










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. v.14 n.2 Mendoza jul./dic. 2007
Bats of Saint Martin, French West Indies/Sint Maarten, Netherlands Antilles
Hugh H. Genoways,1 Scott C. Pedersen,2 Peter A. Larsen,3 Gary G. Kwiecinski,4 and Jeffrey J. Huebschman5
1 University of Nebraska State Museum, W436 Nebraska Hall, University of Nebraska-Lincoln, Lincoln, NE 68588 <hgenoways1@unl.edu>. 2 Department of Biology/Microbiology, South Dakota State University, Brookings, SD 57007 <Scott_Pedersen@sdstate.edu>. 3 Department of Biological Sciences and Museum, Texas Tech University, Lubbock, TX 79409 <plarsen01@yahoo.com>. 4 Department of Biology, University of Scranton, Scranton, PA 18510 <ggk301@Scranton.edu>. 5 Department of Biology, University of Wisconsin-Platteville, Platteville, WI 53818 <huebschj@uwplatt.edu>.
ABSTRACT: Six species of bats have been previously reported from the Antillean island of Saint Martin/Sint Maarten- Artibeus jamaicensis, Brachyphylla cavernarum, Molossus molossus, Tadarida brasiliensis, Noctilio leporinus, and Myotis nigricans nesopolus. Our field research reported herein documents an additional three species of bats from the island for the first time- Monophyllus plethodon, Ardops nichollsi, and Natalus stramineus. Re-examination of the single voucher of Myotis nigricans nesopolus has led us to exclude this species from the fauna of Saint Martin/Sint Maarten. Based on our field research and the study of specimens housed in museum collections, we present information on the eight species of bats that we have documented as occurring on Saint Martin/Sint Maarten. The average rate of fruit bat captures on Saint Martin/Sint Maarten (0.92 bats per net-night-BNN) falls towards the lower end of the range (0.65-2.47 BNN) reported from nearby islands in the northern Lesser Antilles and below the range (2.20-5.93 BNN) reported for mainland populations of Neotropical fruit bats. We discuss possible causes of these decreased population levels and we express some concerns about the future conservation status of the chiropteran fauna of the island.
RESUMEN: Murciélagos de Saint Martin, Indias Occidentales francesas/Sint Maarten, Antillas holandesas. Seis especies de murciélagos han sido registradas previamente para la Isla de Saint Martin/Sint Maarten, en las Antillas: Artibeus jamaicensis, Brachyphylla cavernarum, Molossus molossus, Tadarida brasiliensis, Noctilio leporinus y Myotis nigricans nesopolus. Nuestras investigaciones de campo ha permitido documentar por primera vez la presencia de tres especies adicionales para la Isla: Monophyllus plethodon, Ardops nichollsi y Natalus stramineus. La reexaminación de un único ejemplar de Myotis nigricans nesopolus nos llevó a excluir a esta especie de la fauna de Saint Martin/Sint Maarten. Nuestros estudios de campo y la examinación de ejemplares depositados en colecciones de museos nos ha permitido presentar información sobre las ocho especies de murciélagos que hemos podido documentar en Saint Martin/Sint Maarten. El promedio de captura de murciélagos frugívoros (0.92 murciélagos por noche - BNN) cayó hacia el extremo mas bajo del rango (0.65-2.47 BNN) reportado para islas cercanas al norte en las Antillas Menores, y por debajo del rango (2.20-5.93 BNN) reportado para poblaciones de murciélagos frugívoros neotropicales del continente. Se discuten las posibles causas de esas disminuciones en los niveles poblacionales y se presentan comentarios sobre nuestras preocupaciones sobre el futuro del estado de conservación de la fauna de murciélagos de la isla.
Key words. Chiroptera. Conservation. Distribution. Saint Martin. Sint Maarten.
Palabras clave. Chiroptera. Conservación. Distribución. Saint Martin. Sint Maarten.
INTRODUCTION
The island of St. Martin is the smallest island in the world divided between two sovereign nations. The northern half of the island is a part of France (Saint Martin), being administered as a commune of the overseas department of Guadeloupe. The southern half of the island is part of the Netherlands (Sint Maarten), being administered as part of the Netherlands Antilles through Curaçao. Although this separation was hardly perceptible while working on the island, it added a layer of administrative difficulty in conducting a survey of the chiropteran fauna of the entire island. Over a period of three years (2002-2004), we have documented eight species of bats from the island of which three are reported from Saint Martin/Sint Maarten for the first time (Monophyllus plethodon, Ardops nichollsi, and Natalus stramineus).
The first species of bat recorded from Saint Martin/Sint Maarten was based on individuals referable to Artibeus jamaicensis by Cope (1889). Subsequently, Husson (1960) in a review of the mammals of the Netherlands Antilles added three species of bats to the fauna (Brachyphylla cavernarum, Molossus molossus, and Tadarida brasiliensis) as well as presenting additional information on A. jamaicensis. Finally, Koopman (1968) added the greater fishing bat Noctilio leporinus to the fauna of the island and commented on the other four species known from the island. Husson (1960) also reported a single individual of the bat Myotis nigricans nesopolus; however, our re-examination of this specimen in light of 45 years of additional knowledge of the small Myotis in the Lesser Antilles and northern South America has led us to exclude this species from the fauna of Saint Martin/Sint Maarten. Here, we present information about eight species that comprise the current bat fauna occurring on Saint Martin/Sint Maarten based on our field research and study of specimens housed in museum collections. Based on our work, we present some concerns about the future conservation status of the chiropteran fauna of the island.
METHODS AND MATERIALS
Study area.- Saint Martin/Sint Maarten (18º05'N, 63º12'W) is part of the outer arc of islands of the northern Lesser Antilles. It is a non-volcanic island composed primarily of limestone and igneous diorite. St. Martin occurs on the same submarine plateau as Anguilla and St. Barthélemy and formed one island with them during the height of Pleistocene glaciations. The island is 15 km long and 14 km wide giving it an area of about 85 sq km. The highest point is Pic du Paradis at 425 m. The month with the highest mean temperature in Philipsburg is August at 27.9º C and the coolest months are January and February with a mean of 24.7º C (Stoffers, 1956). Mean annual rainfall is 1000- 1125 mm, but precipitation is very erratic from year to year and long periods of drought occur. Only two ravines contained moving water during our visits and one of these may have been flowing only because of sewage water being discharged into it. Saint Martin/Sint Maarten lies in the Caribbean hurricane zone with the late 1990s being the most active period since the keeping of records (1851) with five named hurricanes-Luis, 1995; Bertha, 1996; Georges, 1998; Jose and Lenny, 1999 (Caribbean Hurricane Network, 2005). The most recent storm to hit the island was Debby in 2000. The most active period for storms for the island is the first week of September (Caribbean Hurricane Network, 2005).
During the early colonial period, the island was under intense agricultural development with sugar cane grown almost to the tops of the hills. Today agriculture has been reduced to a few subsistence areas and pastures. Because of this earlier cultivation, the island is covered with secondary vegetation characterized by no definite structure and wide variation in composition. The few areas of original vegetation are located on the high hills and peaks and these are coming under assault today from development. The primary original vegetation was probably semi-evergreen seasonal forest (Stoffers, 1956). At present, the island is under intense development pressure to expand tourism and to provide space for residential and investment housing and condominiums.
Collecting methods.- Our mist-netting on Saint Martin/Sint Maarten occurred during three periods-12 to 17 July 2002, 31 net nights; 20 to 23 May 2003, 20 net nights; and 14 to 19 May 2004, 35 net nights (86 net nights total). M ist-netting for bats was conducted in a variety of habitats, including ravines, plantations, covered flyways, trails, and access roads. Due to the season and the dry nature of the remaining forest on this island, very few locations had any appreciable fruit in evidence. Five to eight mist-nets of varying lengths were erected at each location and frequently were situated diagonally across a trail or ravine at 20 to 100 m intervals and monitored for four to six hours depending on bat activity and weather.
Caves and rock faces were surveyed for roosting bats. Two active roosts were found-Grotte du Puits des Terres Basses (0.25 km S, 1 km W Mornes Rouges) in the lowlands of the western portion of French St. Martin and a cave complex in the limestone outcrop (Billy Folly) at the eastern edge of Simpson Bay in the southwestern part of Dutch Sint Maarten. Grotte du Puits was visited in 2002 and 2004, and Billy Folly was visited all three years. A third large cave, Devil's Hole, was described by Hummelinck (1979). The fate of this cave is discussed in detail in the species account for Tadarida. Numerous small caves, rock outcrops, and rock shelters around Terres Basses, Cul-de-Sac, and Pic du Paradis were searched for evidence of bat roosts as time permitted during each survey but without effect.
