










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkMastozoología neotropical
versão impressa ISSN 0327-9383versão On-line ISSN 1666-0536
Mastozool. neotrop. v.13 n.1 Mendoza jan./jun. 2006
First record of Rhagomys (Mammalia: Sigmodontinae) in Bolivia
Gabriela Villalpando1, Julieta Vargas1, and Jorge Salazar-Bravo2
1 Museo Nacional de Historia Natural (Colección Boliviana de Fauna), Casilla 8706, La Paz, Bolivia, 2 Department of Biological Sciences, Texas Tech University, Lubbock, TX 79409, USA
Key words. Bolivia. Distribution. Rhagomys.
A recent monograph on the biological diversity of Bolivia concludes that the country is megadiverse (Ibisch and Merida, 2003); this statement is especially true for mammals. Despite being a landlocked country and representing only 6% of the geographic area of South America, Bolivia harbors the 4 th most-diverse mammalian fauna of the continent (Ceballos and Simmonetti, 2002). However, the country remains poorly explored, as reports of new taxa for science or the country have recently been published (Pacheco et al., 2004; Rios-Uzeda et al., 2004; Solmsdorff et al., 2004; Emmons and Patton, 2005). These new records and species, coupled with further advances in the taxonomy and systematics on Neotropical mammals have rendered sections of Anderson 's book Mammals of Bolivia (Anderson, 1997) obsolete in less than a decade after its publication. Accordingly, in this paper, we report a new genus and species of mammal for the fauna of Bolivia.
Measurements of the animal followed Luna and Patterson (2003) except for greatest length of skull (GLS), taken to be represented by the occipito-nasal length, and CML (Condylomolar length), which was not measured because it was not defined either by Luna and Patterson (2003) or any of the references cited therein (Myers et al., 1990; Voss, 1991 or Musser et al., 1998).
Rhagomys longilingua
Luna and Patterson, 2003
The Bolivian record of this distinctive taxon is based on a unique specimen obtained in Parque Nacional y Area Natural de Manejo Integrado Cotapata (PN-ANMI Cotapata, Fig. 1). The senior author caught the individual in a pitfall trap on March 28 th, 2003 at Bajo Hornuni (16° 12.8' S, 67° 53.2 W, 1860 m elevation), prepared the animal as round skin and skull and deposited the specimen in the Colección Boliviana de Fauna, where it was assigned the catalog number CBF 7620. The specimen is a juvenile female, toothwear class TWC 1 of Voss (1991), as judged by the wooly pelage, the incomplete ossification of the basisphenoid-basioccipital suture, and the absence of wear in any of the molars.
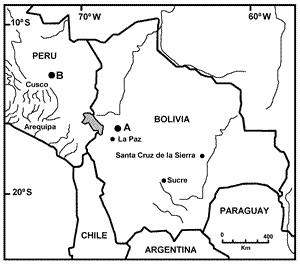
Fig. 1. Map of the localities mentioned in the text. A: Bajo Hornuni in the Parque Nacional y Area de Manejo Integral Cotapata; B: the type locality of Rhagomys longilingua in Peru.
The Bolivian specimen (CBF 7620) is identified as Rhagomys longilingua by the following diagnostic characters: the texture of the pelage is markedly spiny; all digits of manus and pes have broad, squared-shaped, blunt, and callous tips and deep transversal grooves. Finger tips bear a crescent-shaped depression and the hallux bears a nail instead of a claw. Cranially, CBF 7620 bears a large oval-shaped fontanelle in the parapterygoid fossae (Fig. 2). Its M3 has only a paracone and protocone, m2 and m3 are nearly the same size, with distinct metaconid, protoconid, posterolophid and hypoconid elements, and all mandibular teeth (m1-m3) have a posterocrista. The only diagnostic tooth character for R. longilingua missing in CBF 7620 is the anteromedian crista on M1, which in R. longilingua protrudes from the anterolabial conule. Other diagnostic features such as the length of the tongue were not recorded at the time of capture of CBF 7620 and are now missing.

