










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. v.15 n.2 Mendoza jul./dic. 2008
Variación temporal de la quiropterofauna en un bosque de Yungas en Bolivia
M. Isabel Moya1*, Flavia Montaño-Centellas1, Luis Fernando Aguirre1, 2, Julieta Tordoya1, Jesús Martínez 1 y M. Isabel Galarza1
1 Centro de Estudios en Biología Teórica y Aplicada (BIOTA) - Programa para la Conservación de los Murciélagos de Bolivia (PCMB), Casilla 9641, La Paz, Bolivia. * <isamoyadiaz@yahoo.com>
2 Centro de Biodiversidad y Genética, Universidad Mayor de San Simón, Casilla 538, Cochabamba, Bolivia
RESUMEN: Se estudiaron las variaciones temporales de los patrones de riqueza de especies y abundancia de murciélagos de los gremios frugívoro, insectívoro y nectarívoro, durante 25 meses en un bosque montano de Yungas, Bolivia. En la época seca, la abundancia de individuos y la riqueza de especies de frugívoros y nectarívoros fue mayor, mientras que los insectívoros estuvieron mejor representados durante la época húmeda.
Las variaciones temporales en estos parámetros podrían estar relacionadas a la disponibilidad de los recursos alimenticios para cada gremio. Como se muestra en el presente trabajo, estudios a largo plazo son importantes para poder establecer variaciones temporales de la fauna de murciélagos con el fin de tener una idea más real del ensamblaje presente en bosques montanos andinos.
ABSTRACT: Temporal variation in a Bolivian montane Yungas forest bat fauna. We studied temporal variation in species richness and abundance of bats of three trophic guilds: frugivorous, insectivorous, and nectarivorous, over 25 months in a Montane Yungas Forest of Bolivia. Species richness and abundance of frugivorous and nectarivorous bat was greater in dry season than in wet season, whereas insectivore bats were better represented in wet season. These temporal variations could be related to seasonal availability of feeding resources for these guilds. As shown in this study, long term studies are crucial to establish temporal variation in order to have a better view of the assemblages of bats inhabiting Andean Montane Forest.
Palabras clave. Frugívoros; Gremios; Insectívoros; Murciélagos; Nectarívoros.
Key words. Bats; Frugivorous; Guilds; Insectivorous; Nectarivorous.
Los quirópteros constituyen el segundo orden más diverso de mamíferos en el mundo y de mayor éxito adaptativo, con más de 1000 especies (Altringham, 1996; Simmons y Conway, 2003; Simmons, 2005). En las selvas tropicales los murciélagos son particularmente diversos e importantes y representan entre el 40% y el 50% de las especies de mamíferos presentes en estos ecosistemas (Fleming et al., 1972; Fleming, 1988; Fenton et al., 1992; Cosson et al., 1999; Solari et al., 2006). Debido a su especialización trófica, los murciélagos tienen una importante participación en el reciclaje de nutrientes y energía en los ecosistemas (Fleming, 1982; 1988) e interactúan de una forma determinante en la naturaleza ya que ocupan numerosos niveles tróficos (McNab, 1982; Fenton et al., 1992; Galindo-González, 1998; Medellín y Gaona, 1999). En las comunidades de murciélagos tropicales, la riqueza y abundancia de especies dentro de cada gremio trófico es considerable; sin embargo, pueden variar a lo largo del tiempo. Uno de los factores que influye en estas variaciones temporales y, por tanto, en la ocurrencia de especies de murciélagos, es la estacionalidad de la oferta de recursos alimenticios (Dinerstein, 1986). En este sentido, cada gremio trófico tendría variaciones de riqueza de especies y abundancia propias, de acuerdo con el tipo de alimento que consuma.
