










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. v.14 n.1 Mendoza ene./jun. 2007
Fossil record of the Andean rat, Andinomys edax (Rodentia: Cricetidae), in Argentina
Pablo E. Ortiz¹ and J. Pablo Jayat²
¹ Cátedra de Paleozoología II, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Miguel Lillo 205, 4000 San Miguel de Tucumán, Tucumán, Argentina <peortiz@uolsinectis.com.ar>. ² Laboratorio de Investigaciones Ecológicas de las Yungas (LIEY), Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Casilla de Correo 34, 4107 Yerba Buena, Tucumán, Argentina.
ABSTRACT: Andinomys edax (Rodentia, Cricetidae) has a broad distribution in the Central Andean region. The fossil record is restricted to only three localities from Lower-Middle Pleistocene in Bolivia to Upper Holocene in Argentina. Recent field work in northwestern Argentina produced five new paleontological localities: Las Juntas, Catamarca (13.4 - 12.95 kyr BP); Inca Cueva 4, Jujuy (11 - 9 kyr BP); Tafí del Valle, Tucumán (10.25 - 9.75 kyr BP); Los Viscos, Catamarca (1.88 kyr bp); and La Cueva, Jujuy (0.95 kyr BP). The paleontological data indicate that A. edax has been a ubiquitous sigmodontine in northwestern Argentina since the Upper Pleistocene.
RESUMEN: Registro fósil de la rata andina, Andinomys edax (Rodentia: Cricetidae), en Argentina. Andinomys edax presenta una amplia distribución en la región andina central. Su registro fósil está restringido a sólo tres localidades, desde el Pleistoceno Inferior-Medio en Bolivia hasta el Holoceno Superior en Argentina. Recientes trabajos de campo en el noroeste argentino resultaron en cinco nuevas localidades paleontológicas: Las Juntas, Catamarca (13.4 - 12.95 ka AP); Inca Cueva 4, Jujuy (11 - 9 ka AP); Tafí del Valle, Tucumán (10.25 - 9.75 ka AP); Los Viscos, Catamarca (1.88 ka A p); y La Cueva, Jujuy (0.95 ka AP). La información paleontológica indica que A. edax ha sido un sigmodontino ubícuito en el noroeste argentino desde el Pleistoceno Superior.
Key words. Northwestern Argentina. Paleontological record. Quaternary. Sigmodontinae.
Palabras clave. Cuaternario. Noroeste argentino. Registro paleontológico. Sigmodontinae.
Among the Neotropical mammals, the subfamily Sigmodontinae (Rodentia, Cricetidae) is one of the most diverse and complex groups, with ca. 380 species (Musser and Carleton, 2005). At present, the monotypic Andinomys Thomas, 1902, one of the most enigmatic genera of sigmodontines, is treated as a "unique line" of the subfamily or as Sigmodontinae incertae sedis (D'Elía et al., 2005; D'Elía et al., 2006). Andinomys edax Thomas, 1902, has a broad distribution in the Central Andean region from southeastern Peru, through southwestern Bolivia and northernmost Chile to northwestern Argentina (Yepes, 1935; Hershkovitz, 1962; Pine et al., 1979; Heinonen and Bosso, 1994; Anderson, 1997; Mares et al., 1997; Díaz, 1999; Eisenberg and Redford, 1999; Díaz et al., 2000; Ortiz et al., 2000a). This is a very distinctive species because of its peculiar cranial, dental (Hershkovitz, 1962; Steppan, 1995) and molecular characters (D'Elía et al., 2005).
The fossil record of A. edax is poorly known and is restricted to three localities: one from Lower-Middle Pleistocene in Tarija, Bolivia (Marshall et al., 1984; Marshall and Sempere, 1991; Pardiñas et al., 2002), and two from Middle-Upper Pleistocene and Upper Holocene in Argentina (Ortiz and Pardiñas, 2001; Teta and Ortiz, 2002; Pardiñas et al., 2002). In recent years, field work carried out in northwestern Argentina resulted in new paleontological localities for A. edax where several remains were recovered. In this note we report these findings.
