










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkMastozoología neotropical
versão impressa ISSN 0327-9383versão On-line ISSN 1666-0536
Mastozool. neotrop. v.13 n.1 Mendoza jan./jun. 2006
Distribución de Necromys lactens y Phyllotis osilae (Rodentia: Cricetidae: Sigmodontinae) en el noroeste argentino: modelos predictivos basados en el concepto de nicho ecológico
J. Pablo Jayat 1, 2 y Silvia Pacheco 1, 3
1 Laboratorio de Investigaciones Ecológicas de las Yungas (LIEY), Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Casilla de Correo 34, 4107 Yerba Buena, Tucumán, Argentina. 2 Área de Biodiversidad, Fundación ProYungas, 4107 Yerba Buena, Tucumán, Argentina, <pjayat@proyungas.com.ar>. 3 Área de Sistema de Información Geográfica, Fundación ProYungas, 4107 Yerba Buena, Tucumán, Argentina, <pacheco@proyungas.com.ar>
Abstract: Distribution of Necromys lactens and Phyllotis oscilae (Rodentia: Cricetidae: Sigmodontinae) in northwestern Argentina: predictive models based on the concept of ecological niche. We used presence data from field records, museum specimens and literature sources to model the potential distribution of two species of sigmodontine rodents, Necromys lactens and Phyllotis osilae in northwestern Argentina, based on environmental variables. We used the DOMAIN program, which based on presence data defines the environmental space that imposes conditions to the species distribution. We worked with two data sets, one constituted only by reliable data and the other one constituted by these reliable data plus other information considered more doubtful because of its controvertible identification and location. The models obtained from the reliable data set were more efficient in the prediction of distribution than doubtful data, with Kappa index of 0.69 and 0.63 for Necromys lactens and Phyllotis osilae, respectively. These results indicate that both species are restricted to the upper portions of the most important mountains in the region and that their distribution consists of islands or patches immerse in a complex matrix of forest and semi-desert environment. Both species were associated to high altitude grasslands and ecotone environments between 1200 and 3200 m elevation, being P. osilae 's distribution a little more extended than N. lactens.
Key words. Distribution. DOMAIN. Ecological niche model. NW Argentina. Sigmodontine rodents.
INTRODUCCIÓN
El conocimiento preciso de las áreas de distribución de las especies constituye la base sobre la cual se estructuran estudios avanzados acerca de su taxonomía, ecología, biogeografía y evolución. Buenas descripciones de la distribución real y potencial de los distintos taxones tienen implicancias importantes para su conservación, ya que constituyen el fundamento de la identificación de las áreas de simpatría, endemismo y diversidad, así como también de la utilización de hábitat por parte de los mismos.
El conocimiento de las áreas de distribución de las especies de sigmodontinos del noroeste argentino permanece en un estado incipiente. Aunque en las últimas décadas se han realizado aportes importantes que incluyen listas sistemáticas de especies de mamíferos para distintas provincias (Mares et al., 1981; Ojeda y Mares, 1989; Mares et al., 1997; Díaz y Barquez, 1999; Díaz et al., 2000), áreas naturales protegidas (Heinonen y Bosso, 1994; Gil y Heinonen Fortabat, 2003) y localidades aisladas del noroeste argentino (Ortiz et al., 2000), estos aportes expresan la distribución mediante datos "puntuales". Hasta el momento no se ha intentado delimitar un área que represente una hipótesis de distribución regional de alguna especie sobre la base de fundamentos metodológicos explícitos. Algunos intentos de sombreado de las áreas de distribución de las especies presentes en el noroeste argentino como el de los trabajos de Olrog y Lucero (1981), Mares et al. (1989), Barquez et al. (1991) y Díaz y Barquez (2002) se sustentan en el punto de vista particular (generalmente no explícito) de los autores, presentan un grado de resolución espacial muy bajo y/o sólo se restringen a un área particular dentro de la región.
En años recientes, el desarrollo de modelos de distribución de especies basados en el concepto de nicho ecológico (Stockwell y Noble, 1992; Carpenter et al., 1993), el avance de los Sistemas de Información Geográfica y la creciente accesibilidad, vía Internet, a información espacial relacionada con variables ambientales, han aumentado considerablemente nuestra capacidad de predecir las áreas de distribución de las especies. Estos modelos exploran las relaciones existentes entre datos de presencia conocida y variables ambientales que determinan las distribuciones, permitiendo extrapolar el área de distribución hacia zonas geográficas no muestreadas. Estos modelos se han utilizado, además, para evaluar hipótesis biogeográficas (Anderson et al., 2002; Peterson et al., 2002), procesos evolutivos (Peterson et al., 1999; Martínez-Meyer et al., 2003) y para determinar áreas prioritarias de conservación (Feria y Peterson, 2002; Loiselle et al., 2003; Vargas et al., 2004).
En este trabajo determinamos las áreas potenciales de distribución de dos especies de roedores sigmodontinos (Necromys lactens y Phyllotis osilae) en el noroeste argentino, utilizando el programa DOMAIN. Para la construcción de los modelos se utilizaron dos conjuntos de datos, (i) datos de presencia basados en identificaciones seguras de las especies, y (ii) los mismos datos más registros de presencia considerados "dudosos" en cuanto a la identificación de las especies y/o la localización correcta de los mismos. Nuestra experiencia de trabajo de más de diez años en la región nos permitió hipotetizar que las dos especies están limitadas en su distribución a la región de los Pastizales de Neblina (sensu Brown y Grau, 1993) y áreas ecotonales, y que los mapas generados a partir de datos de presencia dudosos serán menos efectivos en la definición del área de distribución.
MATERIALES Y MÉTODOS
Área de estudio
El área de estudio comprende las provincias de Jujuy, Salta, Tucumán, Santiago del Estero y Catamarca, en el noroeste de Argentina. Esta región se caracteriza por una gran heterogeneidad ambiental, consecuencia de la disposición del relieve topográfico y la dirección de los vientos estacionales que producen lluvias diferenciales en distintos sectores. Estas condiciones determinan la presencia de distintas unidades de paisaje que incluyen a las ecorregiones del Chaco Seco, Yungas, Monte de Sierras y Bolsones, Puna y Altos Andes (Burkart et al., 1999).
La cordillera Oriental, el sistema de las Sierras Subandinas, las Sierras Centrales y el extremo norte de las Sierras Pampeanas constituyen los principales accidentes orográficos de la región (Aceñolaza y Toselli, 1981). Los cordones montañosos que forman estos sistemas tienen una orientación norte-sur y están relativamente aislados unos de otros. Las Sierras Subandinas y Centrales muestran una disposición insularizada hacia el este de la región y están rodeadas de bosques de Yungas y Chaco Seco. El resto de los sistemas orográficos son relativamente continuos, alcanzan mayor altitud (algunos superando los 5000 m de altura) y separan a los ambientes boscosos de Yungas y Chaco Seco en la vertiente oriental, de los ambientes abiertos del Monte, Puna y Altos Andes en la vertiente occidental. En la cumbre de todos estos sistemas montañosos, aproximadamente entre los 1500 y 3000 m de altitud, se encuentran los Pastizales de Neblina. Estas comunidades vegetales dominadas por gramíneas y caracterizadas por un importante aporte de humedad durante gran parte del año, forman "islas" de pastizales inmersos en una matriz de bosques y ambientes áridos de altura.
Presencia de especies
Los datos puntuales de distribución de ambas especies se obtuvieron de viajes de campo y de revisión de colecciones sistemáticas y bibliografía especializada. Los datos de campo fueron colectados entre 1998 y 2005 e incluyen observaciones personales de ambas especies y ejemplares colectados y registrados en el catálogo personal de uno de los autores (JPJ) que serán depositados en la Colección de Mamíferos Lillo (CML) en San Miguel de Tucumán, Tucumán, Argentina. Las colecciones estudiadas incluyen la Colección Mamíferos Lillo; el Museo de Ciencias Naturales "Bernardino Rivadavia" (MACN), Capital Federal, Argentina y el Museo Municipal de Ciencias Naturales "Lorenzo Scaglia" (MMPMa), Mar del Plata, Buenos Aires, Argentina. La revisión bibliográfica incluyó todas las publicaciones sobre mamíferos del noroeste argentino desde fines del siglo XIX a la actualidad. Para cada especie se ofrecen datos de especímenes examinados, registros adicionales (Apéndice 3) y se realizan comentarios sobre las localidades de registro tomadas de la bibliografía consideradas dudosas. En el Apéndice 1 se presentan los datos de departamento, provincia y coordenadas geográficas para las localidades de presencia confiable y dudosa. Las coordenadas de las localidades se obtuvieron en el campo, mediante sistemas de posicionamiento global (GPS), y en el laboratorio, a partir de cartas del Instituto Geográfico Militar (IGM) escalas 1:250000 y 1: 500000.
Variables ambientales
Para la construcción de los modelos de distribución se utilizaron ocho variables ambientales que representan características climáticas, topográficas y de vegetación del área de estudio. La resolución espacial de las variables es de 30 arcos segundo, lo cual equivale aproximadamente a 0.72 km² en el área de estudio (Hijmans et al., 2004).