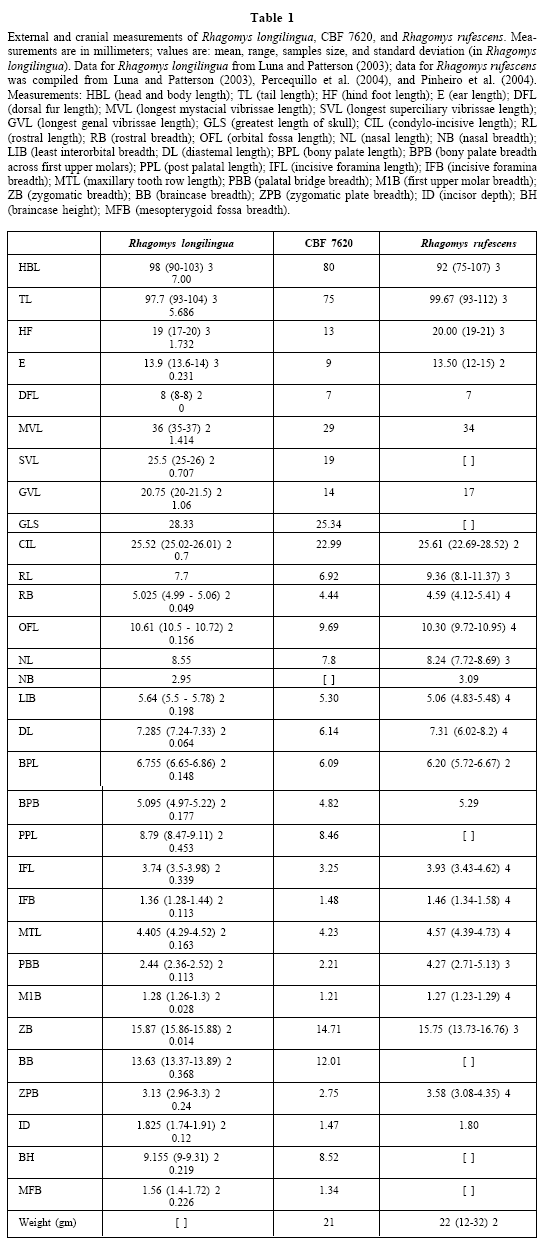
Fig. 2. Dorsal, ventral and lateral views of the cranium and mandible of Rhagomys longilingua from Bolivia (CBF 7620). Scale bar = 5mm
The pelage coloration in CBF 7620 is strongly countershaded, resembling the description of R. longilingua (Luna and Patterson, 2003) except that the pelage in the Bolivian specimen is grayer on the dorsum and rump, with a woolly appearance presumably due to its youth. As in Peruvian R. longilingua the underparts of CBF 7620 are Light Ochraceous-Buff (Ridgway, 1912). However, CBF 7620 has a small patch (5 by 7 mm) of whitish hairs on the chest that is consistently missing in the Peruvian specimens.
The skull of CBF 7620 is strongly built; it bears a short rostrum, posteriorly divergent, beaded supraorbital margins, but only weak postorbital processes (Fig. 2). In profile, the skull appears slightly more convex than shown by Luna and Patterson (2003), but differences may be due to age. Other differences between CBF 7620 and the depicted paratype of R. longilingua include the size of the posterior opening of the alisphenoid canal, which in CBF 7620 is larger across than the strut of bone separating it from the foramen ovale accessorius, whereas it is about half as long in R. longilingua. The shape of the foramen ovale accessorius also differs, being clearly triangular in CBF 7620 as opposed to oval-shaped in R. longilingua. The posterolateral palatal pits depicted in Luna and Patterson (2003) are well defined and appear as single depressions on both sides of the mesopterygoid fossa, whereas on CBF 7620 the right side of the fossa presents three depressions and the left side only one. These and other minor morphological differences could be the result of geographic or interpopulational variation as seen in other groups of sigmodontine rodents (e.g., Musser et al., 1998). No postcranial or soft anatomic details were preserved for CBF 7620.