The fossil material was collected using wet and dry sieves with a 0.1 mm mesh, and identified by comparisons with museum specimens and the literature (Yepes, 1935; Hershkovitz, 1962; Steppan, 1995). Craniodental measurements were taken with digital calipers and recorded to the nearest 0.01 mm (Table 1). The studied material is housed in the following institutions: Colección Paleontología de Vertebrados del Instituto-Fundación Miguel Lillo (PVL), Universidad Nacional de Tucumán, Tucumán, Argentina; Colección del Grupo Yavi (GY), Jujuy, Argentina, and Sección Arqueología del Instituto de Ciencias Antropológicas (ICA), Facultad de Filosofía y Letras, Buenos Aires, Argentina.

The remains were assigned to A. edax based on several characters: interfrontal fontanelles; deep zygomatic plate with anterior border slightly concave and anterodorsal corner projecting as a short spine; mandible robust with coronoid process higher than the articular condyle; anterior margin of masseteric crest extending to the level of protoconid m1; molars large, prismatic and flat crowned, with penetrating flexi and flexids (Figs. 1 and 2).
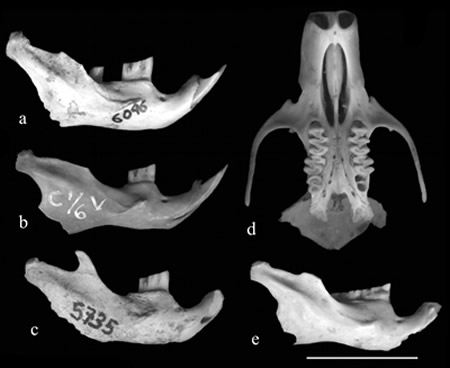
Fig. 1 Andinomys edax. a: external view of right mandible (PVL 6096) from Las Juntas; b: external view of right mandible (GY 1/6 V) from La Cueva; c: external view of right mandible (PVL 5735) from Tafì del Valle; d: palatal view of skull fragment (PVL 5879) from Los Viscos, level 7; e: external view of right mandible (ICA Arqueología 062) from Inca Cueva 4, layer 2. Scale Bar = 10 mm.

Fig. 2. Andinomys edax. a: occlusal view of M1-M3 of PVL 5879, Los Viscos, level 7; b: occlusal view of m1-m2 of PVL 5877, Las Juntas. Scale bar = 2 mm.
Until now, A. edax was known in Argentina from just two fossil localities (Fig. 3) (Ortiz, 2001; Ortiz and Pardiñas, 2001; Teta and Ortiz, 2002). The earliest record comes from La Angostura (26º 56' 25"S, 65º 42' 10"W, 1900 m, Tafí del Valle Department, Tucumán Province), Tafí del Valle Formation, Middle-Upper Pleistocene (Collantes et al., 1993). Recent datings indicate that this geologic unit ranges from 190.5 to 64 kyr BP (Kemp et al., 2003, 2004; Carter-Stiglitz et al., 2006). However, magnetostratigraphic analysis by Schellenberger et al. (2003) suggests that the Tafí del Valle Formation is significantly older, with the base of this formation dating to 1.15 Ma BP. In this site the remains of A. edax were associated with several murid and caviomorph rodent species (Ortiz, 2001; Ortiz and Pardiñas, 2001). The other known locality is the Inca Cueva 5 archaeological site (23° 02' 55" S, 65º 25' 07" W, 3700 m, Humahuaca Department, Jujuy Province). Here, the A. edax remains come from levels dated between 2.12 and 0.72 kyr BP (García, 1991; Teta and Ortiz, 2002).
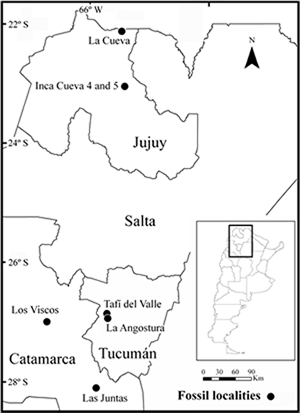
Fig. 3. Map of Northwestern Argentina showing the fossil localities where Andinomys edax has been registered.