Las variables climáticas utilizadas se obtuvieron de la página web de WORLDCLIM (http://biogeo.berkeley.edu/worldclim/worldclim.html) e incluyeron: Rango Medio Diurno de Temperatura (media mensual de la diferencia entre la temperatura máxima y mínima), Rango Anual de Temperatura (diferencia entre la máxima temperatura del mes más cálido y la mínima temperatura del mes más frío) y Precipitación Media Anual. Las variables topográficas, que incluyeron Altura Sobre el Nivel del Mar y Pendiente, fueron derivadas del modelo de elevación digital producido por la NASA (Shuttle Radar Topographic Mission [SRTM]) disponible en ftp://edcsgs9.cr.usgs.gov/pub/data/srtm. Las variables de vegetación consistieron en dos estimaciones del Índice de Vegetación Verde (Normalized Difference Vegetation Index, NDVI) disponible en http://edcdaac.usgs.gov/1KM/1kmhomepage.htlm. Este índice es una medida que refleja la actividad fotosintética de la vegetación (Lillesand y Kiefer, 2000) y sus valores fluctúan entre -1 y 1. Los valores positivos indican áreas con alta cobertura de la vegetación y los valores negativos indican áreas sin vegetación o cubiertas de nubes. Se utilizaron las siguientes variables calculadas sobre valores máximos mensuales (sensu Parra et al., 2004): Verde Medio: número de meses en los que cada píxel tiene un valor entre 109 y 150 (Holben, 1986) y Estacionalidad en NDVI: definido como 100-[Máximo NDVI-Mínimo NDVI]/Máximo NVDI (Hurlbert y Haskell, 2003). Finalmente, se incorporó una variable de unidades de vegetación obtenida mediante una clasificación no supervisada de información espectral de imágenes satelitales. Las categorías de vegetación obte nidas fueron: 1) región Chaqueña y Yungas, 2) ecotono entre Pastizales de Neblina y Yungas, 3) Pastizales de Neblina, 4) Prepuna y Monte, 5) ecotono entre Pastizales de Neblina y región Altoandina y 6) región Altoandina y Puna (Fig. 1). Con el fin de evitar redundancia en el valor explicativo de las variables, se seleccionaron sólo aquellas con valores de correlación de Pearson (r) menores de 0.7 para un valor de P = 0.05 (Tabla 1).
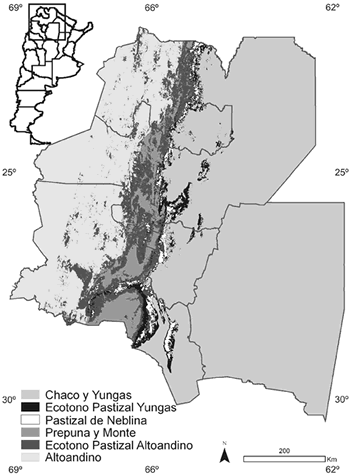
Fig. 1. Distribución de las unidades de vegetación derivada de clasificación de imágenes satelitales para el noroeste argentino usada en la construcción de los modelos de distribución de N. lactens y P. osilae.
Elaboración de los modelos
Distintos modelos han sido utilizados para delimitar las distribuciones de diferentes taxa (Scott et al., 2002), dependiendo la elección de alguno de ellos del objetivo del trabajo, de las características de la distribución de las especies y del tipo de datos disponibles. Para determinar áreas de distribución fuera del rango conocido y con pocos datos de ausencias, como en nuestro caso, el modelo más adecuado es DOMAIN (CIFOR, 1996), ya que trabaja sin datos de ausencia y es especialmente eficiente en modelar distribuciones con alta sensibilidad ya que comete pocos errores de clasificación de presencias verdaderas (Segurado y Araujo, 2004). Además, DOMAIN evita algunos de los problemas encontrados en modelos que tratan las variables climáticas como independientes y que por lo tanto sobreestiman las distribuciones (Carpenter et al., 1993).
En consecuencia, los mapas de distribución potencial se realizaron con el programa DOMAIN, el cual se basa en datos de presencia para definir el espacio ambiental que condiciona la distribución de las especies. El programa utiliza una medida de similitud entre puntos de presencia para asignar valores de clasificación a puntos "incógnita" sobre la base de su proximidad en el espacio ambiental con respecto al sitio con registro de presencia más cercano (Carpenter et al., 1993). DOMAIN realiza la cuantificación de la similitud entre sitios mediante la medida de Gower (Gower, 1971) y genera una matriz continua de valores de similitud que puede visualizarse como un mapa temático de contorno en escala de grises. Los valores de contorno generados por DOMAIN no son entonces valores de probabilidad sino grados de similitud de la clasificación, en los que se establecen umbrales de corte que reflejan la distribución de las especies. El umbral de corte que mejor representa la distribución de una especie puede determinarse objetivamente mediante la valoración de la eficiencia general del modelo (Segurado y Araujo, 2004).
Debido a que en la literatura existen datos de presencia dudosos en cuanto a su identificación taxonómica y/o localización geográfica, se elaboraron dos mapas de distribución para cada especie. El primer mapa se construyó a partir de la selección al azar del 70% de los datos de presencias confiables (18 registros para Necromys lactens y 32 para Phyllotis osilae). El segundo se desarrolló con este 70% de los datos más los datos de presencia dudosos (21 registros para Necromys lactens y 37 para Phyllotis osilae). El 30% restante de las localidades de presencia confiable (8 registros para Necromys lactens y 13 para Phyllotis osilae) se utilizaron en la validación de los modelos, aspecto que se desarrolla en detalle más abajo. Debido a que existen pocos datos de presencia para ambas especies, la proporción de registros utilizados en la construcción y validación de los modelos sigue el criterio de maximizar el número de puntos para validación sin reducir significativamente el número de datos para construcción.
Validación de los modelos
La validación de los modelos implica el contraste de los mismos con información de distribución que incluye registros de presencia y ausencia "confiable" no utilizados en la construcción del modelo. Estos registros se indican en los Apéndices 1 y 2 respectivamente, presentando ambas el mismo tratamiento en cuanto a los datos aportados.
La elección de las localidades de presencia para validación se realizó substrayendo, aleatoriamente, el 30% de los datos de presencia confiables (8 registros para N. lactens y 13 para P. osilae). La ausencia verdadera de un taxón en una localidad determinada es difícil de garantizar. Sin embargo, localidades bien relevadas pueden considerarse como localidades de ausencia confiable. Es entonces que estas localidades se eligieron por haber sido relevadas por diversos autores en más de una oportunidad, por haber sido estudiadas por un autor con un esfuerzo de muestreo considerable o porque son localidades para las que se ha registrado un gran número de ejemplares, lo cual aumenta las posibilidades de registrar todos los taxones presentes en la región. Las ausencias verdaderas incluyeron 27 localidades para los modelos de N. lactens. En el caso de P. osilae se utilizaron 25 localidades para el modelo confiable y 22 para el dudoso. Estas localidades están ampliamente distribuidas en el noroeste argentino (entre 22° 5' y 28° 51' de latitud S y 62° 47' y 66° 19' de longitud W) y ocupan gran parte del gradiente altitudinal (entre los 400 y 4000 m de altitud) y de los ambientes presentes en la región (Chaco Seco, Prepuna-Monte, Puna y todos los pisos altitudinales de Yungas). La fuente de información para estas localidades de ausencia incluye diversas publicaciones sobre mamíferos para la región (Mares et al., 1981; Ojeda y Mares, 1989; Heinonen y Bosso, 1994; Capllonch et al., 1997; Mares et al., 1997; Díaz, 1999; Díaz et al., 2000; Gil y Heinonen Fortabat, 2003), revisión de colecciones y datos propios aún no publicados.
Para evaluar la eficiencia de los mapas de distribución generados, utilizamos los siguientes indicadores: Kappa (medida general de eficiencia del modelo, evalúa en qué medida la ocurrencia predicha es distinta a la debida al azar), Poder Predictivo Positivo (probabilidad de que una predicción de presencia sea correcta), Clasificaciones Correctas (proporción de presencias y ausencias bien predichas con relación al total de las predicciones), error de comisión (el modelo predice que la especie está presente cuando en realidad está ausente), error de omisión (el modelo predice que la especie está ausente cuando en realidad está presente), especificidad (capacidad de predecir ausencias) y sensitividad (capacidad de predecir presencias). Las cualidades de estos indicadores se resumen en el trabajo de Fielding y Bell (1997) y sus valores varían entre 0 y 1. La elección del umbral de corte que mejor representa la distribución se realizó sobre la valoración del desempeño del índice Kappa.
Análisis espaciales
Utilizando técnicas de Sistemas de Información Geográfica (SIG) y las extensiones "Geoprocessing" y "Spatial Analyst" del Programa ArcGIS (v 9) (ESRI, 2005), se determinó el porcentaje del área de distribución representada en cada piso altitudinal y en cada tipo de vegetación. Para esto se realizó la intersección entre el área de distribución predicha por los modelos con el modelo de elevación digital y el mapa de unidades ambientales representadas en la Fig. 1.
RESULTADOS
Registros de distribución
Necromys lactens Thomas, 1918
Los registros de presencia obtenidos para la especie se resumen en el Apéndice 3. En el mismo se discriminan los materiales examinados de los registros adicionales obtenidos de la literatura. La mayoría de los registros obtenidos para esta especie provienen de Pastizales de Neblina y áreas ecotonales en la cabecera de las Yungas. Con muy pocas excepciones (que en su mayoría son consideradas dudosas y se discuten más abajo), las citas ocupan altitudes entre los 1500 m en zonas de ecotono de Pastizales de Neblina y el Bosque Montano (sensu Brown et al., 2001), hasta más de 3000 m en áreas de transición entre estos pastizales y ambientes semiáridos como el desierto de Monte o con pastizales de mayor altura como los ambientes altoandinos.
Aunque el estatus de Akodon orbus Thomas, 1919, es considerado dudoso (Pardiñas et al., en prensa), la descripción original de Thomas y algunas características observables en el cráneo del holotipo hacen suponer que es un sinónimo de N. lactens. Esta opinión ya fue adoptada por otros autores (Cabrera, 1961; Reig, 1987) y, por lo tanto, en este trabajo se incluye la localidad en la construcción del modelo confiable. Debe tenerse en cuenta, sin embargo, que estudios taxonómicos más profundos pueden modificar el estatus de este taxón.