Bolivia cuenta con una alta diversidad de especies de murciélagos (122) con representantes de cada uno de los gremios tróficos conocidos (Aguirre, 1999, 2007). Más específicamente, los bosques yungueños albergan 59 especies de murciélagos, 48% de las especies conocidas para Bolivia (Vargas y Patterson, 2007). Estos bosques se caracterizan por ser marcadamente estacionales, con meses donde la precipitación puede llegar a 680 mm y otros donde no sobrepasa los 15 mm (Arteaga, 2007). Las variaciones estacionales fueron consideradas para analizar la dinámica temporal de tres gremios tróficos de murciélagos (frugívoros, insectívoros y nectarívoros), durante dos años consecutivos.
El estudio fue realizado en el bosque húmedo montano de la Estación Biológica Tunquini, localizada al sudoeste del Parque Nacional y Área Natural de Manejo Integrado Cotapata, La Paz-Bolivia (68º02', 16º20' SO; 68º03', 16º05' NO; 67º43', 16º10' NE; 67º47', 16º18' SE; Ribera-Arismendi, 1995; Moraes, 2001). La zona se caracteriza por presentar laderas de fuerte pendiente y montañas expuestas con diferentes orientaciones (Ribera-Arismendi, 1995; Paniagua-Zambrana et al., 2003). El clima es considerado subtropical húmedo, con una temperatura media anual de 20.4 °C y una precipitación media anual de 2900 mm (Arteaga 2007). La zona tiene una fuerte intervención antrópica, registrándose diferentes tipos de ambientes como bosques de crecimiento secundario, bosques jóvenes con fisonomía de matorral denso, cultivos abandonados, cultivos activos y manchones de bosque de más de 20 años (Paniagua-Zambrana et al., 2003).
Los sitios de muestreo fueron seleccionados considerando las distintas unidades de vegetación. Fueron elegidos dos cultivos, dos barbechos o cultivos abandonados por al menos 5 años, dos bosques jóvenes de 15 y 20 años y dos bosques maduros de más de 25 años, todos los sitios estuvieron ubicados entre los 1400 y 1650 m de altitud. Los muestreos fueron realizados mensualmente entre junio de 2003 y junio de 2005, evitando las noches con luna llena (Reith, 1982; Jones y Rydell, 1994). El método de muestreo se basó en la captura de murciélagos con el uso de redes de neblina colocadas a nivel del sotobosque. Cada sitio fue muestreado dos días cada mes y en cada uno se colocaron 10 redes de neblina (7 redes de 6 m, 2 redes de 9 m y 1 red de 12 m), las redes fueron revisadas cada 45 min. y estuvieron activas desde las 18:00 hasta las 24:30. El esfuerzo de muestreo se midió como el producto del largo de las redes, el total del número de horas que las redes fueron abiertas cada noche, y el número de días de muestreo (Medellín, 1993). Totalizando un esfuerzo mensual promedio de 6305±1102 m*h de red. Cada ejemplar capturado fue identificado a nivel de especie con la ayuda de claves de campo (Anderson, 1993; Aguirre y Anderson, 1997; Vargas et al., 2006), marcado a través de perforaciones en el ala para reconocer las recapturas y finalmente liberado en el mismo sitio de captura.
Las especies fueron caracterizadas en tres gremios tróficos: insectívoros, nectarívoros y frugívoros, según el tipo de ítem que es dominante en sus dietas. El gremio de hematófagos no fue considerado debido al bajo número de capturas (5 individuos) y por tener una sola especie, Desmodus rotundus.
La riqueza de especies de cada gremio fue analizada corrigiéndola por el número de individuos capturados en 25 meses de muestreo. Para eliminar la influencia del orden en el cual los meses fueron adicionados al total, la muestra fue aleatorizada 1000 veces utilizando un análisis de rarefacción a través del programa EcoSim 7.0 (Gotelli y Entsminger, 2005), el cual permite comparar curvas de riqueza de especies basado en la abundancia de individuos capturados. Para determinar si existían diferencias en la abundancia de murciélagos capturados entre los diferentes sitios de muestreo (cultivos, barbechos, bosques secundario joven y bosque secundario maduro) se realizaron pruebas de Kolmogorov Smirnov para dos muestras independientes (Sieguel y Castellán, 1995). Estas pruebas permiten identificar si las tendencias de captura a lo largo del tiempo fueron similares entre los diferentes sitios. Finalmente, la variación temporal de la riqueza de especies y de la abundancia de individuos fue descrita dentro de cada gremio.
