










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. v.16 n.1 Mendoza ene./jun. 2009
ARTÍCULOS Y NOTAS
Latitudinal patterns of species richness distribution in South American carnivores
Monjeau, J. Adrián1*, Jorge A. Tort1, Jorge Márquez1, Pablo Jayat2, Benjamin N. Palmer Fry3, Sofía D. Nazar Anchorena1, Andrés Di Vincenzo1 and Francisco Polop1
1 Instituto de Análisis de Recursos Naturales (IARN), Universidad Atlántida Argentina, Arenales 2740, 7600 Mar del Plata, Buenos Aires, Argentina.
2 Laboratorio de Investigaciones Ecológicas de las Yungas (LIEY), Universidad Nacional de Tucumán, Casilla de Correo 34, 4107 Yerba Buena, Tucumán, Argentina.
3 Pembroke College, University of Cambridge, England.
*[Corresponding author: <amonjeau@parkswatch.org>]
ABSTRACT: The distribution and range of the 45 South American species of Carnivora (except Pinnipedia) currently recognized taxonomically was delimited from a detailed study of the literature. This study analyzes the ecogeographic patterns that operate at a continental scale in this family, which are particularly peculiar for not respecting natural landscape frontiers. The relation between the species distribution and latitudinal bands was evaluated using a Geographical Information System (GIS) with statistical regressions and correlations being carried out between geographical variables, areas, species richness, and latitude. The latitudinal bands with a higher species richness corresponded to the tropical bands with larger quantity of ecoregions, whereas a marked correlation was also found to exist between the area of the latitudinal bands, the area of the ecoregions and the species richness; however, a strong causeless correlation between area and latitude masked the individual contribution of these variables to the species richness. To assess the effect of latitude in the number of carnivore species we conducted a partial correlation analysis upon the number of species per band (SpL) and the latitude after removing the area effect. Species richness declined as latitude increased, but increased as area increased. We conclude that a combination of temperature and area is the predominant factor in the explanation of species richness distribution. This work supports the concept of an energetic gradient (the amount of energy available per surface unit) which gives rise to latitudinal gradients and the area effect.
RESUMEN: Patrones latitudinales de distribución de la riqueza específica en carnívoros sudamericanos. El área de distribución de las 45 especies sudamericanas de Carnivora (excepto Pinnipedia) reconocidas hasta el momento fue delimitada a partir de un detallado estudio de la bibliografía. Este estudio analiza los patrones ecogeográficos que operan a escala continental en este grupo taxonómico, particularmente caracterizado por no respetar las barreras naturales entre paisajes. Se evaluó la correspondencia entre la distribución de especies de carnívoros con bandas latitudinales utilizando un sistema de información geográfica (SIG). Se realizaron correlaciones y regresiones entre variables geográficas, áreas, riqueza de especies y latitud. Las bandas latitudinales con mayor riqueza específica correspondieron a las bandas tropicales con mayor cantidad de ecorregiones. Existe una marcada correlación entre el área de las bandas latitudinales, el área de las ecorregiones y la riqueza de especies. Sin embargo, una fuerte correlación no causal entre área de las bandas y latitud enmascara la contribución del área o la latitud per se a la riqueza de especies. Para estudiar por separado estos efectos se realizaron correlaciones parciales del número de especies por banda latitudinal removiendo el efecto del área. La riqueza de especies aumenta con el área de la banda y disminuye con el aumento de la latitud y de la elevación. La combinación de temperatura y área es el factor común predominante en la explicación de los patrones de variación en la riqueza de especies. Proponemos utilizar el concepto de gradiente energético, donde la riqueza de especies depende de la energía disponible por unidad de superficie, idea que incluye al efecto del área y a los gradientes latitudinales en un mismo patrón.
Key words. Carnivora; Energetic gradient; Macroecology; Neotropics.
Palabras clave. Carnivora; Gradiente energético; Macroecología; Neotrópico.
INTRODUCTION
There are several theories that try to explain why there are more species in the tropics than in temperate or cold areas (reviews in Rosenzweig, 1995; Brown and Lomolino, 1998). Birney and Monjeau (2003), studying various latitudinal changes in American marsupials, found a simple, previously unconsidered explanation to elucidate the well known pattern of decreasing biological diversity with increasing latitude: the extreme minimum temperature of the coldest month (from now on «minimum temperature») seems to be the «hidden guillotine» that reduces the proliferation of species as we come closer to the poles. With this, Birney and Monjeau (2003) explained the flip-side of the diversity phenomenon, that is to say, why there are few species in high latitudes. Another discovery of these authors was that the type of marsupial nutrition altered as latitude increased, with the southernmost all being carnivorous. A further need to explore the generality of these discoveries emerged from this study; would the same explanation be identifiable using the same macroecological analysis and environmental variables in other animal groups? As a way of testing the strength of the «hidden guillotine» rationale, an animal group insensitive to the spatial variations of the landscape was chosen, namely a group less expected to exhibit latitudinal changes. Significantly, Birney and Monjeau (2003) did not separate the effects of area from latitude in their study, although they were aware that both variables strongly correlate in South America. They were unable to truly demonstrate their «hidden guillotine hypothesis» without removing the confounding effect of area in the correlation between minimum temperature and species richness.