Mares et al. (1997) citaron a esta especie (como Bolomys lactens) para Laguna Blanca, 3100 m, en la Puna de la provincia de Catamarca. Esta cita se basó en ejemplares de Akodon leucolimnaeus Cabrera 1926, especie considerada un sinónimo de N. lactens por algunos autores (Gyldenstolpe, 1932; Cabrera, 1961; Reig, 1987). Sin embargo, la asignación de estos ejemplares a Necromys fue cuestionada por Galliari y Pardiñas (1995) quienes examinaron los ejemplares y los consideraron pertenecientes al grupo boliviensis del género Akodon y, por lo tanto, la localidad mencionada no fue incluida en este análisis.
Otros tres registros pueden ser considerados no confiables y sólo fueron incluidos en la construcción del modelo dudoso. Ortiz et al. (2000) citaron a Necromys lactens para Barranca Larga sobre la base de dos ejemplares obtenidos de egagrópilas de lechuzas. Aunque esta localidad se encuentra en un ambiente caracterizado por estos autores como de Monte-Prepuna (correspondientes a la ecorregión Monte de Sierras y Bolsones), existen parches pequeños de Pastizales de Neblina en los alrededores de donde podrían provenir los ejemplares. Debido a que no se tiene la seguridad de la localidad exacta de procedencia de los ejemplares, este registro sólo se incluyó en la construcción del modelo dudoso. Además, en la vertiente occidental de los principales cordones montañosos de la región, sobre la que se encuentra la localidad de Barranca Larga, se ha registrado otra especie de Necromys que no puede ser fácilmente diferenciada de N. lactens mediante materiales preservados en regurgitados de lechuza y, por lo tanto, existen dudas sobre su pertenencia a N. lactens. La localidad de Las Paras (= Las Pavas) resulta dudosa por encontrarse a una altura nunca registrada para esta especie en el noroeste argentino (aunque la especie se conoce para una localidad a 4000 m en Cochabamba, Bolivia). El ambiente de esta localidad se caracteriza por suelos desnudos (pertenecientes a la ecorregión Altos Andes) y resulta por ello atípico para la especie. Adicionalmente, el registro es impreciso debido a que la altura de la localidad fue mencionada de forma solo aproximada por Thomas (1926). Por último, la localidad de Calilegua, mencionada por Díaz (1999), resulta dudosa por ser el único registro de distribución de la especie por debajo de los 1500 m de altitud. Esta localidad se encuentra en el piso altitudinal inferior del bosque de Yungas, aproximadamente mil metros por debajo de los registros de menor altitud para la especie. A pesar de ser un ambiente bien estudiado por los mastozoólogos en las últimas décadas, con la única excepción de este registro, la especie nunca fue documentada en este piso altitudinal. Debido a que N. lactens fue colectada en localidades que se encuentran sobre o en las adyacencias del Cerro Calilegua (Cerro Hermoso y El Duraznillo), no descartamos la posibilidad de que se trate de un error en la trascripción del nombre de la localidad.
Phyllotis osilae J. A. Allen, 1901
Los registros de presencia obtenidos para la especie se resumen en el Apéndice 3. En el mismo se discriminan los materiales examinados de los registros adicionales obtenidos de la literatura.
Esta especie presenta la mayoría de sus registros en Pastizales de Neblina y áreas ecotonales, aunque P. o. nogalaris, una subespecie exclusiva de Argentina, ha sido documentada también en ambientes boscosos en el piso altitudinal superior de las Yungas. El rango altitudinal de los registros obtenidos comprende entre los 1300 y 3200 m.
Son cinco los registros considerados dudosos para esta especie, todos de ambientes atípicos para la especie. El registro de Chumbicha, en el Chaco Serrano (perteneciente a la ecorregión del Chaco Seco), citado por Pearson (1958), ya fue puesto en duda por Mares et al. (1997). El resto de los registros han sido mencionados por Díaz (1999) para la Puna y Prepuna. Díaz (1999) citó a P. o. osilae para Casa Colorada, 2 km al N de Alfarcito, y a P. o. nogalaris para la región de Abra Pampa (CML 1259, 1276, 1280 y 1283). Estos registros extienden la distribución conocida de la especie a localidades ubicadas notoriamente fuera de su hábitat tradicional. En este estudio se examinaron tres de los ejemplares de Abra Pampa (CML 1276, 1280 y 1283) citados por Díaz (1999) como P. o. nogalaris, y que en nuestra opinión corresponden a P. xanthopygus. Phyllotis o. nogalaris es una subespecie de gran tamaño y sólo se conoce para las Yungas. En estudios en curso se compararon medidas craneanas de P. o. nogalaris, incluyendo especímenes estudiados por Pearson (1958), especímenes colectados en Sierra del Centinela (este de la provincia de Jujuy) inequívocamente asignables a la subespecie nogalaris y aquellos de la CML citados por Díaz (1999). Claramente los ejemplares de la CML, todos ellos adultos viejos, son conspicuamente más pequeños comparados con P. o. nogalaris. Además, éstos presentan muchas de las características de P. xanthopygus tales como fosetas posteropalatales ubicadas muy por delante del borde anterior de la fosa mesopterigoides, rostro estrecho, orejas cubiertas con pelos de tinte anaranjado y vibrisas largas y abundantes. Ninguno de estos ejemplares muestra las características morfológicas de P. osilae nogalaris reseñadas por Pearson (1958). Debido a que no se examinaron todos los ejemplares mencionados por Díaz (1999), no podemos descartar definitivamente la presencia de esta especie para algunas localidades de la Prepuna y Puna, y por ello se tratan como dudosas.
Modelos de distribución
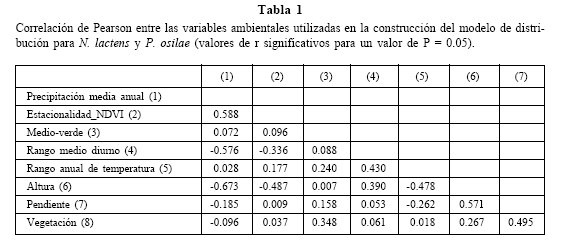
Se obtuvieron dos mapas de distribución potencial para cada especie. Uno de ellos es el resultado de los datos de presencias confiables (Fig. 2A y Fig. 3A) y el otro, es resultado de estos datos más los datos de presencias considerados dudosos (Fig. 2B y Fig. 3B).

Fig. 2. Mapas de distribución potencial de N. lactens. A. Modelo construido sobre la base de datos confiables (umbrales de corte de DOMAIN: 80-82, 82-90 y 90-100). B. Modelo construido sobre la base de datos confiables y dudosos (umbrales de corte de DOMAIN: 90-95 y 95-100).
Fig. 3. Mapas de distribución potencial de P. osilae. A. Modelo construido sobre la base de datos confiables (umbrales de corte de DOMAIN: 84-90 y 90-100). B. Modelo construido sobre la base de datos confiables y dudosos (umbrales de corte de DOMAIN: 90-95 y 95-100).
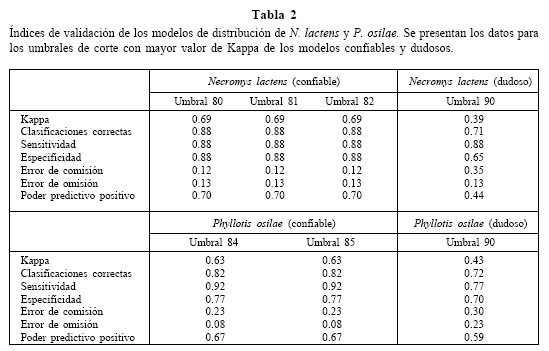
La elección del umbral de corte para los mapas generados se realizó privilegiando los valores más altos del índice Kappa. En el modelo confiable de N. lactens, los umbrales de corte desde 80 a 82 tuvieron el mismo valor de Kappa (y otros indicadores de validación) y por lo tanto se informan los resultados para l os tres valores de corte involucrados. Para P. osilae los umbrales de corte con mayor valor de Kappa en el modelo confiable fueron el de 84 y 85. En el caso de las distribuciones dudosas, el mayor valor de Kappa para ambas especies se obtuvo con el umbral de corte de 90 (Tabla 2, Figs. 2 y 3).
Los modelos confiables de ambas especies siempre presentaron valores mayores de Kappa, con la excepción del umbral de corte de 85 en el que N. lactens muestra un valor más bajo (Fig. 4). Los errores de comisión y omisión para los modelos confiables de las dos especies son relativamente bajos en todos los umbrales de corte analizados y los dos alcanzan su valor mínimo en el nivel de Kappa máximo. Para umbrales de corte mayores con respecto al del Kappa máximo (82 para N. lactens y 84 para P. osilae), el error de omisión aumenta notoriamente debido a que se restringe el área de distribución dejando fuera sitios de presencia verdadera (Fig. 5A). En comparación a los modelos confiables, en los modelos dudosos el error de comisión es siempre más elevado (Fig. 5B), debido a que estos modelos exageran el área de distribución e incluyen sitios de ausencia verdadera.
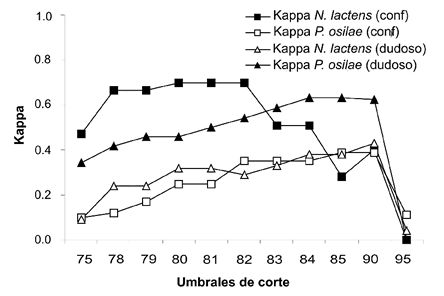
Fig. 4. Valores de Kappa calculados para los modelos basados en datos confiables y dudosos de N. lactens y P. osilae.
