










Servicios Personalizados
Revista

Articulo
 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. v.13 n.1 Mendoza ene./jun. 2006
Dinâmica populacional de marsupiais e roedores no Parque Municipal da Lagoa do Peri, Ilha de Santa Catarina, sul do Brasil
Maurício E. Graipel¹, Jorge J. Cherem¹, Emygdio L. A. Monteiro-Filho² e Luiz Glock³
1 Departamento de Ecologia e Zoologia, Centro de Ciências Biológicas, Universidade Federal de Santa Catarina, Campus Universitário, Trindade; 88.040-970; Florianópolis, SC - Brasil. <graipel@ccb.ufsc.br>. 2 Departamento de Zoologia, Universidade Federal do Paraná, Cx Postal 19020, 81531-990, Curitiba, PR, Brasil. 3 Programa de Pós-Graduação em Biociências, Pontifícia Universidade Católica do Rio Grande do Sul, Brasil.
ABSTRACT: Population dynamics of marsupials and rodents in Parque Municipal da Lagoa do Peri, Santa Catarina Island, southern Brazil. A two-year mark-recapture study of small mammals was conducted in an area of Atlantic Forest hillside in the state of Santa Catarina, Brazil. Four species of marsupial and seven species of rodent were captured during 12 132 trap-nights. Aspects of population dynamics and natural history are discussed for each common species. The sex ratio was biased toward males for all species. Oryzomys russatus, Didelphis aurita and Akodon montensis had the highest population densities, the least amount of variation in population density and an inverse relationship between survivorship rates and population densities. Micoureus paraguayanus, Nectomys squamipes and O. angouya, registered the higher population densities after continual survivorship rates. The recruitment rates were low for most species (< 50%), with some, M. paraguayanus, Lutreolina crassicaudata, Oligoryzomys nigripes, O. angouya and Oxymycterus aff. judex, disappearing occasionally from the study area. The survivorship rates were high for most species (> 50%). Population densities of D. aurita, O. nigripes and O. aff. judex increased in cold and dry periods. The marsupials reproduced seasonally from late winter until the end of the summer, whereas rodents reproduced throughout the year. No species were long-lived, with average life-span < six months.
Key words. Atlantic Forest. Biology. Ecology. Reproduction. Small mammals.
INTRODUÇÃO
A falta de conhecimento acerca da mastofauna brasileira, aliada à drástica redução da Mata Atlântica, uma das florestas tropicais mais ameaçadas do mundo, vem despertando o interesse dos pesquisadores nos últimos anos, promovendo um aumento significativo das pesquisas nesta região (Fonseca e Kierulff, 1989; Stallings, 1989; Olmos, 1991; Bergallo, 1994, Gentile et al., 2000, entre outros).
No estado de Santa Catarina, a maior parte dos estudos sobre mamíferos restringe-se a listas taxonômicas, eventualmente com comentários relacionados a capturas (Azevedo et al., 1982; Sipinski e Reis, 1995; Cimardi, 1996; Cherem e Perez, 1996; Graipel et al., 1997; Avila-Pires, 1999; Wallauer et al., 2000; Quadros e Cáceres, 2001; Cherem et al., 2004). Parâmetros populacionais e comunitários, quando considerados, são pouco abrangentes (Cherem et al., 1996; Voltolini, 1998).
Na Ilha de Santa Catarina, o panorama não é diferente. Dos poucos estudos publicados sobre os mamíferos da Ilha, apenas dois (Graipel et al., 2001; Graipel et al., 2003) apresentam dados sobre sua ecologia, ainda que superficialmente. Além disso, o processo de degradação dos hábitats, aliado à caça, tem levado uma parcela significativa da fauna de mamíferos à extinção local (Caruso, 1990; Olimpio, 1995). Atualmente, suas florestas estão reduzidas a menos de 10% do território insular (Caruso, 1990) e mais de 50% das espécies de médio porte e todas as de grande porte foram extintas (Olimpio, 1995; Graipel et al., 2001). Além das dificuldades para recomposição natural da fauna autóctone em função de barreiras próprias de uma ilha, ainda que continental, há outros impactos como a introdução de fauna exótica (Graipel et al., 2001), que vem trazendo prejuízos consideráveis. A soma desses fatores, certamente, leva ao declínio da biodiversidade (Pianka, 1983), declínio este que deve estar ocorrendo na Ilha de Santa Catarina. A despeito disto, aspectos relacionados à dinâmica populacional de qualquer uma das espécies de mamíferos são desconhecidos.
Em vista disso, foi realizado um estudo de uma comunidade de pequenos mamíferos não voadores em uma área de Mata Atlântica no sul da Ilha de Santa Catarina, durante dois anos, onde foram abordados aspectos da dinâmica populacional e biologia das principais espécies capturadas.
MATERIAL E MÉTODOS
Área de estudo
O estudo foi desenvolvido em uma área de Mata Atlântica (floresta ombrófila densa submontana, de acordo com Klein, 1990), no Parque Municipal da Lagoa do Peri (PMLP; 27º43'S e 48º32'W; Fig. 1), sul da Ilha de Santa Catarina, estado de Santa Catarina, sul do Brasil. Este parque possui aproximadamente 1 500 ha de área florestal (CECCA, 1997).
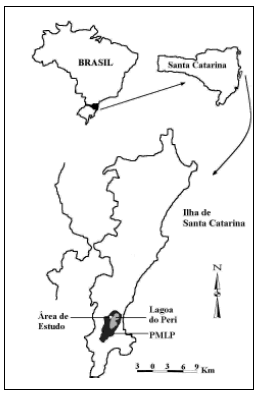
Fig. 1. Localização da área de estudo no Parque Municipal da Lagoa do Peri, Ilha de Santa Catarina, sul do Brasil.
Uma grade de armadilhas foi instalada em uma mata secundária no início da encosta, entre 5 e 60 metros de altitude, na parte oeste da Lagoa do Peri (Fig. 1). Nesta área há um grande número de afloramentos de rochas e o estrato arbustivo forma densos agrupamentos, principalmente de lianas. Esta zona da encosta é caracterizada por uma grande heterogeneidade e um grande número de árvores altas, entre 20 e 30 metros de altura, formando uma cobertura contínua e densa (Klein, 1990).
O clima é mesotérmico úmido, sem estação seca definida, com chuvas distribuídas ao longo de todo o ano e verões quentes. As médias anuais de pluviosidade e temperatura giram em torno de 1 384 mm e 20.5° C, respectivamente. A umidade relativa é alta, com média anual de aproximadamente 82%. Os ventos mais freqüentes são o Sul, ocorrendo principalmente no outono e no inverno e acarretando baixas na temperatura, e o Nordeste, ocorrendo principalmente na primavera e no verão (IPUF, 1978).
Durante o período de estudo, a temperatura média mensal foi de 20.3ºC, variando de 16.1º C em junho de 1998 e julho de 1999 a 24.7º C em março de 1999. A precipitação mensal acumulada foi de 146.3 mm, variando de 21.2 mm, em agosto de 1998, a 319.4 mm, em fevereiro de 1999 (Fig. 2).
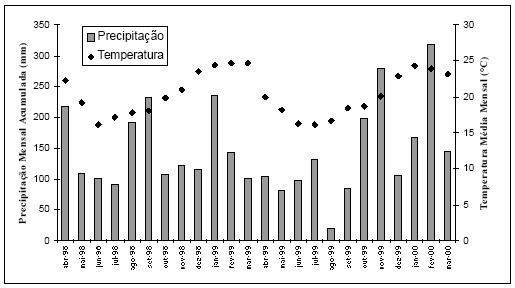
Fig. 2. Variação da precipitação e temperatura no sul da Ilha de Santa Catarina, entre abril de 1998 e março de 2000.
As médias de temperatura e precipitação por estação nos dois anos foram, respectivamente, de 18.6º C e 118.6 mm no outono, 17.4º C e 125.5 mm no inverno, 21º C e 155.2 mm na primavera e 24.2º C e 185.8 mm no verão (Fig. 2) (dados da Estação Meteorológica de Superfície do Destacamento de Proteção ao Vôo de Florianópolis, localizada a 27 o 40'S e 48 o 33'W).
Métodos de campo
De abril de 1998 a março de 2000 foram realizadas amostragens mensais com duração de quatro dias consecutivos cada. Foi utilizada uma grade de 1.26 ha com armadilhas distribuídas em 11 linhas. A primeira e a última linhas possuíam cinco estações, e as demais sete. Cada estação distava 15 metros das outras. Na primeira e na última linhas da grade e na primeira estação das linhas intermediárias foi armada apenas uma armadilha no solo em cada estação. Nas demais estações foram colocadas uma armadilha no solo e outra no sub-bosque. No solo foram utilizadas 20 armadilhas de arame do tipo Young de contenção viva de tamanho médio (450 x 210 x 170 mm, a 30 metros umas das outras) e 53 pequenas (260 x 190 x 90 mm). No sub-bosque, nos primeiros cinco meses de estudo, foram utilizadas 50 armadilhas pequenas amarradas em galhos, lianas ou plataformas de madeira e, posteriormente, 54 pequenas, utilizando-se três métodos distintos (ver Graipel et al., 2003). O esforço total de captura foi de 12 132 armadilhas-noite. Uma compilação detalhada dos dados de captura é apresentada por Graipel et al. (2003). Foram considerados pequenos mamíferos, roedores e marsupiais com peso médio, quando adultos, menor que 1000 g, seguindo Fonseca e Robinson (1990).