Measurements.- Mist-netted bats were measured and examined at the end of the evening and body mass (g), length of forearm (mm), reproductive status, tooth wear, presence of scars, and external parasites were recorded. Length of forearm and cranial measurements (mm) were also taken from museum specimens using digital calipers and weights were recorded in grams. Measurements were taken following Hall (1946), except that greatest length of skull included the incisors, and length of forearm is the distance from olecranon process to tip of carpals with the wing in retracted position. StatView® software package (Sager, 1992) provided standard statistics for each sample and paired t-tests were used to test for differences between groups.
Voucher specimens.- A survey of existing collection materials in natural history museums (American Museum of Natural History, New York, AMNH; Muséum National d'Histoire Naturelle, Paris, France, MNHN; Nationaal Natuurhistorisch Museum, Leiden, The Netherlands, RMNH) yielded an additional 63 specimens of bats from Saint Martin/Sint Maarten. All voucher specimens from the 2002 survey were deposited in the research collections at the University of Nebraska State Museum (UNSM) and those from 2003 and 2004 were deposited in the mammal collections of the Museum of Texas Tech University (TTU).
SPECIES ACCOUNTS
Noctilio leporinus mastivus (Vahl, 1797)
Greater Fishing Bat
Specimens examined (2).-SAINT MARTIN: Grotte du Puits des Terres Basses, 0.25 km S, 1 km W Mornes Rouges, 18º03'48.3"N, 63º08'13.3"W, 33 m, 2 (TTU).
Specimens captured/released (5).-SAINT MARTIN: Grotte du Puits des Terres Basses, 0.25 km S, 1 km W Mornes Rouges, 18º03'48.3"N, 63º08'13.3"W, 33 m, 5.
Additional record.-UNKNOWN: no specific locality (Koopman, 1968).
The greater fishing bat was known previously from St. Martin based on a single individual deposited in the Academy of Natural Sciences of Philadelphia from an unknown location on the island (Koopman, 1968). Davis (1973) assigned circum-Caribbean specimens to the subspecies N. l. mastivus, which was originally described from St. Croix, U. S. Virgin Islands. We have followed this arrangement here. Length of forearm and cranial measurements of our male and female specimens are given in Table 1. These measurements are within the range of those given by Davis (1973) for a sample from the West Indies. Our male specimen was larger than the female in all measurements recorded.
Table 1
Length of forearm and seven cranial measurements (in mm) for seven species of bats from Sint Maarten, Netherlands Antilles/Saint Martin, French West Indies.
Our two specimens were taken on 17 July 2002 in Grotte du Puits des Terres Basses. A general description of this cave is given by Hummelinck (1979). The limestone outcrops throughout this region are covered with low, thorny dry scrub forest that is dense in many areas. The cave occurs at the base of a high limestone cliff (15 m). The cave entrance is about 7 m wide and leads to two rooms in which the ceiling is about 8 m high. This cave was mined for phosphate in the late 19th century and evidence of this work is quite noticeable. In undisturbed portions of the cave, there are deep crevices and rounded solution cavities occupied by bats. Seven individual fishing bats, of which two were preserved and five were released, were taken from one of the highest solution pockets in the western chamber of the cave in 2002. Temperature near the roosting fishing bats was 29º C and the relative humidity was 76%. Other bats roosting in this cave in 2002 and 2004 were A. jamaicensis and B. cavernarum. In 2002, we spotlighted the two saline ponds at Little Bay for the presence of foraging N. leporinus without effect.
Of four females captured two were lactating and two did not evince any gross reproductive activity. Testes lengths of three males were 5, 6, and 8. The four females averaged 58.4 (55.8-61.6) in weight, whereas the three males weighed 71.9, 74.5, and 78.1.
Monophyllus plethodon luciae Miller, 1902
Insular Long-tongued Bat
Specimens examined (13).-SINT MAARTEN: Billy Folly Cave, 18º01'48"N, 63º05'26"W, 104 m, 12 (7 TTU, 5 UNSM); Romeo's Drive, Middle Region, 18º02'50"N, 63º02'35"W, 74 m, 1 (UNSM).
The 2002 capture of a single male near Middle Ground represents the first record of the long-tongued bat from Sint Maarten/Saint Martin. The species is known from the nearby islands of Anguilla (Schwartz and Jones, 1967; Koopman, 1968), St. Kitts (Pedersen et al., 2005), and Nevis (Pedersen et al., 2003) so its presence on the island was not unexpected. The species usually frequents dry habitats where caves are available for daytime roosts. Forearm and cranial measurements of 5 males and one female are given in Table 1. These measurements when compared with a sample from Guadeloupe (Baker et al., 1978) shows that the lower end of the range of measurements matches closely between the samples, but individuals at the upper end of the range of measurements are larger on Sint Maarten/Saint Martin for all measurements except for condylobasal length and mastoid breadth. Single individuals from Nevis (Pedersen et al., 2003) and Montserrat (Pedersen et al., 1996) fall below the range of variation of the sample from Sint Maarten/Saint Martin for length of forearm and greatest length of skull for both individuals and for mastoid breadth and length of maxillary toothrow for the female from Montserrat. We follow Schwartz and Jones (1967) in placing all populations of this species from St. Lucia northward in the Lesser Antilles in the subspecies M. p. luciae, but we recognize that the variation represented among samples in the northern Lesser Antilles call for a revaluation of geographic variation now that larger samples are available. The single female was larger than the mean of the five males in three measurements (length of the forearm, condylobasal length, and length of maxillary toothrow).
Apart from the single male captured near Middle Ground on 14 July 2002, all other specimens were taken at Billy Folly Cave. At Billy Folly Cave, 11 of the 12 specimens taken during visits in July 2002 and May 2003 were males. The single female was captured on 15 July 2002. This segregation of sexes probably indicates that this species forms maternity colonies for reproductive females and their young.
The single male taken from Middle Ground was netted within a grove of genip trees (Melicoecus bijugatus). This locality will be fully described in the species account for Artibeus jamaicensis. The cave complex near Billy Folly is situated within a large limestone outcrop that overlooks both Simpson and Lay bays. This buttress is honeycombed with numerous caves and solution pockets, none of which are readily accessible due to their location within a cliff face or are protected by dense thickets of thorny scrub. Bats were located in an interconnected pair of caves on the western rim of the buttress. The larger, southern-most chamber was well lighted with large openings to the outside through the ceiling and western wall and was littered with bat guano and bones of goats and iguanas. This cavity is extensive with numerous side chambers that penetrate horizontally back into the center of the buttress. Bats were found roosting in the second, northern chamber in both 2002 and 2003. This second chamber was accessible only by a very narrow vertical chimney that descended down into a room of approximately 50 cubic meters. The room was entirely dark with a relative humidity of 89% and a temperature of 29º C while the outside temperature was 32º C. We noted approximately 25 bats in 2002 and only 12 in 2003.
Seven males collected on 21 May 2003 had testes lengths that averaged 3.7 (3-5), whereas five taken on 15 July 2002 had mean testes lengths of 4.5 (3.5-5). The 12 males weighed an average of 16.1 (14.5-18.1). Our single female revealed no gross reproductive activity and weighed 17 when captured.
Brachyphylla cavernarum cavernarum Gray, 1834
Antillean Cave Bat
Specimens examined (51).-SAINT MARTIN: Grotte du Puits des Terres Basses, 0.25 km S, 1 km W Mornes Rouges, 18º03'48.3"N, 63º08'13.3"W, 33 m, 7 (TTU); upper Loteríe Farm, 0.5 km S, 1 km E Rambaud, 18º04'41.4"N, 63º03'22.8"W, 75 m, 8 (3 MNHN, 5 TTU); northeastern slope Pic du Paradis, 18º04'43.4"N, 63º02'54.1"W, 410 m, 1 (TTU). SINT MAARTEN: Lowlands, 35 (AMNH).
Specimens captured/released (10).- SAINT MARTIN: Grotte du Puits des Terres Basses, 0.25 km S, 1 km W Mornes Rouges, 18º03'48.3"N, 63º08'13.3"W, 33 m, 10.
Additional records.- SINT MAARTEN: small cave at Billy Folly (Rojer, 1997: 8); Terre Basse (Timm and Genoways, 2003).
Husson (1960) first reported the Antillean cave bat from this island based on four specimens from Grotte du Puits des Terres Basses on the French side of the island. The species also has been reported from the island by Koopman (1968) and Swanepoel and Genoways (1978) who reported specimens from Lowlands on the Dutch side of the island. Our work has added material from two additional localities both on the French side of the island. Swanepoel and Genoways (1978) assigned members of this species from St. Croix southward to St. Vincent to the nominate subspecies and we have followed that arrangement here. Table 1 presents length of forearm and cranial measurement for 5 males and 5 females from St. Martin. Not surprisingly, the measurements of these specimens closely match the values for sample 19 in the study by Swanepoel and Genoways (1978), which was composed of specimens from St. Martin. Males in our sample were significantly larger than females in greatest length of skull and condylobasal length at the P £ 0.05 and P £ 0.01 levels, respectively. Males also averaged larger than females in all other measurements studied.