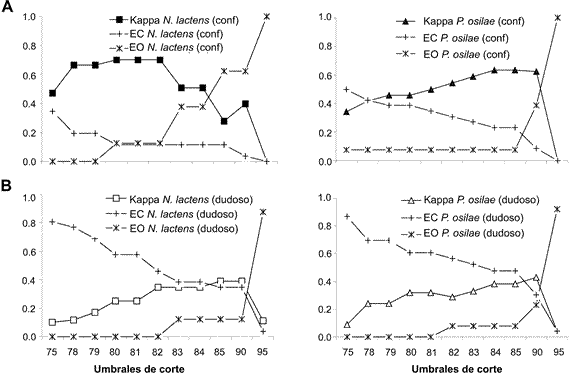
Fig. 5. Valores de Kappa y errores de omisión (EO) y de comisión (EC) para distintos umbrales de corte. A: Modelos confiables para N. lactens y P. osilae. B. Modelos dudosos para N. lactens y P. osilae.
En la Tabla 2 se resumen los valores de los índices utilizados en la validación de los modelos de presencias confiables y dudosas para las dos especies en sus umbrales de corte de mayor valor de Kappa. Los modelos de presencias confiables de ambas especies muestran valores de Kappa, PPP, Clasificaciones Correctas y Especificidad más elevados mientras que los errores de comisión y omisión son bajos. El valor de sensitividad es elevado para los dos tipos de modelos debido a que los de presencias dudosas exageran el área de distribución y por lo tanto predicen bien todas las presencias. Esta sobreestimación de la distribución eleva el valor del error de comisión y disminuye el de omisión.
Los modelos de distribución confiable son similares para ambas especies y muestran una distribución potencial insularizada, restringida a zonas de altura en la cabecera de la Cordillera Oriental, las Sierras Subandinas, las Sierras Centrales y las Sierras Pampeanas. Los modelos predicen distribuciones principalmente restringidas a Pastizales de Neblina lo cual coincide con nuestra predicción. Sin embargo, al sur de la provincia de Jujuy el modelo extiende la distribución de las especies fuera de los Pastizales de Neblina e incluye el valle de cultivos entre los ríos Grande y Perico, así como algunos sectores de Yungas sobre la ruta 9 entre las ciudades de San Salvador de Jujuy y Salta, y sobre las Sierras del Castillejo al SE de la ciudad de Salta.
La distribución obtenida para P. osilae es, en general, más amplia que la de N. lactens, debido a su mayor extensión sobre zonas de ecotono entre Pastizales de Neblina y la región Altoandina en las provincias de Salta y Catamarca.
El modelo dudoso de las dos especies predice, contrariamente, una distribución de grandes áreas continuas que ocupan los sistemas de sierras mencionados y se extienden a ambos lados de los mismos. En el caso de N. lactens, abarca todos los pisos altitudinales de Yungas, además de extensas zonas del Chaco Seco en toda la región y zonas de altura caracterizadas por ambientes de Prepuna-Monte y Puna (Fig. 2B). En tanto, para P. osilae se extiende principalmente sobre ambientes altoandinos, de Puna y Prepuna en la ladera occidental (sobre todo en la provincia de Jujuy y menos en Catamarca) y ambientes chaqueños (principalmente en las provincias de Catamarca y Santiago del Estero) en el extremo sur de la región (Fig. 3B).
De acuerdo con los mapas de distribución confiables obtenidos, P. osilae ocupa una superficie de 3.8 millones de hectáreas, mientras que el área de distribución de N. lactens representa 2.8 millones de hectáreas. Para ambas especies, más del 75% de la distribución predicha se encuentra entre los 1200 y los 3200 m con un pico de ocurrencia entre los 1400 y los 1600 m (Fig. 6). El 60% de la distribución de N. lactens ocurre en Pastizales de Neblina y áreas ecotonales (ecotonos Pastizales-Yungas y Pastizales-Altos Andes de la Fig. 1) y el 40% restante corresponde a la zona del valle de cultivos entre los Ríos Grande y Perico. Al igual que en la especie anterior, hay un alto porcentaje de la distribución de P. osilae (53%) en pastizales y áreas ecotonales, un 40% corresponde a áreas de Yungas del valle de Jujuy y el 7% restante al Monte y Prepuna. La distribución predicha por el modelo para N. lactens es más restringida y se superpone en un 80% con el área de P. osilae. El área de superposición de las dos especies corresponde en un 64% a la zona de Pastizales de Neblina de los cuales el 43% corresponde a Pastizales de Neblina estrictos (Pastizales de Neblina de la Fig. 1).

Fig. 6. Porcentaje del área de distribución de N. lactens y P. osilae en el gradiente altitudinal.
DISCUSIÓN
La aplicación de modelos de distribución de especies en la elaboración de hipótesis de distribución ha crecido rápidamente en los últimos años. El desarrollo de nuevas tecnologías que permiten el análisis de grandes cantidades de datos espaciales, como los Sistemas de Información Geográfica, nuevos paquetes estadísticos y la accesibilidad creciente a bases de datos globales y programas específicos para análisis de distribución, han dado como resultado la proliferación de numerosos trabajos orientados a predecir la presencia de especies a diversas escalas espaciales (e.g., Debinski et al., 2002; Dreisbach et al., 2002; Fertig y Reiners, 2002; van Manen et al., 2002). Estas técnicas permiten hipotetizar el área de distribución de una especie para regiones amplias sin un muestreo intensivo de la misma, lo cual incrementa la eficiencia en la utilización de los recursos.
La construcción de los modelos confiables para ambas especies se realizó sobre la base de relativamente pocos registros de presencia (sobre todo en el caso de N. lactens). Sin embargo, los datos de distribución representan el gradiente altitudinal y latitudinal de la región y los modelos obtenidos son una buena aproximación respecto de nuestra experiencia de campo, con valores de validación (e. g. índice Kappa) comparativamente muy buenos (Fielding y Bell, 1997). Además, la construcción de modelos de distribución de especies con distribuciones restringidas y requerimientos de hábitat específicos como las aquí tratadas, muestran valores bajos de error general (Kappa) (Segurado y Araujo, 2004), por lo que pueden construirse modelos sobre la base de relativamente pocos datos de presencia.
El número de localidades de ausencia confiables utilizadas en la validación es el mínimo recomendado por algunos autores (B. Loiselle, com. pers.). Aunque un número mayor podría mejorar nuestra evaluación de los modelos, en el noroeste argentino es difícil encontrar localidades para las que se hayan realizado muestreos sistemáticos en distintas épocas del año y que por lo tanto presenten series representativas de la mayoría de las especies presentes. Sin embargo, existe información no exhaustiva de distribución proveniente de la literatura y de trabajos de campo que puede dar cuenta de la ausencia de las dos especies estudiadas en ambientes distintos a Pastizales de Neblina y sus ecotonos. De un total aproximado de 250 localidades "extra pastizales" examinadas para todas las unidades de vegetación del noroeste argentino, en ninguna de ellas (excepto los pocos registros mencionados y discutidos en el tratamiento de cada especie), se registraron los taxones tratados. Por el contrario, de 50 localidades de Pastizales de Neblina y áreas ecotonales analizadas, N. lactens estuvo presente en 26 y P. osilae en 45. Todas aquellas localidades de Pastizales de Neblina en las que estas especies no fueron registradas presentan muy pocos ejemplares de sigmodontinos capturados (en algunos casos solo uno o dos) y muchas de ellas se encuentran contiguas a localidades en las que sí se detectaron a partir de muestreos intensivos, sugiriendo que su presencia, en dichos sitios, es altamente probable.
En la elaboración de los modelos se utilizaron ocho variables ambientales relacionadas con temperatura, precipitación, topografía e índices verdes de vegetación. Muchas de estas variables son indicadoras de diferencias entre ambientes desérticos o semidesérticos y ambientes húmedos (como los rangos de temperaturas y los valores de precipitación), de altitud y posición latitudinal (como las variables asociadas con la temperatura), asociadas con el escurrimiento del suelo (éste es el caso de la pendiente de las laderas) e indicadores del tipo de vegetación (índices verdes). Aunque este conjunto de variables permite predecir aceptablemente el área de distribución potencial de ambas especies, nueva información ambiental seguramente mejorará la capacidad de predicción de los mismos.
Aunque los resultados obtenidos muestran que ambas especies se caracterizan por una distribución principalmente restringida a los Pastizales de Neblina y áreas ecotonales, existen sectores en los que las distribuciones se extienden algo más de lo esperado de acuerdo con nuestra experiencia de campo. Esto se observa principalmente en regiones de baja altitud en las Yungas y zonas de transición con el Chaco entre los ríos Grande y Perico, en la provincia de Jujuy, y en sectores adyacentes a esta última región, en la provincia de Salta. La predicción de presencia para estas especies en estas regiones posiblemente responda a que están ocupadas por cultivos y ambientes degradados que presentan un comportamiento similar a los Pastizales de Neblina en cuanto a sus valores de NDVI, y por este motivo el modelo los detecta como áreas ambientalmente apropiadas para la ocurrencia de las especies. Las predicciones de presencia en estas zonas se encuentran, sin embargo, dentro de valores de umbral de similitud inferiores a 85 para la mayoría de los sectores involucrados.
La poca eficiencia general de los modelos dudosos y la ausencia de N. lactens y P. osilae en una gran cantidad de localidades extra-pastizales nos permite suponer que los datos inicialmente considerados dudosos efectivamente consisten en errores en la identificación de las especies o en la localización precisa del sitio de colecta, y que para la elaboración de estos modelos es importante el control de calidad de los registros utilizados.
Aunque las distribuciones obtenidas en este trabajo constituyen hipótesis que deberán ser contrastadas con nuevos datos de campo que las corroboren, son la mejor aproximación con la que contamos hasta el momento y constituyen una herramienta objetiva para elaborar planes de conservación. Modelos de distribución obtenidos para otras especies de la región pueden constituir la base sobre la que se elaboren planes de manejo y conservación que tengan en cuenta características areográficas importantes de las especies como el área total, forma y disposición espacial de los parches ocupados por un taxón, así como también áreas de máxima diversidad y endemismos para la región.