Como iscas, foram utilizados pedaços de banana untados com pasta de amendoim. As iscas eram trocadas quando necessário. As armadilhas foram armadas ao entardecer do primeiro dia e revisadas ao amanhecer dos quatro dias seguintes.
Os animais capturados foram pesados, medidos quanto ao comprimento e largura máximos da cabeça (Cáceres e Monteiro-Filho, 1999), averiguados quanto ao sexo, marcados e soltos, anotando-se sua rota de escape (se pelo chão ou subindo em árvores). Os roedores foram marcados por corte de falanges distais (Fullagar e Jewell, 1965) e os marsupiais por este método ou por uma combinação de furos nas orelhas (Monteiro-Filho e Abe, 1999).
Coletas de roedores e marsupiais realizadas anteriormente na Ilha de Santa Catarina permitiram a identificação prévia por especialistas da maioria das espécies capturadas no PMLP. A coleta de pequenos mamíferos na área de estudo foi realizada apenas quando se tratava do primeiro registro para a Ilha de Santa Catarina ou no caso de indivíduos encontrados mortos nas armadilhas. O material coletado foi depositado na Coleção Científica de Mamíferos da Universidade Federal de Santa Catarina.
Métodos de análise
Para verificar o uso do estrato arbóreo pelas espécies, testou-se a diferença entre o número de capturas (primeiras capturas e recapturas) no solo e sub-bosque, utilizando-se o teste de qui-quadrado (χ ²) para proporções esperadas desiguais (Programa Bioestat 2.0; Ayres et al., 2000), uma vez que o número de armadilhas-noite no sub-bosque (n = 5 124; 42.2%) foi inferior àquele no solo (n = 7 008; 57.8%), e as diferenças entre o número de fugas pelo solo e pelas árvores para cada espécie, utilizando-se o teste de qui-quadrado (χ ²) para proporções esperadas iguais (Programa Bioestat 2.0; Ayres et al., 2000).
A estrutura etária das populações ao longo do tempo foi analisada através da determinação de categorias etárias. Para os marsupiais, estas categorias foram baseadas na erupção dos molares superiores de animais independentes. Indivíduos com apenas o primeiro molar superior eclodido foram considerados infantes (categoria M1), com os dois primeiros, jovens (M2), com os três primeiros, subadultos (M3), e com dentição definitiva, adultos (M4). Para os roedores, foram considerados jovens os machos menores que o menor macho com escrotos desenvolvidos e as fêmeas menores que a menor fêmea com a vulva perfurada, os demais foram considerados adultos, assim como proposto por Bergallo (1995). A estrutura sexual das populações foi analisada através da verificação da existência de diferença significativa na proporção sexual das primeiras capturas de animais independentes e de filhotes no marsúpio, separadamente, através de distribuição binomial (Programa Bioestat 2.0; Ayres et al., 2000).
A flutuação das populações foi verificada através da variação da densidade populacional ao longo do tempo, estimada dividindo-se o tamanho populacional em cada período de amostragem pela área efetiva de amostragem (AEA). O tamanho populacional foi considerado como sendo o número mínimo de animais conhecidos vivos (MNKA; Krebs, 1966), e a estimativa da faixa externa à grade foi calculada através do método de Dice (Fernandez, 1995). A área de atividade de cada espécie, para o cálculo das AEA's, foi obtido através do polígono convexo mínimo (Jennrich e Turner, 1969), utilizando-se o programa Biotas 1.0 Alpha. Foram considerados os indivíduos com pelo menos três capturas não lineares. Indivíduos com área de atividade que incluíam a borda da grade não foram considerados na análise.
Os fatores sobrevivência e recrutamento foram estimados objetivando-se verificar a existência de influência destes sobre a flutuação da densidade populacional de cada espécie. A taxa de sobrevivência mensal foi calculada dividindo-se o MNKAi recapturados após a amostra "i" pelo MNKAi menos o número de animais mortos acidentalmente ou retirados da população. As estimativas de sobrevivência foram padronizadas para um intervalo de 30 dias, usando-se um modelo exponencial negativo (Fernandez, 1995). O recrutamento foi estimado com base nos indivíduos capturados pela primeira vez em cada período, independentemente de terem sido incorporados à população por nascimento ou imigração, dividindo-se os indivíduos capturados sem marcas na amostra "i" pelo total de indivíduos capturados na amostra "i", marcados e sem marcas.
Testou-se a existência de correlação entre densidade populacional de cada espécie de pequeno mamífero e as taxas de sobrevivência e recrutamento dos mesmos, com a temperatura e precipitação, utilizando-se a correlação de Spearman (Programa Bioestat 2.0; Ayres et al., 2000). Foram utilizados dois períodos de amostragem anteriores à amostragem "i" para taxa de sobrevivência e recrutamento, e quatro períodos para temperatura e precipitação, pois o efeito destes fatores nem sempre é imediato e geralmente é indireto (Bergallo e Magnusson, 1999).
O período reprodutivo dos roedores foi determinado pela presença de fêmeas lactantes, prenhes ou com a vulva perfurada e através de machos com escrotos desenvolvidos. O período reprodutivo dos marsupiais foi determinado pela presença de fêmeas lactantes ou com filhotes. Para D. aurita, a relação entre o número de filhotes por ninhada, obtidos neste estudo e na literatura, e a latitude foi verificada através da correlação de Spearman (Programa Bioestat 2.0; Ayres et al., 2000). O número médio de filhotes de D. aurita por ninhada registrados em oito estudos realizados ao longo da Mata Atlântica (Davis, 1947 - 7.2; Fonseca e Kierulff, 1989 - 8.6; Cerqueira et al., 1993 - 6.8; Bergallo, 1994 - 7.3; Gentile et al., 2000 - 7.3; Cherem et al., 1996 - 6.5; Cáceres e Monteiro-Filho, 1997 - 8.1; Graipel e Santos-Filho, 2006 - 8.1) incluindo o número médio de filhotes registrados no PMLP foram comparados usando-se regressão linear simples com a latitude destes mesmos estudos. Sugere-se que o número médio de filhotes (variável dependente) tende a ser maior em maiores latitudes (variável independente) uma vez que condições climáticas limitam a extensão do período reprodutivo.
Para avaliar a residência das espécies na área de estudo, o tempo de permanência de machos e fêmeas de cada espécie, presentes em pelo menos dois períodos, foi testado através do teste "U" de Mann-Whitney (Programa Bioestat, 2.0; Ayres et al., 2000).
Quando relevante, foram considerados os dados agrupados por estações do ano: outono = abril, maio e junho; inverno = julho, agosto e setembro; primavera = outubro, novembro e dezembro; e verão = janeiro, fevereiro e março.
RESULTADOS
Ao final do estudo, 85 marsupiais e 287 roedores foram capturados 421 e 970 vezes, respectivamente. O sucesso de captura foi de 11.5%. Onze espécies de pequenos mamíferos foram capturadas, sendo quatro marsupiais (Didelphimorphia, Didelphidae), Chironectes minimus (Zimmermann, 1780), Didelphis aurita Wied-Neuwied, 1826, Lutreolina crassicaudata (Desmarest, 1804) e Micoureus paraguayanus (Tate, 1931), e sete roedores (Rodentia, Muridae), Akodon montensis Thomas, 1913, Juliomys sp., Nectomys squamipes (Brants, 1827), Oligoryzomys nigripes (Olfers, 1818), Oryzomys angouya (G. Fischer, 1814), Oryzomys russatus (Wagner, 1848) e Oxymycterus aff. judex. Além dessas, dois exemplares de Phyllomys medius (Thomas, 1909) foram encontrados mortos, possivelmente por predadores, próximos à grade.
Espécies capturadas
Chironectes minimus
(gambá-d'água, cuica-d'água)
Apenas um macho (M3) foi capturado em março de 2000 à margem de um córrego.
Didelphis aurita
(gambá)
Foram capturados 56 indivíduos 285 vezes. Capturas no sub-bosque representaram apenas 0.7 % (n = 2, um adulto e um infante) do total de capturas da espécie (χ²285 = 203.75; gl = 1; p < 0.001) e apenas 1.4 % (n = 4, três infantes e um adulto) dos indivíduos subiram em árvores após serem soltos (χ²285 = 269.22; gl = 1; p < 0.001).
