










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. v.15 n.2 Mendoza jul./dic. 2008
Densidad de población y uso de hábitat de Cebus libidinosus (primates, cebidae) en un bosque yungueño de Bolivia
Ninostka Burgoa A.1* y Luis F. Pacheco2,3**
1 Carrera de Biología, Facultad de Ciencias Puras y Naturales, Universidad Mayor de San Andrés,
La Paz, Bolivia.
2 Estación Biológica Tunquini, Instituto de Ecología, Universidad Mayor de San Andrés, Casilla 10077, Correo Central, La Paz, Bolivia.
3 Centro de Estudios en Biología Teórica y Aplicada (BIOTA).
*Cota cota calle 35 A, La Paz, Bolivia <ninostka@gmail.com>.
**Dirección actual: Centro de Postgrado en Ecología y Conservación, Instituto de Ecología, Casilla 10077, Correo Central, La Paz, Bolivia.
RESUMEN: El k'usillo o mono silbador (Cebus libidinosus), es una especie considerada plaga de los cultivos en los Yungas de La Paz, donde ataca los cultivos de cítricos y maíz. Como contribución al manejo de este conflicto, entre mayo y octubre de 2002 y entre mayo y julio de 2003, se hizo una estimación de la abundancia relativa y densidad y el uso de hábitat del k'usillo en el bosque montano de los alrededores de la Estación Biológica de Tunquini (EBT), ubicada dentro del Parque Nacional y Área Natural de Manejo Integrado Cotapata (PN Cotapata). Utilizando el método de conteo en transectas lineales, en seis senderos establecidos, se obtuvo una tasa de encuentro de 1.48 grupos/10 km y 29.15 individuos/10 km de recorrido. La densidad de grupos estimada con el programa Distance 4.1 fue de 2.16 grupos/km2 y 29.1 indiv/km2. Esta densidad es válida sólo para los alrededores de la EBT y no recomendamos su extrapolación a todo el PN Cotapata. Considerando la proporción de bosque y cultivos, el k'usillo usa los cultivos de cítricos con mayor frecuencia de lo esperado, mientras que usa el bosque de acuerdo a su disponibilidad. Esto coincide con las quejas de los pobladores del Chairo en cuanto la preferencia de esta especie por los cultivos.
ABSTRACT: Population density and habitat use of Cebus libidinosus in a montane forest of Bolivia. The Brown Capuchin Monkey, locally known as k'usillo (Cebus libidinosus), is considered a plague for citrus and corn cultivars in the Yungas region of Bolivia. As a contribution to the management of this conflict, between May and October of 2002, and between May and July of 2003, we carried out an estimate of density, and habitat use of a population in the montane forest at the outskirts of the Estación Biológica de Tunquini (EBT), located inside the Parque Nacional de Cotapata and Área Natural de Manejo Integrado Cotapata. We counted monkeys along six trails, used as line transects, and obtained an encounter rate of 1.48 groups/10 km and 29.15 individous /10 km. The density of groups estimated using Distance 4.1 was 2.16 groups/km2 and 29.1 indiv/km2. This density is valid only for the EBT surroundings, and we do not recommend its use for the whole Parque Nacional de Cotapata. Cebus libidinosus used the cultivations (especially citrus) more than expected by its availability which coincides with the perception of the peasants.
Palabras clave: Cotapata; Densidad; Primates; Transectas lineales; Uso de hábitat.
Key words: Cotapata; Density; Habitat; Line transects; Primates.
INTRODUCCIÓN
Uno de los grupos de mamíferos más estudiados en Bolivia es el de los primates. La mayoría de los estudios fueron sobre aspectos de distribución y abundancia (García et al., 1987; García y Tarifa, 1988, 1991; Cameron et al., 1989; Sainz, 1997; Rocha, 1999; Tarifa, 2000; Palomeque, 2001; Gómez et al., 2005; Muñoz y Kopp, 2005), pero también se estudiaron dieta y dispersión de semillas (Pacheco y Simonetti, 2000; Aguilar, 2002), socioecología y comportamiento (Wallace, 1998; Méndez, 1999; Cáceres, 2001; Martínez, 2003). No obstante, hasta el momento sólo dos estudios se han desarrollado en bosques montanos, correspondientes al registro de una nueva especie para Bolivia (Wallace y Painter, 1999) y a un trabajo sobre la ecología trófica de Ateles chamek en el Parque Nacional y Área Natural de Manejo Integrado Cotapata (Apaza, 2002). Ningún trabajo en Bolivia ha reportado sobre el daño que algunos primates pueden causar a los cultivos.
