










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. v.15 n.1 Mendoza ene./jun. 2008
Evaluación de los caracteres diagnósticos de Oxymycterus josei Hoffmann, Lessa y Smith, 2002 (Rodentia: Cricetidae) con comentarios sobre la diferenciación de las especies uruguayas del género
Felipe García-Olaso1
1 Sección Evolución y Sistemática, Facultad de Ciencias, Universidad de la República, Iguá 4225, Montevideo 11400, Uruguay. <fgarola@fcien.edu.uy>
RESUMEN: Actualmente se reconocen dos especies del género Oxymycterus en Uruguay: O. nasutus y O. josei. Estas especies parecen ser un caso particular dentro del género, ya que son simpátridas, genéticamente diferenciadas pero morfológicamente muy similares. Principalmente, dos caracteres diagnósticos fueron descritos para distinguirlas. Este trabajo evalúa la consistencia de esos dos caracteres craneodentarios para diferenciar las especies. Los resultados descartan los pliegues dentarios, pero corroboran al ancho de placa zigomática, como caracteres diagnósticos de O. josei. El actual conocimiento genético, morfológico y geográfico del género en la región sugiere que hay interrogantes históricas que permanecen por ser resueltas.
ABSTRACT: Evaluation of the diagnostic characters of Oxymycterus josei Hoffmann, Lessa y Smith, 2002 (Rodentia: Cricetidae) with comments on the differentiation of the Uruguayan species of the genus. Currently, there are two recognized species of the genus Oxymycterus in Uruguay: O. nasutus and O. josei. These species seem to represent a special case within the genus, as they are sympatric, genetically differentiated but morphologically very similar. Mainly, two diagnostic characters were described to distinguish them. This work evaluates the consistency of those two craniodental characters in differentiating the species. The results discard the dental folds, but support the zygomatic plate width, as diagnostic characters of O. josei. The current genetic, morphologic and geographic knowledge of the genus in the region suggest that historical questions remain to be solved.
Palabras clave. Morfología; Oxymycterus nasutus; Sigmodontinae. Sistemática; Uruguay.
Key words. Morphology; Oxymycterus nasutus; Sigmodontinae; Systematics; Uruguay.
Actualmente se reconocen dos especies del género Oxymycterus en Uruguay: O. nasutus (Waterhouse, 1837) y O. josei Hoffmann, Lessa y Smith, 2002. Para las dos se han propuesto distribuciones geográficas en el sureste del país, siendo en parte simpátridas (Fig.1). Al describir la segunda especie, Hoffmann et al. (2002) observaron que aunque genéticamente (ADN mitocondrial) estas especies constituyen linajes bien diferenciados, son morfológicamente muy difíciles de distinguir. Ciertamente, notaron que eran imposibles de diferenciar externamente y describieron únicamente unos pocos caracteres craneodentarios para distinguir a O. josei de O. nasutus. Según estos autores, además de presentar un cráneo algo más robusto, O. josei presentaría la placa zigomática y la raíz maxilar del arco zigomático más robustas, y pliegues característicos en los primeros molares superior (M1) e inferior (m1). Estas figuras en el patrón de esmalte serían un protostilo y un enterolofo con su correspondiente enterostilo en el M1 y diferencias análogas en el m1 (Hoffmann et al., 2002). Estas figuras de esmalte se desgastan rápidamente con la edad, por lo cual son detectables sólo en individuos jóvenes (Hoffmann et al., 2002).
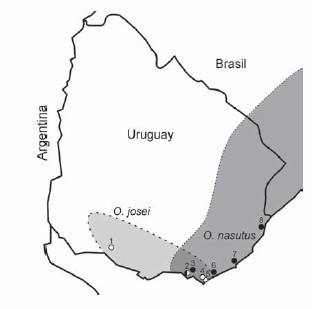
Fig. 1. Mapa de distribuciones propuestas previamente (a partir de localidades mencionadas en Hoffmann et al., 2002) para O. nasutus (gris oscuro) y O. josei (gris claro). Los puntos indican las localidades donde genéticamente se han determinado individuos de O. nasutus (relleno negro), O. josei (relleno blanco) y de ambas especies (relleno blanco y negro, Hoffmann, 1998; G. D'Elía, com. pers.). La numeración corresponde a la indicada en el texto y Apéndice 1 para estas localidades.
En este trabajo se evaluaron los caracteres craneodentarios diagnósticos propuestos por Hoffmann et al. (2002) con el objetivo de esclarecer los criterios de identificación morfológica, y de aportar a la taxonomía alfa de las especies uruguayas del género Oxymycterus.