We found Antillean cave bats in only one cave, Grotte du Puits des Terres Basses, which is a site known to earlier researchers (Husson, 1960). During our visit in 2002, Brachyphylla were found in both chambers of the cave, but in 2004, the maternity colony was confined to the smaller eastern chamber. In contrast to the dry well, lighted western chamber, the eastern chamber was darker, more humid, and had a damp floor covered in guano. Both chambers showed evidence of human disturbance; long wooden poles capable of reaching the roosting bats were left leaning against the walls of both chambers in the vicinity of the roosting bats.
We are not certain of the conditions under which others took individuals at Lowlands, but circumstantial evidence suggests that they were taken from a cave near the south end of Juliana Airport. The original vegetation in this area was probably as around Grotte du Puits des Terres Basses, being low, thorny, dry scrub forest, but the area has become so developed that much of the original vegetation is now gone. This sample includes 17 males and 18 females suggesting that only selected individuals were preserved, but it does indicate that pregnant females and adult males were taken together at this site.
Rojer (1997) reports and presents a photograph of a colony of around 300 individuals of B. cavernarum "in a small cave at Billy Folly." We explored several small caves around Billy Folly point during our field studies in the summers of 2002, 2003, and 2004. The only species of bat that we found occupying these caves was M. plethodon and we never found them in numbers exceeding 25 individuals. We did expend considerable time sifting through guano and dirt to identify bat remains but found little more than the fragmented skulls of 2 or 3 B. cavernarum.
We also captured foraging individuals on two occasions on the slopes of Pic du Paradis. We captured a single adult male along the northeastern slope on the night of 14 May 2004. This individual was taken in a mist net set across the access road to the top of Pic du Paradis on an unseasonably cold night (18.5 Cº). This area was windswept and the vegetation evinced some wind shaping ("krumholz" effect). On the nights of 15 and 16 July 2002, eight B. cavernarum were netted along the western slope of Pic du Paradis in the upper reaches of what is known as Loteríe Farm.
Loteríe Farm can best be described as a refuge protected by its owner in an attempt to preserve the remaining native vegetation on St. Martin and, where possible, the owner is trying to restore native vegetation with an aggressive replanting/reforestation effort. This was the only area that we observed on the island that provided good habitat for bats and other fauna. The deep valley has a small, seasonal running stream and several small catchement pools of water. The flora in this area varied from evergreen to semi-evergreen seasonal forest (Rojer, 1997; Stoffers, 1956). Antillean cave bats were taken in nets set across one of the numerous foot trails on the valley floor and across pools of water. As is typical of the species, large mobs of animals would forage from tree to tree moving up and then down the valley.
Of eight females captured on 30 March 1926 that were examined for reproductive activity, seven were pregnant with embryos averaging 17.4 (13-20) in crown-rump length and one evinced no reproductive activity. Four males from this date had testes that averaged 6.0 (5-8) in length. On 16 May 2004, three lactating females were taken from a maturity colony in Grotte du Puits des Terres Basses. Associated with these females was a newborn male that weighed 14.7 and with a forearm of 36.4. This individual lacked hair over all of its body. Two slightly older young bats had hair visible on the dorsum but none on the venter. These two males weighed 18.0 and 15.8 and had forearms of 41.1 and 40.5. A male taken on the northeastern slope of Pic du Paradis on 15 May 2004 had testes that measured 7 in length. Between 15 and 17 July 2002, two females were captured that were lactating and one revealed no gross reproductive activity. Six males taken during this period had a mean testes length of 6.7 (5.5-9.5). Earlier, Husson (1960) noted that hairless young were found in mid-July in Grotte du Puits des Terres Basses. It appears that B. cavernarum has a reproductive cycle on St. Martin that includes fertilization in February and March, pregnancy in March, April, and May, and parturition extending at least from mid-May to mid-July. This is less synchrony than we anticipated in the reproductive cycle of this bat and there is always the possibility that there are two closely spaced periods of parturition occurring in this cave bat (Wilson, 1979). More data will be needed to resolve this issue.
Seven males captured in May and July had a mean weight of 48.7 (46.0-52.6) and six nonpregnant females taken at these times weighed 48.0 (40.5-53.9) on average.
Ardops nichollsi montserratensis (Thomas, 1894)
Lesser Antillean Tree Bat
Specimens examined (2).-SAINT MARTIN: lower Loteríe Farm, 0.5 km S, 0.5 km E Rambaud, 18º04'40.6"N, 63º03'45.9"W, 59 m, 1 (TTU); upper Loteríe Farm, 0.5 km S, 1 km E Rambaud, 18º04'41.4"N, 63º03'22.8"W, 75 m, 1 (TTU).
The discovery of the Lesser Antillean tree bat is important because Ardops, despite the fact that it has been found on several islands of the inner arc of the northern Lesser Antillean islands, such as Montserrat (Pedersen et al., 1996; Jones and Baker, 1979), Nevis (Pedersen et al., 2003), St. Kitts (Pedersen et al., 2005), and St. Eustatius (Jones and Schwartz, 1967), has not been reported from any islands in the outer arc, including Antigua (Pedersen et al., 2006), Barbuda, Saint Barthélemy, and Anguilla. Length of forearm and cranial measurements of the two females that were captured are given in Table 1. These measurements are slightly smaller than those of a female from Montserrat reported by Pedersen et al. (1996), but closely match those of a female reported from Nevis by Pedersen et al. (2003). We have tentatively assigned our specimens from Saint Martin to A. n. montserratensis on geographic grounds (Jones and Schwartz, 1967) and the matching of these measurements, but we recognize a new revision of this genus would be appropriate.
It is our belief that the Lesser Antillean tree bat occupies a very limited area on Saint Martin that may cover as little as 1.5 square kilometers along the western slope of Pic du Paradis (Loteríe Farm). This area probably extends just to the north of the Ravine de Loteríe and southward to include the upper slopes of the valley that extends to the south and east of Colombier. We do not believe that appropriate habitat extends into Sint Maarten so the species may only be found in Saint Martin. In the area above the great house for Loteríe Farm, the habitat was the least altered that we observe anywhere on the island, some of which we believe may be primary forest (Rojer, 1997; Stoffers, 1956). It was in this patch of forest that the two Ardops were captured on successive nights (16 and 17 May 2004) in mist nets that were located within 100 m of each other along the trail in the bottom of the ravine. This suggests that a favored fruit tree or possibly a roost was located nearby. Both specimens examined were females and weighed 24.0 and 19.6, respectively. Neither revealed gross reproductive activity.
Artibeus jamaicensis jamaicensis Leach 1821
Jamaican Fruit-eating Bat
Specimens examined (68).-SAINT MARTIN: Grotte du Puits des Terres Basses, 0.25 km S, 1 km W Mornes Rouges, 10 (TTU); lower Loteríe Farm, 0.5 km S, 0.5 km E Rambaud, 18º04'40.6"N, 63º03'45.9"W, 59 m, 6 (2 MNHN, 4 TTU); upper Loteríe Farm, 0.5 km S, 1 km E Rambaud, 14 (TTU); northeastern slope Pic du Paradis, 18º04'43.4"N, 63º02'54.1"W, 410 m, 6 (TTU); SW slope Pic du Paradis, 2 (TTU). SINT MAARTEN: Devil's Hole, Simpson Bay, 4 (AMNH); Mary's Fancy, 0.5 km S Cul de Sac, 18º02'29"N, 63º03'47"W, 26 m, 2 (1 TTU, 1 UNSM); Lowlands, 4 (AMNH); Philipsburg, 11 (AMNH); Romeo's Drive, Middle Region, 18º02'50"N, 63º02'35"W, 74 m, 9 (3 TTU, 6 UNSM).
Specimens captured/released (27).- SAINT MARTIN: Grotte du Puits des Terres Basses, 0.25 km S, 1 km W Mornes Rouges, 6; lower Loteríe Farm, 0.5 km S, 0.5 km E Rambaud, 17; upper Loteríe Farm, 0.5 km S, 1 km E Rambaud, 4.
Additional records.- SAINT MARTIN: Deux Frères (Rojer, 1997: 8). SINT MAARTEN: Belvédère (Rojer, 1997: 8); St. Peter (Husson, 1960); Simpson Bay (Husson, 1960).