Carnivores are a group of crucial ecological importance among mammals because they do not seem to respect the barriers that the natural landscapes impose on other groups. The order Carnivora includes most of the terrestrial mammals specialized in predation on other vertebrates. This function of permanent natural disturbance has been recognized as fundamental in the maintenance of biodiversity in terrestrial communities (Soulé and Terborgh, 1999). Furthermore, their energy requirements necessitate large home ranges and give them the most extensive distributional ranges of all mammals (Rapoport, 1982). This last point explains why carnivores are among the groups of organisms that do not seem to recognize the physical barriers that generally cause ecotones among landscapes; however, no species of this family occupies the whole continent, so the factors that limit their geographical distribution continues to be a source of intrigue.
The ecological studies of carnivores are complicated for many reasons. In the current environmental scenario of the South American continent, the historical distributions of the native carnivores have been substantially reduced (Roig, 1991), now inhabiting only a group of small and isolated fractions of what was a broader, continuous area (Redford and Eisenberg, 1989, 1992). A recent study by the Wildlife Conservation Society mapped the last bastions of wilderness in the world, showing that the natural world is only a sprinkle of green («The Last of the Wild») in a matrix of totally modified landscape that the authors named the «Human Footprint» (Sanderson et al., 2002), meaning that environments capable to maintain viable populations of carnivores are becoming a rarity. Added to this, the population density is so low and the behavior so hidden that direct observations are scarce and difficult to obtain. These realities make the obtaining of field data for South American carnivores too laborious and frustrating. For this reason many scientists avoid the study of this family, in preference choosing more abundant groups that are easier to observe, hunt and manipulate by means of experiments, and which have fewer restrictions on the statistical analysis of the data gathered (Terborgh et al., 1999).
The somewhat anomalous behavior of the carnivores in relation to their physical environment could easily be considered as a disposable characteristic for biogeographers when choosing a biological group for testing theories of continental gradients. On the contrary, the very reason for the election of this group lies in its famed insensibility towards ecotones: if the tested hypothesis resists, it is a symptom of its strength (sensu Popper, 1985). The macroecological focus is the ideal recipe, since it analyzes the biological phenomenon at such a scale (the continental scale) that the biogeographic patterns emerge, circumnavigating the aforementioned difficulties encountered in more focused studies (Brown and Maurer, 1989; Brown, 1995; Birney and Monjeau, 2003).
Among these macroecological patterns, the one that has attracted most attention in the biogeographic literature is that of the decrease of species richness with the latitude. The papers that investigate this topic are numerous (see revisions in Brown, 1995; Rosenzweig, 1995; Brown and Lomolino, 1998). In this work, the role of area and latitude in the species richness distribution of terrestrial carnivores of South America is analyzed.
HYPOTHESES
Many hypotheses addressing latitudinal variation in species richness have been proposed (Pianka, 1966; Rohde 1992, Brown and Lomolino 1998). This paper focuses on the contribution of abiotic factors at a continental scale. These factors can be grouped in a few hypotheses, following the lead of Barcena et al. (2004) and Kaufman (1995).
1. Latitude
The species richness per band (SpL) of South American carnivores decreases with latitude (Pianka 1966; Rodhe, 1992; Kaufman, 1995; Brown and Lomolino 1998). The variable is L: average of the latitude range per band. Latitude is a surrogate of real environmental variables related to decrease of temperature towards the poles. The variables associated with latitude per band are: TM = annual mean temperature (°C); TminM = mean minimum temperature (°C); TmaxM = mean maximum temperature (°C); and TR = average of the temperature ranges (°C). The prediction: an inverse relationship between SpL and the selected variables. This hypothesis includes «the hidden guillotine» proposed by Birney and Monjeau (2003), as they suggested that the TminM is the ultimate cause for latitudinal patterns in marsupials. Prediction: an inverse relationship between the dependent variable (SpL) and L and related independent variables.
2. Area
The species richness (SpL) of South American carnivores increases with the area (Arrhenius, 1921; MacArthur and Wilson 1967; Brown and Lomolino, 1998). The variable associated is A: the area of each latitudinal band. Prediction: a direct relationship between SpL and selected variables.
3. Spatial heterogeneity
The more heterogeneous the physical environment is, the more diverse the associated biota will be (Pianka, 1966). Variables associated are BD: number of biomes per band and RdM: average of rainfall differences. Prediction: a direct relationship between SpL and associated variables.
4. Available energy
The species richness is limited by the available energy (Connell and Orias, 1964; Gaston 2000). Interaction of water and energy explains part of the variation in animal and plant species richness (Hawkins et al., 2003). The number of species varies along with the productivity of the environment (Tilman, 1982). Variables associated are TM and RM: mean annual rainfall. Prediction: a direct relationship between SpL and either TM or RM.
5. Combination of several hypotheses
In spite of Birney and Monjeau's (2003)«hidden guillotine» hypothesis, several authors believe that there are no single factors able to prove universality of the latitudinal pattern of species richness. An holistic, multivariate approach is proposed as a general hypothesis that combines several explanations (Kaufman 1995). Variables associated are L and A. Latitude is used as a surrogate of all the variables that co-vary with it, except Area of the band. Area is taken apart from the pool of latituderelated variables because it cannot be invoked as a cause of the latitudinal tendency but is simply a function of the shape of South America. Area is used as a surrogate of variables that co-vary with it. Prediction: the combination of L and A correlates with SpL.