Para evaluar los estados de los caracteres craneodentarios designados por Hoffmann et al. (2002) se analizó la distribución de las especies en base a 25 especímenes, 14 de O. josei y 11 de O. nasutus, provenientes de cinco localidades de Uruguay (Apéndice 1). Estos ejemplares fueron determinados como O. nasutus u O. josei a partir de su genotipo mitocondrial (sensu Hoffmann et al., 2002). Se asumió a estas determinaciones como correctas ya que ambos taxones muestran un claro y marcado diferenciamiento genético (Hoffmann et al., 2002). Para evaluar la presencia de los caracteres dentarios fue necesario clasificar y considerar el nivel de desgaste de los molares. Esto se realizó mediante una modificación de las categorías de edad definidas por Myers (1989) en base a la erupción y desgaste de los molares de especímenes de Akodon. En los individuos analizados, se observó que la erupción del tercer molar (M/m3) es más tardía en Oxymycterus que en Akodon, con relación a la erupción y desgaste del primer y segundo molar. Para esto se definió una nueva clase y se modificaron, únicamente en relación al estado del tercer molar (en lo referente al estado del primer y segundo molar estas clases son idénticas a las de Myers, 1989), las definiciones de algunas clases, como sigue:
Clase de edad 0: Igual a la clase 1 de Myers (1989) excepto que el tercer molar se encuentra dentro del maxilar.
Clase de edad 1: Tercer molar visible, habiendo llegado o llegando a la altura de la superficie oclusal de los demás molares, sin cortar las cápsulas de esmalte de las cúspides.
Clase de edad 2: M/m3 con las capsulas de esmalte cortadas y levemente desgastadas.
Clase de edad 3: M/m3 con la superficie oclusal plana o cóncava con los pliegues menores todavía visibles.
Clase de edad 4 y 5: Igual que en Myers (1989).
Se utilizó la nomenclatura propuesta por Reig (1977) para designar las figuras de esmalte en la superficie de oclusión de los molares. Por último, se consideró que las "diferencias análogas" a las del M1, existentes en el m1, mencionadas por Hoffmann et al. (2002), corresponden a la presencia en O. josei de un anterolófido con su correspondiente metastílido y de un mesolófido con su mesostílido (Hoffmann, com. pers.).
Para evaluar cuantitativamente el ancho de la placa zigomática (ZPW) se tomaron medidas de esta y del largo total del cráneo (GSL) siguiendo a Hoffmann et al. (2002) con calibre digital, promediando tres medidas del mismo ejemplar para cada variable. Para este análisis se utilizaron cráneos de 20 individuos que permitían registrar con certidumbre ambas variables debido a su estado de conservación (Apéndice 1). Los datos del ancho de la placa fueron corregidos para el efecto del tamaño relativo del cráneo, utilizando la variable GSL como variable relacionada al tamaño y con la corrección de Lleonart et al. (2000). La ecuación resultante para la corrección fue: ZPWi* = ZPWi [GSL0/GSLi]b, utilizada para cada individuo i, donde ZPW* corresponde a la corrección de la variable ZPW, GSL0 es la media de la variable GSL y b es el factor de potencia que relaciona a dos variables morfométricas cualesquiera (Lleonart et al., 2000). Este factor fue estimado mediante la regresión lineal de las variables, previa transformación logarítmica. Luego, sobre los datos corregidos se evaluó la existencia de diferencias significativas entre las especies mediante una prueba de U de Mann y Whitney (1947) y Wilcoxon (1945).
En primer lugar, se notó que 12 individuos de los 25 analizados, cinco correspondientes a O. nasutus y siete a O. josei, no presentaban ninguno de los pliegues molares en cuestión, probablemente debido al desgaste avanzado de los molares (Tabla 1). En cuanto a los pliegues superiores, cinco individuos pertenecientes a O. josei, que poseían un grado de desgaste bajo, presentaron protostilo y enterolofo (Fig. 2 a, c). Fue algo novedoso encontrar que un individuo de O. nasutus poseía protostilo y enterolofo (Fig. 2). Con relación a los pliegues inferiores también fue novedoso encontrar que cinco ejemplares de O. nasutus poseían anterolófido y mesolófido (Fig. 2 d), a pesar de que entre los analizados había pocos individuos tan jóvenes como los ejemplares de O. josei que exhibían estos pliegues (Tabla 1). Resumiendo, se encontraron ejemplares de ambas especies que presentaban ambos grupos de pliegues de esmalte dentario.