Fueron capturados un total de 2217 individuos pertenecientes a 29 especies de las familias Phyllostomidae, Vespertilionidae y Molossidae (Tabla 1). Del total de las capturas registradas el 95.5% fueron especies frugívoras de la familia Phyllostomidae, el gremio insectívoros estuvo representado por especies de las familias Vespertilionidae, Molossidae y Phyllostomidae (Micronycteris megalotis); mientras que el gremio nectarívoro, estuvo compuesto por tres especies, Choeroniscus minor, Anoura geoffroyi y Glossophaga soricina (Glossophaginae, Phyllostomidae).
Tabla 1
Especies de murciélagos capturados en dos años de muestreo en los alrededores de la Estación Biológica Tunquini. Gremios tróficos Ia = Insectívoro aéreo, N= nectarívoro, F = Frugívoro y H = Hematófago. Las especies marcadas con un * corresponden a las más abundantes dentro de cada gremio.

Con relación a la riqueza de especies, considerando dos muestras del mismo tamaño (N = 81) el número esperado de especies del gremio de frugívoros es menor (ES(n) = 7.9) que para el gremio de insectívoros (ES(n) = 9) (Fig. 1); es así que, para el total de frugívoros capturados (N = 2101) la curva de acumulación de especies alcanzó claramente una asíntota (Fig. 1), mientras que dentro del gremio insectívoro para el número total de muestra obtenida (N = 81) la curva no llegó a su asíntota (Fig. 1). Ambos grupos, frugívoros e insectívoros, superan en riqueza de especies al gremio de los nectarívoros representado sólo por tres especies.
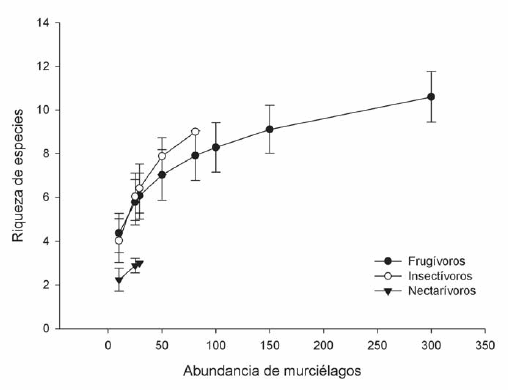
Fig. 1. Riqueza de especies corregida por la abundancia de individuos capturados para los gremios frugívoro, insectívoro y nectarívoro. La gráfica fue truncada para la abundancia de murciélagos frugívoros en 300 capturas de un total de 2000 con el objetivo de visualizar de forma más clara la tendencia de los otros gremios.
Diferencias en la abundancia de murciélagos fueron registradas entre los diferentes tipos de ambientes (Fig. 2 A). En el caso del gremio de murciélagos frugívoros las diferencias fueron entre el bosque secundario maduro y los cultivos (Z = 1.55; P = 0.018) y entre el bosque secundario maduro y los barbechos (Z = 1.83; P = 0.002). Las tendencias en la captura de murciélagos entre los bosques secundarios maduros y los cultivos fueron similares durante la mayor parte del tiempo y solo entre octubre y diciembre de 2004 estas tendencias fueron diferentes entre estos sitios. Las diferencias entre el bosque secundario maduro y los barbechos fueron marcadas durante casi todos los meses siendo los barbechos, donde menos abundancia de murciélagos frugívoros se capturó.
Para el gremio insectívoro, los sitios de mayor abundancia de individuos fueron los cultivos y bosques secundarios maduros (Fig. 2 B). Las diferencias en la abundancia de estos murciélagos fue entre los cultivos y barbechos (Z = 1.69; P = 0.006), y cultivos y bosques secundario joven (Z = 1.55; P = 0.015).