External, cranial and molar morphometric data for the Bolivian specimen demonstrate the smaller size of this individual (Table 1); except for one measurement (IFB), all measurements of CBF 7620 are smaller than reported for Rhagomys longilingua (n=3 for external characters, n=2 for dental and cranial characters). The incisive foramen of the Bolivian specimen is wider than either of the two measured by Luna and Patterson (2003). Because CBF 7620 is a young individual some morphometric differences were to be expected, others may be due to measurers' variation, especially in areas of the skull apparently unaffected by postweaning growth in other sigmodontines (e.g., IBL, IFB, MTL and M1B; Voss 1988).
Ecological notes: The PN-ANMI Cotapata (Fig. 1) is located on the eastern slope of the Andes, some 70 km E of La Paz. The park houses abrupt changes in elevation (from 1100 to 5600 m) with concomitant changes in ecological characteristics and vegetation (Ribera-Arismendi, 1995; Paniagua et al., 2003). CBF 7620 was captured in a humid montane forest on the eastern slope of the Andes, characterized by an annual precipitation of less than 2000 mm and temperatures ranging between 17 and 20 °C. The soils at this locality are markedly acidic (pH below 4.5) with strong saturation of Al 3+, and a thick organic layer (Back et al., 2003).
The dominant vegetation is that of an open forest, between 15 and 30 m tall with a continuous and dense canopy and emergents that reach 40 m. Among the most important arboreal canopy species are Hyeronima alchorneoides, Morus insignis, Liconia triandra, Ficus trigona, Ficus guianensis, Hedyosmum angustifolium, Virola sp., Inga spp., and some Rubiaceae species of the genera Ladenbergia and Bathysa.
The pitfall trap where CBF 7620 was captured was surrounded by abundant fallen leaves, branches and fallen coarse tree trunks. The soil had a thick organic layer and humus-type mull 30 cm deep. The senior author also trapped Akodon fumeus, Oxymycterus paramensis, Neacomys n.sp., Oryzomys yunganus, Oligoryzomys destructor, Oligoryzomys indet., Marmosops noctivagus, and Micoureus constantiae at the site. Vargas et al. (2003) reported Monodelphis adusta from the same general area.
This record extends the distribution of Rhagomys longilingua by ca. 520 km to the southeast of its type locality in the Manu Biosphere Reserve along Rio Cosñipata (13° 06.032'S, 71° 34.125' W, 1900 m elevation). The species ranges from 450 m (in Peru) to 1900 m (in Peru and Bolivia).
As noted by Luna and Patterson (2003), Rhagomys is the first mammalian group to show a disjunct distribution between the Brazilian Atlantic Rainforest (Rhagomys rufescens) and the eastern slopes of the Andes (Rhagomys longilingua). Other groups show the same pattern of distribution; besides frogs of the Hyla parvidens group cited by Luna and Patterson (2003), we found several groups of birds (e.g., the cotingas Phibalura flavirostris and Phyllomyias fasciatus, Phyllomyias sp. [Snow et al., 2004]), seven species of ferns (Smith et al., 1999), and two species of epiphytic cacti (Ibisch, 1996) that show the same pattern. Therefore, the vicariant nature of these distributions may indicate a once continuous distribution in Rhagomys which is now interrupted. Interestingly, it was suggested that such interruption may have occurred around the Plio-Pleistocene as the fluvial terraces of the Chaco, Beni and Pantanal subsided due to tectonic movements (Iriondo, 1993).