MATERIAL AND METHODS
Sources of information
The macroecological studies or those where a continental scale of work is used are mostly based on secondary data from diverse sources of literature. In order to obtain and systematize this information, data was gathered from: 1) The complete list of species of South American Carnivora, compiled by Wilson and Reeder (1993) and upgraded by Wozencraft (2004); 2) The distributional range of each species based on Redford and Eisenberg (1989, 1992), Emmons (1990), Emmons and Feer (1992), Roig (1991), Anderson (1997), Eisenberg and Redford (1999), Jayat et al. (1999) and sources from the internet (http//:www.infonatura.org). The area of continental distribution of each species was defined combining several authors' approaches and the methods proposed by Rapoport and Monjeau (2001). The refinement of the borders of each distribution was carried out with the Arc View 3.2 program; 3) The biome types used for comparing the distribution of each one of the species enumerated in the Appendix were obtained from Redford and Eisenberg (1992), which is a modification of Udvardy (1978), with the following exceptions: for Bolivia Anderson's map was used (1997:75, his figure 470); for Patagonia the classification of landscapes and nomenclature proposed by Monjeau et al. (1998) was used; the biome Pampas was considered as a compact unit, because we found no consistent justification to distinguish between the Argentinean and Uruguayan Pampas, as it is made in Redford and Eisenberg (1992); the «Madeiran» of Udvardy (1978) was included within the more broadly known Amazon for the purpose and scale of this work. The biomes were digitized and georeferenced using Arc View 3.2 and Arc Map 8.2. The overlap between species distributional range and biome type and quantity were made using Arc View 3.2. The information about biome preferences in the Appendix is based on Redford and Eisenberg (1989, 1992), Emmons (1990), Roig (1991), Anderson (1997), Eisenberg and Redford (1999), Jayat et al. (1999) and sources from the internet (http//:www.iucnredlist.org; http//:www.infonatura.org).
Selection, treatment and data analysis
For the meteorological data, the continent was divided in a series of 13 latitudinal bands (Fig. 1). Each band covers 5º of latitude, which is approximately equivalent to 555 km of width, using as a reference the cartography of a Lambert plane projection of South America with the same azimuthal area extracted from a commercial Atlas (Rand McNally Atlas). In this way, the study area consisted of 13 latitudinal bands. The most northerly band was 10° N, which excludes from the study area a small land portion at the north end of the continent. This exclusion was necessary to be able to locate with easiness the projections of latitude published in the maps. Also, the most southern band of 55° S left out a small insular portion for the same reason. These latitudinal bands were numbered one to 13 from North to South. The election of the width of the latitudinal bands was made exclusively in order to compare later to the results with those from Birney and Monjeau (2003). For the assignment of presence of carnivore species in each latitudinal band, it was considered that if some part of the compiled distribution of each species coincided with a latitudinal band, each one of this species should be registered as if its distribution is present in that band. Arc Map 8.2 was used to geo-reference the maps of distribution of each species and to assess the correspondence with biomes, temperature, rainfall, and area of the latitudinal bands. The area of each latitudinal band was calculated from coast to coast, excluding the islands, using the area calculation tool of Arc View 3.2. Data of climate and other environmental data were taken from Prohaska (1976) for Argentina, Uruguay and Paraguay; Miller (1976) for Chile; Johnson (1976) for Peru, Bolivia and Ecuador; Ratisbona (1976) for Brazil; and Snow (1976) for countries located mainly or completely north of the equator in the book of climates of Central America and South America (Schwerdtfeger, 1976). For each latitudinal band five representative places were selected for the following areas: A) Pacific coast, B) a high Andean site, C) the west end of the areas located to the east of the Andes, D) the town closer to the central point in longitude, between places C and E, and E) Atlantic coast. There were no available data for the places corresponding to C in band 1, D in band 10, C and D in band 11, and B and C in bands 12 and 13. For each selected place data of latitude, longitude, elevation, annual mean temperature, annual maximum temperature, annual minimum temperature, mean annual rainfall, average of the minimum monthly rainfall, and average of the maximum monthly temperature were registered. The geographical latitudinal coordinates were transformed into numeric values so that the degrees of the center of the latitude band were expressed using hundredth of minutes (Eg. 7.5° instead of 7º 30'). The temperature differential was calculated as the difference between the annual maximum temperature and the annual minimum temperature and the fluctuation of rainfall was calculated as the difference between the monthly minimum rainfall and the monthly maximum rainfall. There were no mean temperature values (as well as associated parameters) for the B places (Andes), and so these were excluded. The inclusion of these places increased the variance for each band and it influenced the means of each band due to the elevation of each B place, and not because of its latitude. The influence of elevation in rainfall is a lot deeper and predictable than it is on temperature, and, because of this, the values of mean rainfall for each band in our analyses include climatic data for all the selected places.
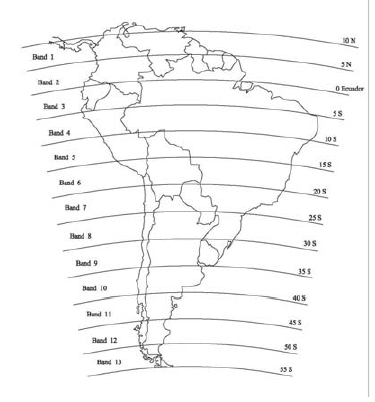
Fig. 1. Map of South America showing the latitudinal bands used in this study.