Cebus libidinosus se encuentra distribuido al este de los Andes desde Colombia y sur de Venezuela hasta Paraguay y norte de Argentina (Castro, 1977; Rylands y Keuroghlian, 1988; Emmons, 1999; Varela, 2003). Ocupa una amplia variedad de hábitats, incluyendo bosques tropicales y subtropicales, selvas lluviosas y bosques montañosos, bosques húmedos perennes, bosques secos, bosques de galería, bosques secundarios, bosques de neblina y manglares, hasta al menos 2700 m de altitud (Forbes et al., 1984; Janson, 1986; Varela, 2003). Esta especie, localmente conocida como k'usillo, prefiere los estratos superior y medio del bosque a una altura de cinco a 20 metros (Janson, 1986; García y Tarifa, 1991; Anderson, 1993; Varela, 2003), aunque no es raro verlo en el suelo y atravesando sabanas o cultivos. En Bolivia ha sido registrado en los departamentos del Beni, La Paz, Cochabamba, Pando, Santa Cruz y Tarija (Anderson, 1993; Emmons, 1999; Roldán, 2001). En nuestro sitio de estudio, el Parque Nacional y Área Natural de Manejo Integrado Cotapata, el k'usillo ha sido registrado hasta los 3000 m (Ribera, 1995), pero es más común por debajo de los 2300 m de altitud. El ámbito de hogar calculado para esta especie oscila entre 90-158 ha y su recorrido diario oscila entre 370 y 2300 m (Varela, 2003). Esta especie es comúnmente cazada para alimentación en varias regiones de Bolivia, pero en nuestro sitio de estudio sólo se le caza cuando se le sorprende dañando los cultivos (Roldán, 2001).
El k'usillo es una de las especies identificadas como dañinas a los cultivos en el Parque Nacional y Área Natural de Manejo Integrado Cotapata (PN Cotapata), donde ataca principalmente cultivos de maíz y cítricos (Pérez y Pacheco, 2006). Según los campesinos, C. libidinosus es más frecuente en los cultivos que en el bosque. La estrategia principal de manejo del problema por parte de los campesinos es el ahuyentamiento de los monos, pero también existen personas que usan veneno y armas de fuego para controlar el daño causado por el k'usillo. Ante tal escenario, la conservación del k'usillo dentro del PN Cotapata sería dependiente, al menos parcialmente, del manejo de su conflicto con la actividad agrícola. Como un aporte en este tema, este trabajo se diseñó para responder dos preguntas: primero ¿cuál es la densidad de C. libidinosus en la zona que bordea los cultivos?, y segundo ¿usa C. libidinosus las zonas de cultivo con mayor frecuencia que las de bosque?
MATERIALES Y MÉTODOS
Área de estudio
El PN Cotapata fue creado el año 1993 y cubre una superficie aproximada de 60 000 ha. con alturas entre 1200 y 5600 m. Se encuentra en la Vertiente Oriental de la Cordillera de los Andes y está ubicado al NE de la ciudad de La Paz, entre las provincias Nor Yungas y Murillo, con coordenadas referenciales 68°02' y 16°20' (SO), 68°03' y 16°05'. (NO), 67°43' y 16°10' (NE) y 67°47' y 16°18' (SE). Pertenece a las subregiones biogeográficas de puna y bosque húmedo montañoso de Yungas (Ribera, 1995).
Este trabajo se realizó en el bosque húmedo montañoso de Yungas, en los alrededores de la Estación Biológica Tunquini (EBT), entre 1500 y 2400 m de altitud (Fig. 1). La zona presenta una época con menor precipitación entre abril y septiembre y una época de lluvias entre octubre y marzo; la precipitación anual en la zona es de aproximadamente 2700 mm por año (EBT, datos no publicados). Las temperaturas medias anuales oscilan entre 12º y 17º C (Ribera, 1995).

Fig. 1. Localización del Parque Nacional y Área Natural de Manejo Integrado Cotapata en el Departamento de la Paz, Bolivia (Extraído y modificado de www.sernap.gov.bo/areasprotegidas/cot_1.htm). EBT: Estación Biológica Tunquini.
El PN Cotapata ha tenido actividad humana desde épocas precolombinas, la cual actualmente es de intensidad moderada y se encuentra concentrada en la región del Valle Chucura-Huarinilla donde la topografía y la accesibilidad permiten cierto uso de la tierra. Una importante proporción del área permanece poco intervenida debido a la dificultad de acceso y la topografía abrupta que la caracteriza, especialmente en el cinturón de bosque nublado (Ribera, 1995). En las áreas circundantes a la EBT la vegetación es heterogénea, debido a la topografía escarpada y el uso agrícola. La vegetación original corresponde a un bosque húmedo montañoso de Yungas (Paniagua et al., 2003), el cual ocupa cerca al 15% de la superficie en el área del PN Cotapata y un 92% del área de estudio. El dosel es denso y continuo, que alcanza entre 15 y 20 m de altura, con emergentes de hasta 40 m. Las familias más representativas son Moraceae, Sapotaceae, Laureaceae, Euphorbiaceae, Meliaceae, Burseraceae, Sapindaceae, Piperaceae, Rubiaceae, Leguminoseae y Araliaceae (Paniagua et al., 2003).
Para el presente estudio, se consideraron dos tipos de hábitat en las áreas circundantes a la EBT: bosque (maduro y secundario) y cultivos (principalmente de cítricos, algunos ya en barbecho o cultivos abandonados). Los Yungas tienen una larga historia de prácticas agrícolas, siendo los principales productos comerciales de la zona la coca (relativamente rara en la zona de estudio), los cítricos y el café (Subcentral 2 de Julio et al., 2005). Los cultivos (naranja, mandarina, lima y café) se concentran cerca de los senderos que llevan a las viviendas y campos de cultivo (Paniagua et al., 2003). Los barbechos jóvenes presentan fisonomía de matorral denso que escasamente sobrepasa los tres m de altura. Con una baja cobertura en el estrato arbóreo, el bosque presenta una altura promedio entre cuatro y cinco m de alto, el dosel es abierto, con pocos emergentes como Hedyosmum racemosum (Chloranthaceae) y Saurauia peruviana (Actinidiaceae), que no superan los 10 m de alto. El estrato inferior está dominado por árboles con un diámetro menor a 10 cm como Baccharis oblanceolata (Compositae), Piper psilophyllum (Piperaceae) y Vernonia ferruginea (Compositae) (Beck et al., 1993; Paniagua et al., 2003).