Fig. 2. Variación de la abundancia de murciélagos entre los diferentes tipos de ambientes donde se realizaron las capturas. A) Frugívoros, B) Insectívoros. Cultivos (•), Barbechos (o), Bosque secundario joven (![]() ), Bosque secundario maduro (
), Bosque secundario maduro (![]() )
)
El gremio frugívoro fue el más abundantes en junio y julio de 2003 y 2004, mientas que durante el 2005 sus abundancias permanecen relativamente constantes. Estos picos de abundancia coinciden con los períodos de fructificación de algunas especies vegetales de los géneros Vismia (Clusiaceae) y Piper (Piperaceae) (Loayza et al., 2006; Arteaga, 2007) el principal alimento de las especies de murciélagos frugívoros más abundantes como Carollia brevicauda, Carollia perspicillata, Sturnira lilium y Sturnira oporaphilum (Tordoya, 2006). De la misma forma, la mayor riqueza de especies de frugívoros fue registrada durante de junio de 2003, mayo, julio y septiembre de 2004 y junio del 2005 (Fig. 3 A, B). Estos murciélagos se constituyen en un grupo faunístico importante para el mantenimiento y regeneración de las selvas tropicales (Fleming y Heithaus, 1981; Fleming y Sosa, 1994) interviniendo en las estrategias reproductivas de especies de plantas pioneras como las del género Piper, Vismia y Solanum (Solanaceae). Estas especies de plantas son abundantes localmente (Paniagua-Zambrana et al., 2003) y ofertan recursos alimenticios a lo largo de todo el año, lo cual favorece la constante dominancia de estas especies
En el gremio insectívoro la mayor abundancia en el 2003 fue en los meses secos (junio y julio), mientras que en el 2004 fue registrado en los meses de mayor humedad (enero, noviembre y diciembre), tendencia que se mantuvo en el 2005 donde se observó una disminución en la abundancia durante la época seca. La mayor riqueza de especies fue registrada durante los meses de diciembre del 2003, enero, octubre, noviembre y diciembre del 2004 y 2005 (Fig. 3 C, D). La mayor abundancia durante los meses más húmedos del 2004 y 2005 podría deberse al inicio de laépoca húmeda la cual influye de forma positiva en la abundancia de insectos (Coley, 1990). Si bien, debido a los hábitos de forrajeo de los murciélagos insectívoros (Kalko y Handley, 2001) la abundancia de estos fue mucho menor a la de los frugívoros, el número de especies insectívoras acechadoras sedentarias (Vespertilionidae; Kalko, 1998), fue mayor que los insectívoros aéreos (Molossidae) probablemente porque estos últimos se caracterizan por volar muy alto y en lugares abiertos mientras que los murciélagos insectívoros asechadores sedentarios tienden a bajar al sotobosque más frecuentemente para buscar su alimento.
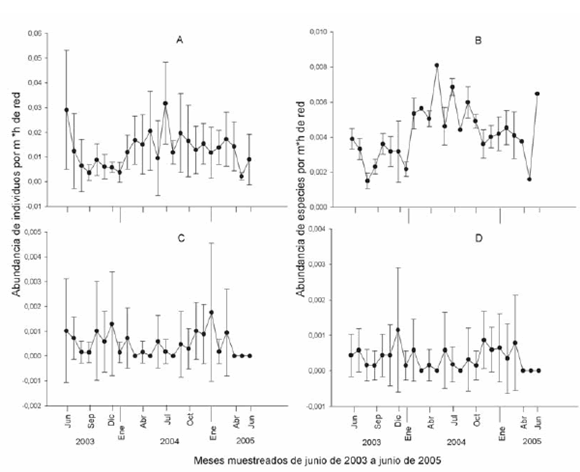
Fig. 3. Variaciones temporales de abundancia y riqueza de especies de murciélagos capturados en los alrededores de la Estación Biológica Tunquini durante 25 meses. (A) Abundancia de frugívoros capturados y (B) riqueza de especies del gremio frugívoro. (C) abundancia de insectívoros capturados y (D) riqueza de especies del gremio insectívoro.