Statistical methods
We first tested each hypothesis separately relying on bivariate analysis. Because some of the variables selected are related with latitude we tested the correlation among them. Simple regressions using the program STATISTICA 5.0 were carried out to find the variable that best predicts the species richness of South American carnivores. To assess the latitudinal trend in species richness, addressing the first and more general hypothesis, we performed correlation analyses between species richness and L. We also made correlations with TM, TminM («the hidden guillotine» of Birney and Monjeau, 2003), TmaxM and TR. To assess the influence of the area in the species richness we correlated the number of species per band versus A (Hypothesis 2). To test the relationship between spatial heterogeneity and the species richness we correlated the number of species per band versus BD, TR and RdM (Hypothesis 3). To test the prediction of the Hypothesis 4 we made a correlation analysis between species richness and TM and RM. To assess the effect of latitude per se in the number of carnivore species we conducted a partial correlation analysis on the number of species per band (SpL) and L after removing for the effect of A. This is akin to evaluating the correlation between the residuals of the simple regression analysis of species number and area with those of latitude and area. Given the macroecological perspective of the study, a demanding significance level of 5% was accepted.
RESULTS AND DISCUSSION
As a result of the existing work, physical data of the South American continent were easily obtained for each latitudinal band (Fig. 1 and Table 1). The updated checklist of the species of South American carnivores is compiled in the Appendix together with information about band and biome distribution. The range of distribution of each species is shown in the group of maps of Fig. 2. As a result of calculating how many species overlap in each latitudinal band, a species richness measurement was obtained for each band. Meteorological information, area of each band, and biome diversity are presented in Table 1.
Table 1. Abiotic variables and species richness considered in this study. Bands = band number from North to South; Latitude = average of the latitude in each band (in degrees and hundredth of minutes); SpL = species richness for each latitudinal band; TM = annual mean temperature (°C); TminM = mean minimum temperature (°C); TmaxM = mean maximum temperature (°C); TR = average of the temperature ranges (°C); RM = mean annual rainfall (mm); RdM = average of rainfall differences (mm); A(x 103) = area of each band in square kilometers; BD = biome diversity per band.


Fig. 2. Distributional ranges of South American carnivores. 1, Puma yagouaroundi; 2, Conepatus humboldtii; 3, Procyon cancrivorus; 4, Lontra provocax; 5, Eira barbara; 6, Lontra felina; 7, Puma concolor; 8, Lontra longicaudis; 9, Leopardus guigna; 10, Pteronura brasiliensis; 11, Lyncodon patagonicus; 12, Potos flavus; 13, Lycalopex vetulus; 14, Galictis cuja; 15, Bassaricyon beddardi; 16, Leopardus braccatus; 17, Leopardus pajeros; 18, Urocyon cinereoargenteus; 19, Chrysocyon brachyurus; 20, Lycalopex griseus; 21, Leopardus tigrinus; 22, Mustela felipei; 23, Conepatus chinga, 24, Leopardus wiedii; 25, Lycalopex sechurae; 26, Bassaricyon gabbii; 27, Lycalopex culpaeus; 28, Mustela africana; 29, Lycalopex gimnocercus; 30, Conepatus semistriatus; 31, Leopardus geoffroyi; 32, Leopardus pardalis; 33, Nasua narica; 34, Cerdocyon thous; 35, Nasuella olivacea; 36, Bassaricyon alleni; 37, Mustela frenata; 38, Tremarctos ornatus; 39, Speothos venaticus; 40, Leopardus colocolo; 41, Atelocynus microtis; 42, Nasua nasua; 43, Panthera onca; 44, Galictis vittata; 45, Leopardus jacobitus.
Table 2 shows the existent correlations among variables. Because latitude correlates with area due to the shape of the continent, there are non causal correlations between area and meteorological variables. Due to the fact that area correlates with biome diversity, we cannot invoke causality in the correlations between biome diversity and latitude and their correlated variables. Table 3 shows the correlations obtained among species richness and the physical variables of the South American continent chosen for this work.
Table 2. Coefficient of correlation among variables considered in this paper. Bold numbers indicate a significant correlation at p<0.05. Latitude variable expresses the mean latitude of each band; TM = annual mean temperature (°C); TminM = mean minimum extreme temperature (°C); TmaxM = mean maximum extreme temperature (°C); TR = average of the temperature ranges (°C); RM = mean annual rainfall (mm); RdM = average of rainfall differences (mm); A (x 103) = area of each band in square kilometers; BD = biomes diversity.
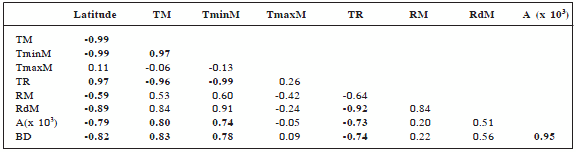
Table 3. Bivariate regression equations of species richness per latitudinal band (SpL) on the environmental variables that were significant (p<0.05). Latitude = average of the latitude in each band (in degrees and hundredth of minutes); TM = annual mean temperature (°C); TminM = mean minimum extreme temperature (°C); TR = average of the temperature ranges (°C); A(x 103) = area of each band in square kilometers; BD = biome diversity per band.