Gran parte del área corresponde a zonas de barbechos más antiguos (bosque secundario entre 10 y 20 años), caracterizados por una vegetación arbórea con una altura mayor (10-15 m), un dosel más abierto y una notable pobreza de especies; destacando por su dominancia Cecropia angustifolia (Moraceae), Piper elongatum (Piperaceae), Ochroma pyramidale (Bombacaceae), Solanum riparium (Solanaceae), Inga adenophylla (Mimosoideae), Inga heterophylla (Mimosoideae), Croton angustifolium (Euphorbiaceae), Hyeronima alchorneoides (Euphorbiaceae), Mabea longifolia (Euphorbiaceae) y Acalypha benensis (Euphorbiaceae) (Beck et al., 1993; Paniagua et al., 2003). Algunas zonas presentan manchones de bosque aparentemente de mayor edad (más de 20 años), estos parches de bosque se diferencian por una mayor altura de dosel, mayor número de estratos y una mayor diversidad de especies, encontrándose individuos de Cecropia pinnatiloba (Moraceae), Cecropia polystachya (Moraceae), Didymopanax morototoni (Araliaceae), Ceiba salmonea (Bombacaceae), Inga adenophylla (Mimosoideae), Inga vellutina (Mimosoideae), Acalypha mapiriensis (Euphorbiaceae), Croton angustifolium (Euphorbiaceae), Bathysa obovata (Rubiaceae), Palicourea guianensis (Rubiaceae) y Pithecoellobium angustifolium (Mimosoideae) (Beck et al., 1993; Paniagua et al., 2003).
Los cultivos ocupan un 7% de extensión dentro del PN Cotapata y un 8% en el área del presente estudio. Entre estos cultivos tenemos al café (Coffea arabica), naranja (Citrus sinensis), lima (Citrus limetta), mandarina (Citrus reticulata), plátano (Musa acuminata), palta (Persea americana), chirimoya (Annona cherimolia), guayaba (Psidium guajava) y siquili (Inga adenophylla) utilizado para dar sombra al café. También se pueden observar algunos cultivos pequeños de walusa (Xanthosoma sagittifolia), yuca (Manihot utilissima), racacha (Arracacia xanthorrhyza), maíz (Zea mayz), locoto (Capsicum pubescens) y pequeñas parcelas con coca (Erythroxylum coca) (Paniagua et al., 2003).
La fauna del bosque de Yungas ha sido descrita por Ribera (1995), Ríos (2001) y Roldán (2001) e incluye los mamíferos arbóreos como Bradypus variegatus (Xenarthra, Bradypodidae), Sciurus ignitus (Rodentia, Sciuridae), Potos flavus (Carnivora, Procyonidae), Ateles chamek (Primates, Atelidae) y Aotus azarae (Primates, Aotidae).
Metodología
Se escogieron seis sendas (Fig. 2, Tabla 1) ya existentes en los alrededores de la EBT, que se limpiaron y acondicionaron para realizar los conteos de C. libidinosus. Estas transectas pasan a través de cultivos y bosque, muestreando el hábitat de manera proporcional con su ocurrencia en el PN Cotapata. Si bien el método de conteos en transecta requiere que estos se hagan a lo largo de sendas rectas y establecidas al azar (Burnham et al., 1980; Rumiz, 1983; Whitesides et al., 1988; Buckland et al., 1993; Cullen y Valladares-Papua, 1997; Rocha, 1999; Thomas et al., 2002), esto no es viable en nuestro sitio de estudio, debido a la abrupta topografía de la zona y el elevado costo de apertura de sendas nuevas. Adicionalmente, en un estudio realizado con cinco especies de lémures, las estimaciones hechas en sendas preestablecidas no difirieron de aquellas hechas en sendas abiertas expresamente para el estudio; excepto para una especie, para la cual se verificaron diferencias en disponibilidad de recursos entre los dos tipos de sendas (Lehman, 2006).
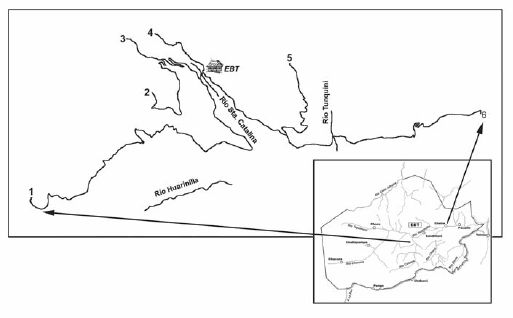
Fig. 2. Esquema del área de estudio, mostrando los senderos establecidos en los alrededores de la Estación Biológica Tunquini (EBT): (1) Sendero Bajo Hornuni 5576 m; (2) Sendero Plataforma 2935 m; (3) Sendero Riachuelo 1833 m; (4) Sendero Inciensal 1063 m; (5) Sendero Palmar 1633 m; (6) Sendero Chairo 4355 m.
Tabla 1
Senderos utilizados en el presente estudio, número de recorridos y total de recorrido para cada sendero (= transecta para el análisis con Distance). Tres de los senderos presentan cultivos. La longitud es expresada en kilómetros y en porcentaje del total recorrido por tipo de hábitat para cada sendero. El esfuerzo total de muestreo es expresado en horas.