Animais adultos foram capturados em todos os períodos de estudo; subadultos, de julho a outubro de 1998 e de julho a dezembro de 1999; jovens de maio a julho 1998, em julho de 1999 e março de 2000; e infantes apenas em abril e dezembro de 1999. Jovens e infantes foram capturados do fim de um período reprodutivo ao início do período seguinte. A proporção sexual foi significativamente favorável aos machos (1.67: 1.00; p < 0.05). Nos meses de julho e agosto de 1998 e 1999, que coincidem com o início do período reprodutivo, entraram na população 12 machos e cinco fêmeas, dos quais nove machos eram subadultos e dois eram adultos, e quatro fêmeas subadultas e uma adulta.
A densidade populacional de D. aurita foi a terceira maior no PMLP, 2.4 ind/ha, com uma das menores variações ao longo do tempo (Tabela 1) e picos no inverno (Fig. 3A), coincidindo com o início do período reprodutivo. Esta espécie registrou a segunda maior taxa de sobrevivência e a menor variação ao longo do tempo (Tabela 1; Fig. 3A). A densidade foi negativa e significativamente correlacionada com a taxa de sobrevivência do mês respectivo (Tabela 2), indicando uma diminuição da taxa de sobrevivência com o aumento da densidade. A taxa de recrutamento foi uma das mais elevadas, apresentando variação intermediária em relação às outras espécies ao longo do tempo (Tabela 1; Fig. 3A). Registrou-se, também, correlação significativamente negativa entre a densidade e a temperatura média mensal do mês respectivo e com um e dois meses de atraso (Tabela 2), indicando um aumento da densidade nos meses de inverno.

Fig. 3. Variação mensal da densidade populacional, taxas de sobrevivência e recrutamento para Didelphis aurita, Micoureus paraguayanus, Akodon montensis, Nectomys squamipes, Oligoryzomys nigripes, Oryzomys russatus, Oryzomys angouya e Oxymycterus aff. judex no período de maio de 1998 a fevereiro de 2000 no Parque Municipal da Lagoa do Peri, Ilha de Santa Catarina, sul do Brasil.
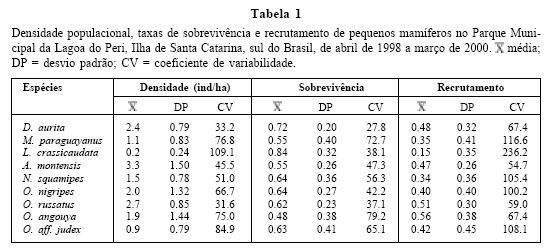
Tabela 2
Análise de correlações entre a densidade populacional de espécies de pequenos mamíferos e os fatores taxas de sobrevivência, taxa de recrutamento, temperatura e precipitação do mês correspondente e de dois a quatro meses antes no Parque Municipal da Lagoa do Peri, Ilha de Santa Catarina, sul do Brasil, de abril de 1998 a março de 2000. rs = Correlação de Spearman; p = probabilidade; n = número de pares; N.S. = não significativo; ░ = correlação negativa; ▒ = correlação positiva.
O primeiro período reprodutivo ocorreu de agosto de 1998 a fevereiro de 1999 e uma fêmea lactante foi capturada em abril de 1999. O segundo período foi de agosto a dezembro de 1999. Em aproximadamente 90% das vezes (n = 8), o primeiro registro de filhotes n o marsúpio foi feito para fêmeas subadultas que nasceram na temporada reprodutiva anterior. Foram registradas, no máximo, duas ninhadas por período reprodutivo no PMLP. Uma fêmea presente na área de estudo durante 14 períodos de amostragem, e capturada em 13, nunca apresentou indícios de reprodução, sendo capturada subadulta no início do primeiro período reprodutivo e adulta no período reprodutivo do ano seguinte.
As ninhadas (n = 15) apresentaram sete filhotes em média (DP = 1,60), com um mínimo de quatro e máximo de nove filhotes. A regressão entre o número médio de filhotes por ninhada com a latitude não foi significativa (n = 8; rs = -0.31; p = 0.450).
A proporção sexual de filhotes no PMLP, em 13 ninhadas, foi exatamente de 1:1. A mortalidade de filhotes no PMLP ocorreu ao longo de todo o período de amamentação, representando 8,1 % do total de filhotes. A média do comprimento total dos filhotes em cada ninhada (n = 4) variou de 27 mm a 52 mm, para as primeiras ninhadas no PMLP, no fim de agosto dos dois períodos reprodutivos.
O tempo médio de permanência de D. aurita no PMLP foi de 148,1 dias, sem se verificar diferença significativa entre os sexos. O percentual de fêmeas residentes também foi superior ao dos machos. Uma fêmea M3 obteve tempo máximo de residência de 631 dias (Tabela 3).
Tabela 3
Tempo médio de permanência de roedores e marsupiais no Parque Municipal da Lagoa do Peri, Ilha de Santa Catarina, Sul do Brasil de abril de 1998 a março de 2000. ![]() média de permanência em dias; DP = desvio padrão; N = Número de indivíduos residentes; Tmáx. = Tempo máximo de permanência; Z(U) = valor de Z para o teste "U" de Mann Whitney; p = probabilidade de erro tipo I; %M = percentual de machos residentes; % F = percentual de fêmeas residentes.
média de permanência em dias; DP = desvio padrão; N = Número de indivíduos residentes; Tmáx. = Tempo máximo de permanência; Z(U) = valor de Z para o teste "U" de Mann Whitney; p = probabilidade de erro tipo I; %M = percentual de machos residentes; % F = percentual de fêmeas residentes.
Lutreolina crassicaudata
(cuica, cuíca-de-cauda-grossa)
Foram capturados sete indivíduos 46 vezes. Todas as capturas ocorreram no solo. Um indivíduo foi morto por cachorro doméstico e outros dois possuíam ferimentos infestados por larvas de moscas e encontravam-se bastante debilitados.
Adultos não foram capturados nos meses de julho, setembro, novembro e dezembro dos dois anos de estudo. Subadultos foram capturados apenas nos meses de março e agosto a outubro de 1998 e março de 1999. A proporção sexual não apresentou desvio do esperado 1:1 (2.50: 1.00; p > 0.05).
Esta foi a espécie com a menor densidade populacional, 0,2 ind/ha, e a que apresentou a maior variação ao longo do estudo (Tabela 1). Nenhuma captura foi registrada entre abril e julho de 1998 e de setembro de 1999 a fevereiro de 2000. Ocorreram picos de densidade no fim do inverno e no fim do verão, sem um padrão definido. Foi a espécie com a maior taxa de sobrevivência e uma das menores variações ao longo do tempo e, por outro lado, teve a menor taxa de recrutamento com a maior variação para o PMLP (Tabela 1).
Apenas na primeira temporada reprodutiva capturou-se uma fêmea com filhotes. Esta apresentava o marsúpio vazio em outubro de 1998, sem evidência de reprodução; não foi capturada em novembro e dezembro de 1998; estava lactante, com os mamilos e marsúpio bem desenvolvidos em 19/01/1999, mas sem filhotes; e, em 25/02/1999, possuía seis filhotes no marsúpio (5 machos e 1 fêmea, com 32 mm de comprimento em média). Em 24/03/2000, os filhotes possuíam em média 95 mm e um filhote macho havia desaparecido do marsúpio. Em 11/04/2000, a fêmea não carregava nenhum filhote. O tempo médio de permanência foi de 87.4 dias, sem diferença significativa entre sexos. O tempo máximo de permanência foi de uma fêmea, 233 dias, enquanto o percentual de machos residentes foi superior ao das fêmeas (Tabela 3).
Micoureus paraguayanus
(catita, guaiquica)
Foram capturados 21 indivíduos 89 vezes. A maior parte das capturas (n = 74; 83.1 %) ocorreu no sub-bosque, sendo registrada diferença significativa em relação às capturas no solo (χ²89 = 60.169; gl = 1; p < 0,001). Quanto às fugas, 50.9 % (n = 45) ocorreram pelas árvores, sem que se registrasse diferença significativa em relação às fugas pelo solo (χ²89 = 0.011; gl = 1; p > 0.05).
A flutuação da população por categoria de idade pareceu responder à sazonalidade na reprodução. No PMLP, adultos foram capturados de março a dezembro, subadultos de janeiro a março e jovens em janeiro de 1999 e fevereiro de 2000. Infantes nunca foram capturados. A proporção sexual foi significativamente favorável aos machos (6.00: 1.00; p < 0.001).
A densidade média foi uma das menores registradas, 1.1 ind/ha, com uma das maiores variações ao longo do tempo entre as espécies capturadas no PMLP (Tabela 1), aumentando no início do verão e com pico de densidade no outono (Fig. 3B). A taxa de sobrevivência média apresentou valores intermediários em relação às demais espécies, com a segunda maior variação ao longo do tempo (Tabela 1; Fig. 3B). A densidade apresentou correlação com a taxa de sobrevivência do mês respectivo e com um mês de atraso (Tabela 2). A taxa de recrutamento ficou entre as menores, apresentando uma grande variação ao longo do tempo (Tabela 1; Fig. 3B). A densidade correlacionou positivamente com a temperatura de dois, três e quatro meses de atraso (Tabela 2).
