414-422. [ Links ]
NOWAK RM. 1999. Walker´s Mammals of the World. Vol. II. 6 th edition. The John Hopkins University Press, Baltimore and London. [ Links ]
PARKER TA and EO WILLIS. 1997. Notes of three tiny grassland flycatchers, with comments on the disappearance of South American fire-diversified savannas. Ornithological. Monographs 48:549-556. [ Links ]
RAY JC. 1998. Temporal variation of predation on rodents and shrews by small African forest carnivores. Journal of Zoology (London) 244:363-370 [ Links ]
RATTER JA, JF RIBEIRO, and S BRIDGWATER. 1997. The Brazilian Cerrado vegetation and its threats to its biodiversity. Annals of Botany 80:223-230. [ Links ]
ROBERTS MW and JL WOLFE. 1974. Social influences on susceptibility to predation in cotton rats. Journal of Mammalogy 55:869-872. [ Links ]
SEMA. 2000. Secretaria de Estado do Meio Ambiente 2000. Atlas das unidades de conservação ambiental do Estado de São Paulo. SEMA. São Paulo. [ Links ]
SILVA SI, I LAZO, E SILVA-ARANGUIZ, FM JAKSIC, PL MESERVE and JR GUTIÉRREZ. 1995. Numerical and functional response of Burrowing Owls to long-term mammal fluctuations in Chile. Journal of Raptor Research 29(4):250-255. [ Links ]
SANTOS EF, EZF SETZ, and N GOBBI. 2003. Diet of the maned wolf (Chrysocyon brachyurus) and its role in seed dispersal on a cattle ranch in Brazil. Journal of Zoology, (London). 260:203-208. [ Links ]
SPRINGER JT and JS SMITH. 1981. Summer food habits of coyotes in Central Wyoming. Great Basin Naturalist 41(4):449-456. [ Links ]
TAYLOR I. 1994. Barn owls. Predator-prey relationship and conservation. Cambridge University Press, Cambridge, UK. [ Links ]
TREJO A and GUTHMANN N. 2003. Owl selection on size and sex classes of rodents: activity and microhabitat use of prey. Journal of Mammalogy 84(2):652-658. [ Links ]
URESK D and JC SHARPS. 1986. Denning habitat and diet of the swift fox in Western South Dakota. Great Basin Naturalist 46(2):249-253. [ Links ]
VIEIRA MV. 1997. Dynamics of a rodent assemblage in a cerrado of southeast Brazil. Revista Brasileira de Biologia 57(1):99-107. [ Links ]
WEBSTER DB and M WEBSTER. 1971. Adaptative value of hearing and vision in kangaroo rat predator avoidance. Brain, Behaviour and Evolution 4:310-322. [ Links ]
WHITE PJ, CA VANDERBILT-WHITE and K RALLS. 1996. Functional and numerical response of kit fox to a short-term decline in mammalian prey. Journal of Mammalogy 77(2):370-376. [ Links ]
ZAR JH. 1984. Biostatistical analysis. 2 nd edition. Prentice Hall, Englewood Cliffs. [ Links ]
Recibido 24 junio 2005.Aceptación final 3 diciembre 2005.