Recently, a phylogenetic analysis of sequences of the IRBP gene suggested that Rhagomys belongs to a clade containing Thomasomys, Rhipidomys and Aepeomys (D'Elia et al., 2006), which corresponds to the tribe Thomasomyini (sensu Smith and Patton, 1999). Because Thomasomys and Aepeomys are exclusively Andean in distribution and the more basal species in Rhipidomys are found in northwestern Amazonia (Costa, 2003), there is a possibility that Rhagomys may have originated in either the premontane forests region of the northern Andes, or the lowland forests of the western Amazonian basin. These areas have also been considered as the potential ancestral areas for the oryzomyines (Weksler, 2006). Therefore, the presence of Rhagomys in southeastern Brazil could have been the result of dispersal across the Amazon basin and Cerrado domains, maybe following the Paraná River basin "pathway" discussed by Costa (2003). Our record of Rhagomys in Bolivia support Luna and Patterson's suggestion that the genus may be continuously distributed across South America, and thus, collectors working in areas between the forested slopes of the Andes and the Atlantic Rainforest, especially in the interior Atlantic Rainforest of Misiones in Argentina, or south eastern Paraguay should be alert to the possible presence of Rhagomys in this area, either R. rufescens or a new sister species. Because 3 of the 4 specimens known of Rhagomys longilingua - including the new record for Bolivia - were obtained with pitfall traps we suggest the use of this kind of traps in their surveys. We concur with Luna and Patterson (2003) on the importance of Rhagomys in understanding the current diversity of the sigmodontine radiation.
We kindly acknowledge the financial support from the following institutions: Instituto de Ecologia, Universidad Mayor de San Andrés (to GV) and Texas Tech University (Grants Program of the Vice-President for Research) and NIH U19 AI4545 2, the ICIDR Opportunity Pool Award (to JSB). Critical reviews by Lucia Luna, Bruce Patterson and an anonymous reviewer contributed to the final version of the manuscript.
LITERATURE CITED
ANDERSON S. 1997. Mammals of Bolivia: taxonomy and distribution. Bulletin of the American Museum of Natural History 231:1-652. [ Links ]
BACK K, M SCHAWE, S BECK, G GEROLD, G GRADSTEIN, and M MORAES. 2003. Vegetación, suelos y clima en los diferentes pesos altitudinales de un bosque montano de Yungas, Bolivia: Primeros resultados. Ecología en Bolivia 38:3-13 [ Links ]
CEBALLOS G and JA SIMONETTI. 2002. Diversidad y conservación de los mamíferos neotropicales, 1ª ed. CONABIO Instituto de Ecología Universidad Nacional Autónoma de México, México. [ Links ]
COSTA LP. 2003. The historical bridge between the Amazon and the Atlantic Forest of Brazil: a study of molecular phylogeography with small mammals. Journal of Biogeography 30: 71-86. [ Links ]
D'ELIA G, L LUNA, EM GONZALEZ, and BD PATTERSON. 2006. On the Sigmodontinae radiation (Rodentia, Cricetidae): An appraisal of the phylogenetic position of Rhagomys. Molecular Phylogenetics and Evolution 38: 558-564. [ Links ]
EMMONS LH and JL PATTON. 2005. A New Species of Oryzomys (Rodentia: Muridae) from Eastern Bolivia. American Museum Novitates 3478:1-26. [ Links ]
IBISCH PL and G MERIDA. 2003. Biodiversidad: la riqueza de Bolivia. Estado de conocimiento y conservación. Editorial FAN, Santa Cruz de la Sierra, Bolivia. [ Links ]
IBISCH PL. 1996. Neotropische epiphytendiversität - das Biespiel Bolivien. Martina Galunder-Verlag, Wiehl, Germany. [ Links ]
IRIONDO M. 1993. Geomorphology and Late Quaternary of the Chaco (South-America). Geomorphology 7: 289-303. [ Links ]
LUNA L and B PATTERSON. 2003. A remarkable new mouse (Muridae: Sigmodontinae) from southeastern Peru: with comments on the affinities of Rhagomys rufescens (Thomas, 1886). Fieldiana, Zoology 101:1-24. [ Links ]