CONCLUSIONES
En este trabajo se destacan dos conclusiones principales. En primer lugar, que las hipótesis de distribución elaboradas mediante datos confiables presentan indicadores de validación que las califican como altamente confiables y son notoriamente más eficientes que aquellas generadas a partir de registros dudosos. Representan, además, una aproximación contrastable y de una muy buena resolución espacial. En segundo término, que los resultados obtenidos indican que ambas especies se encuentran restringidas a las zonas de altura (entre 1500 y 3000 m aproximadamente) de los principales cordones montañosos de la región y delimitan un patrón de distribución insularizado asociado a los Pastizales de Neblina y áreas ecotonales.
Lista de localidades de presencias confiables y dudosas utilizadas para la construcción y validación de las hipótesis de distribución. Entre paréntesis se ofrecen el departamento y la provincia correspondiente a cada localidad, seguido de las coordenadas de la misma. Con asteriscos se indican las localidades utilizadas en la validación, correspondiendo un asterisco para N. lactens, dos para P. osilae y tres para las localidades de validación de ambas especies.
. 1 mi W León, 5800 ft (Doctor Manuel Belgrano, Jujuy) 24° 02' 07.5'' S; 65° 26' 52.5'' W.
. 1 km ENE de Rodeo Pampa, km 59 de Ruta Provincial Nº 7, 3080 m. (Santa Victoria, Salta) 22º 14' 47.7'' S; 65º 03' 4.3'' W. **
. 6 km SW Hwy 9 on Hwy 18, 1524 m. (Ambato, Catamarca) 27º 52' S; 65º 48' W. **
. 9 km NW Bárcena (Tumbaya, Jujuy) 23º 57' S; 65º 30' W. ***
- Bárcena, 9 km NW, 2655 m. (Tumbaya, Jujuy) 23º 57' S; 65º 30' W.
. Above Tafí del Valle, 9500 ft. (Tafí del Valle, Tucumán) 26° 45' 19'' S; 65° 44' 39'' W.
. Abra de Ciénaga Negra, aproximadamente 3 km al SE, 3090 m (Orán, Salta) 23º 19' 49'' S; 64º 53' 32'' W
. Abra Pampa (Cochinoca, Jujuy) 22° 43' S; 65° 42' W.
- La Ciénaga, 3700 m (Cochinoca, Jujuy) 22° 43' S; 65° 42' W.
- La Ciénaga, Abra Pampa (Cochinoca, Jujuy) 22° 43' S; 65° 42' W.
. Agua del Gauchi, aproximadamente 1 km al E del Mogote Los Cocos, 2024 m (Capayán, Catamarca) 28º 46' 52'' S; 66º 18' 43'' W.
. Alto Calilegua, El Duraznillo, 3000 m (Valle Grande, Jujuy) 23º 35' S; 64º 55' W.
- Cerro Calilegua, El Duraznillo, 2600 m (Valle Grande, Jujuy) 23º 35' S; 64º 55' W.
- El Duraznillo, Cerro Calilegua, 2600 m. (Valle Grande, Jujuy) 23º 35' S; 64º 55' W.
. Aproximadamente 2 km al SE de Huaico Hondo, sobre Ruta Provincial Nº 42, al E del Portezuelo, 1992 m (Valle Viejo, Catamarca) 28º 25' 10.9'' S; 65º 32' 40.5'' W. *
. Aproximadamente7 km al NO de la finca de la familia Usandivara, Altos de Medina, 1717 m. (Burruyacú, Tucumán) 22º 23' 13.7'' S; 65º 05' 21.5'' W. **
. Aproximadamente 10 km al E de Los Varela, sobre Ruta Provincial Nº 4, Sierra de Humaya, 2006 m. (Ambato, Catamarca) 27º 56' 12.1'' S; 65º 56' 52.8'' W. **
. Aproximadamente 10 km al S de Hualinchay, sobre el camino a Lara, 2316 m (Trancas, Tucumán) 26º 19' 20.2'' S; 65º 36' 45.5'' W.
. Aproximadamente 15 km al W de Escoipe, sobre Ruta Provincial Nº 33, 2680 m (Chicoana, Salta) 25º 10' S; 65º 49' W. *
- Aproximadamente 15 km al W de Escoipe, sobre Ruta Provincial Nº 33 (Chicoana, Salta) 25º 10' S; 65º 49' W.
. Aproximadamente 16 km de Hualinchay, sobre el camino a Lara, 2750 m (Trancas, Tucumán) 26º 19' 31.9'' S; 65º 38' 51.2'' W.
. Arroyo La Horqueta, 3 km al SE de Lagunas de Yala, 2100 m (Doctor Manuel Belgrano, Jujuy) 24° 7' S; 65° 27' W.
. Bárcena, aproximadamente 3 km al S, sobre Ruta Nacional Nº 9, 1808 m (Tumbaya, Jujuy) 24º 00' 02.0'' S; 65º 26' 51.6'' W. *
. Barranca Larga, 2400 (Belén, Catamarca) 26° 59' 55.5'' S; 66° 43' 34.5'' W.
. Calilegua (Ledesma, Jujuy) 23° 46' 34.5'' S; 64° 46' 16.5'' W.
. Campo Quijano, aproximadamente 5 km al NO, km 30 de la Ruta Nacional Nº 51 (Quebrada del Toro), altura aproximada 1600 m (Rosario de Lerma, Salta) 24º 53' S; 65º 40' W.
. Carapunco, km 81 de la Ruta Provincial N° 307, 2960 m. (Tafí del Valle, Tucumán) 26° 45' 19'' S; 65° 44' 39'' W.
. Casa Colorada, app 2 km al N de Alfarcito, 3034 m (Tilcara, Jujuy) 23° 35' S; 65° 21' W.
. Cerro El Manchao, Los Cajones, 4280 m (Pomán, Catamarca) 28° 16' S; 66° 02' W. **
. Cerro Hermoso (Ledesma, Jujuy) 23º 34' S; 64º 51' W.
- Cerro Hermoso (cercanías), 2800 m (Ledesma, Jujuy) 23º 34' S; 64º 51' W.
. Cerro San Javier (sin localidad específica).
. Chilcayoc (Tumbaya, Jujuy) 23º 56' S; 65º 28' W.
. Chumbicha, 488 (Capayán, Catamarca) 28° 51' 07.5'' S; 66° 14' 31.5'' W.
- Chumbicha, 1600 ft (Capayán, Catamarca) 28° 51' 07.5'' S; 66° 14' 31.5'' W.
. Cuesta del Obispo, aproximadamente 5 km al NE de Piedra del Molino, sobre Ruta Provincial Nº 33, 3174 m. (Chicoana, Salta) 25º 10' 36'' S; 65º 51' 04'' W.
. Cumbre de Mala-Mala, 10800 ft (sin localidad específica).
. El Corralito, aproximadamente 23 Km al SO de Campo Quijano, sobre ruta Nac. Nº 51 (Rosario de Lerma, Salta) 24º 58' S; 65º 48' W. *
. El Rodeo, 1.5 km NE Hwy 4, el: 4500 ft (Ambato, Catamarca) 28° 10' 48'' S; 65° 52' 12'' W.
. Estancia Narvaez, 5,5 Km N Las Chacritas en Ruta Provincial Nº 1, 1782 m. (Ambato, Catamarca) 27º 39' S; 65º 56' W.
. Higuerilla (Valle Grande, Jujuy) 23° 36' S; 65° 05' W. **
- Higuerilla, 2000 m (Valle Grande, Jujuy) 23° 36' S; 65° 05' W.
- Higuerilla, 6600 ft (Valle Grande, Jujuy) 23° 36' S; 65° 05' W.
. Hualinchay, 1,9 km NE, en Ruta Provincial Nº 311, 1732 m (Trancas, Tucumán) 28º 18' S; 65º 35' W. **
- Hualinchay, 2,2 Km NE, en Ruta Provincial Nº 311, 1732 m. (Trancas, Tucumán) 28º 18' S; 65º 35' W.
. Hualinchay, sobre el camino a Cafayate, 1861 m (Trancas, Tucumán) 26º 18' S; 65º 36' W.
. Km 33 de la Ruta Provincial Nº 47, al S de Capillitas, 2500 m (Andalgalá, Catamarca) 27º 25' S; 66º 25' W.
. La Angostura (Tafí del Valle, Tucumán) 26° 56' 25.5'' S; 65° 42' 10.5'' W. *
. La Antena, Sierra del Centinela, al S de El Fuerte, 2350 m. (Santa Bárbara, Jujuy) 24º 17' 56.5'' S; 64º 23' 09.3'' W. **
. La Ciénaga (=Ciénaga?), 8200 ft (Tafí del Valle, Tucumán) 26° 46' S; 65° 39' W. **
. Las Chacritas, aproximadamente 28 km al NNO de Singuil, sobre Ruta Provincial Nº 1, 1888 m (Ambato, Catamarca) 27º 42' 24.2'' S; 65º 54' 40.6'' W. *
. Las Cuevas (Belén, Catamarca) 27° 02' S; 66° 39' W.
. Las Paras, about 4000 m (Chicligasta, Tucumán) 27° 08' 24'' S; 65° 59' 24'' W.
. Las Paras, 3018 (Chicligasta, Tucumán) 27° 08' 24'' S; 65° 59' 24'' W.
- Las Paras, 9900 ft (Chicligasta, Tucumán) 27° 8' 24'' S; 65° 59' 24'' W.
. León (Doctor Manuel Belgrano, Jujuy) 24º 02' 07.5'' S; 65º 26' 01.5'' W.
- León, 1500 m (Doctor Manuel Belgrano, Jujuy) 24º 02' 07.5'' S; 65º 26' 01.5'' W.
- León, 5000 ft (Doctor Manuel Belgrano, Jujuy) 24º 02' 07.5'' S; 65º 26' 01.5'' W.
. León, 1 mile west, 5800 ft (Doctor Manuel Belgrano, Jujuy) 24º 02' 07.5'' S; 65º 26' 52.5'' W.