El presente trabajo se realizó en los años 2002 y 2003. Las observaciones del año 2002 se realizaron entre mayo y octubre y sirvieron principalmente para afinar la toma de datos y estimar el número de días para el muestreo del año 2003, requeridos para obtener un mínimo de 60 observaciones para poder usar el programa Distance (v. 4.1) de manera adecuada en la estimación de densidad (Buckland et al., 1993; Rumiz, 1993; Rocha, 1999; Wallace, 1999; Thomas et al., 2002). También en el año 2002 se midió la longitud de cada sendero y se colocaron marcas cada 100 m para ayudar en el muestreo, y se estimó la disponibilidad de cada tipo de hábitat, midiendo la distancia atravesada por las sendas en cada uno (cultivos y bosque).
Por el bajo número de observaciones obtenidas por un observador en 2002 (16 grupos en total), los muestreos en 2003 se realizaron con tres observadores, los cuales caminaban simultáneamente sendas distintas. Las observaciones se realizaron entre mayo y julio, durante la época de cosecha de cítricos y la de mayor disponibilidad de éstos en las áreas de cultivos. Estos datos se usaron para estimar la densidad poblacional usando la metodología de transectas lineales y el programa Distance (v. 4.1; Burnham et al., 1980; Buckland et al., 1993; Thomas et al., 2002) y para evaluar el uso de hábitat.
Los recorridos se realizaron a pie, a una velocidad aproximada de un km/h, haciendo el menor ruido posible, y realizando paradas breves aproximadamente cada 100 a 200 m, con el fin de escuchar vocalizaciones y otros sonidos que nos ayuden a detectar la presencia de monos (Wallace, 1999). Se tuvo especial cuidado para no contar un grupo dos veces en el mismo sendero. Ante la duda, se desechó la observación.
Los recorridos se hicieron de ida y vuelta, considerando cada recorrido como independiente. Los recorridos de ida se hicieron entre las 8:00 y 12:00 y los de vuelta entre las 14:00 y las 18:00, dependiendo del estado del tiempo y la duración de la ida. Para cada grupo observado se registraron los siguientes datos: hora de encuentro, distancia perpendicular (estimada) del sendero al centro geométrico de cada grupo (Plumptre y Cox, 2006), distancia desde el comienzo del sendero, número de individuos observados, número estimado de individuos y tipo de hábitat donde se encontró el grupo (bosque o cultivo). No se realizaron conteos en condiciones de lluvia o neblina muy densa.
Análisis de datos
La abundancia relativa de C. libidinosus se expresó como el número de grupos y/o individuos por 10 km de transecta (Branch, 1983; Palomeque, 2001). Para estimar la densidad en los alrededores de la EBT, cada grupo fue considerado como un encuentro, con la finalidad de obtener una estimación de la densidad de grupos por km2 (Rocha, 1999). La densidad se estimó estratificando la muestra en senderos con presencia humana (aquellos con cultivos) y senderos sin presencia humana. Para calcular la densidad de individuos/km2 se tomó el promedio del número de individuos contabilizado en aquellos grupos que se logró censar con exactitud (n = 6). Debido a que las transectas no fueron localizadas al azar, consideramos que nuestra estimación es confiable sólo para la zona de los alrededores de la EBT (Cerros Tunquini y Hornuni). No recomendamos su uso para estimar el tamaño de la población de C. libidinosus en el PN Cotapata.
Para determinar el uso de hábitat se comparó la frecuencia de encuentros de grupos entre cultivos y bosques. La disponibilidad de cada tipo de hábitat se estimó midiendo la longitud de cada hábitat que atravesaba cada una de las transectas de conteo (Sainz, 1997; Rocha, 1999; Palomeque, 2001). Las observaciones se analizaron con una prueba de Chicuadrado (Zar, 1999), tomando como frecuencia esperada el número de grupos del total observado que correspondían al porcentaje calculado de disponibilidad para cada hábitat, bajo la hipótesis nula de que el k'usillo utiliza el hábitat de acuerdo a su disponibilidad.
RESULTADOS
El 2002 se hicieron entre ocho y 14 recorridos en cada transecta, con una longitud total recorrida de 207.86 km; en el 2003 se recorrieron 31-37 veces cada transecta, con un total de 588.79 km (Tabla 1). La diferencia en el número de recorridos por transecta se debe a que el estado del tiempo cambiaba entre la mañana y la tarde, resultando en que algunos días sólo se hacía un recorrido en alguna de las transectas.
Sólo seis de los 70 grupos encontrados en 2003 pudieron observarse de manera que el número de individuos del grupo fuera contabilizado con certeza. Para realizar la estimación de la densidad de individuos, sólo se tomó en cuenta el tamaño promedio de estos seis grupos que fueron contados con exactitud (13.5 DS = 6.95 individuos), que da una tasa de encuentro de 29.15 individuos/10 km de recorrido para el área de estudio. La dos mayores tasas de encuentro fueron para senderos sin cultivos, que alcanzaron registros 1.7 a 9.2 veces mayores que las encontradas en los tres senderos con cultivos. La mayor variabilidad se presentó en el sendero más perturbado, el de Chairo (Tabla 2).