Fig. 2. Serie molar superior izquierda de A) O. josei y B) O. nasutus donde se aprecia la presencia de un enterolofo (flecha); y fotografías de la serie molar inferior derecha de C) O. josei y D) O. nasutus donde se observa la presencia de un anterolófido y mesolófido (flechas) en un ejemplar de esta especie. A y C tomadas, y B modificada de Hoffmann et al. 2002 (Copyright © 2002 American Society of Mammalogists. De Journal of Mammalogy por Hoffman et al. Reimpreso con permiso de Alliance Communications Group).
Tabla 1
Ejemplares de Oxymycterus identificados genéticamente de acuerdo a Hoffmann et al. (2002), ordenados por clase de edad definida por el nivel de desgaste de sus molares, para los cuales se indica presencia o ausencia de protostilo y enterolofo (pliegues de esmalte en molares superiores), anterolófido y mesolófido (pliegues de esmalte en molares inferiores). Los superíndices indican: #, determinación realizada por la fotografía publicada en Hoffmann et al. (2002) ya que la mandíbula no se encuentra depositada junto con el cráneo; +, fotografía de serie molar superior e inferior en Hoffmann et al. (2002); *, fotografía de serie molar superior en Hoffmann et al. (2002); ‡, identificación realizada mediante técnicas genéticas por Hoffmann et al. (2002).

Respecto al estudio de la placa zigomática, en general, los ejemplares de O. josei presentaron una placa zigomática más ancha para el mismo largo del cráneo, siendo la relación entre las dos variables similar para las dos especies (Fig. 3). Las medias de la variable ZPW para cada especie, corregida para el tamaño del cráneo, resultaron ser significativamente diferentes entre las especies (U= 0, p<0.001).

Fig. 3. Relación del ancho de placa zigomática (ZPW) de los individuos estudiados, con el largo total del cráneo (GSL), en la especies uruguayas de Oxymycterus. Se puede observar que O. josei presenta una placa zigomática relativamente mayor que O. nasutus.
Los resultados descartan los pliegues dentarios inferiores y superiores como caracteres diagnósticos de O. josei, ya que estos se encuentran, indistintamente, en individuos de O. nasutus. Sólo un individuo de la especie O. nasutus presenta en forma reducida protostilo y enterolofo; sin embargo, esto podría deberse a que no fue posible analizar individuos de O. nasutus de la clase de edad más joven. Por lo tanto, es deseable analizar más individuos de las clases de edad 0 y 1 de esta especie.
Por otra parte, se corrobora al ancho de la placa zigomática como un carácter que es capaz de diferenciar a O. josei de O. nasutus. Esto es de gran importancia ya que lo convierte, hasta el momento, en el único carácter aplicable para la diferenciación morfológica de ambas especies. Sin embargo, debe notarse que para determinar individuos como pertenecientes a uno u otro taxón no es posible utilizar directamente el ancho de la placa zigomática debido al solapamiento en los rangos de variación de ambas especies (Fig. 3). Para determinar ejemplares se hace necesario corregir por el efecto del tamaño relativo mediante algún índice como el empleado en este estudio.
De esta forma las especies uruguayas del género Oxymycterus presentan aún menos caracteres distintivos que lo expresado por Hoffmann et al. (2002) en la diagnosis de O. josei. Esto deja a la recientemente descrita O. josei con escasos caracteres diagnósticos morfológicos, que son exclusivamente internos y de evaluación poco práctica. Este escenario, que es frecuente entre pares de especies de sigmodontinos (ver D'Elía y Pardiñas, 2004), dificulta la identificación morfológica de las especies, necesaria en la implementación de una variedad de estudios.
La invalidez de algunos caracteres morfológicos diagnósticos no sólo hace necesario intentar determinar nuevos criterios para la identificación morfológica de las dos especies de Oxymycterus, sino también revisar las distribuciones geográficas de ambas, propuestas por Hoffmann et al. (2002). Esto se debe a que varias de las localidades incluidas en las distribuciones fueron basadas en ejemplares identificados morfológicamente. Basado en nuestros resultados se pueden reportar especímenes con asignación genética y geográfica certera de la forma O. josei provenientes de Ecilda Paullier (1), Dpto. de San José y de Las Flores (2), Establecimiento El Peñasco (4) y barra del Arroyo Maldonado (5, localidad típica de O. nasutus) en el Dpto. de Maldonado; y de la forma O. nasutus de Las Flores (2), Pan de Azúcar (3) y José Ignacio (6), en el Dpto. de Maldonado y del cruce del Arroyo La Paloma con Ruta 15 (7, ver Apéndice 1 por detalle en nomenclatura del arroyo) y del parque Santa Teresa (8), Dpto. de Rocha (modificado de Hoffmann, 1998 con información de los catálogos de campo de Carlos Altuna y de la Sección Evolución, Facultad de Ciencias, Universidad de la República, Uruguay y comunicación personal de G. D'Elía; ver la Fig. 1 y el Apéndice 1, a dónde corresponde la numeración entre paréntesis). Sería interesante poder corroborar la clasificación a partir de la placa zigomática de los ejemplares hoy en día depositados en museos para poder conocer mejor estas distribuciones. Por otra parte, es importante notar que se han encontrado individuos en la localidad típica de O. nasutus que han sido genéticamente caracterizados como O. josei. Desde que el holotipo de nasutus solo fue analizado morfológicamente (incluyendo los caracteres dentarios ahora invalidados) por Hoffman et al. (2002) y su haplotipo mitocondrial se desconoce, la posibilidad de que, independientemente de si en Uruguay habitan una o dos especies de Oxymycterus, O. josei sea un sinónimo de O. nasutus no puede descartarse.