Como la abundancia de murciélagos nectarívoros es muy baja, es difícil determinar sus patrones de ocurrencia en el área de estudio; sin embargo, es importante resaltar que el mayor número de individuos y de especies fueron capturados en la época seca, en junio de 2003 y abril, mayo y junio de 2004. Esta época coincide con el inicio de la floración de algunas especies de plantas como las del género Inga (Arteaga, 2007). La especie C. minor fue la más abundante en esta zona, a diferencia de lo observado en una zona adyacente situada a menor altitud (1300 m), en la cual, la especie más frecuente fue Glossophaga soricina (Moya, obs. pers.).
La composición de la comunidad de murciélagos en este bosque es el reflejo de un ambiente intervenido antrópicamente y con alta dinámica de formación de claros al interior de los bosques por la caída de árboles y derrumbes debido a la pendiente pronunciada. Las especies que predominan son las oportunistas frugívoras de áreas semiabiertas como C. perspicillata, C. brevicauda, S. lilium y S. oporaphilum, patrón de ocurrencia similar al encontrado por Medellín (2000) en cultivos y bosques tropicales de México. Este patrón es también encontrado en otros bosques neotropicales, y se sugiere que es el resultado del uso de los recursos superabundantes y la capacidad de desplazamiento sobre grandes distancias de las especies frugívoras (Kalko, 1998).
Estos resultados sugieren que las variaciones temporales en la riqueza de especies y abundancia en cada gremio, parecen estar relacionadas a la presencia de sus recursos alimenticios (frutos, néctar o insectos). Los patrones fenológicos de las plantas varían con las fluctuaciones climáticas de un año al otro (Chapman et al., 2005), al igual que la abundancia de insectos pueden responder a los patrones de precipitación o a la actividad antrópica. Para analizar esta hipótesis es importante elaborar trabajos a largo plazo que evalúen las variaciones temporales de la abundancia de individuos o de la riqueza de especies y su relación con la oferta de recursos alimenticios, que permitan comprender mejor la dinámica de la fauna de estos bosques de montaña. Por lo tanto, el estudio de las comunidades de estos animales debería seguir siendo una prioridad para proyectos de investigación y conservación.
AGRADECIMIENTOS
Agradecemos a los miembros del Programa para la Conservación de los Murciélagos de Bolivia (PCMB) y a la comunidad de Chairo por su apoyo y participación en el trabajo de campo. De igual manera a la Estación Biológica Tunquini del Instituto de Ecología por haber facilitado la logística del trabajo. Este proyecto fue financiado por la Fundación John D. y Catherine T. MacArthur y The Whitley Fund for Nature. Los comentarios realizados por la Dra. M. Díaz y otros revisores permitieron estructurar de mejor manera el manuscrito.
LITERATURA CITADA
1. AGUIRRE LF. 1999. Estado de conservación de los murciélagos de Bolivia. Andira 1:2-3. [ Links ]
2. AGUIRRE LF y S ANDERSON. 1997. Clave de campo para la identificación de los murciélagos en Bolivia. Documentos Ecología en Bolivia - Serie Zoología 5:1-38. [ Links ]
3. AGUIRRE LF, X VELEZ-LIENDO, A MUÑOZ y A SELAYA. 2003. Patrones de distribución y zoogeografía de los murciélagos de Bolivia. Revista Boliviana de Ecología 14:3-17. [ Links ]
4. AGUIRRE LF, MI GALARZA, K BARBOZA, A VARGAS, MI MOYA, L SILES, M TERAN, N BERNAL y D PEÑARANDA. 2007. Estado actual de conservación de los murciélagos de Bolivia. Pp. 114-116, en: Historia natural, Distribución y Conservación de los Murciélagos de Boliva (LF Aguirre, ed.). Centro de Ecología y Difusión Fundación Simón I. Patiño. Santa Cruz, Bolivia. [ Links ]
5. ALTRINGHAM JD. 1996. Bats: Biology and Behavior. Oxford University Press, New York. 262 pp. [ Links ]
6. ANDERSON S. 1993. Los mamíferos bolivianos: Notas de distribución y claves de identificación. Instituto de Ecología, La Paz. 159 pp. [ Links ]
7. ARTEAGA LL. 2007. Fenología y producción de semillas de especies arbóreas maderables en un bosque húmedo montano de Bolivia (PN ANMI Cotapata). Revista Boliviana de Ecología 21:55-66. [ Links ]
8. CHAPMAN CA, LJ CHAPMAN, TT STRUHSAKER, AE ZANNE, CJ CLARK y JR POULSEN. 2005. A long-term evaluation of fruiting phenology: importance of climate change. Journal of Tropical Ecology 21:1-14. [ Links ]
9. COLEY PD. 1990. Tasas de herbivorismo en diferentes árboles tropicales. Pp. 191-200, en: Ecología de un bosque tropical, ciclos estacionales a largo plazo. (EG Leigh, AS Rand y DM Windsor, eds.). Balboa, Rep. de Panamá [ Links ].