. Loma Atravesada, aproximadamente 3 km al NO del Puesto de Leandro Vega, NO de Chumbicha, 1369 m. (Capayán, Catamarca) 28° 47' S; 66° 18' W.
. Miyuyoc, 3750 m (Humahuaca, Jujuy) 22° 51' S; 65° 18' W.
. Mountains W of Yala (sin localidad específica).
- Mountains W of Yala, 10000 ft (sin localidad específica).
. Ñorco, 8200 ft (Trancas, Tucumán) 26° 30' 43.5'' S; 65° 32' 58.5'' W. **
. Otro Cerro, 9900 ft (Capayán, Catamarca) 28° 45' S; 66° 17' W.
- Otro Cerro, 3018 m (Capayán, Catamarca) 28° 45' S; 66° 17' W.
- Otro Cerro, 3000 m (Capayán, Catamarca) 28° 45' S; 66° 17' W.
- 45 km west of Chumbicha, about 3000 m. (Capayán, Catamarca) 28º 45' S, 66º 17' W.
. Pampa Verde, aproximadamente 8 km al OSO de Los Toldos y al S del Cerro Bravo, 2400 m (Santa Victoria, Salta) 22º 17' S; 64º 48' W.
. Rinconada, 6 km al N, camino a Timón Cruz, 4286 m (Rinconada, Jujuy) 22° 25' S; 66° 12' W.
. Río Tiraxi, aproximadamente 5 km al S de Tiraxi, sobre ruta provincial N° 29 (Doctor Manuel Belgrano, Jujuy) 23° 55' S; 65° 21' W.
. Río Vallecito, 2896 m (Belén, Catamarca) 27° 17' 34.5'' S; 66° 4' 1.5'' W.
- Río Vallecito, 9500 ft (Belén, Catamarca) 27° 17' 34.5'' S; 66° 4' 1.5'' W
. Ruta 307, km 72 (Tafí del Valle, Tucumán) 26° 48' 52.5'' S; 65° 43' 25.5'' W.
. SW de Hualinchay, 2822 m (Trancas, Tucumán) 26º 20' S; 65º 39' W.
. Tafí del Valle, 7000 ft (Tafí del Valle, Tucumán) 26º 50' 37.5'' S; 65º 42' 34.5'' W. **
. Tafí del Valle, km 78, 2700 m. (Tafí del Valle, Tucumán) 26º 46' 37.5'' S; 65º 43' 43.5'' W. *
. Termas de Reyes, aproximadamente 15 km al N, sobre Ruta Provincial Nº 4 (Doctor Manuel Belgrano, Jujuy) 24º 08' S; 65º 27' W.
. Termas de Reyes, mirador, sobre Ruta Provincial Nº 4 (Doctor Manuel Belgrano, Jujuy) 24º 11' S; 65º 28' W.
. Unión entre las rutas provinciales Nº 9 y 18, 3.4 km al S, sobre Ruta Provincial Nº 18, 1529 m (Paclín, Catamarca) 27º 49' S; 65º 47' W. **
. Valle Encantado, Los Cardones National Park, 3000 m (Chicoana, Salta) 25° 11' 48.5'' S; 65° 50' 33.9'' W.
Lista de localidades de ausencias confiables utilizadas para la validación de las hipótesis de distribución. Entre paréntesis se ofrecen el departamento y la provincia correspondiente a cada localidad, seguido de las coordenadas de la misma. Sin asterisco se indican las localidades utilizadas en la validación de los modelos de ambas especies, con un asterisco las utilizadas sólo en la validación de los modelos de N. lactens y con dos asteriscos las localidades utilizadas sólo en la validación del modelo confiable de P. osilae.
. 6 km SW Santa Victoria, at "El Breal" (extreme NE Salta) (Rivadavia, Salta) 22° 18' S; 62° 47' W.
. 14 km S jct. Hwys 40 and 51, along Hwy 40, 4000 m (La Poma, Salta) 24° 22' S; 66° 14' W.
. 17 km SW Dique Itiyuro, near Yacuiba, 500 m (Gral. José de San Martín, Salta) 22° 01' S; 63° 43' W.
. 24 km NW Agua Blanca, 400 m (Iruya, Salta) 22° 42' S; 64° 34' W.
. Abra de Cañas, 1700 m (Valle Grande, Jujuy) 23° 40' S; 64° 54' W.
. Abra Pampa, 3500 m (Cochinoca, Jujuy) 22° 43' S; 65° 42' W. **
. Aguaray, 700 m (Gral. José de San Martín, Salta) 22° 16' S; 63° 46' W.
. Aguas Negras, 600 m (Ledesma, Jujuy) 23° 45' S; 64° 56' W.
. Alfarcito, 2600 m (Tilcara, Jujuy) 23° 37' S; 65° 23' W. **
. Andalgalá (Andalgalá, Catamarca) 27° 36' S; 66° 19' W.
. Angosto del río Pescado, 650 m (Orán, Salta) 22° 33' S; 64° 32' W.
. Caimancito, 550 m (Ledesma, Jujuy) 23° 44' S; 64° 36' W.
. Chumbicha, 600 m (Capayán, Catamarca) 28° 51' 07.5'' S; 66° 14' 31.5'' W. **
. Cuesta del Hurón, 29 km al O de Cieneguillas sobre ruta provincial N° 64, 3835 m (Santa Catalina, Jujuy) 22 ° 06' S; 66° 03' W.
. El Simbolar, 25 km al SO de Palma Sola, 1000 m (Santa Bárbara, Jujuy) 24° 11' S; 64° 28' W.
. Finca El Carmen, 396 m (Orán, Salta) 23° 06' 51.1'' S; 64° 15' 03.3'' W.
. Finca Falcón, aproximadamente 3 km al NW del pozo de agua N° 5, cercanías del Río Seco, 704 m (Gral. José de San Martín, Salta) 22° 18' 41.4'' S; 63° 58' 7.1'' W.
. Los Chorrillos, 13 km NO, limite N Estancia Los Chorrillos, 1408 m (Burruyacu, Tucumán) 26° 17' S; 64° 59' W.
. Horco Molle, 650 m (Yerba Buena, Tucumán) 26° 47' 38.4'' S; 65° 18' 57.6'' W.
. Hualinchay, 1,9 km NE, en Ruta Provincial Nº 311, 1732 m (Trancas, Tucumán) 26° 18' 44'' S; 65° 35' 52'' W. *
. Maimará, 2500 m (Tilcara, Jujuy) 23° 37' S; 65° 28' W.
. Mesada de Las Colmenas, 1150 m (Ledesma, Jujuy) 23° 42' S; 64° 52' W. *
. Parque Provincial El Cochuna (Chicligasta, Tucumán) 27° 19' 28.4'' S; 65° 55' 46.5'' W.
. Reserva la Florida, 7 km al O. de Ibatín, sobre Río Pueblo Viejo (Monteros, Tucumán) 27° 12' 10.2'' S; 65° 39' 4.5'' W.
. Tres Cruces, 3700 m (Humahuaca, Jujuy) 22° 55' S; 65° 35' W.
. Yavi Chico, 3600 m (Yavi, Jujuy) 22° 5' S; 65° 28' W.
. Villa Nougues, aproximadamente 2 km al N, sobre Ruta Provincial Nº 338, a la altura de la antena repetidora del Canal 10 de televisión (Lules, Tucumán) 26° 50' 33.9'' S; 65° 22' 59.8'' W.
Registros de presencias obtenidos para Necromys lactens y Phyllotis osilae. Los registros se dividen en especímenes examinados y registros adicionales provenientes de la literatura.
Necromys lactens
Especímenes examinados - CATAMARCA: aproximadamente 2 km al SE de Huaico Hondo, sobre Ruta Provincial Nº 42, al E del Portezuelo, 1992 m (JPJ 419); Las Chacritas, aproximadamente 28 km al NNW de Singuil, sobre Ruta Provincial Nº 1, 1888 m (JPJ 552); unión entre las rutas provinciales Nº 9 y 18, 3.4 km al S, sobre Ruta Provincial Nº 18, 1529 m (JPJ 6). JUJUY: Bárcena, 9 km NW, 2655 m (obs. pers. J. P. Jayat); Bárcena, aproximadamente 3 km al S, sobre Ruta Nacional Nº 9, 1808 m (JPJ 630); Cerro Hermoso (MACN 19.517); El Duraznillo, Cerro Calilegua, 2600 m (CML 1736, 1737); La Antena, Sierra del Centinela, al S de El Fuerte, 2350 m (JPJ 952); León (MMPMa 2158, 2161, 2169, 2185, 2190); Termas de Reyes, aproximadamente 15 km al N, sobre Ruta Provincial Nº 4 (JPJ 123). SALTA: 1 km ENE de Rodeo Pampa, km 59 de Ruta Provincial Nº 7, 3080 m (JPJ 1315); Abra de Ciénaga Negra, aproximadamente 3 km al SE, 3090 m (JPJ 721); aproximadamente 15 km al W de Escoipe, sobre Ruta Provincial Nº 33, 2680 m (JPJ 47); El Corralito, aproximadamente 23 Km al SO de Campo Quijano, sobre ruta Nac. Nº 51 (JPJ 152); Pampa Verde, OSO de Los Toldos y al S del Cerro Bravo, 2400 m (JPJ 361). TUCUMÁN: aproximadamente 10 km al S de Hualinchay, sobre el camino a Lara, 2300 m (JPJ 447); aproximadamente 16 km de Hualinchay, sobre el camino a Lara, 2750 m (JPJ 768); Hualinchay, sobre el camino a Cafayate, 1861 m (JPJ 154); SW de Hualinchay, 2822 m (obs. pers. J. P. Jayat); ruta 307, km 72 (MMPMa 2985); Tafí del Valle, km 78, 2700 m (CML 1836); Tafí del Valle, 2800 m (CML 2062).