Según el escenario que aquí se plantea, las especies de Oxymycterus de Uruguay son simpátridas en gran parte de su distribución (incluidas ambas localidades típicas) y, a la vez, prácticamente indistinguibles morfológicamente. Este escenario parece ser un caso particular dentro del género y es interesante para estudiar la ecología, evolución y taxonomía de estas especies. La coexistencia, que se da incluso a escala local (e.g., los ejemplares MNHN 3854 y 3855 en el margen W del Arroyo Tarariras en Las Flores, Dpto. de Maldonado), de dos especies muy similares plantea interrogantes sobre cómo se da la repartición de recursos entre ambas y cómo es la diferenciación de sus nichos ecológicos para minimizar la competencia (MacArthur y Levins, 1967).
En contraposición con lo que pasa con la morfología, O. josei y O. nasutus se diferencian bien genéticamente y son filogenéticamente distantes, según el citocromo b mitocondrial. Si ambos taxones representaran una misma especie biológica, en la que existen dos linajes mitocondriales bien diferenciados, se estaría entonces frente a una discrepancia entre el árbol de genes mitocondrial y el de especies (Tajima, 1983; Pamilo y Nei, 1988). Esta discordancia podría resultar de eventos de separación y reencuentro de poblaciones grandes con fijación alternativa de variantes ancestrales, o de hibridación con otra especie seguida de una gran introgresión. Este último proceso podría haber involucrado a O. rufus, una especie con distribución adyacente a O. josei y que en análisis filogenéticos basados en ADN mitocondrial es recobrada como la especie hermana de ésta. Los haplotipos recobrados en estas especies presentan, entre ellos, valores muy bajos de divergencia observada (1.9%, Hoffmann et al., 2002). Estos valores son comúnmente observados a nivel intraespecífico en varios Sigmodontinae (Smith y Patton, 1999; D'Elía y Pardiñas, 2004).
Para dilucidar si estamos ante dos buenas especies biológicas, que dada su sinmorfía y simpatría constituirían un caso interesante para estudios ecológicos, o ante una sola especie biológica y un buen ejemplo de discordancia entre árbol de especies y árbol de genes, se hace necesario estudiar filogenéticamente nuevos caracteres no ligados al genoma mitocondrial (e.g.: ADN nuclear).
En cualquier caso, toda la información presentada en este trabajo tiene implicancias para el esclarecimiento de la taxonomía alfa del género, así como para entender su biología evolutiva, y deja en claro que los límites de las especies del género Oxymycterus siguen siendo confusos.
Los especímenes examinados se encuentran depositados en la colección de mamíferos del Museo Nacional de Historia Natural y Antropología de Montevideo, Uruguay (MNHN). Los especímenes listados con número de campo CA, correspondiente al catálogo de Carlos Altuna, todavía no poseen número definitivo de dicho museo. Los números entre paréntesis hacen referencia a las localidades ubicadas en la Figura 1 y mencionadas en el texto. Los superíndices indican lo siguiente: ‡, especímenes utilizados para el análisis morfométrico de la placa zigomática; *, especímenes cuya localidad fue erróneamente notada en Hoffmann et al. (2002) y que aquí se enmienda de acuerdo al catálogo de campo original; y §, un espécimen cuyo cráneo se encuentra incorrectamente rotulado CA 451, esto fue verificado con los datos del catálogo original. Una localidad se marca con † ya que existe un problema en la nomenclatura del arroyo en cuestión, que es indicado como Arroyo de la Paloma en la cartografía oficial y como Arroyo de la Palma en un letrero ubicado in situ, variante también aceptada internacionalmente y utilizada en el catálogo original. Este arroyo no debe ser confundido con el que cruza la ruta 15 en el km 18 y en cuyo letrero in situ se lee: Arroyo La Paloma. Se brindan coordenadas obtenidas a partir de la localización en imágenes satelitales de los sitios de colecta (E. P. Lessa com. pers.).