Hypothesis 1
Species richness decreases with latitude
Based on the bivariate approach (Table 3) a marked correlation exists between the species richness per band and latitude (Fig. 3). The pattern of increasing diversity towards the equator is one of the most strongly supported paradigms of biogeography (Darlington, 1957). Ruggiero (1994) and Kaufman (1995) reported correlations between latitude and species diversity of carnivores in South America and the whole American continent, respectively. Our results supported this pattern; however, the underlying process explaining why it occurs still warrants discussion. The best surrogate for latitude is temperature (Table 2). Within temperature variables, the mean minimum extreme (MminT) correlated with latitude equals the mean annual temperature (TM), whereas the mean maximum (MmaxT) does not. In addition to this, we found a strong correlation between MminT and TM but no correlation between TM and MmaxT. That is, the latitudinal gradient must be driven by the minimum extremes of the thermal range, as stated by Birney and Monjeau (2003). We support Birney and Monjeau's (2003) finding of the minimum extreme temperature as a plausible limiting factor explaining why there are fewer species outside the tropics (Blackburn and Gaston 1996), rather than explaining why there are so many species in the tropics. Extremely cold temperatures at high latitudes are the «hidden guillotine» that may indeed reduce the proliferation of species. This idea is related with the available energy hypothesis (Connell and Orias, 1964; Gaston 2000) and also with the environmental favorablenessstress hypothesis discussed in Barcena et al. (2004). Because species richness is favored when the mean values of the environmental variables lie within the optimal range for the physiological needs of the species, low temperatures act as strong selective filters having a direct effect on the number of species able to survive in harsh conditions. The weak side of this explanation is the correlation between Area and MminT (Table 2) in the particular case of South America masking the contribution of this hypothesis per se.
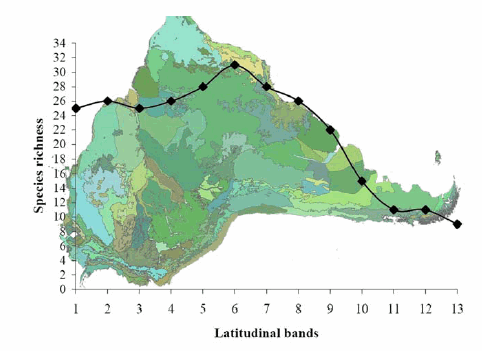
Fig. 3. Relationships between species richness of South American carnivores and latitudinal bands.
Hypothesis 2
Species richness increases with area
Table 3 shows a significant correlation between the area of the latitudinal band and the species richness. Band area is in turn strongly correlated with biome diversity per band (Table 2). Biome diversity is in turn correlated with species richness (Table 3), which we consider the ultimate cause of the area effect in species diversity. This correlation provides an alternative explanation to Hypothesis 1: as the area of the band increases, the number of biomes increases. The more heterogeneous the physical environment contained in the band, the more diverse and complex the associated biota will be, providing the opportunity for carnivore proliferation. This finding is consistent with the well supported relationship between species diversity and area (Arrhenius, 1921; MacArthur and Wilson, 1967; Rosenzweig, 1995; Brown and Lomolino, 1998) and other studies discussing the pattern (Ruggiero, 1999; Ruggiero et al., 1998). Again, the weakness of this explanation is the non-causal correlation between Area and latitude and their related meteorological variables. We cannot invoke the influence of the area per se on species richness in South American carnivores.
Hypothesis 3
The more heterogeneous is the physical environment, the more diverse is the associated biota
Table 3 shows a significant correlation between species richness and the number of biomes per band (BD) and the average of the temperature ranges (TR), both variables assumed to be related to habitat heterogeneity. The hypothesis is supported, but is clearly included in the explanation of hypothesis 2. Because BD and area are correlated, we cannot invoke the independence of hypothesis 3 as an explanatory principle.
Hypothesis 4
The species richness is limited by the available energy
We used a combination of temperature (TM) and precipitation (RM) as surrogates of available energy for carnivores per band. We only found correlation between TM and species richness whereas no correlation exists between species richness and precipitation (Table 3). Our results cannot support this hypothesis solely with these data. The relation with the mean temperature is included in the hypothesis 1. As in Birney and Monjeau (2003) for marsupials, no significant correlation between species richness and rainfall was found in this work during the band analysis. Although a demonstrated relationship exists between productivity and rainfall and at the same time between productivity and species richness (Brown and Lomolino, 1998), in this work there was no clear evidence that rainfall increases the diversity of carnivores. The result goes against what is observed directly in field experience, so it is suggested that the processes and mechanisms that increase the diversity of carnivores with rainfall work at a local or regional scale, in microgradients of humidity that respond to the geology and geomorphology of certain landscapes.
Hypothesis 5
The latitudinal pattern of species richness is better explained by a combination of variables
Latitudinal bands with higher species richness correspond to the tropical ones with more Area and greater biome diversity. Hypotheses 1 and 2 provide alternative explanations of the species richness. There is, however, a strong correlation between area and latitude (Table 2), but of course without any possible causality with biological meaning. In the case of the South American continent, the area of each latitudinal band decreases as the latitude increases, confusing the effects of one or the other variable on the fall of species (Birney and Monjeau, 2003). This correlation masks the contribution of the area or the latitude per se to species richness. To solve this problem we conducted partial correlation analyses on the number of species per band (SpL) and latitude after removing for the effect of area, and on the number of species per band and area after removing the effect of latitude.