Tabla 2
Abundancia relativa de grupos de Cebus libidinosus (número de encuentros /10 km de recorrido) para cada salida y sendero en el 2003 (CV= Coeficiente de Variación). * Senderos que presentan cultivos

Debido al bajo número de grupos observados a distancias mayores a 40 m, se truncaron los datos a esa distancia para su análisis con Distance, lo cual redujo a 52 el número de grupos observados. Se dejó que Distance 4.1 calculara una variedad de posibles estimadores y agrupaciones de datos y eligiera el modelo más adecuado para estimar la densidad, con base en el Criterio de Información de Akaike (AIC), siguiendo los protocolos sugeridos por Buckland et al. (1993), Peres (1999), Thomas et al. (2002), Gómez et al. (2003) y H. Gómez, WCS/Bolivia, comentario personal (2005). El modelo que mejor se ajustó a los datos fue el modelo "half-normal cosine". La densidad estimada fue de 2.16 ± 0.50 grupos/km2. El intervalo de confianza (95%) para la estimación indica que la densidad podría estar entre 1.28 y 3.64, con un ancho efectivo de la transecta de 20.44 m. Con esta densidad de grupos, la densidad de individuos sería de 29.1 (rango de17.3 a 49.1) ind./km2 (Tabla 3).
Tabla 3
Resumen de los resultados obtenidos con el programa Distance 4.1 para la estimación de densidad global de grupos de Cebus libidinosus, estratificada por presencia y ausencia humana y por cultivos y bosques, en los alrededores de la EBT.

El análisis de uso de hábitat se hizo solamente con las observaciones de las transectas que tenían tanto cultivos como bosque en su trayecto. Los monos mostraron una mayor presencia en los cultivos (12.1% del área), casi el doble (frecuencia observada = 8.0, frecuencia esperada = 4.1) de lo esperado de acuerdo con su disponibilidad; mientras que fueron ligeramente menos frecuentes en el bosque de lo esperado por su disponibilidad (87.9% del área; frecuencia esperada 29.9, frecuencia observada = 26; χ2 = 4.21, gl=1; p< 0.05).
DISCUSIÓN
Para realizar este estudio se utilizaron senderos ya establecidos en los alrededores de la EBT y, aunque estos no hayan sido establecidos al azar (Buckland et al., 1993; Thomas et al., 2002), se consideró que sí son representativos del área de estudio, puesto que atraviesan bosques y cultivos en proporción a la disponibilidad de cada uno de estos hábitats en el área general de estudio. El hecho que las transectas no fueran rectas es algo imposible de controlar en nuestro sitio de estudio y consideramos que esto no invalida nuestra estimación.
Es interesante que, si bien Cebus libidinosus usa las zonas de cultivo con mayor frecuencia de lo esperado, la mayor tasa de encuentro se dio en áreas sin cultivos (transectas Inciensal y Riachuelo), que tuvieron densidades relativas dos a nueve veces mayores que las transectas con menor densidad relativa, ambas en zonas con cultivos. Esto sugiere que, cuando los grupos de monos habitan cerca a los cultivos, tienden a usar estos como fuente de alimento, probablemente más que el propio bosque maduro; sin embargo, las mayores abundancias estarían en zonas alejadas de los cultivos. Esto podría ser resultado de dos factores no excluyentes: 1) la abundancia de monos en zonas de cultivo está reducida por la cacería de control que realiza la gente, y 2) el comportamiento de los monos que habitan zonas de cultivo es más cauteloso, lo cual podría haber ocasionado menor visibilidad de los mismos en las transectas que incluían cultivos. En todo caso, las observaciones apoyan la percepción del campesino en el sentido de que los monos son más frecuentemente vistos cerca a los cultivos, ya que los habitantes locales no caminan mucho por el bosque que está alejado de sus cultivos.
La densidad relativa de C. libidinosus en nuestro sitio de estudio es intermedia con relación a las reportadas en la literatura disponible, que abarca un rango entre 0.2 y 55 individuos/ km2 (Struhsaker et al., 1975; Klein y Klein, 1976; Izawa, 1976; Neville et al., 1976; Izawa y Yoneda, 1981; Freese et al., 1982; Terborgh, 1983; Terborgh y Jansen, 1984; Cámeron et al., 1989; Tabla 4); incluyendo zonas sin perturbación humana. Lamentablemente la bibliografía obtenida no indica el tipo de vegetación y/o la altitud para todos los estudios. Sin embargo, es posible que este estudio sea el que se ha realizado a mayor altitud. Esto último es importante, pues si bien el sitio de estudio representa un ambiente relativamente extremo para C. libidinosus en cuanto a la altitud, esto no implicó una densidad reducida con relación a sitios a menor altitud; lo cual da pie a especular que los ambientes yungueños no son menos productivos en cuanto a los recursos que requiere C. libidinosus.
Tabla 4
Tamaño medio de tropa (individuos) y densidad de Cebus libidinosus [= Cebus apella] expresado por distintos autores. La densidad se expresa como individuos/km2, con excepción de los casos que se indican con * donde es gr/km2.

Los campesinos indican que el k'usillo constituye una plaga porque logran alimentarse de una gran cantidad de frutos en un día, esto fue verificado por nuestras observaciones en campo y los datos de uso de hábitat, que muestran que los monos presentan marcada preferencia por los cultivos. Por lo tanto, nuestros resultados apoyan la percepción de la gente en cuanto a que C. libidinosus prefiere los cultivos en este sitio de estudio.