Oxymycterus josei.-URUGUAY: Maldonado: (2) margen Oeste del Arroyo Tarariras, Las Flores (34º49'00"S, 55º19'10"O): MNHN 3838‡ Holotipo, MNHN 3846, MNHN 3847, MNHN 3848‡, MNHN 3849‡, MNHN 3850‡, MNHN 3854‡, MNHN 3857‡, MNHN 4280‡, MNHN 4282‡, MNHN 4283‡; (4) Est. El Peñasco (34º50'48"S, 54º57'07"O): CA 479‡*, CA 480‡*,CA 481‡*
Oxymycterus nasutus.-URUGUAY: Maldonado: (2) margen Oeste del Arroyo Tarariras, Las Flores (34º49'00"S, 55º19'10"O): MNHN 3855‡; (3) Arroyo El Renegado, 3 Km. al E de Pan de Azúcar (34º45'00"S, 55º11'50"O): MNHN 4281‡; (6) 5 Km. al NNO del Faro de José Ignacio (34º38'39"S, 54º39'31"O): MNHN 4279‡*, CA 459‡§; Rocha: (7) Ruta 15 Km. 10, Arroyo de la Paloma† (34º35'10"S, 54º10'44"O): CA 614‡, CA 615‡, CA 617‡, CA 618, CA 627, CA 629‡, CA 630.
AGRADECIMIENTOS
Agradezco a Enrique Lessa, Guillermo D'Elía, Federico Hoffmann y dos revisores anónimos por sus comentarios que mejoraron sensiblemente este trabajo. También agradezco a Alejandro D'Anatro por su consejo sobre el análisis morfométrico y a Lucía Franco por su ayuda con las figuras. Por otra parte, les estoy agradecido a Javier González, Enrique González, Álvaro Mones y Arturo Toscano por facilitarme la revisión de colecciones en el Museo Nacional de Historia Natural y Antropología de Montevideo.
LITERATURA CITADA
1. D'ELÍA G y UFJ PARDIÑAS. 2004. Systematics of Argentinean, Paraguayan, and Uruguayan swamp rats of the genus Scapteromys (Rodentia, Cricetidae, Sigmodontinae). Journal of Mammalogy 85:897-910. [ Links ]
2. HOFFMANN FG. 1998. Sistemática de los ratones hocicudos del género Oxymycterus (Rodentia: Sigmodontinae) del Uruguay. Tesis de Maestría, PEDECIBA, Facultad de Ciencias, Universidad de la República, Montevideo, Uruguay. [ Links ]
3. HOFFMANN FG, EP LESSA y MF SMITH. 2002. Systematics of Oxymycterus with description of a new species from Uruguay. Journal of Mammalogy 83:408-420. [ Links ]
4. LLEONART J, J SALAT y GJ TORRES. 2000. Removing allometric effects of body size in morphological analysis. Journal of Theoretical Biology 205:85-93 [ Links ]
5. MACARTHUR RH y R LEVINS. 1967. The limiting similarity, convergence and divergence of coexisting species. American Naturalist 101:377-385. [ Links ]
6. MANN HB y DR WHITNEY. 1947. On a test of whether one of 2 random variables is stochastically larger than the other. Annals of Mathematical Statistics 18:50-60. [ Links ]
7. MYERS P. 1989. A preliminary revision of the varius group of Akodon (A. dayi, dolores, molinae, neocenus, simulator, toba, and varius). Pp. 5-54, en: Advances in Neotropical mammalogy (K Redford y JF Eisenberg, eds.). Gainesville, Florida, Sandhill Crane Press. [ Links ]
8. PAMILO P y M NEI. 1988. Relationships between gene trees and species trees. Molecular Biology and Evolution 5:568-583 [ Links ]
9. REIG OA. 1977. A proposed unified nomenclature for the enameled components of the molar teeth of the Cricetidae (Rodentia). Journal of Zoology 181:227- 241. [ Links ]
10. SMITH MF y JL PATTON.1999. Phylogenetic relationships and the radiation of sigmodontine rodents in South America: Evidence from cytochrome b. Journal of Mammalian Evolution 6:89-128. [ Links ]
11. TAJIMA F. 1983. Evolutionary relationship of DNA sequences in finite populations. Genetics 105:437- 460. [ Links ]
12. WILCOXON F. 1945. Individual comparisons by ranking methods. Biometrics Bulletin 1:80-83. [ Links ]
Recibido 31 julio 2006.
Aceptación final 20 agosto 2007.