Nenhuma fêmea foi capturada com filhotes ou com indícios de reprodução. Verificou-se um padrão sincronizado de envelhecimento da população no PMLP. As fêmeas estiveram ausentes da área de estudo entre agosto e dezembro. A entrada de fêmeas na população aconteceu entre janeiro e abril de 1999, sendo duas jovens e uma subadulta.
O tempo médio de permanência foi de 82.1 dias, sem diferença significativa entre sexos. Um maior percentual de fêmeas foi considerado residente. Contudo, o tempo máximo de residência foi de um macho, 263 dias (Tabela 3), o qual permaneceu ausente da grade por oito meses.
Akodon montensis
(rato-do-mato)
Akodon montensis foi a segunda espécie mais capturada. No total, 73 indivíduos foram capturados 225 vezes. Todas as capturas ocorreram no solo.
Adultos (com comprimento de cabeça para machos ≥30 mm e fêmeas ≥31 mm) foram capturados ao longo dos 24 meses, enquanto jovens, apenas em maio, junho, setembro a dezembro de 1998 e abril a junho de 1999. A proporção sexual foi significativamente favorável aos machos (2.17: 1.00; p < 0.001). A. montensis apresentou a maior densidade média no PMLP (3.3 ind/ha; Tabela 1) e uma das menores flutuações ao longo do tempo (Tabela 1), sem um padrão definido de distribuição ao longo do tempo (Fig. 3C). As taxas de sobrevivência e de recrutamento apresentaram valores intermediários em relação às outras espécies, com variação intermediária ao longo do tempo para a taxa de sobrevivência e o menor coeficiente de variabilidade entre todas as espécies capturadas para a segunda (Tabela 1; Fig. 3C). Não ocorreu correlação significativa entre densidade populacional e taxa de sobrevivência, recrutamento, temperatura ou precipitação (Tabela 2).
Fêmeas reprodutivamente ativas foram capturadas ao longo de todo o estudo, exceto em junho de 1998, abril e maio de 1999. Machos com escrotos desenvolvidos foram capturados em todos os meses. Foram observadas duas ninhadas, uma com três e outra com cinco filhotes.
O tempo médio de permanência de A. montensis no PMLP foi de 87.2 dias, sem diferença significativa entre sexos, apesar de o percentual de fêmeas residentes ter sido superior ao de machos. O tempo máximo de permanência foi de uma fêmea, 233 dias (Tabela 3).
Nectomys squamipes
(rato-d'água)
Foram capturados 31 indivíduos 199 vezes. Todas as capturas no PMLP aconteceram no solo e 98.5 % a menos de 15 m de distância da água.
Adultos (com comprimento de cabeça para machos ≥45 mm e fêmeas ≥47 mm) foram capturados em todos os meses do ano. Jovens foram capturados de setembro a novembro de 1998 e 1999 e em fevereiro e março de 1999 e 2000. A proporção sexual foi significativamente favorável aos machos (2.44: 1.00; p < 0.01). A densidade média foi de 1.5 ind/ha, aumentando durante os meses mais quentes, podendo ser considerada intermediária em relação às demais espécies, assim como a flutuação ao longo do tempo (Tabela 1; Fig. 3D). A espécie apresentou uma das maiores taxas de sobrevivência com flutuação intermediária ao longo do tempo (Tabela 1; Fig. 3D). A densidade foi significativamente correlacionada com a taxa de sobrevivência com um e dois meses de atraso, e com a temperatura para o mês respectivo e com um mês de atraso (Tabela 2), provocando um aumento da população nos meses mais quentes do ano, com picos populacionais precedidos de períodos de alta na taxa de sobrevivência. A taxa de recrutamento foi uma das menores, com variação intermediária ao longo do tempo em relação às demais espécies (Tabela 1; Fig. 3D).
Fêmeas reprodutivamente ativas foram capturadas ao longo de todo o estudo, com exceção de junho e julho de 1998 e 1999. Nenhum macho reprodutivamente ativo foi registrado entre maio e agosto nas duas temporadas, com exceção de um indivíduo capturado em julho de 1998. Registrou-se uma fêmea lactante em fevereiro e recapturada prenhe em maio de 1999, equivalendo a um intervalo entre ninhadas de pelo menos três meses.
O tempo médio de permanência foi de 74.6 dias, sem diferença significativa entre sexos. O tempo máximo de permanência foi de 256 dias para uma fêmea, contudo, um maior percentual de machos foi considerado residente (Tabela 3).
Juliomys sp.
(rato-do-mato)
Somente um macho adulto foi capturado em uma armadilha instalada no sub-bosque em junho de 1998.
Oligoryzomys nigripes
(rato-do-mato)
Foram capturados 38 indivíduos 105 vezes. Em dezembro de 1998, um macho jovem foi encontrado morto na mesma armadilha com um macho adulto. Em junho de 1999, um macho jovem foi capturado junto com uma fêmea adulta. De todas as capturas, 15.2 % (n = 16) aconteceram no sub-bosque (χ²105 = 105.00; gl = 1; p < 0.001).
Adultos (com comprimento máximo da cabeça para os machos ≥25.7 mm e para as fêmeas ≥26.7 mm) foram capturados na área de estudo de maio de 1998 a fevereiro de 1999 e de junho a dezembro de 1999, enquanto jovens, em julho e setembro de 1998 e junho e julho de 1999. A proporção sexual foi significativamente favorável aos machos (1.71:1.00; p < 0.05). Registraram-se valores intermediários para a densidade populacional e também para a flutuação ao longo do tempo em relação às demais espécies (Tabela 1; Fig. 3E). A densidade média foi de 2.0 ind/ha (Tabela 1), com picos de densidade durante os meses de inverno e praticamente ausência nos meses mais quentes e úmidos do ano (Fig. 3E), o que é evidenciado pelas correlações obtidas (Tabela 2).
A taxa de sobrevivência foi elevada, com flutuação intermediária ao longo do tempo em relação às demais espécies (Tabela 1; Fig. 3E). A taxa de recrutamento e a variação ao longo do tempo apresentaram valores intermediários (Tabela 1; Fig. 3E). Registrou-se correlação significativa positiva entre a densidade e o recrutamento do mês respectivo e anterior, negativa com a temperatura média mensal do mês respectivo, com um e dois meses de atraso e com a precipitação mensal com um mês de atraso (Tabela 2). A flutuação desta população foi caracterizada principalmente pela entrada de indivíduos na população, principalmente durante o inverno.
Sempre que fêmeas foram capturadas, entre os meses de maio e dezembro, com exceção de junho e novembro de 1998, pelo menos uma encontrava-se reprodutivamente ativa. Da mesma maneira, sempre que machos foram capturados, entre os meses de junho e fevereiro, pelo menos um possuía escroto aumentado. O intervalo entre gestações para uma fêmea foi de dois meses.
O tempo médio de permanência na área de estudo foi de 128.5 dias, sem diferença significativa entre sexos. Um percentual superior de fêmeas (53.8 %) foi considerado residente. O tempo máximo de permanência foi de 385 dias para um macho (Tabela 3), que não foi capturado por aproximadamente 10 meses.
Oryzomys russatus
(rato-do-mato)
Esta foi a espécie com o maior número de indivíduos registrados, 75, capturados 228 vezes. Todas as capturas aconteceram no solo.
Adultos (com comprimento máximo da cabeça para os machos ≥37.5 mm e para as fêmeas ≥34.5 mm) estiveram presentes ao longo dos 24 meses, enquanto jovens em abril e maio de 1998 e 1999 e outubro de 1998. A proporção sexual foi favorável aos machos, desviando do esperado 1:1 (1.78: 1.00; p < 0.001).
A densidade média foi uma das maiores, 2.7 ind/ha, com a menor variação entre as espécies capturadas no PMLP (Tabela 1), sem um padrão de distribuição definido ao longo do tempo (Fig. 3F). A taxa de sobrevivência apresentou valores intermediários, com uma das menores flutuações ao longo do tempo (Tabela 1; Fig. 3F). Registrou-se correlação significativa e negativa entre densidade e taxa de sobrevivência com o mês correspondente, com um e dois meses de atraso (Tabela 2). O. russatus apresentou uma das maiores taxas de recrutamento, com uma baixa flutuação ao longo do tempo (Tabela 1; Fig. 3F). A densidade correlacionou significativamente com o recrutamento do mês correspondente e com um mês de atraso (Tabela 2).
Ambos os sexos foram registrados reprodutivamente ativos ao longo de todo o estudo. Uma fêmea foi capturada lactante em outubro de 1999, prenhe em novembro de 1999, lactante em dezembro de 1999 e novamente prenhe em janeiro de 2000. A produtividade desta fêmea foi, possivelmente, de três ninhadas em um intervalo inferior a seis meses.