In this paper we add five new paleontological localities with A. edax remains in Catamarca, Jujuy and Tucumán provinces. All the studied specimens are listed in Appendix 1. The oldest remains were found in Late Pleistocene loessic sediments dated between 13.4 and 12.95 kyr BP in Las Juntas (28° 06' 36"S, 65° 53' 54"W, 1562 m, Ambato Department, Catamarca Province), consisting of 16 maxillae and 18 mandibles. The remains from Inca Cueva 4, layer 2 (23° 02' 55"S, 65º 25' 07"W, 3650 m, Humahuaca Department, Jujuy Province), come from the Pleistocene-Holocene boundary (dated between 11 and 9 kyr BP; Yacobaccio, 1991) and consist only of two mandibles. In the Tafí del Valle site (26° 51' 43"S, 65° 43' 06"W, 2007 m, Tafí del Valle Department, Tucumán Province) we recovered six mandibles and one isolated frontal from sediments of the Pleistocene-Holocene boundary dated between 10.25 and 9.75 kyr BP (Ortiz and Jayat, in prep.). We report several A. edax remains belonging to seven individuals from Late Holocene of Los Viscos, level 7 (26° 59' 55"S, 66° 43' 34"W, 2400 m, Belén Department, Catamarca Province), dated to 1.88 kyr bp. Finally, from La Cueva archaeological site (22º 07' 28"S, 65º 27' 58"W, 3440 m, Yavi Department, Jujuy Province), we studied several cranial and mandibular remains from 0.95 kyr BP level (Ortiz, 2001).
Andinomys edax is present in seven of the nine Late Quaternary micromammal localities known for northwestern Argentina. However, is not an abundant taxon in any of these assemblages, showing low frequencies ranging from 6 to 11% (Ortiz, 2001; Ortiz and Pardiñas, 2001; Teta and Ortiz, 2002; Ortiz and Jayat, in prep.). Likewise, at present, this species is not abundant and its frequency values are even lower than in fossil assemblages. In owl pellet samples recovered in many localities in the region, values for A. edax do not exceed 2%. It also comprises a minority of captures (P. Jayat, pers. obs.).
Nowadays, A. edax inhabits several environments such as subtropical mountain forests (Capllonch et al., 1997), high-altitude grasslands (Díaz, 1999; Díaz and Barquez, 1999; Ortiz et al., 2000a; Jayat et al., in prep.), and semiarid areas in Puna and Prepuna (Díaz, 1999; Ortiz et al., 2000a). Its wide distributional range in such contrasting environmental units indicates its ecological plasticity. Similar conclusions can be obtained from the fossil record. All fossil localities for A. edax are located in areas where the species occurs today, despite the dramatic environmental fluctuations that happened during the Late Quaternary. Paleontological data indicate that Late Pleistocene glacial cycles, reflected by the displacement of vegetation belts on mountain slopes, would have had minor influence on A. edax distribution. Some of the fossil associations of micromammals where A. edax is present, such as La Angostura (Ortiz and Pardiñas, 2001), Tafí del Valle (Ortiz and Jayat, in prep.) and Las Juntas (Ortiz et al., in prep.), indicate colder and drier Late Pleistocene paleoenvironmental conditions than at present. Neotomys ebriosus, Reithrodon auritus and Abrocoma cinerea are present in these assemblages, rodent species that today inhabit higher elevation environments in this region (see Pardiñas and Galliari, 2001; Pardiñas and Ortiz, 2001; Braun and Mares, 2002). On the other hand, the Late Holocene Inca Cueva 5 assemblage, where A. edax is relatively well represented (11%), suggests more humid environmental conditions than today (Teta and Ortiz, 2002). The Quaternary glacial cycles have impacted micromammal communities in the high Andes of northwestern Argentina with changes in the frequency and composition of species in the assemblages (Ortiz and Pardiñas, 2001; Pardiñas and Ortiz, 2001; Pardiñas et al., 2002; Ortiz, 2003; Ortiz and Jayat, in prep.), resulting in the extinction of at least one species (Ortiz et al., 2000b). Notwithstanding the continuous faunistic turnover, A. edax has been a ubiquitous sigmodontine in northwestern Argentina since Upper Pleistocene times.