The relationship between the species richness per band and latitude after removing the effect of area shows a non significant correlation (partial r = -0.417, p=0.17). The relationship between the species richness per band and the area of the band per se (after removing the effect of latitude) is also not statistically significant (partial r = -0.402, tobs associated is 1.38, c P=0.19).
A multiple regression analysis between species richness per band as dependant variable and latitude + area + latitude x area shows that the effect of latitude and its interaction with area are both highly significant:
SpL= 28.5 - 0.47 x L - 0.001 x A 0.0004 x L x A (R2= 0.91, p<0.001) (1)
Fig. 4 shows how the species richness decreases towards the pole (high latitude) and decreases with the area of the band. The combined effect of both variables in species richness becomes apparent south of the tropics. Area and latitude are correlated (Table 2) and their combined effect on species richness per band is highly significant; however, the contribution of each variable per se cannot be separated by partial correlation.
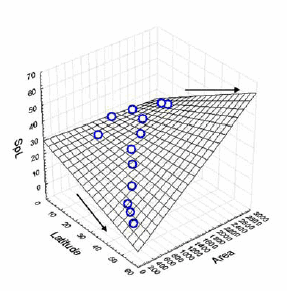
Fig. 4. Relationship between species richness of South American carnivors and the combined effect of latitude and area.
Many studies on North American diversity gradients have been carried out, where the area of each band increases as the latitude increases and in spite of it, the biodiversity equally falls (see reviews in Brown, 1988; Brown and Lomolino, 1998; Rosenzweig, 1995; a clear example can be seen in Kaufman, 1995: 330, figure 5). On the other hand, it is well known that the tropical countries of Central America, such as Costa Rica, Panama or Guatemala, are very specious in spite of their small surface. Extreme counter-examples are Antarctica and the Holarctic region, with great areas available but hosting very few terrestrial species. In the same way it can be deduced that the decrease of species with latitude in the world's oceans cannot be due to the species-area relationship and, on the other hand, can be explained by the theory of the minimum temperature suggested by Birney and Monjeau (2003), as well as the continental pattern. This would suggest that the area has important effects on species richness but does not exercise a significant influence on the whole latitudinal pattern of species fall. The result of our work is perhaps a particular case driven by the shape of South America.
In searching for the universality of the latitudinal pattern, our studies suggest an explanation for South America that should be tested worldwide: the latitudinal pattern of species richness may be driven by the amount of energy (temperature) per surface unit (area), as stated in equation 1. The extensive distributional ranges of large carnivores (Fig. 2) related to their high energetic demands may be one of the reasons explaining why they are the first to go extinct on islands across the world and in isolated national parks. The higher the latitude, the lower the minimum extreme temperatures (Table 1). This lower available energy per surface unit can result either in a shutdown of food for long periods or in an unsustainable balance between energetic demands and prey available.
For an homoeothermic organism such as a carnivore, the major thermodynamic challenge consists of obtaining more energy from its prey than that it expends in catching them. As the minimum temperature becomes more extreme with latitude, the maintenance of the constant internal temperature becomes more and more expensive and the diversity of prey becomes scarcer as a result of the same studied pattern. These two factors can lead to a morpho-functional specialization in the capture of very specific prey (reduction of the niche width), or indeed the development of resistance mechanisms such as hibernation, fat accumulation or becoming granivorous (all nonexistent in South American Eutherian carnivores). These processes all reduce the adaptative possibilities that support cladogenetic production (the «hidden guillotine»), in stark contrast to what happens in the relatively benign tropics.
Future directions of this research
It is important to recognize that this type of analysis considers the comparative method suitable to hypothesize on the adaptative character of the distribution of species regarding latitude. On the other hand, if the «hidden guillotine» is invoked as one of the key factors impeding the proliferation of species, our next work will focus on studying the relationship between elevation and carnivore species richness in South America (since temperature is a common physical factor inversely correlated with latitude and altitude). The equivalence between elevation and latitude is obvious for an educated traveler: the summit habitat of equatorial volcanoes such as Chimborazo or Cotopaxi being as hostile for life as Antarctica.
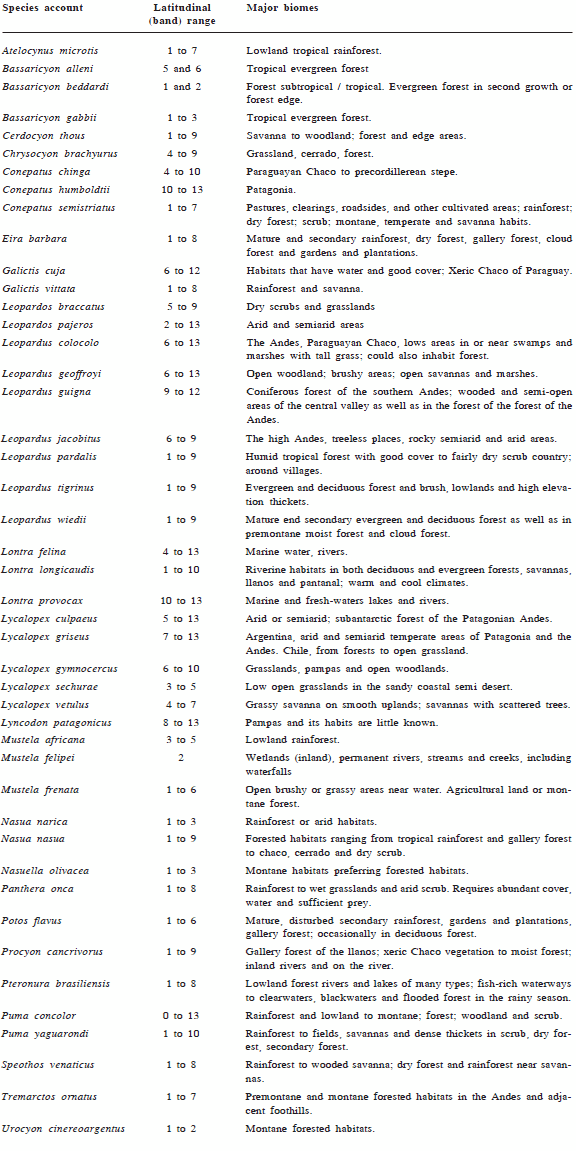
Species account and their correspondence with latitudinal bands and major biomes.