This was the most common and widespread species of bat occurring on the island during our work. The Jamaican fruit-eating bat was the first species of bat known from St. Martin when Cope (1889) described two adult males from the island as a new species under the name Dermanura eva. Rehn (1901) described the cotypes in the collections of the Academy of Natural Sciences of Philadelphia in more detail using the name Artibeus eva. Rehn (1901) provides the length of forearm of one cotype (ANSP 5784) as 62 and cranial measurements of the second cotype (ANSP 5783) including greatest length of skull 27.6 and zygomatic breadth 17.0. These measurements fall within the range of variation for our specimens from the island (Table 1) and support the placement of Dermanura eva as a junior synonym of Artibeus jamaicensis as was first done by Andersen (1908) and was continued by Hall (1981). Subsequently, Husson (1960) and Koopman (1968) have reported additional specimens of this species from the island. Length of forearm and cranial measurements for 5 males and 5 females are given in Table 1. These measurements reveal no significant secondary sexual differences between individuals in this sample. Genoways et al. (2001) have reviewed morphological variation in Antillean populations of this species. Our sample from Sint Maarten/Saint Martin (Table 1) most closely matches the measurements of other samples of A. j. jamaicensis; in fact, the mean values of our sample are almost an exact match for the means of samples from Puerto Rico and Dominica (Genoways et al., 2001).
A maternity colony of Jamaican fruit-eating bats was discovered in the eastern room of Grotte du Puits des Terres Basses where we also found the greater fishing bats. Directly beneath the roosting bats, the floor of this well-lit space was covered with almond seeds (Terminalia cattapa) and mango pits (Mangifera indica). Among the females present, some had not given birth while others were carrying newborn on 17 July 2002. Jamaican fruit-eating bats were roosting in the same general area as the fishing bats, but in a lower part of the ceiling and thus were more easily disturbed. We examined four specimens that were taken from the Devil's Hole cave near Simpson Bay (Hummelinck, 1979), but this valuable cave no longer exists (Rojer, 1997; see species account for Tadarida). Among the specimens of this species preserved in the American Museum of Natural History is a non-volant young male from Devil's Hole, which would indicate that Jamaican fruit-eating bats reproduced in this cave. The conditions under which Artibeus from Lowlands and Philipsburg were collected are unknown.
The remaining specimens taken during our research were netted at night in foraging areas. The species seemed particularly abundant in the area of Pic de Paradis and Loteríe Farm. The native vegetation was less disturbed in these areas than elsewhere on the island, with native fruit trees and bushes present as well as a few introduced fruit trees such as mango and banana (Musa). At Loteríe Farm, fruit bats were taken at lower elevations where the vegetation was more open and non-native fruit trees were more abundant. However, individuals also were captured high on the northeastern slope of the peak as they flew along an access road on Pic du Paradise.
The specimens from Middle Ground were taken in nets set within a small grove of genip trees below Romeo Drive in a ravine called Cherry Gut by the local residents. Cherry Gut, which was criss-crossed by foot paths and contained accumulations of human refuse indicating that it was being used as a landfill site, was the most highly disturbed habitat that we worked on the entire island. Low scrub and weedy plants covered the flanks of the ravine, and over-browsing by goats prevented all but the mature genips from growing in this area. The fact that we collected nine individuals of the Jamaican fruit-eating bat in two years of work in this region is a tribute to the ability of this species to tolerate human disturbance. Certainly, Artibeus feeds on genip fruit, which may draw the bats into this orchard, but the trees also provide a sheltered corridor between Middle Region and the nearby lowlands of the eastern part of the French side of the island.
Mary's Fancy is an idiosyncratic semi-abandoned cluster of cottages, trails, a swimming pool, stream, and a mature collection of large trees and flowering plants including almonds, torchwood (Fagara trifoliate), Barbados cherry (Malpighia puncicifolia), coconut palms (Cocos nucifera), banana, mangrove (Rhizophora mangle and Avicennia nitida), and other small native trees and shrubs. It exists as an oasis for wildlife amidst the urban clutter of Cul-du-Sac. However, only two Jamaican fruit-eating bats were captured here in two nights of netting in 2002 and 2003 a result probably due to heavy road traffic nearby, little fruit on the trees, and dense vegetation, which denied the bats access to any available understory fruits.
An adult female taken on 26 March 1926 showed no signs of reproductive activity as did a female taken on 30 March 1926, but a second female from this date was lactating. All seven of the adult females taken on 13 April 1926 in Philipsburg were lactating. Three subadult individuals from these dates are preserved in collections. Of the five females taken on 15-16 May 2004, one was lactating and the other four were pregnant with single embryos that averaged 26.3 (23-32) in crown-rump length. Eight females were captured during our work between 13 and 17 July 2002. Of these 8 females, four were pregnant, with two having near-term embryos that measured 38 and 40 in crown-rump length, whereas two of the other females evinced no gross reproductive activity and two were lactating. One of the lactating females from Grotte du Puits des Terres Basses on 17 July 2002 was accompanied by a young pup that had a forearm length of 39 as compared with mother's 62. A nearly naked juvenile male taken on 13 April 1926 had a forearm length of 35.5, whereas a young male from this date was covered in juvenile pelage and possessed a forearm that was 54.2 long. Another young male taken from the Devil's Hole on 26 March 1926 was covered in juvenile pelage, but had a forearm that was only 47.1 long.
Adult males had testes lengths of 6 and 8 on 30 March 1926 and 7 on 13 April 1926. Five adult males taken from Pic du Paradis and Loteríe Farm, Saint Martin, on 15-16 May 2004 had testes that had a mean length of 6.5 (6-8), whereas four adult males collected on 21-22 May 2003 from Mary's Fancy and Middle Region, Sint Maarten, had testes that averaged 8.2 (7-10).
The Jamaican fruit-eating bat is known to follow a bimodal polyestrous reproductive cycle in Central America (Wilson, 1979) and Jamaica (Genoways et al., 2005). On Jamaica pregnant females were present in January, February, and March and in July and August and parturition was expected in March-April and early May and July, August, and September (Genoways et al., 2005). The reproductive data for this species on Sint Maarten/Saint Martin matches the bimodal pattern in data for March-April and July reproduction, but finding pregnant females in mid-May does not match the bimodal pattern. Having four of five females pregnant at this time is surprising. Given the size of the embryos, they were not near-term, but certainly would have been born before July. This may indicate that the bimodal polyestrous cycle is not as highly synchronous on St. Martin as it is elsewhere in the geographic range of the species. Alternatively, this could represent a breakdown in the bimodal polyestrous reproductive cycle with females producing as many young in a year as possible in response to low population levels on the island or resource availability (Fleming, 1971). This observation deserves further study to verify its occurrence and to clarify its significance.
Nine males taken in mid-May had a mean weight of 41.0 (37.9-45.5). A lactating female captured on 16 May weighed 40.4 and four females with mid-term embryos based on crown-rump length weighed 46.7, 46.8, 49.1, and 54.4. Twelve July-taken adult males weighed an average of 41.2 (36.3-46.7), whereas two non-pregnant adult females taken during this time period weighed 36.0 and 42.0 and a lactating female weighed 43.4. Four females with near-term embryos when captured between 14 to 17 July weighed 55.5, 57.0, 57.2, and 57.6.
A lactating adult female (AMNH 72482) captured on 13 April revealed evidence of alopecia on the top and sides of its head as well as on the chin and extending onto the upper chest area. An adult female (AMNH 72261; length of forearm = 59.3) taken at Devil's Hole on 26 March had heavily worn dentition, which appeared to be the result of advanced age of the individual. An adult male taken at Middle Region (TTU 102081) was missing the tip of its upper left canine tooth. The tooth was either broken or had unusually heavy wear, and evidently was infected with the abscess draining through the external side of the alveolus just lateral to the nares.
The presence of the M3/m3 is known to vary in populations of Artibeus jamaicensis (Genoways et al., 2001). In 10 specimens that we examined from Sint Maarten/Saint Martin, the upper M3 was absent in all individuals and the lower m3 was present in seven individuals, absent in both lower jaws of one individual, and was present in the right dentary and absent in the left dentary of two individuals. The upper M3 being absent matches other populations of A. j. jamaicensis, but the percentage of individuals with the lower m3 present is below the 90% or higher levels in other samples of this taxon.
Natalus stramineus stramineus Gray 1838
Lesser Antillean Funnel-eared Bat
Specimens examined (0).
Additional record.-SINT MAARTEN: Billy Folly Cave, 18º01'48"N, 63º05'26"W, 104 m.
During the first visit to the Billy Folly Cave system in 2002, one member of our party (Pedersen) observed and attempted to collect the skeletal remains of a small bat in the southernmost chamber. The partial cranium, jaw, and some post-cranial elements were located on a protected small rock shelf about a meter above the floor of the cave. Although these fragments proved to be too fragile for collection, Pedersen was able to identify them as Natalus stramineus based on the distinctive shape of the cranium and dentary. This portion of the cave was relatively open and a headlight was not necessary to see the remains. This is not the typical cave situation in which we have observed colonies of Natalus on other islands in the northern Lesser Antilles.