10. COSSON JF, J PONS y M MASSON. 1999. Effects of forest fragmentation on frugivorous and nectarivorous bats in French Guiana. Journal of Tropical Ecology 15:515-534. [ Links ]
11. DINERSTEIN E. 1986. Reproductive ecology of fruit bats and the seasonality of fruit production in a Costa Rican cloud forest. Biotropica 18:307-318. [ Links ]
12. FENTON MB, LD ACHARVA, D HICKEY, MB MERRIMAN, C OBRIST, MK SIME y DM ADKINS. 1992. Phillostomid bats (Chiroptera: Phyllostomidae) as indicators of habitat disruption in the Neotropics. Biotropica 24:440-446 [ Links ]
13. FLEMING TH. 1982. Foraging strategies of plant-visiting bats. Pp. 287-325, en: Ecology of bats (TH Kunz, ed.). New York. Plenum Press. [ Links ]
14. FLEMING TH. 1988. The short-tailed fruit bat. The University of Chicago Press Chicago. 365 pp. [ Links ]
15. FLEMING TH y ER HEITHAUS. 1981. Frugivorous bats, seed shadows, and the structure of tropical forests. Biotropica Supplement 13:45-53. [ Links ]
16. FLEMING TH y VJ SOSA. 1994. Effects of nectarivorous and frugivorous mammals on the reproductive success of plants. Journal of Mammalogy 75:845- 851. [ Links ]
17. FLEMING TH, ET HOOPER y DE WILSON. 1972. Three Central American bat communities: structure reproductive cycles and movement patterns. Ecology 53:555-569 [ Links ]
18. GALINDO-GONZALES J. 1998. Dispersión de semillas por murciélagos: su importancia en la conservación y regeneración del bosque tropical. Acta Zoológica Mexicana 73:57-74. [ Links ]
19. GOTELLI NJ y GL ENTSMINGER. 2005. EcoSim: Null models software for ecology. Version 7. Acquired Intelligence Inc. y Kesey-Bear. Jericho, VT 05365. [ Links ]
20. JONES G y J RYDELL. 1994. Foraging strategy and predation risk as factors influencing emergent time in echolocating bats. Philosophical Transactions of the Royal Society of London Series B. 346:445- 455. [ Links ]
21. KALKO EKV. 1998. Organization and diversity of tropical bats communities through space and time. Zoology 101:281-297. [ Links ]
22. KALKO EKV y CO HANDLEY-JR. 2001. Neotropical bats in the Canopo: diversity, community structure, and implications for conservation. Plant Ecology 153:319-333. [ Links ]
23. LOAYZA AP, RS RIOS y DM LARREA-ALCAZAR. 2006. Disponibilidad de recurso y dieta de murciélagos frugívoros en la Estación Biológica Tunquini, Bolivia. Ecología en Bolivia 41:7-23. [ Links ]
24. MCNAB BK. 1982. Evolutionary alternatives in the physiological ecology of bats. Pp. 151-200, en: Ecology of bats (Kunz TH ed.) Plenum Press. New York. [ Links ]
25. MEDELLIN RA. 1993. Estructura y diversidad de una comunidad de murciélagos en el neotrópico húmedo mexicano. Pp. 333-354, en: Avances en el estudio de los mamíferos de México (RA Medellin y G Ceballos, eds.). Publicaciones especiales, Vol. 1. Asociación Mexicana de Mastozoología, México. [ Links ]