O tempo médio de permanência na área de estudo foi de 71.0 dias, sem diferença significativa entre sexos (Tabela 3). O percentual de machos residentes foi superior ao de fêmeas, e o tempo máximo de permanência foi de 231 dias para um macho (Tabela 3).
Oryzomys angouya
(rato-do-mato)
No total, 48 indivíduos foram capturados 158 vezes. Considerando todas as capturas, 15.2% (n = 24) aconteceram em armadilhas instaladas no sub-bosque (χ²158 = 48.222; gl = 1; p < 0,001) e 4.5 % dos indivíduos (n = 7), ao serem soltos, subiram em árvores (χ²158 = 131.241; gl = 1; p < 0.001).
Foram registrados adultos (com comprimento máximo da cabeça para os machos ³ 39.5 mm e para as fêmeas ³ 45 mm) em todos os meses do ano, enquanto jovens estiveram ausentes apenas em julho, agosto e janeiro de 1998 e 1999. A proporção sexual foi favorável aos machos, desviando do esperado 1:1 (2.00: 1.00; p < 0.01). A densidade média foi de 1.9 ind/ha, valor este considerado intermediário em relação às demais espécies (Tabela 1). Apresentou variação intermediária ao longo do tempo (Tabela 1), sem um padrão definido de distribuição (Fig. 3G). A taxa de sobrevivência foi elevada, registrando-se a maior variação ao longo do tempo (Tabela 1; Fig. 3G). A densidade foi positiva e significativamente correlacionada com a taxa de sobrevivência do mês correspondente, com um e dois meses de atraso e três meses de atraso com a temperatura (Tabela 2). A taxa de recrutamento foi a maior observada, apresentando uma das menores variações ao longo do tempo (Tabela 1; Fig. 3G).
Fêmeas adultas não foram capturadas apenas em dezembro de 1998 e 1999. Nos demais meses, pelo menos em um dos dois anos de estudo, foram registradas fêmeas reprodutivamente ativas. Machos com escroto desenvolvido foram capturados em todos os meses do ano. Uma fêmea foi capturada prenhe em setembro de 1999 e novamente em janeiro de 2000.
O tempo médio de permanência foi de 92.4 dias. O tempo máximo de permanência foi de 387 dias para uma fêmea. Contudo, um percentual superior de machos foi considerado residente (Tabela 3).
Oxymycterus aff. judex
(rato-focinhudo, rato-do-brejo)
Foram capturados 21 indivíduos 54 vezes, sempre nas áreas mais úmidas e com vegetação alterada. Todas as capturas aconteceram no solo.
Adultos (com os máximo da cabeça para os machos ≥47.8 mm e para as fêmeas ≥50.2 mm) foram capturados nos meses de maio, junho e novembro de 1998, janeiro de 1999, de maio a outubro de 1999 e janeiro de 2000, enquanto os jovens, de maio a julho de 1998, abril e novembro de 1999 e fevereiro de 2000. Os jovens predominaram durante o inverno de 1998, estando ausentes no inverno seguinte, quando a população atingiu seu maior tamanho. A proporção sexual não desviou do esperado 1:1 (2.00: 1.00; p > 0.05).
A densidade média foi uma das menores, 0.9 ind/ha e a variação ao longo do tempo foi uma das maiores registradas (Tabela 1), com picos de densidade no inverno (Fig. 3H).
As taxas de sobrevivência e de recrutamento apresentaram valores intermediários em relação às demais espécies, com grande variação ao longo do tempo (Tabela 1; Fig. 3H). Registrou-se correlação significativa entre a densidade e o recrutamento do mês correspondente e com dois meses de atraso. A densidade correlacionou-se significativamente com a temperatura média mensal, negativamente com o mês correspondente e positivamente com quatro meses de atraso, e negativamente com a precipitação mensal acumulada com um mês de atraso (Tabela 2).
Fêmeas reprodutivamente ativas foram capturadas em julho de 1999 e janeiro de 2000 com a vulva perfurada e duas estavam lactantes em outubro de 1999. Todos os machos adultos capturados estavam com os escrotos aumentados.
O tempo médio de permanência na área de estudo foi de 76.2 dias. O tempo máximo de permanência foi de 180 dias para uma fêmea e também um percentual superior de fêmeas foi considerado residente (Tabela 3).
DISCUSSÃO
A comunidade de pequenos mamíferos do PMLP
No PMLP foi capturado um maior número de espécies (63.64%) e de indivíduos (77.15%) de roedores em relação aos marsupiais (36.36% das espécies; 22.85% dos indivíduos), como observado previamente para a maioria dos estudos na região Neotropical (ver Stallings, 1989; Voltolini, 1998).
A comunidade do PMPL foi caracterizada principalmente por espécies terrícolas, exceção feita a M. paraguayanus, capturada principalmente no sub-bosque. D. aurita, O. nigripes e O. angouya foram capturados também no sub-bosque, contudo, um maior número de capturas aconteceu no solo (Graipel, 2003). As capturas de um único indivíduo de Juliomys sp. em sub-bosque e de C. minimus em uma armadilha próxima a um curso d'água correspondem aos hábitos arborícola e semi-aquático descritos respectivamente para estes gêneros (Graipel, 2003; Monteiro-Filho e Graipel, 2005).
Dinâmica populacional
No PMLP observou-se uma marcante sazonalidade no aparecimento das diferentes categorias de idade dos marsupiais, enquanto os roedores não apresentaram um padrão bem definido no recrutamento de jovens.
Para a maioria das espécies do PMLP, onde há uma grande extensão florestal, a proporção sexual foi desviada para os machos, o que normalmente é atribuído aos maiores deslocamentos que eles realizam e considerado um padrão entre os mamíferos (e.g., Ralls et al., 1986; Chepko-Sade e Halpin, 1987). Para D. aurita, uma das espécies com um maior número de capturas e uma boa caracterização quanto à categoria de idade, o recrutamento de machos subadultos foi o principal responsável pelo desvio na proporção sexual da espécie, o que pode ser atribuído à dispersão dos indivíduos quando alcançam esta idade. Em ilhas pequenas (e.g., Cherem et al., 1996; C. Oliveira, com. pessoal), onde a dispersão de machos é, no mínimo, improvável, verifica-se uma igualdade na proporção sexual. Em fragmentos florestais isolados (e.g., Graipel e Santos-Filho, 2006; Cáceres e Monteiro-Filho, 1998; Quental et al., 2001; Fernandez et al., 2003), onde a emigração (mas não a imigração) de machos pode ocorrer com uma maior freqüência, o desvio foi favorável às fêmeas. Esses dados e o maior tempo de permanência das fêmeas da maioria das espécies no PMLP (Tabela 3) corroboram a hipótese do maior deslocamento e/ou dispersão dos machos. Estas condições, segundo Roper (2003), não acarretam em preocupações importantes para a conservação das espécies.
As estratégias demográficas mostraram-se marcantes para as várias espécies capturadas no PMLP. Aquelas com maiores níveis populacionais, D. aurita, A. montensis e O. russatus, apresentaram as menores flutuações (CV < 50%; Tabela 1). As taxas de sobrevivência registraram queda com o aumento da densidade, apesar de a correlação não ter sido significativa para A. montensis, caracterizando-as como espécies dependentes da densidade (Yoccoz et al., 2001). Por outro lado, para M. paraguayanus, N. squamipes e O. angouya, as espécies com maiores flutuações (CV > 50%; Tabela 1), verificou-se um aumento da densidade populacional após períodos contínuos de incremento da taxa de sobrevivência (Fig. 3A, 3B e 3G), como também registrado por Gentile et al. (2000) para N. squamipes. Para Bergallo e Magnusson (1999), o aumento da taxa de sobrevivência de N. squamipes teve relação direta com a disponibilidade de frutos, de forma semelhante esta também ocorreu no PMLP (Cáceres et al., 2002). De um modo geral, as taxas de sobrevivência podem ser consideradas relativamente altas para as espécies do PMLP. Apenas para O. angouya verificou-se valor inferior a 50% (Tabela 1).
Embora as taxas de recrutamento para todas as espécies tenham sido relativamente baixas (< 50 %; Tabela 1), houve uma quase total substituição dos indivíduos de M. paraguayanus, L. crassicaudata, O. nigripes, O. angouya e O. aff. judex ao longo do período de estudo. Apenas para O. russatus, O. aff. judex e O. nigripes, o recrutamento teve relação direta com o aumento dos níveis populacionais.
A flutuação da maioria das populações parece estar relacionada com fatores climáticos, que podem interferir na disponibilidade de alimentos (ver adiante em reprodução) fazendo com que fossem registrados crescimentos contínuos nos níveis populacionais da maioria das espécies a partir do verão, com uma tendência significativa para elevação até o início do inverno. Além disso, os picos populacionais para D. aurita, O. nigripes e O. aff. judex foram registrados nos períodos mais frios e secos, quando os recursos alimentares utilizados por estas espécies, como frutos e insetos (Fonseca et al., 1996), diminuem grandemente (Bergallo e Magnusson, 1999; Cáceres et al., 2002). Isto poderia propiciar ainda um maior número de capturas e uma maior probabilidade de captura (Smith e Blessing, 1969). Nesses períodos, a imigração de jovens pode ter um papel determinante nos níveis de densidade. O aumento da densidade de D. aurita nesses períodos, no PMLP, foi influenciado principalmente pelo recrutamento de machos subadultos, coincidindo com o início do período reprodutivo da espécie.