Dávalos (2005) and Tejedor et al. (2005) presented evidence that unrecognized taxa of Natalus exist within currently named populations in the West Indies. Goodwin (1959), in his revision of members of the subgenus Natalus, restricted the type locality of Natalus stramineus to Antigua rather than Lagoa Santa, Minas Gerais, Brazil, as was done by earlier authors. This decision later was reconfirmed by Handley and Gardner (1990) and Tejedor (2006). Tejedor (2006) restricted N. stramineus to the northern Lesser Antilles, ranging from Dominica to Anguilla, and in a canonical analysis of external and cranial characters, demonstrated that "the sample from Dominica is morphometrically distinct from that of the remaining islands." Although Tejedor (2006) did not use it, the name N. s. dominicensis Shamel (1928) is available for the population on Dominica, leaving the nominate subspecies, N. s. stramineus, as the appropriate name to apply to the other populations in the northern Lesser Antilles, including those on Sint Maarten/Saint Martin.
We are not able to state with certainty whether populations of Natalus are extant on the island. We believe that the remains observed in Billy Folly Cave had been deposited there in the last 50 years, but with the environmental changes that have occurred on Sint Maarten/Saint Martin over this period of time, this species could have been lost from the fauna. Our work revealed a limited number of caves on the island and Hummelinck (1979) described only two caves on the island. The loss of the largest cave, Devil's Hole (or Airmouse Cave), could have had a negative impact on the status of this species, which is dependent on dark, moist caves as day roosts (Genoways et al., 2001).
Tadarida brasiliensis antillularum (Miller, 1902)
Brazilian Free-tailed Bat
Specimens examined (6).-SAINT MARTIN: lower Loteríe Farm, 0.5 km S, 0.5 km E Rambaud, 18º04'40.6"N, 63º03'45.9"W, 59 m, 4 (TTU). SINT MAARTEN: Mary's Fancy, 0.5 km S Cul de Sac, 18º02'29"N, 63º03'47"W, 26 m, 2 (UNSM).
Additional record.-SINT MAARTEN: Bat Cave, Devil's Hole, Simpson Bay (Husson, 1960; Hummelinck, 1979).
This free-tailed bat was first reported from Sint Maarten by Husson (1960) based on 24 female specimens collected in the Bat Cave at Devil's Hole by Hummelinck (see Hummelinck, 1979). Although this large collection from a single place on Sint Maarten would seem to indicate that Tadarida was abundant on the island, we found the species to be quite rare being represented by only 6 individuals from two localities, one located on each side of the island. Length of forearm and cranial measurements for 5 males and 1 female are reported in Table 1. The length of forearm of specimens from Sint Maarten/Saint Martin appears to average larger than those of a sample from the island of Dominica (Genoways et al., 2001), but the cranial measurements seem to match closely between these samples. We continue to follow Schwartz (1955) in assigning all Lesser Antillean population of this species to the subspecies T. b. antillularum until a comprehensive study of geographic variation in Antillean populations is undertaken. Owen et al. (1990) presented evidence of a close relationship between T. b. cynocephala from the southeastern United States and Antillean populations of T. brasiliensis. Further data are needed to fully explore the taxonomic and biogeographic implications if such a relationship exists.
The measurements of our female specimen were less than the mean of the sample of males for four measurements-length of forearm, greatest length of skull, condylobasal length, and zygomatic breadth, but only in the last measurement was its value below the range of measurements for the males. In the other four measurements, the values for the female were the same as the mean for the sample of males.
The single most important known site for Brazilian free-tailed bats on the island was the Bat Cave at Devil's Hole. This cave is described in detail by Hummelinck (1979). He found this natural cave to consist of a single room of about 25 by 15 m. The room was accessible because part of the ceiling had collapsed, but bowl-shaped solution holes were evident in the remaining portions of the original roof. The cave had been mined in the late 19th century so its original shape was unknown. The floor of the cave was covered in guano and pools of water. Some of these pools were tidal/brackish because the eastern shore of Simpson's Bay was approximately 100 m from the cave. This main chamber contained a nursery colony composed primarily of adult female Tadarida brasiliensis. Hummelinck (1979) mentions that the Devil's Hole had been "adapted to tourist purposes" just before the appearance of his publication. Hummelinck (1979) also describes two near-cylindrical mining shafts in the immediate vicinity of the Bat Cave but these shafts contained no bats.
We were unable to find Devil's Hole during our field research in 2002 and 2003, but we continued to try to determine its location during our third season of work in 2004. The next to the last day of our work on the island we came to an active construction site in the quickly developing housing area above the southeast shore of Simpson's Bay. We inquired of some of the workers at the site about their knowledge of the Devil's Hole. They quickly responded that it was here in front of us. According to their account, the main part of the cave had been filled-in during the early 1990s; however, the two mine shafts still remained although they were being backfilled as we watched. We were able to quickly explore them and found that the entrances were about 75% filled with rock rubble and trash. There were no bats in these mine shafts and there was no evidence of guano on the floors, apparently the bats did not relocate here when the main chamber was filled in. The loss of this maternity cave due to resort development must have been a major blow to the population of the Brazilian free-tailed bats on the island and undoubtedly accounts, at least in part, for why this free-tailed bat is relatively rare on St. Martin.
During our work in Sint Maarten, we captured two adult male Brazilian free-tailed bats on 13 July 2002 in a 10-m mist net that was set over the abandoned swimming pool at Mary's Fancy. The pool was located in an opening to the sky in this heavily wooded area. The pool was half-filled with one end covered in water hyacinths. Four individuals of Pallas' mastiff bats (M. molossus) also were taken over the pool. When we returned to this pool in 2003, it was completely covered in water plants and no bats were captured.
On the French side of the island, we captured four individuals at Loteríe Farm-two males and a female on 16 May 2004 and one male on 18 may 2004. They were all taken in nets set in open areas between widely scattered large trees in the vicinity of the farm buildings. Short grass covered the ground between the trees. Specimens of M. molossus and A. jamaicensis also were taken in this area.
The testes lengths of three May-taken males were 2.5, 3.0, and 3.0 and those of two July-taken males were 2.0 and 2.5. The single female in our sample was taken on 16 May 2004, evinced no gross reproductive activity, and weighed 9.1. The weights of the five males were, respectively, 9.2, 8.8, 9.1, 9.4, and 9.4.
Molossus molossus molossus (Pallas, 1776)
Pallas' Mastiff Bat
Specimens examined (23).-SAINT MARTIN: lower Loteríe Farm, 0.5 km S, 0.5 km E Rambaud, 18º04'40.6"N, 63º03'45.9"W, 59 m, 8 (1 MNHN, 7 TTU); northeastern slope Pic du Paradis, 18º04'43.4"N, 63º02'54.1"W, 410 m, 1 (TTU); southwestern slope Pic du Paradis, 18º04'N, 63º03'W, 350 m, 2 (TTU). SINT MAARTEN: Mary's Fancy, 0.5 km S Cul de Sac, 18º02'29"N, 63º03'47"W, 26 m, 6 (2 TTU, 4 UNSM); Philipsburg, 2 (AMNH); Romeo's Drive, Middle Region, 18º02'50"N, 63º02'35"W, 74 m, 4 (1 TTU, 3 UNSM).
Specimens captured/released (33).-SAINT MARTIN: lower Loteríe Farm, 0.5 km S, 0.5 km E Rambaud, 18º04'40.6"N, 63º03'45.9"W, 59 m, 33.
Additional record.-SINT MAARTEN: Philipsburg (Husson, 1960); ruins of "One Hoe" [18º03'N, 63º02'W] (Rojer, 1997: 8).
Husson (1960) was the first to report this free-tailed bat from Sint Maarten based on 21 specimens collected by Hummelinck in Philipsburg in 1955. Koopman (1968) reported an additional two specimens from Sint Maarten in the collection of the American Museum of Natural History, which undoubtedly were the two specimens from Philipsburg listed above. This mastiff bat was more abundant and widespread on the island during our work than the other free-tailed species, T. brasiliensis.
Husson (1962) restricted the type locality of M. molossus to the island of Martinique, which lead Dolan (1989) to apply the name M. m. molossus to this species throughout the Lesser Antilles. Length of forearm and cranial measurements for five males and five females from Sint Maarten/Saint Martin are given in Table 1. These measurements compare favorably with samples from Antigua (Pedersen et al., 2006), Nevis (Pedersen et al., 2003), and St. Kitts (Pedersen et al., 2005). Based on the work of these authors, we have assigned our material to the nominate subspecies. Males were significantly larger than females only for condylobasal length at the P £ 0.05. Males averaged larger, although not at a significance level equal to or below P £ 0.05, than females in all other measurements except length of forearm and postorbital constriction for which the sexes had the same mean values.
All of the specimens of Pallas' mastiff bat taken during our work were netted in foraging areas. However, Husson (1960) reported that the first specimens collected from the island were taken by Hummelinck from the attics of schools and houses, whereas Rojer (1997) reported this species as roosting among the ruins at "One Hoe." These are typical roost sites for this species elsewhere in the West Indies (Gannon et al., 2005; Genoways et al., 2005; Pedersen et al, 2005, 2006). Foraging M. molossus were taken in nets along with T. brasiliensis over the abandoned swimming pool at Mary's Fancy and the open grassy area with isolated single large trees at lower Loteríe Farm and along with A. jamaicensis in the ravine at Romeo's Drive in Middle Region. Elsewhere, individuals were netted over roads along the northeastern and southwestern slopes of Pic du Paradis, which we believe were used as protected flyways to move from one side of the island to the other.