26. MEDELLIN RA y O GAONA. 1999. Seed dispersal by bats and birds in forest and disturbed habitats in Chiapas, Mexico. Biotropica 31:432-441. [ Links ]
27. MEDELLIN RA, M EQUIHUA y M AMIN. 2000. Bat diversity and abundance as indicators of disturbance in Neotropical rainforests. Conservation Biology l4:1666-1675. [ Links ]
28. MORAES M. 2001. La Estación Biológica de Tunquini: un sitio favorable para el entrenamiento e investigación biológica y ecológica de los bosques montano de Bolivia. Ecología en Bolivia 35:1-2. [ Links ]
29. PANIAGUA-ZAMBRANA N, C MALDONADOGOYZUETA y C CHUMACERO-MOSCOSO. 2003. Mapa de vegetación de los alrededores de la Estación Biológica Tunquini, Bolivia. Ecología en Bolivia 38:15-26. [ Links ]
30. REITH CC. 1982. Insectivorous bats fly in shadow to avoid moonlight. Journal of Mammalogy 63:685- 690. [ Links ]
31. RIBERA-ARISMENDI MO. 1995. Aspectos Ecológicos, del uso de la tierra y conservación en el Parque Nacional y Área Natural de Manejo Integrado Cotapata. Pp.3-15, en: Caminos de Cotapata (CB Morales, ed.). Instituto de Ecología, FUND-ECO (Fundación para el Desarrollo de la Ecología) y FONAMA (Fondo Nacional del Medio Ambiente)- IEA (International Energy Agency). [ Links ]
32. SIEGEL S y NJ CASTELLAN. 1995. Estadística no paramétrica, aplicada a las ciencias de la conducta. Ed. Trillas. México 437 pp. [ Links ]
33. SIMMONS NB. 2005. Order Chiroptera. Pp. 312-529, en: Mammal species of the World: a taxonomic and geographic reference (Wilson DE y DM Reeder, eds.). Vol. 1. Johns Hopkins University. Baltimore, MD. [ Links ]
34. SIMMONS NB y TM CONWAY. 2003. Evolution of ecology diversity. Pp.493-535, en: Bat Ecology (TH Kunz y MB Fenton, eds.). The University of Chicago Press, Chicago. [ Links ]
35. SOLARI S, V PACHECO, L LUNA, PM VELAZCO y BD PATTERSON. 2006. Mammals of the Manu Biosphere Reserve. Pp. 13-22, en: Mammals and birds of the Manu Biosphere Reserve, Peru (BD Patterson, DF Stotz y S Solari, eds.). Fieldiana: Zoology, new series. [ Links ]
36. TORDOYA JE. 2006. Sobreposición de dieta de murciélagos frugívoros en cuatro tipos de ambientes en la Estación Biológica Tunquini (E.B.T.). Tesis Inédita de licenciatura. Universidad Mayor de San Andrés, La Paz, Bolivia 90 pp. [ Links ]
37. VARGAS A y PATTERSON B.D. 2007. Comunidades de Murciélagos Montanos de Boilivia. Pp. 82-88, en: Historia natural, Distribución y Conservación de los Murciélagos de Boliva (LF Aguirre, ed.). Centro de Ecología y Difusión Fundación Simón I. Patiño. [ Links ]
38. VARGAS A, MI GALARZA y LF AGUIRRE. 2006. Protocolo para el estudio de comunidades de murciélagos (Phyllostomidae). Pp. 12-22, en: Métodos estandarizados para el estudio de murciélagos en Bosques Montanos (I Galarza y LF Aguirre, eds.). BIOTA (Centro de Estudios en Biología Teórica y Aplicada). Cochabamba, Bolivia [ Links ]
Recibido 20 diciembre 2005.
Aceptación final 10 mayo 2008.