Além disso, registrou-se o surgimento de espécies ausentes no PMLP em outras estações do ano, como no caso de O. nigripes e O. aff. judex. Oliveira (1985) registrou uma seqüência de ocupação em direção de hábitats alternativos por roedores, quando ocorria aumento de densidade ou uma menor disponibilidade de recursos. Esta condição poderia criar um padrão de imigração a partir de suas áreas de origem, como observado no Chile por Meserve et al. (2001), e conseqüentemente levar a uma proporção sexual desfavorável às fêmeas, por serem mais residentes, como observado no PMLP para M. paraguayanus, O. nigripes e O. angouya, e por Wolff (1993) para pequenos mamíferos.
Para M. paraguayanus, N. squamipes, O. angouya e O. aff. judex, houve uma tendência significativa para elevação da densidade em períodos mais quentes e úmidos, geralmente com alguns meses de atraso (Fig. 3B, 3D, 3G e 3H), possivelmente devido à entrada na população de indivíduos nascidos a partir do fim do inverno e primavera.
Reprodução
A relação entre a reprodução sazonal de algumas espécies de pequenos mamíferos e fatores climáticos tem sido considerada por alguns autores (e.g., Streilein, 1982; Bergallo, 1994; Bergallo e Magnusson, 1999), estando principalmente associada aos ciclos anuais de chuvas (Heideman e Bronson, 1990). A reprodução nesses períodos possivelmente está associada a uma maior disponibilidade de alimentos (Atramentowicz, 1982; Julien-Laferrière e Atramentowicz, 1990; Bergallo e Magnusson, 1999). Para os marsupiais D. aurita e L. crassicaudata, a reprodução ocorreu durante o inverno e a primavera, sendo registrados filhotes no marsúpio ainda no verão. Contudo, a entrada de infantes na população, período em que ocorre o desmame e considerado o mais crítico na vida destes animais (Tyndale-Biscoe, 1979), coincide com o início do período de chuvas, na primavera e verão. Estas espécies apresentaram um padrão reprodutivo poliéstrico sazonal, que pode ser considerado comum aos marsupiais neotropicais de maior porte em áreas tropicais e subtropicais (Fleming, 1973; Fonseca e Kierulff, 1989; Cerqueira et al., 1993). Entre os roedores, o mesmo foi verificado para N. squamipes, apesar de a reprodução para esta espécie ter se concentrado em períodos mais quentes e chuvosos do ano, estendendo-se por mais tempo. A. montensis, O. russatus e O. angouya apresentaram poliestria panestacional, mais comum entre os roedores (Pereira et al., 1993).
Tempo de permanência
Uma vez que a distribuição das armadilhas em grades favorece o estudo de populações por proporcionar mais recapturas que em transecções (Monteiro-Filho e Graipel, 2005), seria esperado também um maior tempo de permanência para animais capturados em grades, simplesmente pelo fato de as armadilhas cobrirem uma maior superfície da área de estudo, contemplando assim, toda ou uma grande parcela da área de atividade de muitos dos indivíduos. Neste sentido, seria esperado um maior tempo de permanência para as espécies capturadas no presente estudo, com o uso de grade, em relação a espécies similares capturadas em transecções por Fonseca e Kierulff (1989) e D'Andrea et al. (1999). Como observado em outros estudos (e.g., Fonseca e Kierulff, 1989; D'Andrea et al., 1999; O'Connell, 1989), registrou-se, em geral, um maior tempo de permanência das fêmeas no PMLP,resultado que pode estar relacionado ao fato de as fêmeas de pequenos mamíferos serem mais territoriais que os machos (Wolff, 1993).
Diversidade e conservação
O número de espécies de pequenos mamíferos capturados no PMLP foi inferior ao de outros estudos em áreas de Mata Atlântica (e.g., Fonseca e Kierulff, 1989, n = 19; Stallings, 1989, n = 17). Contudo, Fonseca e Kierulff (1989) e Stallings (1989) utilizaram um maior esforço de amostragem e abrangeram uma maior heterogeneidade de hábitats, o que geralmente implica em maior riqueza de espécies (August, 1983; Voss e Emmons, 1996).
Por se localizar em uma ilha, uma menor riqueza de espécies no PMLP em relação ao continente poderia ser esperada (Whittaker, 1998). Contudo, trata-se de uma ilha relativamente grande e muito próxima ao continente, tendo se isolado possivelmente há menos de 8 000 anos. Isto possibilitaria uma maior similaridade nos parâmetros comunitários entre ilha e continente, como verificado por Bergallo et al. (1998) ao comparar uma área de Mata Atlântica no continente no litoral sul de São Paulo (Estação Ecológica da Juréia-Itatins) e uma ilha próxima (Ilha do Cardoso).
Bergallo et al. (1998) registraram sete espécies de pequenos mamíferos em uma área de Mata Atlântica na Ilha do Cardoso. Este menor número de espécies em relação ao PMLP poderia ser atribuído ao menor esforço de amostragem de Bergallo et al. (1998), ao menor tamanho daquela Ilha (225 km2), aproximadamente a metade do tamanho da Ilha de Santa Catarina (451 km2; Gaplan, 1986), e/ou ao fato de uma menor diversidade de espécies ser normalmente encontrada em florestas primárias, pois estas geralmente possuem uma menor heterogeneidade de ambientes e podem abrigar um menor número de espécies (Fonseca, 1989). Bergallo et al. (1998) utilizaram apenas armadilhas no solo, o que poderia também refletir a baixa riqueza, porém apenas uma espécie no PMLP foi capturada exclusivamente no sub-bosque.
No entanto, no PMLP (floresta secundária), o número de espécies foi inferior àquele obtido por Voltolini (1998; n = 16) (floresta primária) no continente próximo, na mesma latitude que o PMLP (27º43'S - 28º13'S e 49º59'W - 48º33'W; 27º43'S e 48º32'W, respectivamente), mesmo com um menor esforço de amostragem nesta última área. Dentre as espécies capturadas por Voltolini (1998), Brucepattersonius iheringi, Delomys sublineatus e Thaptomys nigrita têm sido registradas apenas em ambientes de encosta com altitudes mais elevadas (Vieira e Monteiro-Filho, 2003). O estudo conduzido por Voltolini (1998) foi realizado acima de 400 m de altitude, enquanto a área de estudo no PMLP localiza-se abaixo de 100 m de altitude. Relação similar quanto à altitude foi observada por Vieira e Monteiro-Filho (2003), trabalhando em duas áreas de florestas primárias, onde registraram 11 espécies na área mais baixa (100 m) e 16 espécies na área mais elevada (900 m).
Uma redução na riqueza da comunidade de pequenos mamíferos no PMLP pode também ter sido influenciada pela degradação ambiental na Ilha de Santa Catarina, como demonstram os dados de Caruso (1990). Originalmente as áras de floresta ombrófila densa ocupavam cerca de 313 km². Em 1938, estavam reduzidas a 52.65 km² e, em 1978, a somente 39.9 km², considerando-se as florestas que sofreram apenas desmatamento seletivo de algumas árvores adultas. Esta redução da cobertura florestal é apontada como um dos principais fatores que levaram à extinção das espécies de grande porte e de aproximadamente 50 % das espécies de médio porte (Olimpio, 1995; Graipel et al., 2001).
Assim, os fatores altitude e degradação pretérita do ambiente foram provavelmente as principais causas da menor riqueza de espécies de pequenos mamíferos observada no PMLP em relação ao continente próximo (Voltolini, 1998). Além disso, a perturbação da composição desta comunidade, evidenciada pela alta densidade de D. aurita, pode ser um reflexo da redução da diversidade de carnívoros da Ilha, como observado por Fonseca e Robinson (1990) para uma área de Mata Atlântica no sudeste do Brasil.
Entretanto, a avaliação da extinção de pequenos mamíferos na Ilha de Santa Catarina é uma tarefa muito difícil, pois não se dispõe de registros anteriores à década de 1980, ao contrário do que pôde ser verificado para mamíferos de médio e grande porte, em função dos registros bibliográficos, em sítios arqueológicos e por entrevistas (Olimpio, 1995; Graipel et al., 2001).
A partir da década de 1940 até o início da década de 1990, verificou-se uma regeneração significativa da cobertura florestal na Ilha (Caruso, 1990; Veado, 2004). Esta condição, associada com a diminuição da caça, é percebida com a confirmação da ocorrência de espécies de mamíferos de médio porte até então citadas somente em entrevistas (Graipel et al., 2001), como a irara Eira barbara e o ouriço Sphiggurus villosus. É possível que pequenos felinos e outros carnívoros considerados extintos na Ilha ocorram em baixas densidades e tenham seus níveis populacionais aumentados no futuro, minimizando as perturbações verificadas na assembléia de pequenos mamíferos do PMLP. No entanto, nos últimos dez anos, um aumento da alteração e fragmentação florestal, decorrente da especulação imobiliária, tem sido novamente verificado na Ilha (Veado, 2004), o que pode vir a comprometer os avanços na conservação deste ecossistema obtidos nas últimas décadas.