Examined specimens of Andimonys edax from the five fossil localities
Las Juntas: two right mandibles with m1-m3 (PVL 5866, PVL 6096), one right mandible with m1 (PVL 5867), eight right mandibles without molars (PVL 5868, PVL 5872, PVL 5873, PVL 6094, PVL 6099, PVL 6100, PVL 6101, PVL 6106), four left maxillae with zygomatic plate without molars (PVL 5869, PVL 6097, PVL 6111, PVL 6112), three right maxillae with M1 (PVL 5870, PVL 6092, PVL 6107), one left mandible with m3 (PVL 5871), one right maxilla with M1-M2 (PVL 5874), two right mandibles with m3 (PVL 5875, PVL 6093), several isolated incisors and molars (PVL 5876), one left mandible with m1-m2 (PVL 5877), two left mandibles without molars (PVL 6095, PVL 6104), one left mandible with m1 (PVL 6098), five right maxillae without molars (PVL 6102, PVL 6103, PVL 6105, PVL 6109, PVL 6110), one left maxilla with M1 (PVL 6108) and two left maxillae without molars (PVL 6210, PVL 6213).
Inca Cueva 4, layer 2: one left mandible with incisor (ICA Arqueología 031) and one right mandible with m1-m3 (ICA Arqueología 062).
Tafí del Valle: two right mandibles with m1 (PVL 5735, PVL 5764), one left mandible with incisor and m1 (PVL 5763), two left mandibles without molars (PVL 5765; PVL 5766) and one isolated frontal (PVL 5767).
La Cueva: one right mandible with m1 and m3 (GY 1/6 V); two left maxillae (GY 1/6 IX; GY 1/6 XV); one left maxilla with M1 (GY 1/6 XVII); one left maxilla and 1 left mandible (GY 1/6 XVIII); isolated M2 and m3 (GY 1/6 XXXIX); one isolated M1 (GY 1/7 V); one isolated M2 (GY 1/7 VIII); one isolated M3 (GY 1/7 IX); one right maxilla, one left maxilla, one left mandible, several isolated upper and lower molars (GY 1/7 XIII); one left mandible (GY 2/6 VI); one right mandible with m1 (GY 2/6 IX); isolated upper and lower molars (GY 2/6 XII); one right maxilla and isolated M3 (GY 2/7 XI).
Los Viscos, level 7: two skull fragments (PVL 5878; PVL 5879), one left mandible with m1-m2 (PVL 5880), three left mandibles with m1 (PVL 5881, PVL 5886, PVL 5884), two left mandibles with m1-m3 (PVL 5882, PVL 5885), one left mandible with m3 (PVL 5883), one right mandible (PVL 5887), three right mandibles with m1 and m3 (PVL 5888, PVL 5891, PVL 5893), two right mandibles with m1 (PVL 5889, PVL 5892), 1 right mandible with m1-m2 (PVL 5890), 1 left maxilla with zygomatic plate (PVL 5894), two left maxillae with M1-M2 and zygomatic plate (PVL 5895, PVL 5896), one left maxilla with M1-M3 (PVL 5897), two right maxillae with M1-M2 (PVL 5898, PVL 5899) and isolated molars and incisors (PVL 5900).
ACKNOWLEDGMENTS
We thank J. Kulemeyer and U. Pardiñas for providing La Cueva materials, and H. Yacobaccio and M. Mondini for Inca Cueva 4 archaeological remains. J. Babot, J. Ballesteros, D. García Lopez, C. Herrera, D. Juri, C. Madozzo Jaén, D. Miotti, G. Ortiz Tempel, and F. Scacchi helped us during field works. A. Malizia reviewed the English version. N. Nasif, M.G. Vucetich, S. Vizcaíno and an anonymous reviewer provided insightful comments and suggestions that helped improve this manuscript. Financial support was provided by Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT), Consejo de Investigaciones de la Universidad Nacional de Tucumán (CIUNT) and Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET). We thank the institutional support of Cátedra de Paleozoología II and Laboratorio de Investigaciones Ecológicas de Las Yungas (LIEY), Universidad Nacional de Tucumán.