MUSSER GG, MD CARLETON, E BROTHERS, and AL GARDNER. 1998. Systematic studies of oryzomyine rodents (Muridae, Sigmodontinae): Diagnoses and distributions of species formerly assigned to Oryzomys " capito ". Bulletin of the American Museum of Natural History 236: 1-376. [ Links ]
MYERS P, JL PATTON and MF SMITH. 1990. A review of the boliviensis group of Akodon (Muridae: Sigmodontinae), with emphasis on Peru and Bolivia. Miscellaneous Publications, Museum of Zoology, University of Michigan 177: 1-104. [ Links ]
PACHECO V, S SOLARI and PM VELAZCO. 2004. A new species of Carollia (Chiroptera: Phyllostomidae) from Peru and Bolivia. Occasional Papers, Museum of Texas Tech University 236:1-15. [ Links ]
PANIAGUA N, C MALDONADO and C CHUMACERO. 2003. Mapa de Vegetación de los alrededores de la estación Biológica de Tunquini, Bolivia. Ecología en Bolivia 38:15-26 [ Links ]
PERCEQUILLO A, P GONCALVES and J DE OLIVEIRA. 2004. The re-discovery of Rhagomys rufescens (Thomas, 1886), with a morphological re-description and comments on its systematic relationships based on morphological and molecular (cytochrome b) characters. Mammalian Biology 69:238-257. [ Links ]
PINHEIRO P, P HARTMANN and L GEISE. 2004. New record of Rhagomys rufescens (Thomas 1886) (Rodentia: Muridae). Zootaxa 431:1-11 [ Links ]
RIBERA-ARISMENDI MO. 1995. Aspectos Ecológicos del uso de la tierra y conservación en el Parque Nacional y Área de Manejo Integrado Cotapata. In: Caminos de Cotapata (C Morales, ed.). Instituto de Ecología, FUND-ECO, FONAMA-EIA. La Paz, Bolivia. [ Links ]
RIDGWAY R. 1912. Color standards and color nomenclature. The author, Washington, D.C. [ Links ]
RIOS-UZEDA B, R WALLACE and J VARGAS. 2004. La jayupa de la altura (Cuniculus taczanowskii, Rodentia, Cuniculidae), un nuevo registro de mamífero para la fauna de Bolivia, Mastozoologia Neotropical 11: 109-114. [ Links ]
SALAZAR-BRAVO J, T TARIFA, LF AGUIRRE, E YENSEN, and T YATES. 2003. Revised checklist of Bolivian mammals. Occasional Papers, The Museum of Texas Tech University, 220:1-27. [ Links ]
SMITH AR, M KESSLER and J GONZALES. 1999. New records of pteridophytes from Bolivia. American Fern Journal 89: 244-266. [ Links ]
SMITH MF and JL PATTON. 1999. Phylogenetic relationships and the radiation of sigmodontine rodents in South America: Evidence from cytochrome b, Journal of Mammalian Evolution 6: 89-128. [ Links ]
SNOW D, M BROOKE and B WALTHER. 2004. Family Cotingidae in Handbook of the Birds of the World. Vol 9: Cotingas to Pipits and Wagtails (J Del Hoyo, A Elliot and D Christie, eds.). Lynx Editions, Barcelona. [ Links ]
SOLMSDORFF K, D KOCK, C HOHOFF, and N SACHSER. 2004. Comments on the genus Galea MEYEN 1833 with description of Galea monasteriensis n. sp. from Bolivia (Mammalia, Rodentia, Caviidae). Senckenbergiana Biologica 84:137-156. [ Links ]
VARGAS J, T TARIFA and C CORTEZ. 2003. Nuevos registros de Monodelphis adusta y Monodelphis kunsi (Didelphimorphia: Didelphidae) para Bolivia. Mastozoología Neotropical 10: 123-131. [ Links ]
VOSS RS. 1988. Systematics and ecology of ichthyomyine rodents (Muroidea): Patterns of morphological evolution in a small adaptive radiation. Bulletin of the American Museum of Natural History 188: 259-493. [ Links ]
VOSS RS. 1991. An introduction to the Neotropical muroid rodent genus Zygodontomys. Bulletin of the American Museum of Natural History 210: 1-113. [ Links ]
WEKSLER M. 2006. Phylogenetic relationships of the Oryzomyini (Muroidea: Sigmodontinae): Separate and combined analyses of morphological and molecular data. Bulletin of the American Museum of Natural History 296: 1-149. [ Links ]
Recibido 9 setiembre 2005.
Aceptación final 31 enero 2006.