AGRADECIMENTOS
Agradecemos a Ivo R. Ghizoni Jr., Claudio R. Graipel, Carlos H. Oliveira e aos estudantes de graduação da UFSC pela ajuda nos trabalhos de campo. A FLORAM (Fundação Municipal do Meio Ambiente de Florianópolis), e aos fiscais do Parque Municipal da Lagoa do Peri pela autorização e prontidão na prestação de auxílio em vários momentos. A Nilton C. Cáceres pelas revisões dos manuscritos. Este trabalho foi parcialmente financiado pelo programa PICDT/CAPES.
LITERATURA CITADA
ATRAMENTOWICZ M. 1982. Influence du milieu sur l'activité locomotrice et la reproduction de Caluromys philander (L.). Revue d'Ecologie 36:376-395. [ Links ]
AUGUST PV. 1983. The role of habitat complexity and heterogeneity in structuring tropical mammal communities. Ecology 64:1495-1507. [ Links ]
AVILA-PIRES FD. 1999. Mamíferos descritos do estado de Santa Catarina, Brasil. Revista Brasileira de Zoologia 16:51-62. [ Links ]
AYRES M, MJR AYRES, DL AYRES e AS SANTOS. 2000. BioEstat 2.0: aplicações estatísticas nas áreas das ciências biológicas e médicas. Belém: Sociedade Civil Mamirauá, CNPq, Brasília. [ Links ]
AZEVEDO TR, D EL ACHKAR, MF MARTINS e A XIMENEZ. 1982. Lista sistemática dos mamíferos de Santa Catarina conservados nos principais museus do estado. Revista Nordestina de Biologia 5:93-104. [ Links ]
BERGALLO HG. 1994. Ecology of a small mammal community in an Atlantic Forest area in southeastern Brazil. Studies on Neotropical Fauna and Environment 29:197-217. [ Links ]
BERGALLO HG. 1995. Comparative life-history characteristics of two species of rats, Proechimys iheringi and Oryzomys intermedius, in an Atlantic Forest of Brazil. Mammalia 59:51-64 [ Links ]
BERGALLO HG, CF VERA Y CONDE, EB BITTENCOURT, DEP BOSSI e CFD ROCHA. 1998. As similaridades nos parâmetros comunitários de pequenos mamíferos de duas áreas de Mata Atlântica do sul de São Paulo, sudeste do Brasil. Pp. 939-949, em: Anais do VIII Seminário Regional de Ecologia, Volume VIII, São Carlos. [ Links ]
BERGALLO HG e WE MAGNUSSON. 1999. Effects of climate and food availability on four rodent species in southeastern Brazil. Journal of Mammalogy 80:472-486. [ Links ]
CÁCERES NC e ELA MONTEIRO-FILHO. 1997. Reprodutive biology of the common opossum, Didelphis marsupialis (Mammalia: Marsupialia), in southern Brazil. Brenesia 47-48:117-124. [ Links ]
CÁCERES NC e ELA MONTEIRO-FILHO. 1998. Population dynamics of the common opossum, Didelphis marsupialis (Mammalia, Marsupialia), in southern Brazil. International Zeitschrift für Säugetierkunde 63:169-172. [ Links ]
CÁCERES NC e ELA MONTEIRO-FILHO. 1999. Tamanho corporal em populações naturais de Didelphis (Mammalia: Marsupialia) do sul do Brasil. Revista. Brasileira de Biologia 59:461-469. [ Links ]
CÁCERES NC, IR GHIZONI-JÚNIOR e ME GRAIPEL. 2002. Diet of two marsupials, Lutreolina crassicaudata and Micoureus paraguayanus, in a coastal Atlantic Forest island of Brazil. Mammalia 66:331-340. [ Links ]
CARUSO MML 1990. O desmatamento na Ilha de Santa Catarina de 1500 aos dias atuais. 2 a.ed. Editora da UFSC, Florianópolis. [ Links ]
CECCA (Centro de Estudos, Cultura e Cidadania). 1997. Unidades de Conservação e áreas protegidas da Ilha de Santa Catarina: caracterização e legislação. Editora Insular, Florianópolis. [ Links ]
CERQUEIRA R, R GENTILE, FAS FERNANDEZ e PS D'ANDREA. 1993. A five-year population study of an assemblage of small mammals in Southeastern Brazil. Mammalia 57: 507-517. [ Links ]
CHARLES-DOMINIQUE P, M ATRAMENTOWICZ, M CHARLES-DOMINIQUE, H GÉRARD, A HLADIK, CM HLADIK e MF PRÉVOST. 1981. Les mammifères frugivores arboricoles nocturnes d'une foret guyanaise: inter-relations plantes-animaux. Revue d'Ecologie (Terre et Vie) 35:341-435. [ Links ]
CHEPKO-SADE BD e ZT HALPIN. 1987. Mammalian dispersal patterns: the effects of social structure on population genetics. University of Chicago Press, Chicago. [ Links ]
CHEREM JJ e DM PEREZ. 1996. Mamíferos terrestres de floresta de araucária no município de Três Barras, Santa Catarina, Brasil. Biotemas 9:29-46. [ Links ]
CHEREM JJ, ME GRAIPEL, ME MENEZES e M SOLDATELI. 1996. Observações sobre a Biologia do Gambá (Didelphis marsupialis) na Ilha de Ratones Grande, Estado de Santa Catarina, Brasil. Biotemas 9:47-56. [ Links ]
CHEREM JJ, PC SIMÕES-LOPES, SL ALTHOFF, ME GRAIPEL. 2004. Lista dos mamíferos do estado de Santa Catarina, sul do Brasil. Mastozoología Neotropical 11(2):151-184. [ Links ]
CIMARDI AV. 1996. Mamíferos de Santa Catarina. FATMA, Florianópolis. [ Links ]
D'ANDREA PS, R GENTILE, R CERQUEIRA, CEV GRELLE, C HORTA e L REY. 1999. Ecology of small mammals in a Brazilian rural area. Revista Brasileira de Zoologia 16:611-620. [ Links ]
DAVIS DE. 1947. Notes on the life histories of some Brazilian mammals. Boletim do Museu Nacional-Zoologia 76:1-8. [ Links ]
EISENBERG JF. 1981. The mammalian radiations. The University of Chicago Press, Chicago. [ Links ]
FERNANDEZ FAS. 1995. Métodos para estimativas de parâmetros populacionais por captura, marcação e recaptura. Oecologia brasiliensis 2:1-26. [ Links ]
FERNANDEZ FAS, CS BARROS e M SANDINO. 2003. Biased sex ratios in populations of the woolly mouse opossum Micoureus paraguayanus in Atlantic Coastal Forest fragments. Natureza & Conservação 1:78-84. [ Links ]
FLEMING TH. 1973. The reproductive cycles of three species of opossums and other mammals in the Panama Canal Zone. Journal of Mammalogy 54:439-455. [ Links ]
FLEMING TH. 1975. The role of small mammals in tropical ecosystems. Pp. 269-298, em: Small mammals: their productivity and population dynamics (FB Golley, K Petrusewicz e L Ryszkowski, eds.). Cambridge University Press, Cambridge. [ Links ]
FONSECA GAB. 1989. Small mammal species diversity in brazilian tropical primary and secondary forest of different sizes. Revista Brasileira de Zoologia 6:381-422. [ Links ]
FONSECA GAB e MCM KIERULFF. 1989. Biology and natural history of Brazilian Atlantic Forest small mammals. Bulletin of the Florida State Museum, Biological Sciences 34:99-152. [ Links ]
FONSECA GAB e JG ROBINSON. 1990. Forest Size and Structure: Competitive and predatory effects on small mammal communities. Biological Conservation 53:265-294. [ Links ]
FONSECA GAB, G HERRMANN, YLR LEITE, RA MITTERMEIER; AB RYLANDS e JL PATTON. 1996. Lista anotada dos mamíferos do Brasil. Occasional Papers in Conservation Biology 4:1-38. [ Links ]
FULLAGAR PJ e PA JEWELL. 1965. Marking small rodents and the difficulties of using leg rings. Journal of Zoology 147:224-228. [ Links ]
GAPLAN. 1986. Atlas de Santa Catarina. Gabinete de Planejamento e Coordenação Geral, Rio de Janeiro. [ Links ]
GENTILE R, PS D'ANDREA, R CERQUEIRA e LS MAROJA. 2000. Population dynamics and reproduction of marsupials and rodents in a Brazilian rural area: a five-year study. Studies on Neotropical Fauna and Environment 35:1-9. [ Links ]
GRAIPEL ME, JJ CHEREM, ME MENEZES, M SOLDATELI, DA MACHADO e PCA GARCIA. 1997. Vertebrados da Ilha de Ratones Grande, Santa Catarina, Brasil. Biotemas 10:105-122. [ Links ]