AGRADECIMIENTOS
Este estudio fue financiado por Wildlife Conservation Society, como parte del proyecto "Ecología de Mamíferos Grandes en el Parque Nacional y Área Natural de Manejo Integrado Cotapata", del Centro de Estudio en Biología Teórica y Aplicada (BIOTA). Un agradecimiento especial a Ruddy Ticona y Maritza Callisaya por su ayuda en el campo. También deseamos agradecer a la comunidad de Chairo por su afecto y a la administración del PN Cotapata por el apoyo logístico. Los comentarios de Humberto Gómez y Damián Rumiz contribuyeron a clarificar el manuscrito. Este trabajo fue presentado en el Primer Congreso Nacional de Mastozoología en Bolivia, realizado el 2005 en la ciudad de Cochabamba, Bolivia.
LITERATURA CITADA
1. AGUILAR MS. 2002. Dispersión de semillas por tres especies de primates Ateles chamek, Cebus apella y Saimiri sciureus en un bosque de la Estación Biológica del Beni. Tesis de Licenciatura inédita. Facultad de Ciencias Puras y Naturales. Universidad Mayor de San Andrés. La Paz, Bolivia. [ Links ]
2. ANDERSON S. 1993. Los mamíferos bolivianos: Notas de distribución y claves de identificación. Instituto de Ecología y Colección Boliviana de Fauna. Universidad Mayor de San Andrés. 159 pp. [ Links ]
3. APAZA AE. 2002. Comportamiento alimentario de Ateles chamek (Cebidae) y disponibilidad de frutos en época húmeda en los Yungas del PN-ANMI Cotapata. Tesis de Licenciatura inédita. Facultad de Ciencias Puras y Naturales. Universidad Mayor de San Andrés, La Paz, Bolivia. [ Links ]
4. BECK S, T KILLEEN y E GARCÍA. 1993. Vegetación de Bolivia. Pp. 6- 24, en: Guía de Árboles de Bolivia ( T Killeeen, E García y S Beck, eds). Herbario Nacional de Bolivia-Missouri Botanical Garden. Ed. Quipus. La Paz, Bolivia. [ Links ]
5. BRANCH LC. 1983. Seasonal and habitat differences in the abundance of primates in the Amazon (Tapajós) National Park, Brazil. Primates 24:424-431. [ Links ]
6. BUCKLAND ST, DR ANDERSON, KP BURNHAM y JL LAAKE. 1993. Distance sampling estimating abundance of biological populations. Ed. Chapman & Hall, London 446 pp. [ Links ]
7. BURNHAM KP, DR ANDERSON y JL LAAKE. 1980. Estimation of density from line transect sampling biological population. Wildlife Monographs Washington, D.C. 72:1-202. [ Links ]
8. CÁCERES CP. 2001. Determinación del comportamiento de Cebuella pygmaea en zonas con diferente influencia de las actividades humanas en el departamento de Pando. Tesis de Licenciatura inédita. Facultad de Ciencias Puras y Naturales. Universidad Mayor de San Andrés, La Paz, Bolivia. [ Links ]
9. CAMERON R, C WILTSHIRE, C FOLEY, N DOUGHERTY, X ARAMAYO y L REA. 1989. Goeldi's monkey and other primates in northern Bolivia. Primate Conservation 10:62-70. [ Links ]
10. CASTRO NR. 1977. Lineamientos para la conservación de primates en el Perú. Pp. 228-247, en: Primera conferencia internacional sobre la conservación y utilización de primates americanos no humanos en las investigaciones biomédicas (Lima, Perú, 2-4 de junio de 1975) OPS. Washington D.C. 3/7. [ Links ]
11. CULLEN LJR y C VALLADARES-PADUA. 1997. Métodos para estudos de ecologia, manejo e conservação de primatas na natureza. Pp. 239-269, en: Manejo e conservação de vida silvestre do Brasil (C Valladares-Padua, R Bodmer y L Cullen, organizadores). MCT (Ministério da Ciência e Tecnología), CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico). Directoria de Unidades de Pesquisa. Sociedade Civil Mamirauá. Ed. Litera Maciel Ltda. Rio de Janeiro. [ Links ]
12. DEFLER T y D PINTO. 1985. Censusing primates by transect in a forest of know primate density. International Journal of Primatology. 6:243-259. [ Links ]
13. EMMONS LH. 1999. Mamíferos de los bosques húmedos de América tropical, una guía de campo. Editorial FAN (Fundación Amigos de la Naturaleza), Santa Cruz de la Sierra, Bolivia 298 pp. [ Links ]
14. FORBES P, B MACKEITH y R PERBERDY. 1984. All the world's animals: primates. NewYork. Toronto: Torstar Books. [ Links ]
15. FREESE CH, PG HELTNE, NR CASTRO y G WHITESIDES. 1982. Patterns and determinants of monkey densities in Peru and Bolivia, with notes on distribution. International Journal of Primatology 3:53-90. [ Links ]
16. GARCÍA JE, V CASTELLÓ y M CORVILLO. 1987. Primeras apreciaciones de la densidad de Cebus apella y Saimiri sciureus en la Estación Biológica Beni, Bolivia. Ecología en Bolivia 10:15-27. [ Links ]
17. GARCÍA JE y T TARIFA. 1988. Primate survey of the Estación Biológica Beni, Bolivia. Primate Conservation 9:97-100 [ Links ]
18. GARCÍA JE y T TARIFA. 1991. Estudio de la comunidad de primates en la reserva de la biosfera "Estación Biológica del Beni", Bolivia. Ecología en Bolivia 7:1-14. [ Links ]
19. GÓMEZ H, G AYALA, RB WALLACE y F ESPINOZA. 2003. Densidad de la ardilla roja amazónica (Familia Sciuridae, Sciurus spadiceus) en el valle del río Tuichi (Parque Nacional y Área Natural de Manejo Integrado Madidi, La Paz, Bolivia). Ecología en Bolivia 38:79-88. [ Links ]