Registros adicionales - CATAMARCA: 45 km west of Chumbicha, about 3000 m (Thomas, 1919, como Akodon orbus); Barranca Larga, 2400 m (Ortiz et al., 2000); Laguna Blanca, 3100 m; Otro Cerro, 3000 m (Mares et al., 1997). JUJUY: 9 km NW Bárcena; Alto Calilegua, El Duraznillo, 3000 m; Calilegua; Cerro Calilegua, El Duraznillo, 2600 m; Cerro Hermoso (Díaz, 1999); Cerro Hermoso (cercanías), 2800 m (Heinonen y Bosso, 1994); Chilcayoc; León, 1500 m (Thomas, 1918, como Akodon lactens; Díaz, 1999). SALTA: aproximadamente 15 km al W de Escoipe, sobre Ruta Provincial N° 33 (Díaz et al., 2000); Valle Encantado, Los Cardones National Park, 3000 m (Ortiz et al., 2000). TUCUMÁN: La Angostura (Ortiz y Pardiñas, 2001, como Necromys cf. N. lactens); Las Paras, about 4000 m (Thomas, 1926, como Bolomys negrito).
Phyllotis osilae
Especímenes examinados - CATAMARCA: Agua del Gauchi, aproximadamente 1 km al E del Mogote Los Cocos, 2024 m (JPJ 1228); aproximadamente 10 km al E de Los Varela, sobre Ruta Provincial Nº 4, Sierra de Humaya, 2006 m (JPJ 1312); aproximadamente 2 km al SE de Huaico Hondo, sobre Ruta Provincial Nº 42, al E del Portezuelo, 1992 m (JPJ 412); El Rodeo, 1.5 km NE of Hwy 4, el: 4500 ft (CML 3448); Estancia Narvaez, 5,5 km N Las Chacritas en Ruta Provincial Nº 1, 1782 m (obs. pers. J. P. Jayat); km 33 de la Ruta Provincial Nº 47, al S de Capillitas, 2500 m (JPJ 674); Las Chacritas, aproximadamente 28 km al NNW de Singuil, sobre Ruta Provincial Nº 1, 1888 m (JPJ 567); Loma Atravesada, aproximadamente 3 km al NO del puesto de Leandro Vega, NO de Chumbicha (JPJ 1157); unión entre las rutas provinciales Nº 9 y 18, 3,4 km al S, sobre Ruta Provincial Nº 18, 1529 m (JPJ 19). JUJUY: Barcena, 9 km NW, 2655 m (obs. pers. J. P. Jayat); Bárcena, aproximadamente 3 km al S, sobre Ruta Nacional Nº 9, 1808 m (JPJ 632); La Antena, Sierra del Centinela, al S de El Fuerte, 2350 m (JPJ 933); Termas de Reyes, aproximadamente 15 km al N, sobre Ruta Provincial Nº 4 (JPJ 291); Termas de Reyes, mirador, sobre Ruta Provincial Nº 4 (JPJ 122). SALTA: 1 km ENE de Rodeo Pampa, km 59 de Ruta Provincial Nº 7, 3080 m (JPJ 1313); Abra de Ciénaga Negra, aproximadamente 3 km al SE, 3090 m (JPJ 722); aproximadamente 15 km al W de Escoipe, sobre Ruta Provincial Nº 33, 2680 m (JPJ 51); Campo Quijano aproximadamente 5 Km al NO, Km 30 de la ruta Nacional 51 (Quebrada del Toro), altitud aproximada 1600 msnm (JPJ 91); Cuesta del Obispo, aproximadamente 5 km al NE de Piedra del Molino, sobre Ruta Provincial Nº 33, 3174 m (JPJ 1210). TUCUMÁN: SW de Hualinchay, 2822 (obs. pers. J. P. Jayat); aproximadamente 10 km al S de Hualinchay, sobre el camino a Lara, 2300 m (JPJ 435); aproximadamente 16 km de Hualinchay, sobre el camino a Lara, 2750 m (JPJ 746); aproximadamente 7 km al NO de la finca de la familia Usandivara, Altos de Medina, 1717 m (JPJ 810); Carapunco, km 81 de la Ruta Provincial N° 307, 2960 m (JPJ 1063); Hualinchay, 1,9 km NE, en Ruta Provincial Nº 311, 1732 m (obs. pers. J. P. Jayat); Hualinchay, 2,2 Km NE, en Ruta Provincial Nº 311, 1732 m (obs. pers. J. P. Jayat); Hualinchay, sobre el camino a Cafayate, 1861 (JPJ 173).
Registros adicionales - CATAMARCA: 6 km SW Hwy 9 on Hwy 18, 1524 m (Mares et al., 1997); 45 km west of Chumbicha, about 3000 m (Thomas, 1919); Cerro El Manchao, Los Cajones, 4280 m (Mares et al., 1997); Chumbicha, 1600 ft (Pearson, 1958); Chumbicha, 488; Las Cuevas; Las Paras, 3018 (Mares et al., 1997); Las Paras, 9900 ft (Pearson, 1958); Otro Cerro, 3018 m (Mares et al., 1997); Otro Cerro, 9900 ft (Pearson, 1958); Río Vallecito, 2896 m (Mares et al., 1997); Río Vallecito, 9500 ft (Pearson, 1958). JUJUY: 1 mi. W León, 5800 ft (Pearson, 1958); 9 km NW Bárcena; Abra Pampa; Arroyo La Horqueta, 3 km al SE de Lagunas de Yala, 2100 m; Casa Colorada, app 2 km al N de Alfarcito, 3034 m; Cerro Calilegua, El Duraznillo, 2600 m; Chilcayoc; Higuerilla (Díaz, 1999); Higuerilla, 2000 m (Thomas, 1921; Díaz, 1999); Higuerilla, 6600 ft (Pearson, 1958); La Ciénaga, 3700 m; La Ciénaga, Abra Pampa; León; León, 1 mile west, 5800 ft (Díaz, 1999); León, 1500 m (Thomas, 1918); León, 5000 ft (Pearson, 1958); Miyuyoc, 3750 m; Mountains W of Yala (Díaz, 1999); Mountains W of Yala, 10000 ft (Pearson, 1958; Díaz, 1999); Rinconada, 6 km al N, camino a Timón Cruz, 4286 m; Río Tiraxi, app 5 km al S de Tiraxi, sobre ruta provincial N° 29 (Díaz, 1999). TUCUMÁN: above Tafí del Valle, 9500 ft (Pearson, 1958); Cerro San Javier (Capllonch et al., 1997); Cumbre de Mala-Mala, 10800 ft; La Ciénaga (=Ciénaga?), 8200 ft; Ñorco, 8200 ft; Tafí del Valle, 7000 ft (Pearson, 1958).
AGRADECIMIENTOS
Numerosas personas colaboraron en la realización de este trabajo. D. Miotti, G. Namen, P. Ortiz, E. Martín, J. Torres, D. Juri, K. García, M. E. Mosca, L. Ferre, S. Sabate, O. Díaz, W. Villafañe (Almita), U. Pardiñas, M. E. Fanjul y M. Quoirín colaboraron en las tareas de campo. Agradecemos también a los curadores de las colecciones estudiadas que gentilmente permitieron el acceso a los materiales depositados en las mismas. Parte de estos estudios fueron financiados mediante una beca de formación de postgrado otorgada por el CONICET y un subsidio otorgado por la Fundación ProYungas a uno de los autores (J. P. Jayat). Comentarios realizados por B. Loiselle, H. R. Grau, P. Blendinger, P. Ortiz e I. Gasparri significativamente mejoraron la calidad del manuscrito. Por último, agradecemos los comentarios y sugerencias de los revisores del manuscrito, J. J. Morrone y T. E. Espinosa.