GRAIPEL ME, JJ CHEREM e A XIMENEZ. 2001. Mamíferos terrestres não voadores da Ilha de Santa Catarina, sul do Brasil. Biotemas 14:109-140. [ Links ]
GRAIPEL ME. 2003. Contribuição ao estudo da mastofauna do estado de Santa Catarina, sul do Brasil. Tese de doutorado, Pontifícia Universidade Católica do Rio Grande do Sul, Rio Grande do Sul. [ Links ]
GRAIPEL ME, JJ CHEREM, PRM MILLER e L GLOCK. 2003. Trapping small mammals in the forest understory: a comparison of three methods. Mammalia 67:551-558. [ Links ]
GRAIPEL ME e M SANTOS-FILHO. 2006. Reprodução e dinâmica populacional de Didelphis aurita Wied-Neuwied (Mammalia: Didelphimorphia) em ambiente periurbano na Ilha de Santa Catarina, Sul do Brasil. Biotemas 19:65-73. [ Links ]
HEIDEMAN PD e FH BRONSON. 1990. Photoperiod, melatonin secretion, and sexual maturation in a tropical rodent. Biology of Reproduction 43:745-750. [ Links ]
IPUF - Instituto de Planejamento Urbano de Florianópolis. 1978. Plano Diretor do Parque Municipal da Lagoa do Peri. Relatório Final. Florianópolis. [ Links ]
JENNRICH RJ e FB TURNER. 1969. Measurement of non-circular home range. Journal of Theoretical Biology 22:227-237. [ Links ]
JULIEN-LAFERRIÈRE D e M ATRAMENTOWICZ. 1990. Feeding and reproduction of three didelphid marsupials in two Neotropical Forests (French Guiana). Biotropica 22:404-415. [ Links ]
KLEIN RM. 1990. Estrutura, composição florística, dinamismo e manejo da "Mata Atlântica" (Floresta Ombrófila Densa) do sul do Brasil. II Simp. Ecossist. Costa Sul e Sudeste Brasileira: Estrutura, Função e Manejo, São Paulo 1:259-286. [ Links ]
KREBS CJ. 1966. Demographic changes in fluctuating populations of Microtus californicus. Ecological Monographs 36:239-273. [ Links ]
MASSOIA E. 1973. Observaciones sobre el género Lutreolina en la República Argentina (Mammalia - Marsupialia - Didelphidae). Revista de Investigaciones Agropecuarias, INTA 10:13-20. [ Links ]
MESERVE PL, WB MILSTEAD e JR GUTIÉRREZ. 2001. Results of a food addition experiment in a north-central Chile small mammal assemblage: evidence for the role of "bottom-up" factors. Oikos 94:548-556. [ Links ]
MONTEIRO-FILHO ELA e VS DIAS. 1990. Observações sobre a biologia de Lutreolina crassicaudata (Mammalia, Marsupialia). Revista Brasileira de Biologia, 50:393-399. [ Links ]
MONTEIRO-FILHO ELA e AS ABE. 1999. Catchability of the white-eared opossum, Didelphis albiventris, in a disturbed area of southeastern Brazil. Arquivos de ciencias veterinarias e zoologia da UNIPAR 2:31-35. [ Links ]
MONTEIRO-FILHO ELA e ME GRAIPEL. 2005. Captura e Marcação. Pp. 17-27, em: Os marsupiais do Brasil: biologia, ecologia e evolução (NC Cáceres e ELA Monteiro-Filho, eds.). Editora da Universidade Federal do Mato Grosso do Sul, Campo Grande. [ Links ]
O'CONNELL MA. 1989. Population dynamics of neotropical small mammals in seasonal habitats. Journal of Mammalogy 70:532-548. [ Links ]
OLIMPIO J. 1995. Conservação da fauna de mamíferos silvestres da Ilha de Santa Catarina: Aspectos biogeográficos, históricos e sócio-ambientais. Dissertação de mestrado, Universidade Federal de Santa Catarina, Santa Catarina, Brasil. [ Links ]
OLIVEIRA LFB. 1985. Estrutura e ordenação espaço-temporal de uma congregação de roedores no sul da planície costeira do Rio Grande do Sul, Brasil (Mammalia, Rodentia, Cricetidae). Dissertação de mestrado, Universidade Federal do Rio Grande do Sul, Rio Grande do Sul, Brasil. [ Links ]
OLMOS F. 1991. Observations on the behavior and population dynamics of some Brazilian Atlantic Forest rodents. Mammalia 55:555-565. [ Links ]
PEREIRA LA, WA CHAGAS e JE COSTA. 1993. Ecologia de pequenos mamíferos silvestres da Mata Atlântica, Brasil. I. Ciclos reprodutivos de Akodon cursor, Nectomys squamipes e Oryzomys nigripes (Rodentia, Cricetinae). Revista Brasileira de Zoologia 10:389-398. [ Links ]
PIANKA ER. 1983. Evolutionary Ecology. Harper & Row, 3rd ed., New York. [ Links ]
QUADROS J e NC CÁCERES. 2001. Ecologia e conservação de mamíferos na Reserva Volta Velha, SC, Brasil. Acta Biológica Leopoldensia 23:213-224. [ Links ]
QUENTAL TB, FAS FERNANDEZ, ATC DIAS e FS ROCHA. 2001. Population dynamics of the marsupial Micoureus paraguayanus in small fragments of Atlantic Coastal Forest in Brazil. Journal Tropical Ecology 17:339-352. [ Links ]
RALLS K, HP HARVEY e AM LYLES. 1986. Inbreeding in natural populations of birds and mammals. Pp. 35-56, em: Conservation biology - the science of scarcity and diversity (ME Soulè, ed.). Sinauer Associates, Sunderland, USA. [ Links ]
ROPER JJ. 2003. Desvio nas razões sexuais ou análises tendenciosas? Natureza & Conservação 1:19-21. [ Links ]
SIPINSKI EAB e NR REIS. 1995. Dados ecológicos dos quirópteros da Reserva Volta Velha, Itapoá, Santa Catarina, Brasil. Revista Brasileira de Zoologia 12:519-528. [ Links ]
SMITH GC e RW BLESSING. 1969. Trap response and food availability. Journal of Mammalogy 50:368-369. [ Links ]
STALLINGS JR. 1989. Small mammal inventories in an eastern Brazilian park. Bulletin of the Florida State Museum, Biological Sciences 34:153-200. [ Links ]
STREILEIN KE. 1982. The ecology of small mammals in the semiarid Brazilian caatinga. III. Reproductive biology and population ecology. Annals of Carnegie Museum 51:251-269. [ Links ]
TALAMONI SA e MM DIAS. 1999. Population and community ecology of small mammals in southeastern Brazil. Mammalia 63:167-181. [ Links ]
TYNDALE-BISCOE CH. 1979. Ecology of small marsupials. Pp. 343-379, em: Ecology of small mammals (DM Stoddart, ed.). Chapman & Hall Ltd., London. [ Links ]
VEADO RA. 2004. Os sistemas naturais do Município de Florianópolis. Pp. 47-53, em: Atlas do Município de Florianópolis (MDA Bastos, coord.). IPUF, Florianópolis, Brasil. [ Links ]
VIEIRA EM e ELA MONTEIRO-FILHO. 2003. Vertical stratification of small mammals in the Atlantic rain Forest of southeastern Brazil. Journal of Tropical Ecology 19:501-507. [ Links ]
VOLTOLINI JC. 1998. Estratificação vertical de marsupiais e roedores na Floresta Atlântica do Sul do Brasil. Dissertação de Mestrado. Instituto de Biociências da USP, São Paulo. [ Links ]
VOSS RS e LH EMMONS. 1996. Mammalian diversity in Neotropical lowland rainforests: a preliminary assessment. Bulletin of the American Museum of Natural History 230:1-115. [ Links ]
WALLAUER JP, M BECKER, LG MARINS-SÁ, LM LIERMANN, SH PERRETTO e V SCHERMACK. 2000. Levantamento dos mamíferos da Floresta Nacional de Três Barras - Santa Catarina. Biotemas 13:103-127. [ Links ]
WHITTAKER RJ. 1998. Island Biogeography - Ecology, Evolution and Conservation. Oxford Universituy Press, Oxford. [ Links ]
WOLFF JA. 1993. Why are female small mammals territorial? Oikos 68:364-370. [ Links ]
YOCCOZ NG, NC STENSETH, H HENTTONEN e AC PRÉVOT-JULLIARD. 2001. Effects of food addition on the seasonal density-dependent structure of bank vole Clethrionomys glareolus populations. Journal of Animal Ecology 70:713-720. [ Links ]
Recibido 13 diciembre 2004.
Aceptación final 17 noviembre 2005.