20. GÓMEZ H, G AYALA y RB WALLACE. 2005. Biomasa de primates y ungulados en bosques amazónicos subandinos en el Parque Nacional y Área Natural de Manejo Integrado Madidi (La Paz, Bolivia). Pp. 22; en: Resumen del Primer Congreso Nacional de Mastozoología en Bolivia. Cochabamba, Bolivia. [ Links ]
21. IZAWA K. 1976. Group sizes and composition of monkeys in the upper Amazon basin. Primates 17:367- 399. [ Links ]
22. IZAWA K y M YONEDA. 1981. Habitat utilization of non-human Primates in a forest of western Pando, Bolivia. Kyoto University Overseas Research. Report of new world monkeys. Kyoto University Primate Research Institute 13-22. [ Links ]
23. JANSON CH. 1986. Ecological and social consequences of food competition in brown capuchin monkeys. Unpublished Ph.D. dissertation, University of Washington, Seattle. http:/primate.wisc\Black-capped Capuchin (C. apella). htm (Extraído el 2002). [ Links ]
24. KLEIN LL y DJ KLEIN. 1976. Neotropical Primates: Aspects of habitat usage, population density and regional distribution in La Macarena, Colombia. Pp. 70-78, en: Neotropical Primates: Field studies and conservation (RW Thorington Jr. y PG Heltne, eds). National Academy of Sciences, Washington, D.C. [ Links ]
25. LEHMAN SM 2006. Effects of Transect Selection and Seasonality on Lemur Density Estimates in Southeastern Madagascar. International Journal of Primatology 27:1041-1057. [ Links ]
26. MARTINEZ JF. 2003. Funciones ecológicas de las vocalizaciones de Cebuella pigmaea (Callitrichidae) en el oeste del departamento de Pando. Tesis de Licenciatura inédita. Facultad de Ciencias Puras y Naturales. Universidad Mayor de San Andrés. La Paz, Bolivia. [ Links ]
27. MÉNDEZ PF. 1999. Influencia de la distribución y abundancia de frutos sobre el tamaño de subgrupos de Ateles chameck en la EBB, Bolivia. Tesis de Licenciatura inédita. Facultad de Ciencias Puras y Naturales. Universidad Mayor de San Andrés. La Paz, Bolivia. [ Links ]
28. MUÑOZ RA y DM KOPP. 2005. Abundancia de primates diurnos en la Estación Biológica Tahuamanu, Pando - Bolivia. Pp. 28, en: Resumen del Primer Congreso Nacional de Mastozoología en Bolivia. Cochabamba, Bolivia. [ Links ]
29. NEVILLE M, N CASTRO, A MÁRMOL y J REVILLA. 1976. Censusing primate populations in the reserved Area of the Pacaya and Samiria rivers, department Loreto, Peru. Primates 17:151-181. [ Links ]
30. PACHECO LF y JA SIMONETTI. 2000. Genetic structure of a mimosoid tree deprived of its seed disperser, the spider monkey. Conservation Biology 14:1766-1775 [ Links ]
31. PALOMEQUE ES. 2001. Estimación de abundancia y preferencia de hábitat del marimono (Ateles chamek) en el área de Chalalán (Parque Nacional y Área Natural de Manejo Integrado Madidi). Tesis de Licenciatura inédita. Facultad de Ciencias Puras y Naturales. Universidad Mayor de San Andrés. La Paz, Bolivia. [ Links ]
32. PANIAGUA ZN, GC MALDONADO y MC CHUMACERO. 2003. Mapa de vegetación de los alrededores de la Estación Biológica de Tunquini, Bolivia. Ecología en Bolivia 38:15-26 [ Links ]
33. PERES CA. 1999. General guidelines for stardardizing line-transect Surveys of tropical forest primates. Neotropical Primates 7:11-16. [ Links ]
34. PEREZ E y LF PACHECO. 2006. Damage by large mammals to subsistence crops within a protected area in a montane forest of Bolivia. Crop Protection 25:933-939 [ Links ]
35. PLUMPTRE AJ y D COX. 2006. Counting primates for conservation: primate surveys in Uganda. Primates 47:65-73 [ Links ]
36. RIBERA MO. 1995. Aspectos ecológicos del uso de la tierra y conservación en el Parque Nacional y Área de Manejo Integrado Cotapata. Pp 1-84, en: Caminos de Cotapata (CB Morales, ed.). Instituto de Ecología,FUND-ECO (Fundación para el Desarrollo de la Ecología) y FONAMA (Fondo Nacional del Medio Ambiente), IEA (International Energy Agency), La Paz, Bolivia. [ Links ]
37. RÍOS B. 2001. Presencia de mamíferos terrestres medianos y grandes en el Parque Nacional y Área Natural de Manejo Integrado Cotapata a través del uso de métodos indirectos. Ecología en Bolivia. 35:3- 16. [ Links ]
38. ROCHA NE. 1999. Censo de primates y evaluación de la metodología de líneas de transectas en Lago Caimán, Parque Nacional Noel Kempff Mercado. Tesis de Licenciatura inédita. Facultad de Ciencias Agrícolas, Universidad Autónoma René Moreno. Santa Cruz, Bolivia. [ Links ]
39. ROLDÁN AI. 2001. Manual de los mamíferos silvestres. Parque Nacional y Área Natural de Manejo Integrado Cotapata, Bolivia. WCS (Wildlife Conservation Society), National Audubon Society, BIOTA (Centro de Estudios en Biología Teórica y Aplicada). [ Links ]
40. RUMIZ D. 1983. Aplicación de métodos de censado para Alouatta caraya en hábitats fraccionados. Comparación de métodos de transecta. Pp. 233-239, en: Simposio de Primatología. Arequipa, Perú [ Links ].