ACKNOWLEDGMENTS
Research was supported by the Consejo Nacional de Investigaciones Científicas y Técnicas, the Instituto de Análisis de Recursos Naturales, Universidad Atlántida Argentina, and the Laboratorio de Investigaciones Ecológicas de las Yungas (LIEY), Universidad Nacional de Tucumán. Juanjo Morrone provided detailed editing of previous versions of this work, Jesús Olivero and an anonymous reviewer provided invaluable help with comments, discussion, new references and good ideas. We thank Marcelo Kittlein for his advice in statistics. We are especially grateful to the late Elmer Birney for the discussion of this ideas and general theory on latitudinal trends. John Terborgh and Eddy Rapoport provided enlightening brainstorming on latitudinal patterns.
LITERATURE CITED
1. ANDERSON S. 1997. Mammals of Bolivia, taxonomy and distribution. Bulletin of the American Museum of Natural History Nro. 231, New York. [ Links ]
2. ARRHENIUS O. 1921. Species and area. Journal of Ecology 9:95-99. [ Links ]
3. BARCENA S, R REAL, J OLIVERO, and JM VARGAS. 2004. Latitudinal trends in breeding waterbird species richness in Europe and their environmental correlates. Biodiversity and Conservation 13:1997- 2014. [ Links ]
4. BIRNEY EC and JA MONJEAU. 2003. Latitudinal patterns in South American Marsupial Biology, Pp. 293-313, in: Carnivorous with pouches: Biology of carnivorous marsupials (M Jones, C Dickmann and M Archer, eds.). SCIRO Publishing, Inc., Melbourne, Australia. [ Links ]
5. BROWN JH. 1988. Species diversity. Pp. 57-88, in: Analytical biogeography (AA Myers y PS Giller eds.). Chapman and Hall, London. [ Links ]
6. BROWN JH. 1995. Macroecology. Chicago University Press, Chicago. [ Links ]
7. BROWN JH and M LOMOLINO. 1998. Biogeography. Second Edition. Sinauer Associates, Sunderland, Mass. [ Links ]
8. BROWN JH and BA MAURER. 1989. Macroecology: the division of food and space among species on continents. Science 243:1145-1150. [ Links ]
9. CONNELL JH and E ORIAS. 1964. The ecological regulation of species diversity. American Naturalist 98:399-414. [ Links ]
10. DARLINGTON PJ. 1957. Zoogeography: the geographical distribution of animals. John Wiley & Sons, New York. [ Links ]
11. EISENBERG JF and K REDFORD. 1999. Mammals of the Neotropics. Vol. 3. The Central Neotropics. Ecuador, Peru, Bolivia, Brazil. The University of Chicago Press. Chicago and London. [ Links ]
12. EMMONS LH. 1990. Neotropical rainforest mammals. A field guide. University of Chicago Press, Chicago. [ Links ]
13. EMMONS LH and F FEER. 1997. Neotropical rainforest mammals. A field guide, Second edition. University of Chicago Press, Chicago. [ Links ]
14. GASTON KJ. 2000. Global patterns in biodiversity. Nature 405: 220-227. [ Links ]
15. HAWKINS BA, R FIELD, HV CORNELL, DJ CURRIE, JF GUEGAN, DM KAUFMAN, JT KERR, GG MITTELBACH, T OBERDORFF, EM O´BRIEN, EE PORTER, and JRG TURNER. 2003. Energy, water, and broad-scale geographic patterns of species richness. Ecology 84:3105-3117. [ Links ]
16.JAYAT JP, RM BARQUEZ, MM DÍAZ, and PJ MARTINEZ. 1999. Aportes al conocimiento de la distribución de los carnívoros del noroeste de Argentina. Mastozoología Neotropical 6:15-30. [ Links ]
17. JOHNSON AM. 1976. The climate of Peru, Bolivia and Ecuador. Pp. 147-218, in: Climates of Central and Southern America, World Survey of Climatology. (W Schwerdtfeger, ed.). Elsevier, Amsterdam. [ Links ]
18. KAUFMAN DM. 1995. Diversity of new world mammals: Universality of the latitudinal gradients of species and bauplans. Journal of Mammalogy 67:322-334. [ Links ]
19. MACARTHUR RH and EO WILSON 1967. The theory of island biogeography. Monographs in Population Biology 1:1-203. [ Links ]
20. MILLER A. 1976. The climate of Chile. Pp. 113-145, in: Climates of Central and Southern America World Survey of Climatology. (W Schwerdtfeger, ed.). Elsevier, Amsterdam. [ Links ]
21. MONJEAU JA, EC BIRNEY, L GHERMANDI, RS SIKES, L MARGUTTI, and C PHILLIPS. 1998. Plants, small mammals, and the hierarchical landscape classification of Patagonia. Landscape Ecology 13:285-306. [ Links ]