LITERATURA CITADA
ACEÑOLAZA FG y A TOSELLI. 1981. Geología del Noroeste Argentino. Universidad Nacional de Tucumán, Argentina 212 pp. [ Links ]
ANDERSON R, M GÓMEZ-LAVERDE y A PETERSON. 2002. Geographical distributions of spiny pocket mice in South America: insights from predictive models. Global Ecology and Biogeography 11:131-141. [ Links ]
BARQUEZ RM, MA MARES y RA OJEDA. 1991. Mamíferos de Tucumán. Oklahoma Museum of Natural History, Norman, Oklahoma. [ Links ]
BROWN AD, HR GRAU, LR MALIZIA y A GRAU. 2001. Argentina. Pp. 623-659, en: Bosques Nublados del Neotrópico (M Kappelle y A D Brown, eds.). Instituto Nacional de Biodiversidad (INBio), Costa Rica. [ Links ]
BROWN AD y HR GRAU. 1993. La Naturaleza y el Hombre en las Selvas de Montaña. Proyecto GTZ- Desarrollo Agroforestal en Comunidades Rurales del Noroeste Argentino, Salta. [ Links ]
BURKART R, NO BÁRBARO, RO SÁNCHEZ y DA GÓMEZ. 1999. Eco-regiones de la Argentina. APN-PRODIA. Buenos Aires. [ Links ]
CABRERA A. 1926. D os roedores nuevos de las montañas de Catamarca. Revista Chilena de Historia Natural 30:319-321. [ Links ]
CABRERA A. 1961. Catálogo de los mamíferos de América del Sur. Parte II. Revista del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Zoología 4(2):309-732. [ Links ]
CAPLLONCH P, AG AUTINO, MM DÍAZ, RM BARQUEZ y M GOYTIA. 1997. Los mamíferos del Parque Biológico Sierra de San Javier, Tucumán, Argentina: Observaciones sobre su sistemática y distribución. Mastozoología Neotropical 4:49-71. [ Links ]
CARPENTER G, AN GILLISON y J WINTER. 1993. DOMAIN: a flexible modelling procedure for mapping potential distributions of plants and animals. Biodiversity and Conservation 2:667-680. [ Links ]
CIFOR. 1996. DOMAIN V 1.4. Center for International Forestry Research, Bogor, Indonesia. http://www.cgnet.org/cifor. [ Links ]
DEBINSKI DM, ME JAKUBAUSKAS, K KINDSCHER, EH SAVERAID y MG BORGOGNONE. 2002. Predicting meadow communities and species occurrences in the greater Yellowstone ecosystem. Pp. 499-506, en: Predicting species occurrences, issues of accuracy and scale (JM Scott, PJ Heglund, ML Morrison, JB Haufler, MG Raphel, WA Wall y FB Samson, eds.). Island Press. [ Links ]
DIAZ MM. 1999. Mamíferos de la Provincia de Jujuy: Sistemática, distribución y ecología. Tesis doctoral inédita, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Argentina 640 pp. [ Links ]
DIAZ MM, JK BRAUN, MA MARES y RM BARQUEZ. 2000. An update of the taxonomy, systematics, and distribution of the mammals of Salta province, Argentina. Occasional Papers Sam Noble Oklahoma Museum of Natural History, University of Oklahoma 10:1-52. [ Links ]
DÍAZ MM y RM BARQUEZ. 1999. Contributions to the knowledge of the mammals of Jujuy Province, Argentina. The Southwestern Naturalist 44:324-333. [ Links ]
DÍAZ MM y RM BARQUEZ. 2002. L os mamíferos de Jujuy, Argentina. Editorial L O L A, Argentina. [ Links ]
DREISBACH TA, JE SMITH y R MOLINA. 2002. Challenges of modeling fungal habitat: When and where do you find chanterelles? Pp. 475-481, en: Predicting species occurrences, issues of accuracy and scale (JM Scott, PJ Heglund, ML Morrison, JB Haufler, MG Raphel, WA Wall y FB Samson, eds.). Island Press. [ Links ]
ESRI. 2005. ArcGIS (v 9). Environmental Systems Research Institute, Inc. [ Links ]
FERIA T y A PETERSON. 2002. Prediction of bird community composition based on point-occurrence data and inferential algorithms: a valuable tool in biodiversity assessments. Diversity and Distributions 8:49-56. [ Links ]
FERTIG W y WA REINERS. 2002. Predicting presence/absence of plant species for range mapping: A case study from Wyoming. Pp. 483-489, en: Predicting species occurrences, issues of accuracy and scale (JM Scott, PJ Heglund, ML Morrison, JB Haufler, MG Raphel, WA Wall y FB Samson, eds.). Island Press. [ Links ]
FIELDING AH y J F BELL. 1997. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environmental Conservation 24:38-49. [ Links ]
GALLIARI CA y UFJ PARDIÑAS. 1995. La identidad de Akodon leucolimnaeus Cabrera (Rodentia, Sigmodontinae). X° Jornadas Argentinas de Mastozoología, Resúmenes: 28. [ Links ]
GIL G y S HEINONEN FORTABAT. 2003. Lista comentada de los mamíferos del Parque Nacional Baritú (Salta, Argentina). Acta Zoológica Lilloana 47:117-135. [ Links ]
GOWER JC. 1971. A general coefficient of similarity and some of its properties. Biometrics 27:857-874. [ Links ]
GYLDENSTOLPE N. 1932. A manual of Neotropical sigmodont rodents. Kungliga Svenska Vetenskapsakademiens Handlingar 3:1-164. [ Links ]
HEINONEN S y A BOSSO. 1994. Nuevos aportes para el conocimiento de la mastofauna del Parque Nacional Calilegua (Provincia de Jujuy, Argentina). Mastozoología Neotropical 1:51-60. [ Links ]
HIJMANS R, S CAMERON y J PARRA. 2004. WorldClim, version 1.2. A square kilometer resolution database of global terrestrial surface climate. http://biogeo.berkeley.edu/. [ Links ]
HOLBEN BN. 1986. C haracteristics of maximun-value comosite images for temporal AVHRR data. International of Remote Sensing 7:1435-1445. [ Links ]
HURLBERT A y J HASKELL. 2003. The effect of energy and seasonality on avian species richness and community composition. American Naturalist 161:83-97. [ Links ]
LILLESAND TM y R KEIFER. 2000. Remote sensing and image interpretation, 4th edition. John Wiley and Sons, INC. New York, New York. [ Links ]
LOISELLE B, C HOWELL, C GRAHAM, J GOERCK, T BROOKS, K SMITH y P WILLIAMS. 2003. Avoiding pitfalls of using species distribution models in conservation planning. Conservation Biology 17:1591-1600. [ Links ]
MARES MA, RA OJEDA, JK BRAUN y RM BARQUEZ. 1997. Systematics, distribution, and ecology of the mammals of Catamarca Province, Argentina. Pp. 89-141, en: Life among the muses: Papers in honor of James S. Findley (T L Yates, W L Gannon y D E Wilson, eds.). Special Publication, The Museum of Southwestern Biology 3. [ Links ]
MARES MA, RA OJEDA y MP KOSCO. 1981. Observations on the distribution and ecology of the mammals of Salta Province, Argentina. Annals of the Carnegie Museum of Natural History 50:151-206. [ Links ]
MARES MA, RA OJEDA y RM BARQUEZ. 1989. Guía de los mamíferos de la provincia de Salta, Argentina. University of Oklahoma Press, Norman, Oklahoma. [ Links ]
MARTÍNEZ-MEYER E y A NAVARRO-SIGÜENZA. 2003. Evolution of seasonal ecological niches in the Passerina buntings (Aves: Cardinalidae). Proceedings of the Royal Society, London B 271:1151-1157. [ Links ]
OJEDA RA y MA MARES. 1989. A biogeographic analysis of the mammals of Salta province, Argentina. Patterns of species assemblage in the neotropics. Special Publications, The Museum Texas Tech University 27:1-66. [ Links ]
OLROG CC y MM LUCERO. 1981. Guía de los mamíferos argentinos. Ministerio de Cultura y Educación, Fundación Miguel Lillo, Tucumán. [ Links ]
ORTIZ PE y UF PARDIÑAS. 2001. Sigmodontinos (Mammalia, Rodentia) del Pleistoceno tardío del Valle de Tafí (Tucumán, Argentina): taxonomía, tafonomía y reconstrucción paleoambiental. Ameghiniana 38:3-26. [ Links ]
ORTIZ PE, S CIRIGNOLI, DH PODESTÁ y UF PARDIÑAS. 2000. New records of sigmodontine rodents (Mammalia: Muridae) from high-Andean localities of northwestern Argentina. Biogeographica 76:133-140. [ Links ]
PARRA J, C GRAHAM y J FREILE. 2004. Evaluating alternative data sets for ecological niche models of birds in the Andes. Ecography 27:350-360. [ Links ]
PARDIÑAS UFJ, G D'ELIA, P TETA, P ORTIZ, JP JAYAT y S CIRIGNOLI. En prensa. Akodontini Vorontsov, 1959 (sensu D'Elía, 2003). En: Mamíferos de Argentina: sistemática y distribución (RM Barquez, MM Díaz y RA Ojeda, eds.). Sociedad Argentina para el Estudio de los Mamíferos. [ Links ]
PEARSON OP. 1958. A taxonomic revision of the rodent Genus Phyllotis. University of California Publications in Zoology 56:391-496. [ Links ]
PETERSON A, J SOBERÓN y V SÁNCHEZ - CORDERO. 1999. Conservatism of ecological niches in evolutionary time. Science 285:1265-1267. [ Links ]
PETERSON A, L BALL y K COHOON. 2002. Predicting distributions of Mexican birds using ecological niche modelling methods. Ibis 144:E27-E32. [ Links ]
REIG OA. 1987. An assessment of the systematics and evolution of the Akodontini, with the description of new fossil species of Akodon (Cricetidae, Sigmodontinae). Pp. 347-399, en: Studies in Neotropical Mammalogy (BD Patterson y RM Timm, eds.). Fieldiana, Zoology, New Series 39. [ Links ]
SCOTT J, P HEGLUND, M MORRISON, J HAUFLER, M RAPHAEL, W WALL y F SAMSON. 2002. Predicting species occurrence: issues of accuracy and scale. Island Press, Washington DC. [ Links ]
SEGURADO P y MB ARAUJO. 2004. An evaluation of methods for modeling species distributions. Journal of Biogeography 31:1555-1568. [ Links ]
STOCKWELL D y I NOBLE. 1992. Induction of sets of rules from animal distribution data: a robust and informative method of data analysis. Mathematics and Computers in Simulation 33:385-390. [ Links ]
THOMAS O. 1918. On small mammals from Salta and Jujuy collected by Mr. E. Budin. Annals and Magazine of Natural History 9:186-193. [ Links ]
THOMAS O. 1919. On small mammals from Catamarca "Otro Cerro" North-eastern Rioja, collected by Sr. E. Budin. Annals and Magazine of Natural History 9:489-500. [ Links ]
THOMAS O. 1921. On a further Collection of Mammals from Jujuy obtained by Sr. Budin. Annals and Magazine of Natural History 9:608-617. [ Links ]
THOMAS O. 1926. Two new mammals from north Argentina. Annals and Magazine of Natural History 9:311-313. [ Links ]
VAN MANEN FT, JD CLARK, SE SCHLARBAUM, K JOHNSON y G TAYLOR. 2002. A model to predict the occurrence of surviving butternut trees in the southern Blue Ridge Mountains. Pp. 491-497, en: Predicting species occurrences, issues of accuracy and scale (JM Scott, PJ Heglund, ML Morrison, JB Haufler, MG Raphel, WA Wall y FB Samson, eds.). Island Press. [ Links ]
VARGAS JH, T CONSIGLIO, PM JORGENSEN y TB CROAT. 2004. Modelling distribution patterns in a species-rich plant genus, Anthurium (Araceae), in Ecuador. Diversity and Distributions 10:211-216. [ Links ]
Recibido 5 agosto 2005.
Aceptación final 1º abril 2006.