41. RYLANDS AB y RA MITTERMEIER. 1982. Conservation of primates in Brazilian Amazonia. International Zoo Yearbook 22:17-37. [ Links ]
42. RYLANDS AB y A KEUROGHLIAN. 1988. Primate population in continuous forest and forest fragments in central Amazonia. Acta Amazónica 18(3-4):291- 307. [ Links ]
43. SAINZ BA. 1997. Censo de primates en un área de explotación forestal del Bajo Paragua. Tesis de Licenciatura inédita. Facultad de Ciencias Agrícolas. Universidad Autónoma Gabriel Rene Moreno. Santa Cruz, Bolivia. [ Links ]
44. STRUHSAKER TT, K GLANDER, H CHIRIVI y NJ SCOTT. 1975. A survey of primates and their habitats in northern Colombia. Pp 43-78, en: Primates Censusing Studies in Perú and Colombia. Pan American Health Organization. World Health Organization. Washington DC. [ Links ]
45. SUBCENTRAL 2 DE JULIO, SUBCENTRAL PACALLO Y ESTACIÓN BIOLÓGICA TUNQUINI INSTITUTO DE ECOLOGÍA. 2005. Plan de desarrollo rural sostenible. Sector del valle del río Huarinilla, Parque Nacional y Área Natural de Manejo Integrado Cotapata. Instituto de Ecología, Universidad Mayor de San Andrés, La Paz, Bolivia. [ Links ]
46. TARIFA T. 2000. Abundancia, distribución y estado de conservación del marimono (Ateles chamek) en la Reserva de la Biosfera Estación Biológica del Beni, Bolivia. Pp. 275-296, en: Biodiversidad, conservación y manejo en la región de la reserva de la biosfera Estación Biológica del Beni, Bolivia/ Biodiversity, conservation and management in the region of the Beni biological station biosphere reserve, Bolivia (Herrera-MacBryde, OF Dallmeier, B MacBryde, JA Comiskey y C Miranda, eds). Smithsonian Iinstitution/Man and Biosphere Program. Series No. 4, Smithsonian Institution, Washington, D.C. [ Links ]
47. TERBORGH J. 1983. Five new world primates: A study in comparative ecology. Princeton University Press. Princeton. New Jersey. [ Links ]
48. TERBORGH J y CH JANSEN. 1984. Ecology of primates in south-eastern Peru. National Geographic Society. Research Reports. [ Links ]
49. THOMAS L, T BUCKLAND, K BURHAM, D ANDERSON, J LAAKE, D BORCHERS y S STRINDBERG. 2002. Distance sampling. Pp. 544-552, en: Enciclopedia of environments (EL- Shaarawi a y W Piegorsch, eds.). Ed. John Wiley & Sons Ltd. Chichester. Volumen 1. [ Links ]
50. VARELA N. 2003. Aproximación a la biología, manejo y medicina de los monos maiceros. Boletín del Grupo de Estudio de Animales Silvestres (GEAS). Bogotá, Colombia. http://urras.portalveterinaria.com. 5(4):13 (Acceso en julio de 2002). [ Links ]
51. WALLACE RB. 1998. The behavioural ecology of black spider monkeys in north-eastern Bolivia. Tesis doctoral inédita. University of Liverpool. Liverpool, Reino Unido. [ Links ]
52. WALLACE RB. 1999. Transectas Lineales: Recomendaciones para el diseño, práctica y análisis. P. VI-1 - VI-14, en: Técnicas de investigación para el manejo de fauna silvestre: un manual del curso dictado con motivo del III Congreso Internacional sobre manejo de fauna silvestre en la amazonía (L Painter, D Rumiz, D Guinart, R Wallace, B Flores y W Townsend, eds.)., Santa Cruz de la Sierra, Bolivia. BOLFOR Documento Técnico 82/1999, Santa Cruz, Bolivia. [ Links ]
53. WALLACE RB y RLE PAINTER. 1999. A new record for Bolivia: an apparently isolated population of common woolly monkeys representing a southern range extension for the genus Lagothrix. Neotropical Primates 7:111-112. [ Links ]
54. WHITESIDES G, JF OATES, SM GREEN y RP KLUBERDANZ. 1988. Estimating primate densities from transects in a west African rain forest: a comparison of techniques. Journal of Animal Ecology 57:345 - 367. [ Links ]
55. ZAR JH. 1999. Biostatistical Analysis, fourth ed. Prentice-Hall International, Englewood Cliffs, New Jersey 662 pp. [ Links ]
Recibido 30 enero 2006.
Aceptación final 17 mayo 2008.