LITERATURE CITED
ANDERSON S. 1997. Mammals of Bolivia, taxonomy and distribution. Bulletin American Museum of Natural History 231:1-652. [ Links ]
BRAUN JK and M MARES. 2002. Systematics of the Abrocoma cinerea species complex (Rodentia: Abrocomidae), with a description of a new species of Abrocoma. Journal of Mammalogy 83:1-19. [ Links ]
CAPLLONCH P, A AUTINO, MM DÍAZ, RM BARQUEZ, and M GOYTIA. 1997. Los mamíferos del Parque Biológico Sierra de San Javier, Tucumán, Argentina: observaciones sobre su sistemática y distribución. Mastozoología Neotropical 4:49-71. [ Links ]
CARTER-STIGLITZ B, SK BANERJEE, A GOURLAN, and E OCHES. 2006. A multi-proxy study of Argentina loess: Marine oxygen isotope stage 4 and 5 environmental record from pedogenic hematite. Palaeogeography, Palaeoclimatology, Palaeoecology 239:45-62. [ Links ]
COLLANTES M, J POWELL, and J SAYAGO. 1993. Formación Tafí del Valle (Cuaternario superior), provincia de Tucumán (Argentina): litología, paleontología y paleoambientes. XII Congreso Geológico Argentino y II Congreso de Exploración de Hidrocarburos, Actas 2:200-206. [ Links ]
D'ELÍA G, UFJ PARDIÑAS, and P MYERS. 2005. An introduction to the genus Bibimys (Rodentia: Sigmodontinae): Phylogenetic position and alpha taxonomy. Pp. 211-246, in: Mammalian diversification: from Chromosomes to Phylogeography (A Celebration of the Career of James L. Patton) (E Lacey and P Myers, eds.). University of California Publication in Zoology, Berkeley, 133. [ Links ]
D'ELÍA G, P TETA, and UFJ PARDIÑAS. 2006. Incertae Sedis. Pp. 197-202, in: Mamíferos de Argentina: sistemática y distribución (RM Barquez, MM Díaz, and RA Ojeda, eds.). Sociedad Argentina para el Estudio de los Mamíferos. [ Links ]
DÍAZ MM. 1999. Mamíferos de la Provincia de Jujuy: sistemática, distribución y ecología. Unpublished Ph.D. dissertation, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán. [ Links ]
DÍAZ MM and RM BARQUEZ. 1999. Contributions to the knowledge of the mammals of Jujuy Province, Argentina. The Southwestern Naturalist 44:324-333. [ Links ]
DÍAZ MM, J BRAUN, M MARES, and RM BARQUEZ. 2000. An update of the taxonomy, systematics, and distribution of the mammals of Salta Province, Argentina. Occasional Papers, Sam Noble Oklahoma Museum of Natural History 10:1-52. [ Links ]
EISENBERG JF and KH REDFORD. 1999. Mammals of the Neotropics. The Central Neotropics. Ecuador, Peru, Bolivia, Brazil. Volume 3. University of Chicago Press, Chicago. [ Links ]
GARCÍA L. 1991. Variabilidad funcional de los sitios con cerámica en cuevas y aleros de la Quebrada de Inca Cueva (Jujuy). Shincal 3:64-68. [ Links ]
HEINONEN S and A BOSSO. 1994. Nuevos aportes para el conocimiento de la mastofauna del Parque Nacional Calilegua (Provincia de Jujuy, Argentina). Mastozoología Neotropical 1:51-60. [ Links ]
HERSHKOVITZ P. 1962. Evolution of Neotropical cricetine rodents (Muridae) with special reference to the phyllotine group. Fieldiana: Zoology 46:1-524. [ Links ]
KEMP RA, PS TOMS, JM SAYAGO, E DERBYSHIRE, M KING, and L WAGONER. 2003. Micromorphology and OSL dating of the basal part of the loess-paleosol sequence at La Mesada in Tucuman province, Northwest Argentina. Quaternary International 106-107:111-117. [ Links ]
KEMP RA, M KING, PS TOMS, E DERBYSHIRE, JM SAYAGO, and M COLLANTES. 2004. Pedosedimentary development of part of a Late Quaternary loess-palaeosol sequence in northwest Argentina. Journal of Quaternary Science 19:567-576. [ Links ]
MARES MA, RA OJEDA, JK BRAUN, and RM BARQUEZ. 1997. Systematics, distribution, and ecology of the mammals of Catamarca Province, Argentina. Pp. 89-141, in: Life among the Muses: Papers in Honor of James S. Findley (TL Yates, WL Gannon, and DE Wilson, eds.). Museum of Southwestern Biology, University of New Mexico, Albuquerque. [ Links ]