22. PIANKA ER. 1966. Latitudinal gradients in species diversity: a review of concepts. American Naturalist 100:33-46. [ Links ]
23. POPPER KR. 1985. Búsqueda sin término. Editorial Tecnos, Madrid. [ Links ]
24. PROHASKA F. 1976. The climate of Argentina, Paraguay and Uruguay. Pp. 13-122, in: Climates of Central and Southern America. World Survey of Climatology (W Schwerdtfeger, ed.). Elsevier, Amsterdam. [ Links ]
25. RAPOPORT EH. 1982. Areography, geographical strategies of species. Pergamon Press, Oxford. [ Links ]
26. RAPOPORT EH and JA MONJEAU. 2001. Areografía. Pp. 23-30, in: Introducción a la biogeografía en Latinoamérica: Teorías, conceptos, métodos y aplicaciones (J Llorente Bousquets y JJ Morrone, eds.). Siglo XXI, México. [ Links ]
27. RATISBONA C. 1976. The climate of Brazil. Pp. 219- 293, in: Climates of Central and Southern America World Survey of Climatology (W Schwerdtfeger, ed.). Elsevier, Amsterdam. [ Links ]
28. REDFORD KH and JF EISENBERG. 1989. Mammals of the Neotropics. Vol. 1. Mammals of the northern Neotropics: Panama, Colombia, Venezuela, Guyana, Suriname, French Guiana. University of Chicago Press, Chicago. [ Links ]
29. REDFORD KH and JF EISENBERG. 1992. Mammals of the Neotropics. Vol. 2. The Southern Cone. Chile, Argentina, Uruguay, Paraguay. University of Chicago Press, Chicago. [ Links ]
30. RHODE K. 1992. Latitudinal gradients in species diversity: the search for the primary cause. Oikos 65:514- 527. [ Links ]
31. ROIG VG. 1991. Desertification and distribution of mammals in the southern cone of South America. Pp. 239-279, in: Latin American mammalogy: History, biodiversity, and conservation (Mares y Schmidly, eds.), University of Oklahoma Press, Norman and London. [ Links ]
32. ROSENZWEIG ML. 1995. Species diversity in space and time. Cambridge University Press, Cambridge. [ Links ]
33. RUGGIERO A. 1994. Latitudinal correlates of the size of mammalian geographical ranges in South America. Journal of Biogeography 21:545-559. [ Links ]
34. RUGGIERO A. 1999. Spatial patterns in the diversity of mammal species: A test of the geographic area hypothesis in South America. Ecoscience 6:338-354. [ Links ]
35. RUGGIERO A, JH LAWTON, and TM BLACKBURN. 1998. The geographic ranges of mammalian species in South America: spatial patterns in environmental resistance and anisotropy. Journal of Biogeography 25:1093-1103. [ Links ]
36. SANDERSON E, M JAITEH, M LEVY, K REDFORD, A WANEEBO, and G WOOLMER. 2002. The human footprint and the last of the wild. Bioscience 52: 891-904 [ Links ]
37. SCHWERDTFEGER W. 1976. Climates of Central and Southern America. World Survey of Climatology (W Schwerdtfeger, ed.). Elsevier, Amsterdam. [ Links ]
38. SNOW JW. 1976. The climate of Central and Northern South America. Pp. 295-403, in: Climates of Central and Southern America World Survey of Climatology (W Schwerdtfeger, ed.). Elsevier, Amsterdam. [ Links ]
39. SOULE ME and J TERBORGH. 1999. Continental Conservation. Island Press, Washington D.C. [ Links ]
40. TERBORGH J, JA ESTES, P PAQUET, K RALLS, D BOYD-HEGER, BJ MILLER, and R NOSS. 1999. The role of top carnivores in regulating terrestrial ecosystems. Pp. 39-64, in: Continental Conservation (ME Soulé y J Terborgh, eds.). Island Press, Washington D.C. [ Links ]
41. TILMAN D. 1982. Resource competition and community structure. Princeton University Press, Princeton, New Jersey. [ Links ]
42. UDVARDY MDF. 1978. World biogeographical provinces. CoEvolution Quarterly (based on MDF Udvardy: International Union for Conservation of Nature and Natural Resources, Occasional Paper Nro. 18, 1975). Sausalito, California. [ Links ]
43. WILSON DE and DM REEDER. 1993. Mammals species of the world. A taxonomic and geographic reference. Smithsonian Institution Press, Washington DC. [ Links ]
44. WOZENCRAFT WC. 2004. Order Carnivora. A taxonomic and geographic reference. IUCN Carnivora Specialist Group. IUCN, Gland, Switzerland. [ Links ]
Recibido 28 febrero 2007.
Aceptado 26 marzo 2008.
Editor asociado: JJ Morrone