Of the 10 females taken between 15 and 22 May during our work on the island, eight revealed no gross reproductive activity, whereas single females taken on 16 and 18 May carried embryos that measured, respectively, 7.5 and 6.0 in crown-rump length. Four males taken during this same time period had testes that averaged 3.4 (2-5) long. Three females, prepared as specimens on 13 and 14 July, revealed that one taken on each date was pregnant carrying a single embryo that measured, respectively, 22 and 9 in crown-rump length, whereas the third female evinced no gross reproductive activity. Four males taken during this same time period had testes that averaged 4.5 (4-5) long. Eleven of 12 mastiff bats captured and released on the night of 15 May 2002 were females. Ten of the females were judged to be pregnant with the eleventh female being small, weighing only 9.7 and being judged as non-reproductive. Because Genoways et al. (2005) on Jamaica found females of this species at various stages of reproduction and others reproductively inactive during the same period, they concluded that the reproductive cycle most closely resembled an aseasonal polyestrous pattern (Wilson, 1979). Our data from St. Martin are not in conflict with this conclusion, but far more data are needed to confirm this conclusion.
The eight non-pregnant females taken during May weighed on average 12.1 (9.9-16.0), whereas the non-pregnant female taken on July 14 weighed 10.6. In comparison, the 10 pregnant females that were captured and released in May weighed on average 13.7 (12.6-14.4). The four males taken in May weighed on average 12.2 (11.2-13.0) and the four males taken in July weighed on average 13.1 (11.9-13.7).
SPECIES REMOVED FROM FAUNA
Myotis nesopolus Miller, 1900
Specimen examined (1).-SINT MAARTEN: Cul de Sac, 1 (RMNH).
Husson (1960) first reported this specimen from Sint Maarten under the scientific name Myotis nigricans nesopolus. Since this publication, the taxonomy of the Myotis nigricans group has undergone considerable change, particularly by LaVal (1973). LaVal (1973) assigned West Indian bats of this genus to two species- Myotis dominicensis from the island of Dominica and Myotis martiniquensis from the islands of Martinique and Barbados. LaVal (1973) did not comment on this specimen from St. Maarten or two specimens from Grenada (Koopman, 1968). Masson and Breuil (1992) subsequently have reported a Myotis from Guadeloupe that presumably would be assigned to M. dominicensis, at least on geographic grounds. Koopman (1968) initially assigned the specimens from Grenada to M. n. nigricans and Genoways et al. (1998) added a third specimen for the island and supported Koopman's taxonomic assignment. Therefore, as is currently understood, three species of small Myotis occur in the Lesser Antilles and the species most closely approaching Sint Maarten is M. dominicensis known from Guadeloupe about 240 kilometers to the south.
The measurements of the specimen from St. Maarten (see below) do more closely resemble those of M. dominicensis and M. nigricans than they do the measurements of the larger-sized M. martiniquensis (LaVal, 1973: 35). However, length of forearm of the specimen from St. Maarten at 31.1 is considerably shorter than averages of the former two species at 34.0 and 34.8, respectively. The single characteristic that LaVal (1973) found that distinguished M. dominicensis was a low ratio of the length of the tibia to the forearm. LaVal (1973) stated "The ratio of 0.35 [for M. dominicensis ] is exceeded by all other series of Neotropical Myotis I have examined. A large majority of the series of all species fall within the range of 0.38 to 0.42, except Myotis larensis, with a ratio of 0.48." With a tibia length of 16.0, the ratio of the length of the tibia to the forearm for the specimen from St. Maarten is 0.51. This ratio would certainly argue against assigning the specimen from St. Maarten to M. dominicensis and would seem to align it with M. larensis, which is a species described by LaVal (1973) from the dry coastal areas of northern Venezuela characterized by small size, but very long third metacarpal, tibia, and greatest length of skull in comparison with the forearm. Genoways and Williams (1979) demonstrated that M. larensis was best considered to be a subspecies of M. nesopolus, a species originally described from Curaçao and not treated by LaVal in his study. Table 2 gives values for these three ratios for seven taxa of Myotis from Caribbean islands and Caribbean coast of South America. The values of the specimen from St. Maarten clearly fall with those of the taxa of Myotis nesopolus from coastal Venezuela, Curaçao, and Bonaire.
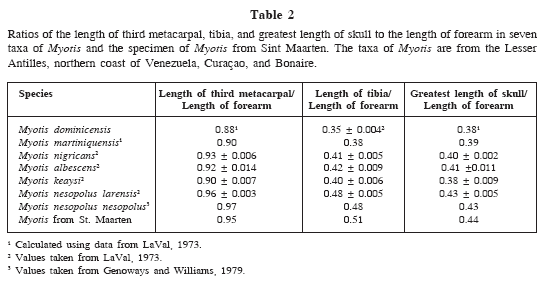
Morphologically, the specimen from St. Maarten most closely resembles Myotis nesopolus from two islands just off the northern coast of Venezuela and the adjacent mainland. This brings the discussion back to Husson (1960) who stated that "It can't be said with certainty whether the specimen from St. Martin really belongs to M. nesopolus; however, the specimen resembles M. nesopolus more than it does M. nigricans dominicensis." We are in full agreement with this statement. If we accept that the specimen from St. Maarten is a representative of M. nesopolus, we have a species occurring on the northeastern edge of the Caribbean Sea and two islands just off the north coast of South America more than 900 kilometers to the southwest. This is not a pattern of distribution known for any other species of Antillean bat so an explanation of this distribution pattern needs to be considered.
We only know from Husson (1960) that the specimen was found in an old building in Cul de Sac on the Dutch side of the island. It was obtained by F. A. da Costa on 20 September 1946. In searching for an explanation for this distribution pattern, it is difficult to ignore the fact that the Curaçao, Bonaire, and Sint Maarten are, and have been, members of the Netherlands Antilles. Also the fact that the specimen was collected just following World War II when traffic among the islands was high needs to be considered. Our tentative conclusion based on the facts and circumstantial evidence is that the specimen from Sint Maarten represents an individual that was accidentally transported from Curaçao or Bonaire to the island. Our work on the island provided no evidence of a resident population of Myotis, although such a negative hypothesis can't be proven. We believe at the present time it is best to remove Myotis from the chiropteran fauna of Sint Maarten pending any evidence to the contrary.
External and cranial measurements of the male from St. Maarten (RMNH 17107) are as follows: length of forearm, 31.1; length of third metacarpal, 29.4; length of tibia, 16.0; greatest length of skull, 13.6; condylobasal length, 12.7; zygomatic breadth, 7.6; postorbital constriction, 3.2; mastoid breadth, 6.7; length of maxillary toothrow, 5.1; breadth across upper molars, 4.9.
DISCUSSION
Faunal composition. -Data presented herein provide records of three species of bats new to the fauna of the island of Saint Martin/Sint Maarten- Monophyllus plethodon, Ardops nichollsi, and Natalus stramineus. With the addition of these three species and the omission of Myotis nigricans nesopolus, the chiropteran fauna of the island is composed of eight species. Ecologically, this simple chiropteran fauna includes representatives of four families-Noctilionidae, Phyllostomidae, Natalidae, and Molossidae-including one piscivore (N. leporinus), one omnivore (B. cavernarum), one pollenivore/nectarivore (M. plethodon), two frugivores (A. nichollsi, A. jamaicensis), and three insectivorous species (N. stramineus, T. brasiliensis, and M. molossus).
As our knowledge of the bats occurring on the Lesser Antillean islands north of Guadeloupe increases, an interesting biogeographical pattern may be emerging. The chiropteran fauna of Nevis (Pedersen et al., 2003) exactly matches that of Saint Martin/Sint Maarten. The faunas of St. Kitts (Pedersen et al., 2005) and Antigua (Pedersen et al., 2006) each are known to have seven species of bats. The only species missing from St. Kitts that is known from Saint Martin/Sint Maarten is Natalus stramineus, and Ardops nichollsi is not present on Antigua. The former is not a surprise given the difficulty of capturing this funnel-eared bat, but the latter is more puzzling given the abundance of Antillean tree bats on several adjacent islands.
On Montserrat the fauna contains an additional two species- Sturnira thomasi and Chiroderma improvisum -that do not occur on any island north of Guadeloupe except for the small populations on Montserrat. These data seem to indicate that the bat fauna of Montserrat with its 10 species has a stronger biogeographic relationship with Guadeloupe than with other islands in the northern Lesser Antilles. The other islands in the northern Lesser Antilles would appear to share a fauna that contains at most eight species of bats such as those present on Nevis and Saint Martin/Sint Maarten. We need data from additional islands to confirm this observation, but many of these are either small-Saba, Sint Eustatius, and Saint Barthélemy-or are low lying-Anguilla and Barbuda. It will be interesting to learn if these islands share this same fauna of eight species.