MARSHALL L and T SEMPERE. 1991. The Eocene to Pleistocene vertebrates of Bolivia and their stratigraphic context: a review. Pp. 631-652, in: Fósiles y Facies de Bolivia. Vol. I - Vertebrados (R Suarez-Soruco, ed.). Revista Técnica de Yacimientos Petrolíferos Fiscales Bolivianos 12(3-4). [ Links ]
MARSHALL L, A BERTA, R HOFFSTETTER, R PASCUAL, OA REIG, M BOMBIN, and A MONES. 1984. Mammals and Stratigraphy: Geochronology of the continental mammal-bearing Quaternary of South America. Paleovertebrata, Mémoire Extraordinaire 1-76 [ Links ]
MUSSER GG and MD CARLETON. 2005. Superfamily Muroidea. Pp: 894-1531, in: Mammal Species of the World, a taxonomic and geographic reference (DE Wilson and DM Reeder, eds.). 3rd Ed. Smithsonian Institution Press, Washington. [ Links ]
ORTIZ PE. 2001. Roedores del Pleistoceno superior del valle de Tafí (Provincia de Tucumán), implicancias paleoambientales y paleobiogeográficas. Unpublished Ph.D. dissertation, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán. [ Links ]
ORTIZ PE. 2003. Fossil record and distribution of Cavia tschudii (Caviidae, Rodentia) in Argentina. Mammalia 67:607-611. [ Links ]
ORTIZ PE and UFJ PARDIÑAS. 2001. Sigmodontinos (Mammalia, Rodentia) del Pleistoceno tardío del Valle de Tafí (Tucumán, Argentina): taxonomía, tafonomía y reconstrucción paleoambiental. Ameghiniana 38:3-26. [ Links ]
ORTIZ PE, S CIRIGNOLI, DH PODESTÁ, and UFJ PARDIÑAS. 2000a. New records of sigmodontine rodents (Mammalia: Muridae) from high-Andean localities of northwestern Argentina. Biogeographica 76:133-140. [ Links ]
ORTIZ PE, UFJ PARDIÑAS, and SJ STEPPAN. 2000b. A new fossil phyllotine (Rodentia: Muridae) from northwestern Argentina and the relationships of Reithrodon group. Journal of Mammalogy 81:37-51. [ Links ]
PARDIÑAS UFJ, G D´ELÍA, and PE ORTIZ. 2002. Sigmodontinos fósiles (Rodentia, Muroidea, Sigmodontinae) de América del Sur: estado actual de su conocimiento y prospectiva. Mastozoología Neotropical 9:209-252. [ Links ]
PARDIÑAS UFJ and CA GALLIARI. 2001. Reithrodon auritus. Mammalian Species 664:1-8. [ Links ]
PARDIÑAS UFJ and PE ORTIZ. 2001. Neotomys ebriosus, an enigmatic South American rodent (Muridae, Sigmodontinae): its fossil record and present distribution in Argentina. Mammalia 65:244-250. [ Links ]
PINE RH, S MILLER, and M SCHAMBERGER. 1979. Contributions to the mammalogy of Chile. Mammalia 43:339-376. [ Links ]
SCHELLENBERGER A, F HELLER, and H VEIT. 2003. Magnetostratigraphy and magnetic suceptibility of the Las Carreras loess-paleosol sequence in Valle de Tafí, Tucumán, NW-Argentina. Quaternary International 106-107:159-167. [ Links ]
STEPPAN SJ. 1995. Revision of the tribe Phyllotini (Rodentia: Sigmodontinae), with a phylogenetic hypothesis for the Sigmodontinae. Fieldiana, Zoology, new series 80:1-112. [ Links ]
TETA P and PE ORTIZ. 2002. Micromamíferos andinos holocénicos del sitio arqueológico Inca Cueva 5, Jujuy, Argentina: tafonomía, zoogeografía y reconstrucción paleoambiental. Estudios Geológicos 58:117-135. [ Links ]
YACOBACCIO H. 1991. Sistemas de asentamiento de los cazadores-recolectores tempranos de los Andes centro-sur. Unpublished Ph.D. dissertation, Facultad de Filosofía y Letras, Universidad Nacional de Buenos Aires. [ Links ]
YEPES J. 1935. Consideraciones sobre el género " Andinomys " (Cricetinae) y descripción de una forma nueva. Anales del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" 38:333-348. [ Links ]
Recibido 14 noviembre 2006.
Aceptación final 16 marzo 2007.