Conservation.-Although the bat fauna of Saint Martin/Sint Maarten is not unique, we must express considerable concern about the future of each species. If all feeding guilds are considered, bat capture rates on Sint Maarten/Saint Martin (1.63 bats per net per night - BNN) fall well below capture rates reported from mainland populations (2.59-6.65; Fenton et al., 1992; Findley, 1993) and are among the lowest noted on islands throughout the Lesser Antilles that the authors have surveyed (1.55-3.75; Pedersen et al., 2005). If only fruit bats are evaluated, capture rates during our three surveys on Sint Maarten/Saint Martin (0.92 BNN) are again on the low end of those rates reported from adjacent islands in the northern Lesser Antilles (0.65-2.47), and fall well below fruit bat captures per net-night in the Neotropics of Central America (2.20-5.93; Pedersen et al. 2005). We use this BNN metric conservatively and avoid its use as an estimate of population size per se, rather BNN is best used as an approximation of bat activity at a particular location. Indeed, Fenton et al. (1992) showed that phyllostomid bats are useful indicators of habitat disruption; they documented a 21% decrease (4.20 > 3.29 BNN) in fruit bat capture rates in their comparison of undisturbed with disturbed habitats in Mexico. Island populations of fruit bats appear more susceptible to habitat disruption than are mainland populations. Indeed, if contrasts between pre- and post-hurricane surveys are made using comparable data from Montserrat, captures rates decrease by as much as 60 to 80% after a natural disaster (Pedersen et al., 1996; Pedersen, 2001). Our observations of the remaining forest of Saint Martin/Sint Maarten and the conditions for bats in particular were not very encouraging and suggest an anthropogenic ecological disaster.
In several areas on the island, we encountered accumulating piles of garbage and human waste. The long-term concern is that these areas will lose all native vegetation and will be of no more use to the wildlife and more importantly to humans, including tourists. The watertable of the island has been reduced to the point that we observed only two small flowing streams and the one through Dutch Cul de Sac was so polluted with oil and phosphates that bats could not use it as a water source. We noted very few freshwater ponds on the island and most reservoirs were covered, leaving resort and residential swimming pools as the only source of freshwater available to bats and other wildlife.
In conjunction with the desire for expansion of the tourism industry, there is a major building boom on the island. Residential building of upscale homes, villas, and condominiums is progressing at a rapid pace, especially on the Dutch half of the island. On the French side the construction of access roads and private drives on the steep slopes of Pic du Paradis (425 m) have created great scars on the landscape and severely altered the ability of the soil to store water. Private residences are found at 410 m on the northeastern slope and up to 350 m on the southwestern slope. The subsequent loss of native vegetation impacts not only the food resources available to bats, but also deprives some species (Ardops nichollsi and Artibeus jamaicensis) of roost sites.
The removal of native vegetation has probably had an impact on the insect fauna of the island, which was suggested by Steadman et al. (1984) as a possible cause of the loss of two species of insectivorous bats on Antigua. The most striking example of potential reduction of an insectivorous species was the low numbers of the normally over-abundant Molossus molossus of which we took only 49 specimens in our three years of work giving a capture rate of 0.57 bats per net night (M. molossus -Montserrat: 0.98; St. Kitts: 1.14; St. Eustatius: 1.97; Nevis: 2.41 BNN). The loss of a diversity of food sources in the form of insects seems the most likely reason for the reduction of this species on Saint Martin/Sint Maarten because their preferred roost sites associated with tile and corrugated steel roofs appear to be abundant on the island.
Certainly, the heaviest blow to the chiropteran fauna of the island was the filling of the Devil's Hole cave and its two associated tunnels. This was the largest known bat cave on the island and had reproductive populations of at least Tadarida brasiliensis and Artibeus jamaicensis and probably also could have served Brachyphylla cavernarum. Half of the species of bats on the island- M. plethodon, B. cavernarum, N. stramineus, and T. brasiliensis -are obligate cave roosting species, meaning that they require caves for day roosts and nursery colonies of newborn bats. Noctilio leporinus and Artibeus jamaicensis are facultative cave species, meaning that they will use caves when available for reproduction, day roosts, and to escape harsh environmental condition, such as hurricanes. The few scattered remains of the funnel-eared bat, Natalus stramineus, were found in Billy Folly Cave just up slope from the former location of Devil's Hole. We question the current presence of this species on island because the Devil's Hole would have been a far more appropriate environment for the species than is provided by Billy Folly Cave. We are certain that there are other small caves on this island, but they are not the large cave complexes that we have found on adjacent islands, such as Anguilla or Barbuda.
The most important thing that can be done to conserve chiropteran populations on Saint Martin/Sint Maarten is to protect all of the remaining cave resources on the island. The caves and their immediate environs should be preserved as natural areas so that the impact of human activity and development can be limited in the future. Caves are very highly specialized ecosystems that possess a significant amount of biological diversity. The caves are certainly very important to the continued survival of many species of bats on Saint Martin/Sint Maarten, but on the other hand, bats are extremely important to these ecosystems because they perform the same role for caves as green plants do for non-cave situations. It is the dropped food items and guano of bats that are primary sources of energy entering these ecosystems. To maintain a healthy set of cave ecosystems with their rich biodiversity, it will be extremely important to maintain healthy bat populations on Saint Martin/Sint Maarten (Genoways et al., 2005).
We don't wish to paint only a dark picture of the future of bat conservation on Saint Martin/Sint Maarten. There are many dedicated people working in a variety of conservation projects across the island (primarily concerning marine ecosystems) and our work would have been impossible without their full cooperation. They labor under severely limited funding with limited personnel. The Dutch have an excellent terrestrial conservation plan that was developed in the mid-1990s (Rojer, 1997), but during our survey work, it was not apparent that the plan had been implemented. This plan deserves official support and implementation to help preserve the overall biodiversity of the island. A real conservation bright spot was Loteríe Farm in French Saint Martin. The owners of this private property are making significant progress in preserving and restoring the native vegetation of the property. This is an exciting development because we believe the most important areas for preservation on the island are headwaters of the Loterie and Columbier valleys along the western slopes of Pic du Paradis. These valleys retain the most extensive areas of native and advanced secondary vegetation remaining on the island. They should have the highest priority for preservation. Indeed, only at Loteríe Farm did we encounter the Lesser Antillean endemic species Ardops nichollsi.
ACKNOWLEDGMENTS
We wish to thank the following for financial and material support throughout the project: Totten Trust, Hagan Fund, Andy Caballero, Paul Ellinger, and Elsje Bosch (Nature Foundation Sint Maarten), Anne Marie Scot (St. Maarten/Martin Hotel & Tourist Association), and Robert and Dottie Chertok with Dale and Alice Vanwormer (American University of the Caribbean Medical School). We also wish to thank the following for their advice and assistance in the field: Hans van Buel, Eric Dubois-Millot (Nature Foundation Sint Maarten), Adam Brown (EPIC), and Anya Hartpence and Betsy South (South Dakota Sate University). We would be remiss not to give special thanks to Andy Caballero who aided our work far beyond his official duties with logistical support, arranging accommodations, and general guidance for our work on the island.
We thank local authorities for their assistance in arranging collecting permits and access to public lands: Andy Caballero, Paul Ellinger, and Elsje Bosch (Nature Foundation Sint Maarten), Ronan Kirsch and Jean-Marc Pons (Museum National d'Histoire Naturelle, Paris), and Nadia Roseau (Bureau de l'Environnement, Prefecture de la Region Guadeloupe, Basse-Terre, Guadeloupe).
The patience and courtesy extended to us by the estate owners are most gratefully acknowledged, especially William J. (BJ) Welch (Loteríe Farm-Rain Forest) and Elizabeth Reitz (Mary's Fancy). Curatorial support was provided by the Division of Zoology, University of Nebraska State Museum, with special thanks to Thomas Labedz and Patricia Freeman, and the Museum of Texas Tech University, with special thanks to Heath Garner and Robert J. Baker. We also wish to thank the following curators for allowing us access to specimens housed in their research collection: Chris Smeenk, Nationaal Natuurhistorisch Museum, Leiden (RMNH); Nancy B. Simmons, American Museum of Natural History (AMNH); Jean-Marc Pons, Museum National d'Histoire Naturelle (MNHN); Robert J. Baker, Texas Tech University (TTU); Patricia Freeman, University of Nebraska State Museum (UNSM).
LITERATURE CITED
ANDERSEN K. 1908. A monograph of the chiropteran genera Uroderma, Enchisthenes, and Artibeus. Proceedings of the Zoological Society of London 1908: 204-319. [ Links ]
BAKER RJ, HH GENOWAYS, and JC PATTON. 1978. Bats of Guadeloupe. Occasional Papers of the Museum, Texas Tech University 50:1-16. [ Links ]
CARIBBEAN HURRICANE NETWORK. 2005. St. Maarten/St. Martin (TNCM). Accessed November 21 at <http://stormcarib.com/climatology/> [ Links ].
COPE ED. 1889. On the Mammalia obtained by the Naturalist Exploring Expedition to southern Brazil. American Naturalist 23:128-150. [ Links ]
DAVALOS LM. 2005. Molecular phylogeny of funnel-eared bats (Chiroptera: Natalidae), with notes on biogeography and conservation. Molecular Phylogenetics and Evolution 37:91-103. [ Links ]
DAVIS WB. 1973. Geographic variation in the fishing bat, Noctilio leporinus. Journal of Mammalogy 54:862-874. [ Links ]
DOLAN PG. 1989. Systematics of Middle American mastiff bats of the genus Molossus. Special Publications of the Museum, Texas Tech University 29:1-71. [ Links ]
FENTON MB, L ACHARYA, D AUDET, MB HICKEY, C MERRIMAN, MK OBRIST, and D SYME. 1992. Phyllostomid bats (Chiroptera: Phyllostomidae) as indicators of habitat disruption in the Neotropics. Biotropica 24:440-446. [ Links ]
FINDLEY JS. 1993. Bats: a community perspective. Cambridge University Press, Cambridge, UK, xi + 167 pp. [ Links ]
FLEMING TH. 1971. Artibeus jamaicensis: delayed embryonic development in a Neotropical bat. Science 171:402-404. [ Links ]
GANNON MR, A KURTA, A RODRIGUEZ-DURAN, and MR WILLIG. 2005. Bats of Puerto Rico. Texas Tech University Press, vi + 239 pp. [ Links ]
GENOWAYS HH, RJ BAKER, JW BICKHAM, and CJ PHILLIPS. 2005. Bats of Jamaica. Special Publications of the Museum, Texas Tech University 48:1-155. [ Links ]
GENOWAYS HH, CJ PHILLIPS, and RJ BAKER. 1998. Bats of the Antillean island of Grenada: a new zoogeographic perspective. Occasional Papers of the Museum, Texas Tech University 177:1-28. [ Links ]
GENOWAYS HH, RM TIMM, RJ BAKER, CJ PHILLIPS, and DA SCHLITTER. 2001. Bats of the West Indian island of Dominica: natural history, areography, and trophic structure. Special Publications of the Museum, Texas Tech University 43:1-43. [ Links ]
GENOWAYS HH and SL WILLIAMS. 1979. Notes on bats (Mammalia: Chiroptera) from Bonaire and Curaçao, Dutch West Indies. Annals of Carnegie Museum 48:311-321. [ Links ]
GOODWIN GG. 1959. Bats of the subgenus Natalus. American Museum Novitates 1977:1-22. [ Links ]
HALL ER. 1946. Mammals of Nevada. University of California Press, Berkeley, xi + 710 pp. [ Links ]
HALL ER. 1981. The mammals of North America. John Wiley & Sons, New York, 1:xv + 1-600 + 90. [ Links ]
HANDLEY CO, J r and AL GARDNER. 1990. The holotype of Natalus stramineus Gray (Mammalia: Chiroptera: Natalidae). Proceedings of the Biological Society of Washington 103:966-972. [ Links ]
HUMMELINCK PW. 1979. Caves of the Netherlands Antilles. Publications of the Foundation for Scientific Research in Surinam and the Netherlands Antilles 97:1-176. [ Links ]
HUSSON AM. 1960. Mammals of the Netherlands Antilles. Natuurwetenschappelijke Wekgroep Nederlandse Antillen, Curaçao 12:viii + 1-170. [ Links ]
HUSSON AM. 1962. The bats of Suriname. Zoölogische Verhandelingen, Rijksmuseum van Natuurlijke Historie, Leiden, The Netherlands 58:1-282. [ Links ]
JONES JK, J r and RJ BAKER. 1979. Notes on a collection of bats from Montserrat, Lesser Antilles. Occasional Papers of the Museum, Texas Tech University 60:1-6. [ Links ]
JONES JK, J r and A SCHWARTZ. 1967. Bredin-Archbold-Smithsonian Biological Survey of Dominica. 6. Synopsis of bats of the Antillean genus Ardops. Proceedings of the United States National Museum 124(3634):1-13. [ Links ]
KOOPMAN KF. 1968. Taxonomic and distributional notes on Lesser Antillean bats. American Museum Novitates 2333:1-13. [ Links ]
LAVAL RK. 1973. A revision of the Neotropical bats of the genus Myotis. Science Bulletin, Natural History Museum of Los Angeles County 15:1-54. [ Links ]
MASSON D and M BREUIL. 1992. Un Myotis (Chiroptera, Vespertilionidae) en Guadeloupe (Petites Antilles). Mammalia 56:473-475. [ Links ]
OWEN RD, RK CHESSER, and DC CARTER.1990. The systematic status of Tadarida brasiliensis cynocephala and Antillean members of the Tadarida brasiliensis group, with comments on the name Rhizomops Legendre. Occasional Papers of the Museum, Texas Tech University 133:1-18. [ Links ]
PEDERSEN SC. 2001. The impact of volcanic eruptions on the bat populations of Montserrat, BWI. American Zoologist 40:1167A. [ Links ]
PEDERSEN SC, HH GENOWAYS, and PW FREEMAN. 1996. Notes on the bats from Montserrat (Lesser Antilles), with comments concerning the effects of Hurricane Hugo. Caribbean Journal of Science 32:206-213. [ Links ]
PEDERSEN SC, HH GENOWAYS, MN MORTON, JW JOHNSON, and SE COURTS. 2003. Bats of Nevis, northern Lesser Antilles. Acta Chiropterologica 5:251-267. [ Links ]
PEDERSEN SC, HH GENOWAYS, MN MORTON, GG KWIECINSKI, and SE COURTS. 2005. Bats of St. Kitts (St. Christopher), northern Lesser Antilles, with comments regarding capture rates of Neotropical bats. Caribbean Journal of Science 41:744-760. [ Links ]
PEDERSEN SC, HH GENOWAYS, MN MORTON, VJ SWIER, PA LARSEN, KC LINDSAY, RA ADAMS, and JD APPINO. 2006. Bats of Antigua, northern Lesser Antilles. Occasional Papers of the Museum, Texas Tech University 249:1-18. [ Links ]
REHN JAG. 1901. Notes on Chiroptera. Proceedings of the Academy of Natural Sciences of Philadelphia 52:755-759. [ Links ]
ROJER A. 1997. Biological inventory of St. Maarten. Carmabi Foundation, Postbus 2090, Curaçao, Nederlandse Antillean, 28 pp. Accessed at http://www.mina.vomil.an/Publ/RojerSXMsurvey.html. [ Links ]
SAGER S. 1992. StatView®. Abacus Concepts, Inc., Berkeley, CA, x + 466 p. [ Links ]
SCHWARTZ A. 1955. The status of the species of the brasiliensis group of the genus Tadarida. Journal of Mammalogy 36:106-109. [ Links ]
SCHWARTZ A and JK JONES, J r. 1967. Bredin-Archbold-Smithsonian Biological Survey of Dominica. 7. Review of bats of the endemic Antillean genus Monophyllus. Proceedings of the United States National Museum 124(3635):1-20. [ Links ]
SHAMEL HH. 1928. A new bat from Dominica. Proceedings of the Biological Society of Washington 41:67-68. [ Links ]
STEADMAN DW, GK PREGILL, and SL OLSON. 1984. Fossil vertebrates from Antigua, Lesser Antilles: evidence for late Holocene human-caused extinctions in the West Indies. Proceedings of the National Academy of Science USA 81:4448-4451. [ Links ]
STOFFERS AL. 1956. The vegetation of the Netherlands Antilles. Studies on the Flora of Curaçao and Other Caribbean Islands 1:1-142. [ Links ]
SWANEPOEL P and HH GENOWAYS. 1978. Revision of the Antillean bats of the genus Brachyphylla (Mammalia: Phyllostomatidae). Bulletin of the Carnegie Museum of Natural History 12:1-53. [ Links ]
TEJEDOR A. 2006. The type locality of Natalus stramineus (Chiroptera: Natalidae): implications for the taxonomy and biogeography of the genus Natalus. Acta Chiropterologica 8:361-380. [ Links ]
TIMM RM and HH GENOWAYS. 2003. West Indian mammals from the Albert Schwartz Collection: biological and historical information. Scientific Papers of the Natural History Museum, The University of Kansas, 29:1-47. [ Links ]
WILSON DE. 1979. Reproductive patterns. Pp. 317-378, in: Biology of bats of the New World Family Phyllostomatidae, Part III (RJ Baker, JK Jones, Jr., and DC Carter, eds.). Special Publications of the Museum, Texas Tech University 16:1-441. [ Links ]
Recibido 26 marzo 2006.
Aceptación final 24 abril 2007.

